1. Introduction
Post-transcriptional regulation is fundamental for
determining the appropriate time and place at which a given mRNA is
translated and its half-life. Processes as different as cell fate
determination during embryogenesis, differentiated cell response to
physiologic or stress cues, and stabilization of memories depend on
pre-localization and the regulated translation of specific sets of
mRNAs. RNA is thus the carrier of the two messages that are to be
translated into the protein and of the detailed information
concerning where and whether it is to be translated (1). Moreover, while mRNAs encoding
housekeeping proteins, such as cytoskeletal components, have on
average short decay rates of days, mRNAs encoding regulatory
proteins, such as transcription factors, have half-lives of minutes
(2). Deregulation of both
short-lived mRNA stability and/or localization/translation may
cause marked alterations in cell function, ranging from cancer to
cell death.
Post-transcriptional regulation largely depends on
the activity of a number of general and specific RNA binding
proteins (RBPs) that recognize consensus sequences and/or
structural features, mainly located in the 3′-untranslated region
(3′-UTR) of mRNAs but that can also be present in the
5′-untranslated- (5′-UTR) and coding sequences. Several RBPs bind
primary transcripts (heterogeneous nuclear RNAs: hnRNAs) following
transcription (Fig. 1), leading
to the assembly of nascent heterogeneous nuclear ribonucleoprotein
complexes (hnRNPs). In the context of hnRNPs, hnRNA undergoes
3′-end cleavage and the addition of a poly(A) tail, and these
events promote transcription termination and the assembly of a
dynamic enzymatic complex, the spliceosome. The spliceosome, which
contains five small nuclear RNPs (snRNPs) and numerous proteins, is
responsible for intron selection and removal, and the appropriate
joining of exons (3). RNA protein
interactions in the spliceosome are crucial for determining which
sequences should be retained in the mature message, and function as
checkpoints before mRNA transport to the cytoplasm. Conformational
perturbations within the spliceosome can control alternative
splicing decisions (4). Notably,
the molecular interactions within the spliceosome may be even more
complex during maturation of pre-mRNAs containing very large
introns, for which nested splicing events have been recently
suggested (5), as well as of
mRNAs containing ultra-short introns (6). The fact that crucial decisions are
taken during maturation of the primary transcript suggests that the
nuclear history of mRNA can be determined for its stability and
translation into the cytoplasm (7). The nucleo-cytoplasmic traffic of
RNPs occurs through the nuclear pore complex (NPC), and requires
nuclear export signals (NES), present on RBPs. The directionality
of nucleo-cytoplasmic traffic depends on remodelling of the mRNPs
on the cytoplasmic side of the nuclear envelope (8). Several RBPs also contain nuclear
localization signals (NLS), and shuttle between the nucleus and the
cytoplasm. Modulation, by post-translational modifications, of RBP
ability to enter the nucleus and/or to exit from it may be one of
the earliest regulatory steps in the pathway leading to the
translation of a given mRNA.
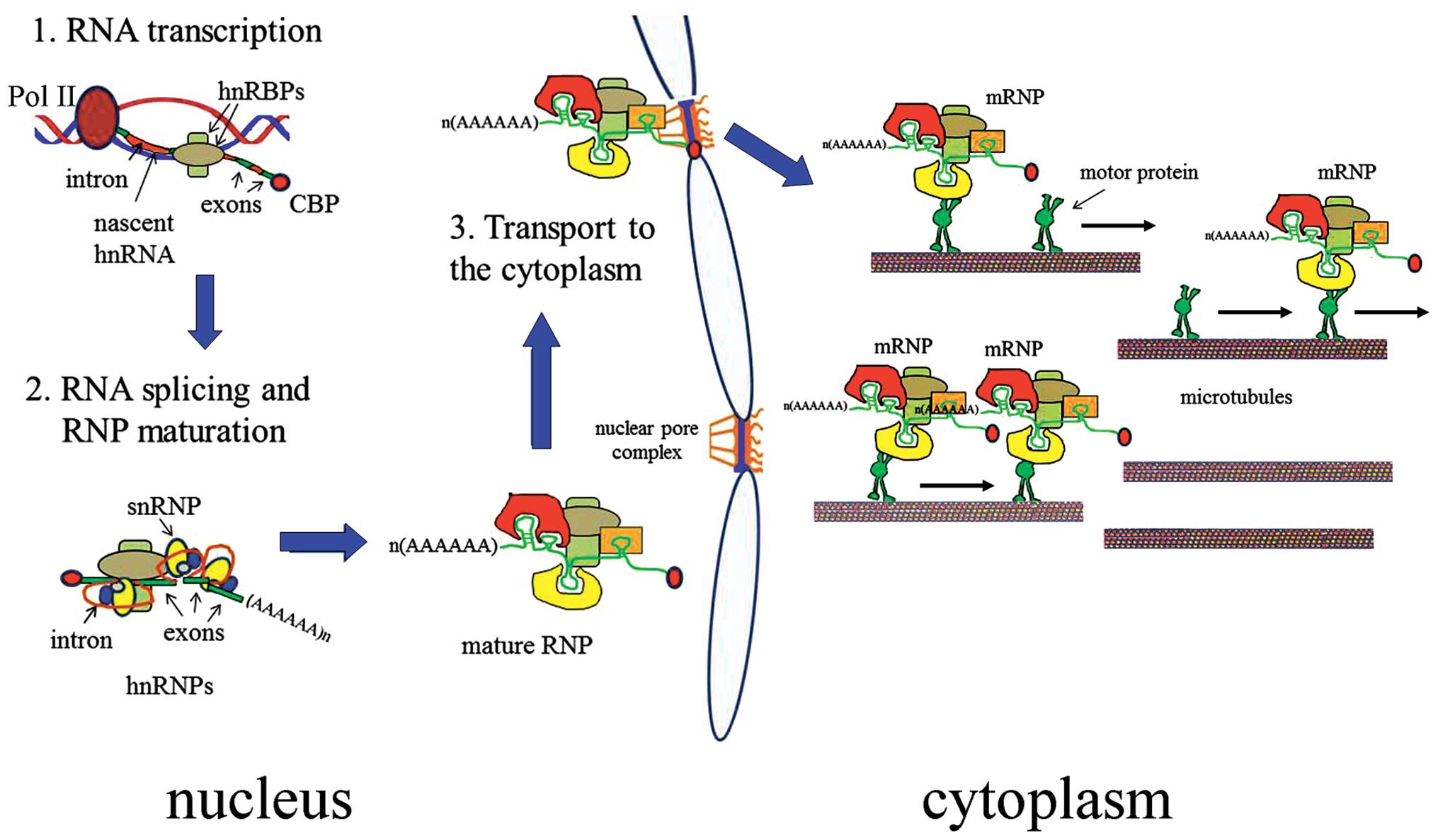 | Figure 1Post-transcriptional fate of mRNA.
During transcription (step 1) by RNA Polymerase II (Pol II), the
nascent heterogeneous nuclear primary transcript (nascent hnRNA) is
complexed with RNA-binding proteins (RBPs) (hnRBPs), some of which
are also involved in splicing (step 2), together with small nuclear
ribonucleoparticles (snRNPs). In the splicing process, introns are
removed and exons are joined together, to obtain a mature mRNA,
complexed with RBPs [mature RNA-protein particles (RNP)], ready to
be transported to the cytoplasm (step 3), through interaction with
proteins of the nuclear pore complex. In the cytoplasm, RNPs attach
to cytoplasmic motor proteins, which mediate the
microtubule-dependent transport of mRNA to its final
destination. |
Once in the cytoplasm, mRNPs are capable of binding
to motor proteins, which mediate their transport, in a
translationally inactive form, along the cytoskeletal filaments. At
their final destinations, mRNAs are immediately translated or
stored until a signal-dependent remodelling of mRNPs occurs.
Molecular mechanisms allowing the selective translation of
pre-localized mRNAs are of the most importance in both the
developing and adult nervous system, and can be at the basis of
learning and memory processes.
2. Regulatory signals and regulatory
factors
General
Post-transcriptional regulation depends on the
sequences/structures present on mRNA (cis-acting elements),
and trans-acting factors, and the ability to recognize them.
In addition, RNA protein interactions can be modulated over time by
extracellular signal transduction pathways that target RBPs,
inducing their post-translational remodelling.
Cis-acting determinants present on
mRNA
RNA is a highly dynamic molecule able to fold into
complex secondary and tertiary structures, based on standard
C:G/A:U base pairs as well as on non-Watson-Crick interactions
(9–14) generating different conformers with
different biological activities. During folding, RNA has the
tendency to be trapped in inactive tridimensional structures
(15) and it is not easy to
envisage thermodynamically predominant isoforms of a given mRNA
(16,17). Two main classes of RNA-binding
proteins cooperate in vivo with RNA in the acquisition of
functional structures: i) RNA chaperones, such as RNA helicases of
the DEAD-box family, or unwindases (18–20), which prevent and/or resolve
misfolded structures (15,21–23),
and ii) tertiary structure-binding proteins, which are able to
recognize and stabilize the ‘correctly’ folded RNAs (induced fit)
(16).
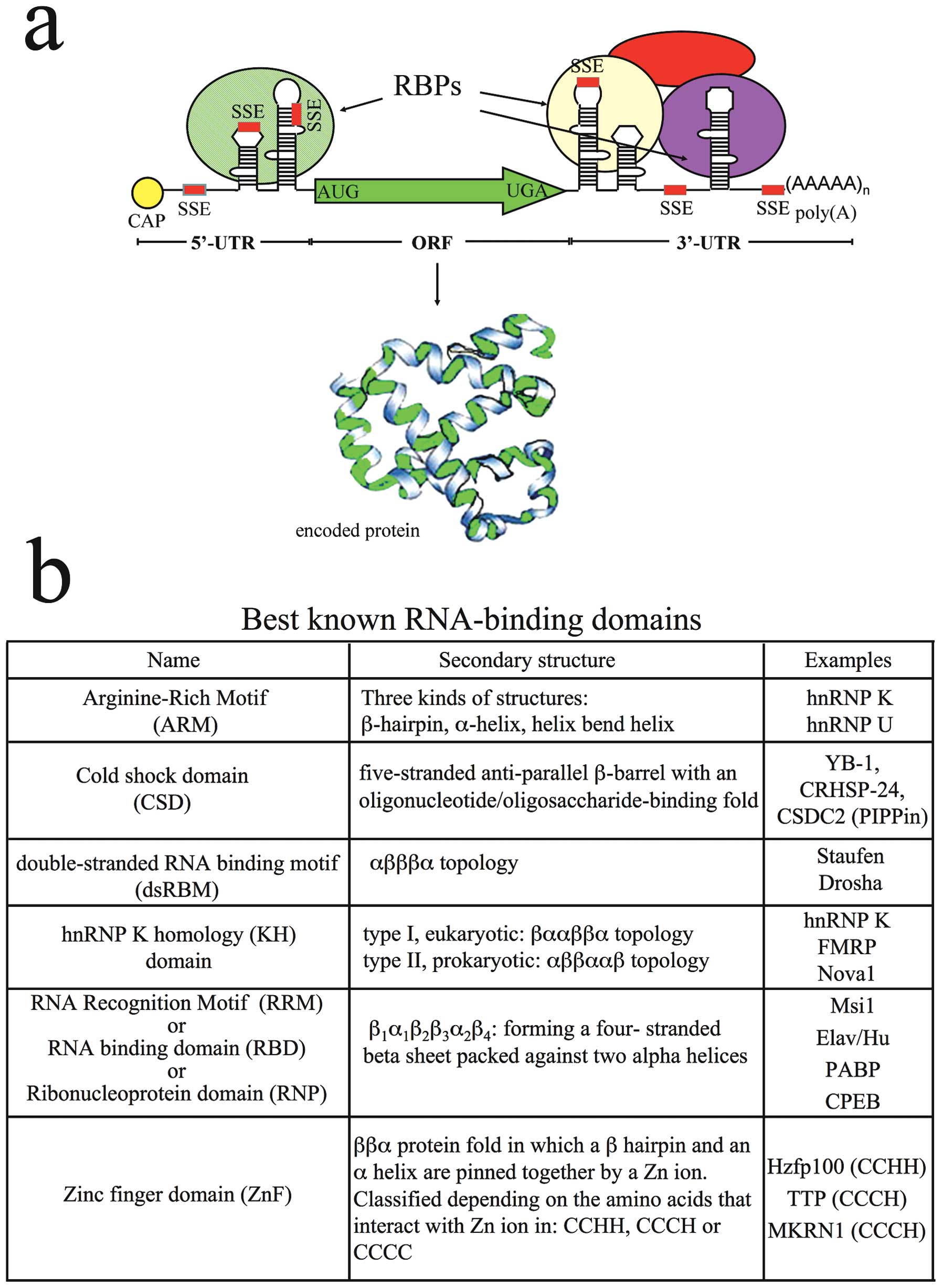
In general terms, cis-acting determinants or
‘zipcodes’ can be formed by simple sequences and/or
secondary/tertiary structure elements (24,25). Fig.
2a is a schematic drawing of mRNA structural organization.
Zipcodes may occasionally function independently of the precise
sequence context in which they are normally found: they are capable
of driving the localization of any reporter message to which they
are fused. However, not all the zipcodes are position-independent.
Some RNAs contain multimers of a single motif, while others contain
multiple cis-acting determinants; in both cases, multiple
zipcodes mediate multiple steps in the localization process by
acting synergistically (25).
Trans-acting factors: RNA-binding
proteins (RBPs) and non-coding RNAs (ncRNAs)
Cis-acting determinants are recognized by
RBPs that assemble on RNA, forming RNA protein particles (RNPs) of
different and regulated composition. In most cases,
trans-acting factors recognize a specific secondary
structure in the RNA, often a hairpin stem-loop structure, along
with a small number of specific anchoring nucleotides (25). RNA recognition and binding is
mediated by different families of RNA-binding domains, a summary of
which is shown in Fig. 2b.
Most RBPs contain more than one RNA-binding domain,
which has been shown to act synergistically by intra-molecular-
(26,27) or inter-molecular-cooperativity
(28). Conversely, the same RBP
is able to recognize different sequence elements within the target
mRNA, and recognition can require the correct spatial positioning
of these sequences (29). More
importantly, binding of all elements can affect overall affinity of
the RBP for the target mRNA. This cooperative effect depends on
significant unfolding or the structural changes the mRNA undergoes
after the first contact with the RBP (26,30). This observation is important when
we consider that mRNA also interacts with non-coding RNAs, the
regulatory role of which has been recently recognized (see
below).
A widely diffused RNA-binding domain is the
ribonucleoprotein (RNP) motif or RNA recognition motif (RRM). RRMs
contain ~90 amino acids and one or more copies of it were initially
identified in proteins that bind heterogeneous nuclear (hn)
pre-mRNA (hnRNP) (12,31). Proteins present in hnRNPs also
contain a variable number of Arg-Gly-Gly (RGG) repeats that in most
cases seem to be non-specific binding domains, acting in
cooperation with other RNA-binding domains (31). In alternative or in addition to
RRM and RGG motifs, hnRNPs may contain one or more domains known as
K-homology (KH) after hnRNP K, the first protein in which the motif
was identified (32).
Several RBPs contain an RNA-binding domain with a
preference for double-stranded stretches of RNA (dsRNA-binding
motif: DSRM). The prototypic DSRM was found in Staufen (Stau), a
protein required for maternal mRNA localization in
Drosophila egg (33). Stau
is also involved in the regulation of mRNAs required for memory
formation in both Drosophila and Aplysia (34,35). In mammals, two Stau isoforms are
present, which bind to distinct, although overlapping, sets of
mRNAs (36). Recently, Stau has
been reported to regulate pyramidal cell spine morphology via NMDA
receptor-mediated synaptic plasticity (37). Moreover, it probably forms a
functional complex with the fragile X mental retardation protein
(FMRP) and the 43 kDa transactive response DNA-binding protein
(TDP-43), both of which are related to nervous system pathologies
(see below) (38).
The most abundant protein family in the mammalian
genome is probably constituted by zinc-finger-containing proteins
(39), which contain several
types of Cys-Cys (CC) or Cys-His (CH) motifs, and were considered
primarily as transcriptional regulators. The first identified
zinc-finger containing protein, TFIIIA, is necessary for the
transcription, by RNA polymerase III, of the genes encoding
ribosomal 5S RNA, but is also able to bind ribosomal 5S RNA itself
in the 7S ribonucleoparticles stored in the cytoplasm of amphibian
oocytes. Similarly, many other members of the family are able to
bind the two classes of nucleic acids or RNA only (40,41).
In mammals, one example of CCHH zinc finger
domain-containing RBP is hZFP100, involved in the processing of
histone mRNAs (42), while a CCCH
domain characterizes tristetraprolin (TTP), which participates in
mRNA decay (43). TTP and the
other members of the TIS11 family have a crucial role in
post-transcriptional regulation, targeting for degradation
ARE-containing mRNAs (44,45).
In rat brain, an interaction between PABP (poly-A binding protein)
and Makorin 1 (MKRN 1), a RING zinc finger protein, has been
recently demonstrated (46). By
binding to PABP and dendritic mRNAs, MKRN1 was able to regulate
translation at synapses in response to stimuli inducing synaptic
plasticity (46).
Another domain, found in RBPs as well as in
DNA-binding proteins, is the cold-shock domain (CSD), first
identified in bacterial RNA chaperones and then found in a number
of eukaryotic proteins (Y-box proteins), which is able to interact
with single-stranded DNA and/or RNA (47). In one of the best studied
CSD-containing proteins, the Xenopus protein FRGY2, the CSD was
shown to be important for the sequence-specific RNA-binding, while
a second tail domain was involved in translation repression
(48). Y-box protein (YB)-1, the
prototypic member of the CSD-containing protein family, is both a
transcription factor and a major component of mRNPs. We recently
demonstrated that in nuclear extracts from rat brain, YB-1
interacts with a group of proteins that bind the mRNA encoding the
histone variant H1°. Among these proteins another CSD-containing
protein, CSD-C2, also known as PIPPin (49), was present (50).
Proteins that do not contain any conventional
RNA-binding domains have been shown to bind RNA. The variety of
trans-acting domains should therefore be larger than
expected. Examples of proteins, already known for other
well-established functions, which are also able to bind RNA,
include thymidylate synthase (51), mitochondrial glutamate
dehydrogenase (52), cytosolic
glyceraldehyde-3-p dehydrogenase (53), and the calmodulin-binding protein
PEP-19 (also known as PCP-4 in humans) (54).
Besides RNA-binding proteins, regulation of the mRNA
metabolism also involves non-coding RNAs (ncRNAs). A high
proportion of complex genomes gives rise to ncRNAs, among which the
most widely studied class is that of miRNAs, small RNAs of ~22
nucleotides (nt) that recognize specific sequences present in the
3′-UTR of target mRNAs. By binding their target mRNAs, miRNAs
mediate post-transcriptional gene silencing, through an increase of
mRNA decay rate and/or inhibition of translation (55–58). Deregulation of miRNA functions is
involved in cancer development (59–61) and in a number of other human
diseases, including neurological syndromes (62).
Biogenesis of miRNA is a multi-step process that
starts in the nucleus with the transcription of a long precursor
(pri-miRNA). Following the sequential action of the RNase III
enzymes Drosha and DICER, a dsRNA of ~22 nt is produced in the
cytoplasm, resulting in the formation of the RNA-induced silencing
complex (RISC) in which only one strand of the dsRNA is inserted
that contains members of the Argonaute (Ago) protein subfamily as
catalytic endonuclease components (56).
An increasing number of functional interactions have
been identified between RBPs and miRNAs. In general terms, RBPs and
miRNAs may cooperate or counteract in the regulation of a given
mRNA. Examples of the two types of RBPs-miRNA interplay have been
identified in different types of cancer cells (63). The role of miRNAs in neurons has
also been identified and previously assessed (64,65,66). Since a number of RBPs are
uniquely, or almost uniquely, expressed in the nervous system, the
possibility exists that they are capable of regulating the
interaction among mRNAs and certain groups of miRNAs in a specific
way, in turn regulating the establishment and/or maintenance of the
unusual functions of this tissue.
Signaling pathways that control mRNA
localization and metabolism
All the steps of RNA metabolism, from splicing and
maturation in the nucleus to final degradation, are affected by the
cellular microenvironment and by extracellular signals, such as
hormones, growth factors, physical and chemical stress stimuli, as
well as neurotransmitters (Fig.
3). All these cues are known to affect mRNA stability and
translation by triggering signaling pathways that cause
post-translational modifications of specific RBPs. A variety of
post-translational modifications affecting RBPs have been
identified, including serine/threonine phosphorylation, proline
hydroxylation, arginine/lysine methylation, lysine ubiquitination
or SUMOylation and lipidation. These chemical modifications
influence affinity and/or specificity of protein-protein and/or
RNA-protein interactions, and are consequently critical for the
rapid remodelling of ribonucleoprotein complexes and for the
stability and localization of mRNAs (2,67).
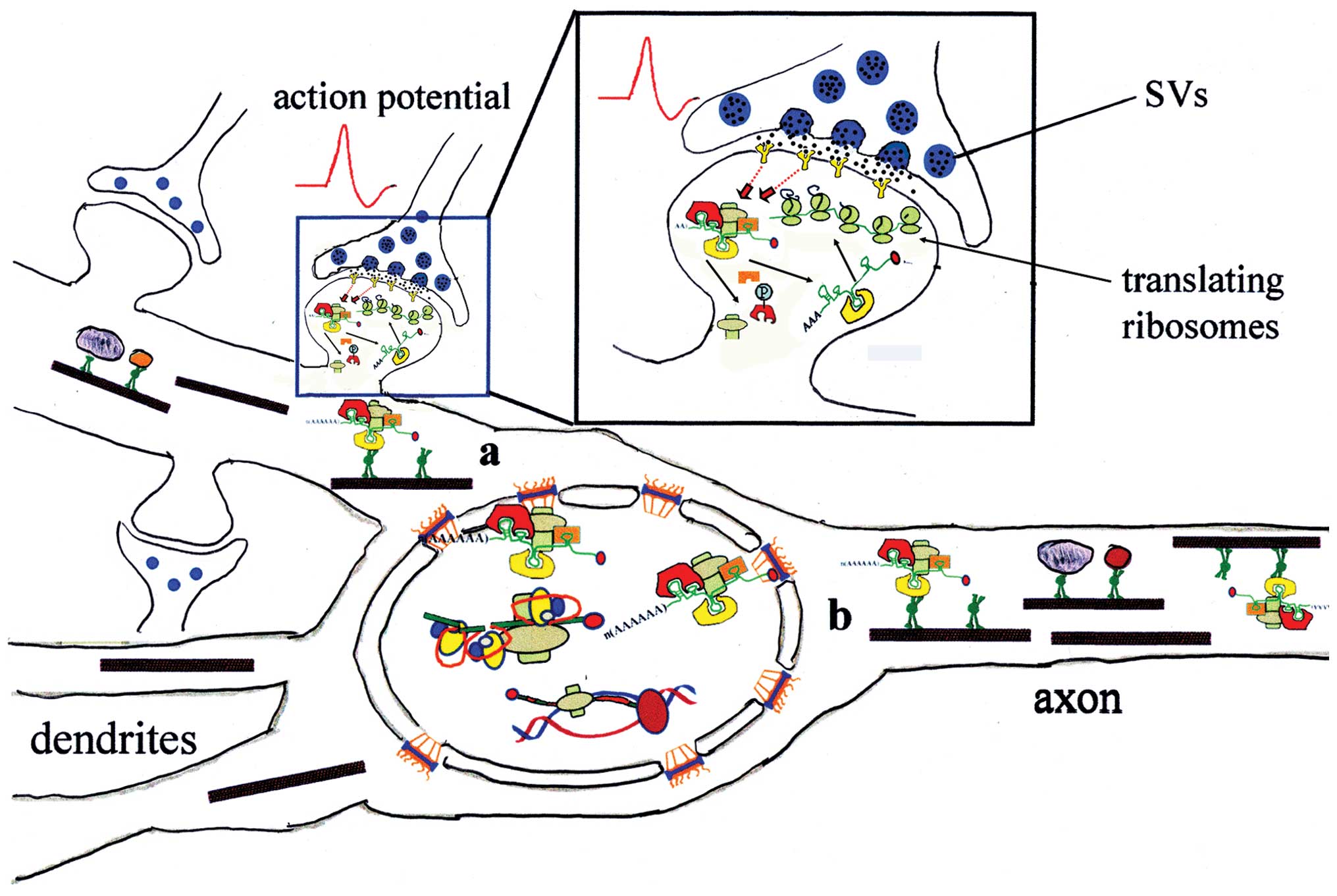 | Figure 3RNA traffic in neurons. As shown in
Fig. 1, at the time of
transcription, nascent RNA was already complexed with RNA-binding
proteins (RBPs) to form RNA-protein particles (RNPs), which then
migrate to the cytoplasm through the nuclear pore complex. In the
cytoplasm, through interaction with motor proteins, RNPs are
transported to their final destinations, where they can pause in a
silent state, until they are activated by a signal. Some components
of RNPs can then be cycled back to the nucleus. When exiting the
nucleus, however, two different peripheral destinations are
available to RNPs: a) dendrites or b) axon. Localized translation
of mRNA may be controlled by synaptic activity (action potential).
In response to neurotransmitters and/or neuromodulators, which bind
to their receptors on the post-synaptic element, signal
transduction pathways can be activated (red dotted lines), which
target RBPs, inducing post-translational modifications, such as
phosphorylation. RBP modifications in turn cause remodelling of the
RNPs, with the release of some RBPs and recruitment of mRNA to
ribosomes. Newly synthesized proteins accumulate at the
post-synaptic sites, thus inducing long-lasting modifications. SVs,
neurotransmitter-containing synaptic vesicles. Synthesis,
maturation, and transport of RNPs are shown as in Fig. 1. |
Localized mRNAs are transported close to the plasma
membrane or organelles, suggesting that membrane-bound kinases play
an important role in the regulation of the translation of localized
transcripts, in response to extracellular signals (68). Extracellular cues also activate
retrograde pathways that may even reach the nucleus, inducing the
modification of chromatin organization of genes and, in turn, gene
expression. In this manner, responses to extracellular signals and
neural activity may also be converted into a stable modification of
the cell phenotype.
At least three major mechanisms are known for
dynamic modification of chromatin organization: i) chromatin
remodelling by ATP-dependent complexes (69); ii) post-translational modification
of histone proteins (70), and
iii) replacement of histone isotypes present in chromatin with
other isotypes (71). During rat
brain development, at least histones H3.3, a core histone variant,
and H1°, a linker histone variant, have been shown to accumulate
during the terminal differentiation of nerve cells (72,73), in the absence of gene activity
changes. Post-transcriptional regulation of the expression of these
two histones probably depends on the activity of a group of
RNA-binding proteins (74,49),
some of which have been identified and cloned (75,76). Among these proteins, one was
already known as Purkinje cell-expressed peptide (PEP-19, or PCP-4
in humans), but is also expressed in a neuron-specific manner in
other brain regions (77,78).
PEP-19 is a calmodulin-binding protein that is known
to bind H1° mRNA (54).
Similarly, PIPPin (also known as CSDC2) binds H3.3 and H1° mRNAs
(49); it is a phosphoprotein
that can be phosphorylated by different kinases (79, unpublished
data), and that interacts with other proteins already known to bind
RNA (50). It may therefore be
hypothesized that, in response to calcium-dependent neuronal
activity, proteins of this type are modified. Their
post-translational modification may in turn change their
interaction with histone variant mRNAs, thus inducing their
translation and accumulation of the corresponding proteins. Histone
variants could finally enter chromatin, thus modifying the
structural organization and expression of specific genes.
3. Messenger RNA trafficking and
localization in neurons
Mature transcripts can be localized through at least
three mechanisms: i) local protection from degradation, ii)
diffusion and local anchoring, and iii) direct transport by
interaction with the cytoskeleton and cytoskeleton-associated motor
proteins (80–82). Subcellular pre-localization of
mRNAs and locally regulated translation offer cells at least three
important advantages: i) energy saving: a high number of protein
molecules can be obtained locally by transporting and translating a
single mRNA molecule; ii) efficacy/safety of protein production:
some proteins may be harmful to the cells when synthesized in the
incorrect location; the myelin basic protein (MBP), a major
component of the axon-wrapping myelin sheet, produced in the
central nervous system (CNS) by oligodendrocytes, is, for example,
a sticky protein that potentially interacts with any cell membrane
if produced in the cell body (12,83,84); other proteins, such as actin,
tubulin and microtubule-associated proteins, exist as different
isoforms, all able to form multimers/polymers; their localized
synthesis allows the formation of only the right multimers; iii)
differential translation regulation: in response to local
signaling, proteins may be synthesized only in the compartment
exposed to the signal (85,86).
Pre-localization of mRNA is determined in polarized
cells (87). The most asymmetric
cells in the body are neurons, which consist of a cell body or
soma, several branched dendrites and a long axon that can be
1,000-fold the diameter of the cell body (88). The most noteworthy property of
nerve cells is the variety of morphologically different
sub-compartments. Therefore, how a neuron, with only one cell
nucleus and thus a single supply of active genes, obtains such a
motley distribution of proteins if of interest and should be
investigated.
mRNA pre-localization in dendrites
Through utilization of different experimental
approaches, such as in situ hybridization, and amplification
of mRNAs present in neuronal processes, the presence in dendrites
of a large number of mRNAs was clearly demonstrated. Among the
dendritically localized messages the most intensively studied are
those encoding microtubule-associated protein 2 (MAP2), the
brain-derived neurotrophic factor, the α subunit of the
calcium/calmodulin-dependent protein kinase II (CaMKIIα), the
activity-regulated cytoskeleton-associated protein (Arc), the NMDAR
NR1 subunit, and the AMPA receptor (84,89,90). Dendritic RNA transport is specific
and can be regulated by neuronal activity in a rapid manner
(91,92).
Information required for the successful mRNA
transport to dendrites is sometimes contained in a single zipcode,
as in the case of the non-protein coding, dendritically localized
BC1 mRNA, which contains, at its 5′-end, a 62 nt zipcode, able to
fold into a single stem-loop and to drive microtubule-dependent
transport (25,93–95). Other mRNAs have more complex
localization signals. The MAP2 mRNA, for example, contains a 640 nt
zipcode element in its 3′-UTR, probably created with distinct
sub-elements that fold into multiple stem-loops (96), and are able to mediate individual
steps of RNP assembly and localization process. Similarly, the mRNA
encoding the myelin basic protein in oligodendrocytes contains two
distinct elements: i) A2RE, an 11 nt element found only in
transcripts that are localized (97); it forms the binding site for hnRNP
A2 and is necessary and sufficient for transport (98,99), and ii) a 1 kb long element,
essential for appropriate localization of a protein-coding reporter
RNA, and probably involved in RNA anchoring (25). Of note, the A2RE element was also
identified in the neuronal mRNAs encoding CaMKIIα, Arc and
neurogranin, all of which were assembled in the same granules
containing hnRNP A2, and targeted to dendrites (100). Conversely, it has also been
reported that the dendritically localized mRNAs encoding MAP2,
CaMKIIα, and β-actin assemble in distinct RNPs, which contain very
few RNA molecules (101,102).
Besides positively acting elements, elements that
inhibit transport also exist. Localization of CaMKIIα depends on
the 3′-UTR (103), which
contains three positive cis-acting regions: i) a stretch of
~1200 nt, in the latter half of the 3′-UTR, sufficient for
localization (104); ii) one
element present in the distal 170 nt of the UTR, which contains
cytoplasmic polyadenylation elements (CPEs) (105), and requires a wild-type
CPE-binding protein (CPEB); iii) a third zipcode, within the first
94 nt of the 3′-UTR, which contains the already mentioned, 30-nt
long sequence with high homology to an element present in
neurogranin mRNA. Other elements in the CaMKIIα mRNA, on the
contrary, inhibit its transport, in the resting state, while
inhibition is eliminated in the depolarized neurons as
activity-dependent derepression is critical for their dendritic
targeting (106).
Taken together, these findings suggest that assembly
and transport of mRNAs involves different signals and highly
specific RNA scanning by regulatory proteins. This complexity is
likely at the origin of the several types of RNPs that have been
described, such as transport RNPs, stress granules (SGs),
processing bodies (P-bodies), and P-body-like structures (107–112). Composition of the complexes may
vary with growth conditions, and different mRNA subsets are
potentially present in different classes of particles; conversely,
the same mRNA species may be found in multiple mRNP complexes.
Based on these observations, Keene and Tenenbaum suggested that
RBPs may regulate mRNAs as subpopulations during cell growth and
development. Their model predicted that functionally related genes
were regulated as groups or post-transcriptional operons, by
specific mRNA-binding proteins that recognize sequence elements
common among the mRNAs (113).
One of these ‘master’ RBPs that controls the expression of several
mRNAs at a time is the Fragile X mental retardation protein (FMRP),
which regulates dendritic mRNA transport and local protein
synthesis on the post-synaptic side of synapses. Over 400 mRNAs
have been reported to associate with FMRP. FMRP interacts with
kinesins and is able to repress translation of its client mRNAs
both in vitro and in vivo. More recently, FMRP has
also been found in axons and growth cones, where it seems to be
involved in responses induced by Semaphorin-3A (Sema3A) (114).
The specific interaction between RNAs and RBPs
suggests the existence of an ‘RNA signature’ that
characterizes each transcript (84,100). For example, the BC1 RNA
assembles with proteins to form an RNP involved in the transport of
dendritic mRNAs (93); the signal
responsible for its dendritic targeting is a motif at its 5′-end,
which folds into a single stem-loop instead of into a cloverleaf
structure, as expected on the basis of its sequence similarity to
tRNA. Notably, tRNAs also migrate to dendrites, but some of them
remain and function in the cell body. It has been suggested that
its structural properties allow BC1 RNA to be transported to
dendrites more efficiently than tRNAs (94).
Further sources of cis-acting signal
variability are alternative splicing and alternative
polyadenylation site selection. These processes are capable of
generating different RNA isoforms with different targeting
specificities. In brain neurons, two distinct pools of
brain-derived neurotrophic factor (BDNF) transcripts are produced
by the differential use of polyadenylation sites, i.e., mRNA
molecules with the longer 3′-UTR, but not those with the shorter
one, contain localization elements, which target them to dendrites
(115). At the same time, the
longer 3′-UTR also mediates BDNF translation repression in the
resting state and activity-dependent translation (116). In a recent study, it has been
found that the longer 3′-UTR contains an AU-rich element (ARE) that
interacts with the neuronal RBP HuD. Such an interaction is
necessary and sufficient for the stabilization of the longer mRNA
isoform (117).
A shorter and longer 3′-UTR species also exists in
the case of CaMKIIα mRNA, although the long mRNA is much more
abundant. Again, the sequence required for dendritic targeting has
been mapped downstream to the first polyadenylation site (104), suggesting that the shorter mRNA
isoform is confined to the cell body, while the longer mRNA is
transported to dendrites. It therefore seems that the strategy of
alternative 3′-UTRs may be commonly adopted by genes encoding
proteins with important functions in the cell body and
dendrites.
As mentioned, cis-acting elements interact
with RBPs that, by recognizing and binding them, control mRNA
localization and stability and thus play an important role in the
development and maintenance of the nervous system. Moreover,
coordinated expression of neural genes may be obtained by assembly
of their mRNAs in common mRNP complexes, which can contain
different RBPs under different cell states and at different times
(118). This process is of
particular importance in modulating synaptic plasticity which seems
to depend on the activity-induced translation of hundreds of
locally targeted mRNAs, present in neuronal processes, where they
drive a variety of specific functions (119).
For instance, mammalian zipcode-binding protein 1
(ZBP1) is required for the dendritic targeting of β-actin mRNA both
in developing and mature neurons (120–122). Knockdown of ZBP1 in cultured
hippocampal neurons reduced the dendritic levels of ZBP1 and
β-actin mRNA and impaired the growth of dendritic filopodia in
response to BDNF treatment (120). This observation is important
when considering that the formation and density of dendritic
filopodia during neuronal development are likely to be connected to
the process of synaptogenesis (120,123–125). Phosphorylation of ZBP1, in
response to extracellular signals such as BDNF, by the
membrane-bound Src tyrosine kinase results in a decreased affinity
of ZBP1 for its bound mRNA and consequent local activation of mRNA
translation (126–128). However, only ~50% of β-actin
mRNA colocalizes with ZBP1 (92),
thus suggesting the involvement of other RBPs in the
localization/translation regulation of this mRNA. Another RBP
required is the Src-associated with a mitosis of 68 kDa (Sam68):
knocking down Sam68 in neuronal cultures or interfering with its
binding to β-actin mRNA causes a deficit of β-actin mRNA in
dendrites and in spine density (122), while loss of Sam68 has been
connected with the pathogenesis of neurodegenerative fragile X
tremor/ataxia syndrome (FXTAS) (129,130).
The zipcode-binding protein, ZBP2, was identified in
chicken embryo brain by RNA affinity chromatography (through
binding to an RNA fragment containing the zipcode) (131). ZBP2 is a predominantly nuclear
protein that also affects β-actin mRNA localization in the
cytoplasm; it is the chicken homologue of the human KH
domain-containing splicing regulatory protein (KSRP), a protein
involved in pre-mRNA splicing (132). The homologue of ZBP2 in rat is
the MAP2-RNA trans-acting protein (MARTA1), which binds the
3′-UTR of MAP2 mRNA, a dendritically localized neuronal transcript
(133,134). ZBP2 has been found to bind the
nascent β-actin zipcode co-transcriptionally and to facilitate
binding of ZBP1 to the zipcode (135). It provides an example of how
interactions between RNA-binding proteins and RNA serve to recruit
and stabilize additional proteins to form a large RNP.
Another well-studied factor involved in mRNA
localization and, probably, modulation of translation is the
already mentioned HnRNP A2 which binds MBP mRNA, allowing its
correct localization in the oligodendrocyte processes in which
myelination occurs (127). hnRNP
F has been found to be a component of MBP transport granules, and
to cooperate with HnRNP A2 in regulating MBP expression. Activity
of this factor is controlled through phosphorylation by the Fyn
kinase (136). HnRNP A2 was
found in dendrites, in association with other hnRNPs, in large
macromolecular complexes (neuronal transport granules) that contain
mRNA, pre-mRNA splicing factors, and mRNA export factors (137). The presence of proteins with
different roles in RBP granules is a common observation: for
example, the complex responsible for dendritic targeting of a
CaMKII reporter contains PSF (polypyrimidine tract-binding
protein-associated splicing factor), hnRNPU and Staufen 1
RNA-binding proteins (127).
mRNA pre-localization in axons
When discussing mRNA localization determinants in
neurons, a challenging issue comes from the finding that mRNAs can
be also localized in axons. For a long time, this possibility was
not considered since the translational machinery did not seem to be
present at significant levels in axons. Moreover, mechanisms
ensuring protein transport from cell bodies to the axonal
compartment were, on the contrary, present and efficient, thus
making apparently unnecessary localized protein synthesis. However,
findings of previous studies have demonstrated axonal localization
and translation of several mRNA (138–144), including the β-actin mRNA
(145–147), which depends on the zipcode
recognized by ZBP1 for localization (148).
Together with β-actin transcript, mRNAs for which
axonal localization has been reported include those for oxytocin
and vasopressin (149,150), β-tubulin (151), actin-depolymerizing factor
ADF/cofilin (152), the
microtubule-associated proteins 1b (MAP1b) (153) and τ (154), the cytoskeleton regulating
protein RhoA (155), the κ
opioid receptor (156) and heat
shock proteins (157). Notably,
among axonally localized mRNAs, transcripts encoding transcription
factors have been found, an example being CREB mRNA (158). Moreover, after injury, the
population of localized mRNA enlarges to include many transcripts
encoding proteins of the translational apparatus and trans-membrane
receptors (159). Thus, as with
dendrites, a supply of mRNAs exists in axons, and some of these
transcripts can be translated under special circumstances (160). In addition, the pool of axonal
mRNAs markedly changes during development: in embryonic axons
exclusively transcripts belonging to the ‘cellular assembly and
organization category’ were found. These transcripts can be further
subdivided into smaller subsets of cytoskeletal-related mRNAs and
transport of vesicles/trafficking-related mRNAs (161). These observations suggest that,
at least in the growing axons, localized protein synthesis may
allow dynamic remodelling of the axons during their progression
through the extracellular environment.
Localization and translation of mRNA in axons still
pose two main problems. The first one concerns the organization and
regulation of the translational machinery: axoplasm structural
domains have been recently described (periaxoplasmic ribosomal
plaques: PARPs) which contain ribosomes attached to a superficial
plaque-like structural matrix, together with β-actin mRNA, ZBP-1,
SRP54, myosin Va and kinesin II molecular motor proteins. Rapid
axoplasmic transport of microinjected heterologous radiolabeled BC1
RNA to putative PARP domains suggested that the anchored
translation machinery potentially represents the destination of
specifically sorted mRNAs (162). Moreover, multiple translation
components, including ribosomal subunits and initiation factors,
interact with the trans-membrane receptor (DCC) for netrin-1,
suggesting that their activity can be regulated by extracellular
signals (163). The possibility
that ribosomes/mRNAs could be at least in part horizontally
transferred into axons from surrounding glial cells was also
discussed (142,164). In addition, it has been reported
that proximal segments of transected sciatic nerves accumulate
newly synthesized RNA in axons, and that these mRNAs are actually
synthesized in Schwann cells and then transferred to neurons
through a mechanism that requires actin cytoskeleton and myosin-Va
(165).
The second problem concerns specific transport to
dendrites and/or axons for those mRNAs that use at least some
identical zipcodes for both localizations. Preferential
localization depends on the entire supply of RBPs assembled in a
given RNP, including those necessary for anchoring RNPs to the
cytoskeleton. Several aspects of microtubule organization, among
which the specific microtubule-associated proteins (MAPs) and motor
proteins, are different between axons and dendrites. These
differences may be at the origin of specific sorting of cargoes and
their differential targeting in neurons (143,166).
4. Locally regulated stabilization and/or
translation of pre-localized mRNAs in neurogenesis
General
Neurogenesis is an extremely complex, multi-step
process through which self-renewing undifferentiated neural stem
cells obtain different and integrated differentiated phenotypes
(167). This process also occurs
in the adult nervous system following ischemic insult or damage
(168,169), and has been implicated in
learning, neuronal plasticity and memory formation in hippocampus
(170,171).
Regulation of mRNA metabolism during
development
During development, neurons sprout processes or
growth cones, which explore their environment and guide pathfinding
over extremely long distances. It is now clear that at least some
aspects of axon guidance require axonal mRNA translation (138–140,142–147). For example, Welshhans and
Bassell developed and used a new in vitro turning assay
allowing these authors to demonstrate that growth cones exhibit
protein synthesis-dependent attraction to netrin-1 and BDNF. This
attraction is lost in neurons lacking ZBP1; concomitantly,
BDNF-induced β-actin mRNA localization is also attenuated. ZBP1 is
also necessary for netrin-1-induced local translation of β-actin
mRNA (148).
Responses of growth cone to guidance cues such as
netrin-1, BDNF, and Sema3A proceed through cycles of
desensitization and resensitization that are critical for
navigation over large distances. Notably, in isolated growth cones,
the resensitization step is eliminated by treatment with
translation inhibitors, while the sensitization step is affected by
inhibitors of endocytosis, but not by translation inhibitors
(160,172,173). Thus, only some steps of growth
cone response to guidance cues require translation; when required,
however, the activation of translation is triggered by signal
transduction pathways which involve different kinases, including
phosphatidylinositol-3-kinase (PI3K), mammalian target of rapamycin
(mTOR), and mitogen-activated kinases (MAPKs) (160,174).
On the other hand, in the absence of induction
signals, as well as during the translocation to mRNA final
destinations, translation is inhibited. Some RBPs, such as ZBP1,
block translation initiation by inhibiting recruitment of the 60S
subunit of the ribosome. In other cases, RBPs modulate the length
of poly(A) tail (174,175). Conversely, the cytoplasmic
polyadenylation element-binding protein (CPEB) binds its
recognition element (CPE) in the 3′-UTR of its target mRNAs and
promotes polyadenylation-induced translation (176). Block of CPEB function in
hippocampal neurons reduces the translation of β-catenin, induced
in the growth cones by neurotrophin 3 (NT3), an effect that is
probably mediated by calcium ions and CaMKII (177).
Translational regulation by RBPs can also be
influenced by miRNAs; these latter molecules are capable of binding
RBPs which are, in turn, able to regulate their abundance (65,174,178–180). Several miRNAs are
developmentally regulated in mammalian neurons and have been
identified in axons (181,182) and dendrites (65).
RBPs are expressed in region-specific patterns in
developing brain, suggesting that they are crucial in the
establishment of cell type-specific functions (174,183). For example, the splicing
regulator Rbfox1 (A2BP1), which may involved in the control of
neuronal excitation and seizures, in the mammalian brain (184), is differentially expressed at
different developmental stages and in different brain regions
(185). In cortical neurons,
A2BP1 accumulates in the nucleus, the cell body and dendrites,
while in cultured hippocampal neurons it is mainly present in the
nucleus but also evidenced in the proximity of synapses (185).
During cell division, asymmetric segregation of RBPs
into one daughter cell also contributes to specify its fate, and to
generate a cell lineage. For instance, it has been recently
demonstrated that the RNA-binding protein Stau2 is asymmetrically
distributed during progenitor divisions in the developing mouse
cortex, thus determining the asymmetrical distribution of the mRNAs
to which it binds (186,187). In the radial glial precursor,
Stau2 forms a complex with other RBPs, such as Pumilio 2 (Pum2) and
DEAD box 1 (DDX1), and with a number of mRNAs, including those
encoding β-actin, mammalian prospero (prox1) (187), and the E3 ubiquitin ligase
tripartite motif protein 32 (Trim32) (186). Perturbation of the complex
induces premature differentiation of the radial glial precursors
into neurons and mislocalization of the target mRNAs (187). Thus, it seems that asymmetric
localization of RBPs can regulate the balance of stem cell
maintenance versus cell differentiation, as well as cell fate.
Musashi-1 (Msi1) RBP was first reported to play a
role in the development of the Drosophila adult external
sensory organ (188). In
mammals, Msi1 is considered a specific marker for neural/progenitor
stem cells (189–193). It acts by suppressing
translation of mRNAs encoding differentiation-inducing proteins,
such as the membrane protein Numb which is involved in the
Notch/Delta signalling cascade (118,191,194,195), and the cyclin-dependent kinase
inhibitor p21WAF-1 (196).
The embryonic lethal abnormal vision (ELAV)-like
proteins form a highly conserved RNA-binding protein family,
functionally involved in the stabilization of mRNAs which bear
AU-rich elements (ARE). As with their Drosophila homologue,
neuronal-specific proteins ELAV (nELAVL, originally HuB, HuC, and
HuD) are necessary and sufficient for inducing neuronal
differentiation in mammalian neural precursors (197–199) and are therefore considered
markers of post-mitotic neurons (200). The nELAVL proteins were
discovered through studies on paraneoplastic neurologic disorders
(PND) (201). Some common
tumors, such as small-cell lung, ovarian and breast cancers,
produce proteins normally expressed in the nervous system; thus,
the antitumor immune response is also directed against nerve cells,
triggering neurologic symptoms (202). A number of reports have
demonstrated that the overexpression of ELAVL proteins induces the
stabilization of ARE-containing mRNAs. Ratti and colleagues found
that nELAV proteins colocalize with, and bind to, Msi1 mRNA, in the
adult mouse subventricular zone and in cultured neural
stem/progenitor cells. They suggested that Msi1 and nELAV
RNA-binding activities might be complementary and exert a different
function on their target mRNAs. In other words, nELAV stabilization
of the Msi1 transcript may prolong its expression during the
gradual passage from proliferation to differentiation. This would
allow the stem/progenitor cell to continue to divide even after
Msi1 transcriptional inactivation (118). The post-transcriptional
regulatory action of nELAV proteins on Msi1 mRNA seems to be
evolutionarily restricted to mammals, and active from embryonic to
adult neurogenesis. In adult neurogenic areas, nELAV activity might
be transiently induced by spatial- and/or temporal-restricted
signals. It has also been suggested that upregulation and
cytoskeletal translocation of nELAV proteins, and the subsequent
positive effects on Gap43 mRNA levels may be involved in synaptic
plasticity and learning processes (see below), in the rat dentate
gyrus (203,204).
As mentioned, nELAV proteins are considered early
markers of neuronal commitment and are expressed in a specific
temporal order in the developing nervous system of vertebrates. In
the adult mouse, specific patterns of expression of each nELAV
member have been reported in different neuronal types, both in the
central and peripheral nervous systems, thus suggesting a role of
nELAV proteins in the maintenance of various types of postmitotic
neurons: for instance, HuC is strongly expressed in all neurons of
neocortex, while HuD is prevalent in the large projection cells of
the internal pyramidal layer, and HuB is detectable only in
scattered neurons (205,206).
Of the target mRNAs regulated by the nELAV proteins
some, in turn, encode proteins involved in mRNA metabolism,
including mRNA which encodes the neuro-oncological ventral antigen
1 (Nova1) (207), a
neuron-specific splicing factor that controls several mRNAs
important for synaptic function by alternative maturation (208,209). Nova1 mRNA stability and
translation are positively controlled by nELAV proteins. Moreover,
PKC-dependent nELAV phosphorylation induces the recruitment of
Nova1 mRNA to polysomes (207).
These findings suggest that, as in the case of transcription
factors, a hierarchy of RNA-binding proteins exists whose members
are expressed as part of a regulatory cascade.
Regulation of mRNA metabolism in learning
and memory
The most noteworthy implication of locally
regulated mRNA translation is its possible involvement in long-term
changes that accompany the complex processes of learning and
memory. Although Hu proteins were originally identified as early
markers of neuronal differentiation (210), persistence of HuB, HuC and HuD
mRNAs in the adult nervous system, especially in the hippocampus
and neocortex, indicates that they are also involved in adult
neuronal plasticity and spatial learning (200,204,211).
In mammals, HuD can form dimers and trimers that
associate with bound mRNAs, with cytoskeleton proteins and with
other ELAV-like proteins to form complexes (204) which, in turn, may associate with
polysomes to form translationally competent complexes involved in
learning (206) and in long-term
memory storage and recall. The parallel increase of cytoskeleton
proteins and HuD in the CA1 hippocampal region suggests a common
mechanism of regulation.
A learning-specific increase of rodent nELAV
proteins was demonstrated in both cell bodies and proximal
dendrites of hippocampal pyramidal cells. Notably, the concomitant
upregulation of HuD and GAP-43 expression, together with
co-localization of HuD and GAP-43 mRNA, have been described in
hippocampal pyramidal cells only in rodents that had learned two
different spatial discrimination paradigms (204,206). An effect of HuD in learning and
memory has also been suggested by using human HuD-overexpressing
mice, which do not show any motor/coordination deficits or apparent
developmental abnormalities (212). Although HuD expression increases
after learning, constitutive overexpression of this protein leads
to deficits in behavioural tests such as contextual fear
conditioning (CFC) and Morris water maze. These results suggest
that HuD levels are tightly regulated and any alteration results in
abnormal mRNA stabilization and cognitive impairment (212). Moreover, HuD is probably
involved in the stabilization of GAP-43 in growth cones, where
granules containing HuD also colocalize with GAP-43 mRNA and
ribosomes present in these regions (213). HuD was found to be associated
in vivo with large cytoplasmic granules in the neuronal cell
body and smaller granules in dendrites. Both types of granules were
also stained with the ribosomal marker Y10B, suggesting that they
also contain ribosomes (203).
The nELAV proteins seem to represent the final
target of a cellular cascade that involves PKC, and induces the
downstream stabilization of specific mRNAs, implicated in memory
trace formation (206). The role
of PKC was confirmed in cultured hippocampal neurons, where
phosphorylation of HuD regulates BDNF, NGF, NT-3 and GAP-43 mRNAs
stability and protein expression. HuD was suggested to be
negatively regulated by CARM1 methyl transferase, whose activity is
controlled by PKC-dependent phosphorylation (214).
Besides nELAV proteins, which have been the most
extensively studied RBPs, a few other proteins are involved in
learning and memory. Iijima and colleagues, for instance,
identified Hematopoietic Zinc Finger (Hzf), an RBP which regulates
the dendritic localization of mRNAs in neuronal cells. This protein
is highly expressed in Purkinje cells and binds to the 3′-UTR of
type 1 inositol 1,4,5-trisphosphate receptor (IP3R1) mRNA.
Post-transcriptional regulation by Hzf affects synaptic plasticity
and motor learning in cerebellum, because Hzf−/− mice
exhibit severe impairments in motor coordination and motor learning
related to cerebellar functions (215,216).
Obviously, the importance of RBP expression is
paralleled by the importance of RNA cis-acting elements
recognized by these proteins. Mutations in the 3′-UTR of mouse
CamKIIα mRNA cause in vivo the disruption of dendritic
localization and translation of CamKIIα, specifically affecting
long-term memory formation, suggesting that CamKIIα may be a
component of the translation system necessary for memory
consolidation (217).
5. RBPs linked to human disease
Considering their critical functions in
development, it is not surprising that altered RNA protein
interactions lead to severe pathologies, such as neurodegenerative
diseases and complex neurologic syndromes. Fragile X syndrome
(FXS), the most common form of inherited mental retardation,
depends on mutation of a gene, located in the X chromosome, which
encodes the already mentioned RNA-binding protein FMRP. Most
clinical cases of FXS result from hyper-expansion and methylation
of CGG repeats within the promoter of fmr1 that cause deficit of
its expression. FMRP is involved in the transport and translational
regulation of several dendrite-localized mRNAs, including that
encoding Arc (218,219), a protein required for
hippocampal long-term depression (LTD) and long-term potentiation
(LTP) (220), and the amyloid
precursor protein (APP) (221,222), thus FMRP function is related to
long-term synaptic plasticity. FMRP and its target mRNAs form
granule-like complexes that are transported with a
kinesin-dependent mechanism along microtubules (223).
Mice bearing a ko-fmr1 gene show specific defects
in trace fear memory and LTP loss in the anterior cingulate cortex
and lateral amygdala, while retaining normal locomotion capacity
and pain sensitivity (224). The
interaction between fmr1 and mGluR5, a Gp1 glutamate receptor,
whose increased activity is one of the consequences of FMRP loss of
function, was studied in mice bearing a ko version of fmr1, crossed
with mice which showed a reduced mGluR5 expression. The results
revealed that, in animals with 50% reduction of the activity of
mGluR5, the symptoms of FXS were reduced, counterbalancing the
defects caused by fmr1 mutation (225). The neuronal hyperexcitability
linked to a lack of FMRP, which generates epileptogenic responses,
resembles the effects of a lack of the already mentioned BC1 RNA,
suggesting that BC RNAs and FMRP have similar and overlapping modes
of action in translational repression at synapses (226).
FMRP deficiency was reported to affect the
proliferation rate in cultured adult neural progenitor/stem cells
(aNPCs), isolated from ko-fmr1 mice. Moreover, mutated aNPCs have a
higher tendency to differentiate into astrocytes as compared to
aNPCs derived from normal brains. Notably, in mutated aNPCs, the
Wnt signaling pathway as a whole is altered, and this alteration
can be at the origin of the observed modification of cell
differentiation balance (227).
In vivo, deletion of FMRP from aNSCs leads to deficits in
learning tasks that depend on adult hippocampal neurogenesis
(228). Using ko-fmr1 mice, a
marked impairment of cognitive functions was demonstrated,
accompanied by a decrease of post-synaptic proteins, and in
particular the NMDA receptor, although the relationship between the
two events has not yet been established (229).
Alterations similar to those observed in
FMRP-mutated mice were also found in translin knockout mice.
Similar to FMRP, translin is an RNA-binding protein that is able to
associate with microtubules and motor proteins, and is involved in
the transport and translation of mRNA localized in dendrites
(230). Similarly, huntingtin
(Htt) and the Htt-associated protein 1 (HAP1) have been reported to
be part of dendritic large transport granules which also contain
microtubule-dependent motors and transport β-actin mRNA (109).
Another degenerative pathology that may involve
altered RNA-protein interactions is frontotemporal lobar
degeneration (FTLD). FTLD has been classified into two subtypes: i)
the first one is characterized by accumulation of the
microtubule-associated protein τ (FTLD-τ) and ii) the second one is
characterized by inclusion bodies that do not contain τ, but
contain ubiquitin (FTLD-U) (231). Recently, a major component of
FTLD-U inclusion bodies has been identified as TAR DNA-binding
protein-43 (TDP-43), a protein of 43 amino acids encoded on
chromosome 1, that can bind both DNA and RNA and functions as a
splicing factor. Besides FTLD, amyotrophic lateral sclerosis (ALS)
and other related neurodegenerative disorders show inclusions of
TDP-43 and are now known as TDP-43 proteinopathies (232). TDP-43 extracted from tissues of
FTLD or ALS is often hyperphosphorylated, ubiquitinated and poorly
soluble (231). Of note, several
mutations in ALS have been found in the gene encoding angiogenin
(ANG), a secreted RNase (233).
Angiogenin and TDP-43 have been identified in stress granules,
assemblies of different RBPs and mRNAs. Moreover, angiogenin
stimulates the transcription of rRNA and is responsible for the
production of stress-induced small RNAs (234). As mentioned in a previous
section, TDP-43 has also been found in ribonucleoprotein complexes,
which also contain Stau and FMRP (38).
As a final example, proximal spinal muscular
atrophy (SMA) is an autosomal recessive disease caused by
alteration of the Smn1 gene, that encodes SMN, a protein present in
the cells as part of a complex which also contains Sm proteins and
U snRNAs, the core components of the pre-mRNA splicing machinery.
Recently, it has been reported that KH-type splicing regulatory
protein (KSRP), a multifunctional protein closely related to chick
ZBP2 and rat MARTA 1, and involved in the exosome-mediated
degradation of ARE-containing mRNAs, interacts with SMN. This
interaction is lost in the naturally occurring mutations of SMN,
found in Type 1 SMA patients (235). Another partner of SMN is the
ribonucleoprotein-R (hnRNPR), a protein involved in the axonal
transport of β-actin mRNA. Loss of β-actin mRNA in axonal growth
cones of hnRNPR-depleted motor neurons resembles the condition
observed in SMN-deficient motor neurons (236).
6. Conclusion and future directions
As reviewed in the previous sections, a number of
studies have previously clearly demonstrated the importance of mRNA
localization and the regulated translation in neuronal functions
ranging from axon pathfinding in development to nerve regeneration
in the adult nervous system, as well as in the long-lasting changes
involved in learning and memory.
However, a number of issues remain to be resolved.
It is, for example, still uncertain whether and how mRNAs are
actually anchored in the active sites of cell processes, e.g.,
whether silent RNPs are stably associated with specific synapses.
An interesting model has been proposed by Doyle and Kiebler
(84), who suggested that RNPs
containing the mRNAs necessary, for example, for specialized
dendritic functions are not strictly anchored in a single site but,
instead, continuously move, patrolling groups of synapses in
dendrites. RNPs should then be specifically captured and modified
in order to allow translation of the cargo mRNAs only from sites in
which synaptic activity is high. They named their model ‘sushi
belt model’, after the conveyor belt used in most Japanese
sushi restaurants: the belt conveys sushi to all the potential
consumers; however, the hungriest ones consume more items. In the
same manner, synapses recently activated should be ‘hungrier’ than
silent ones and are likely to capture/modify RNPs (84).
The most puzzling question is how a given mRNA is
specifically delivered to dendrites (pathway ‘a’ in Fig. 3) or to the axon (pathway ‘b’ in
Fig. 3). The
microtubule-associated proteins MAP2 and τ are enriched in
dendrites and axons, respectively; however, it remains to be
determined whether these and/or other cytoskeleton-associated
proteins have a role in selecting RNPs. As mRNAs already associate
with some RBPs in the nucleus, whether RNA final destination have
been decided from the beginning remains to be clarified, and if
this occurs specificity factors should be found among nuclear
proteins.
It seems that glial cells are involved in
regulating localized translation in axons, through the horizontal
transfer of ribosomes (142). An
attractive working hypothesis can thus be that glial cells also
transfer into neurons regulatory factors, including transcription
factors and RBPs, by sorting them to membrane microvesicles and/or
to exosomes, which are then shed from glial cells and fuse with
neuronal plasma membranes, delivering their cargoes into nerve
cells.
It also remains to be defined how miRNA and RBPs
cooperate in regulating mRNA metabolism in neurons. In these latter
cells, ncRNAs may have an additional role in mRNA silencing during
transport and storage processes. In conclusion, mRNAs, miRNAs and
RBPs form dynamic complexes in which part of the components
probably have only structural functions, while others confer upon
mRNA the ability to be specifically sorted to a given sub-cellular
compartment and/or to be translated in response to extracellular
signals. In other words, RNPs may be regarded as ‘mobile
nucleosomes’, possibly originating from an ancient RNA world.
Acknowledgements
The authors have been supported by the Università
degli Studi di Palermo, Palermo, Italy.
References
|
1
|
Mauger DM, Siegfried NA and Weeks KM: The
genetic code as expressed through relationships between mRNA
structure and protein function. FEBS Lett. 587:1180–1188. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Thapar R and Denmon AP: Signaling pathways
that control mRNA turnover. Cell Signal. 25:1699–1710. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Will CL and Lührmann R: Spliceosome
structure and function. Cold Spring Harb Perspect Biol. 3:pii:
a003707. 2011.
|
|
4
|
Chiou NT, Shankarling G and Lynch KW:
hnRNP L and hnRNP A1 induce extended U1 snRNA interactions with an
exon to repress spliceosome assembly. Mol Cell. 49:972–982. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Suzuki H, Kameyama T, Ohe K, Tsukahara T
and Mayeda A: Nested introns in an intron: evidence of multi-step
splicing in a large intron of the human dystrophin pre-mRNA. FEBS
Lett. 587:555–561. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Sasaki-Haraguchi N, Shimada MK, Taniguchi
I, Ohno M and Mayeda A: Mechanistic insights into human pre-mRNA
splicing of human ultra-short introns: potential unusual mechanism
identifies G-rich introns. Biochem Biophys Res Commun. 423:289–294.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Matsumoto K, Wassarman KM and Wolffe AP:
Nuclear history of a pre-mRNA determines the translational activity
of cytoplasmic mRNA. EMBO J. 17:2107–2121. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Stewart M: Nuclear export of mRNA. Trends
Biochem Sci. 35:609–617. 2010. View Article : Google Scholar
|
|
9
|
Ramos A, Gubser CC and Varani G: Recent
solution structures of RNA and its complexes with drugs, peptides
and proteins. Curr Opin Struct Biol. 7:317–323. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Linder P and Tuite MF: The versatility of
RNA structure and function. In: Jacques Monod Conference: New
insights into the mechanism of mRNA translation: the significance
of RNA structure; Aussois, France. 22–26 March 1999; Trends Genet.
15. pp. 302–303. 1999, View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Caprara MG and Nilsen TW: RNA: versatility
in form and function. Nat Struct Biol. 7:831–833. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Derrigo M, Cestelli A, Savettieri G and Di
Liegro I: RNA-protein interactions in the control of stability and
localization of messenger RNA (Review). Int J Mol Med. 5:111–123.
2000.PubMed/NCBI
|
|
13
|
Butcher SE and Pyle AM: The molecular
interactions that stabilize RNA tertiary structure: RNA motifs,
patterns, and networks. Acc Chem Res. 44:1302–1311. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hajdin CE, Bellaousov S, Huggins W,
Leonard CW, Mathews DH and Weeks KM: Accurate SHAPE-directed RNA
secondary structure modeling, including pseudoknots. Proc Natl Acad
Sci USA. 110:5498–5503. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Doetsch M, Schroeder R and Fürtig B:
Transient RNA-protein interactions in RNA folding. FEBS J.
278:1634–1642. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Weeks KM: Protein-facilitated RNA folding.
Curr Opin Struct Biol. 7:336–342. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Liu L and Chen SJ: Coarse-grained
prediction of RNA loop structures. PLoS One. 7:e484602012.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Fuller-Pace FV: RNA helicases: modulators
of RNA structure. Trends Cell Biol. 4:271–274. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
De la Cruz J, Kressler D and Linder P:
Unwinding RNA in Saccharomyces cerevisiae: DEAD-box,
proteins and related families. Trends Biochem Sci. 24:192–198.
1999.
|
|
20
|
Linder P: mRNA export: RNP remodeling by
DEAD-box proteins. Curr Biol. 18:R297–R299. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Herschlag D: RNA chaperones and the RNA
folding problem. J Biol Chem. 270:20871–20874. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Clodi E, Semrad K and Schroeder R:
Assaying RNA chaperone activity in vivo using a novel folding trap.
EMBO J. 18:3776–3782. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Grohman JK, Gorelick RJ, Lickwar CR, Lieb
JD, Bower BD, Znosko BM and Weeks KM: A guanosine-centric mechanism
for RNA chaperone function. Science. 340:190–195. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kislauskis EH and Singer RH: Determinants
of mRNA localization. Curr Opin Cell Biol. 4:975–978. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Jambhekar A and Derisi JL: Cis-acting
determinants of asymmetric, cytoplasmic RNA transport. RNA.
13:625–642. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Lunde BM, Moore C and Varani G:
RNA-binding proteins: modular design for efficient function. Nat
Rev Mol Cell Biol. 8:479–490. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Doyle M and Kiebler MA: A zipcode
unzipped. Genes Dev. 26:110–113. 2012. View Article : Google Scholar
|
|
28
|
Dienstbier M, Boehl F, Li X and Bullock
SL: Egalitarian is a selective RNA-binding protein linking mRNA
localization signals to the dynein motor. Genes Dev. 23:1546–1558.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Patel VL, Mitra S, Harris R, Buxbaum AR,
Lionnet T, Benowitz M, Girvin M, Levy M, Almo SC, Singer RH and
Chao JA: Spatial arrangement of an RNA zipcode identifies mRNAs
under post-transcriptional control. Genes Dev. 26:43–53. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Chao JA, Patskovsky Y, Patel V, Levy M,
Almo SC and Singer RH: ZBP1 recognition of β-actin zipcode induces
RNA looping. Genes Dev. 24:148–158. 2010.
|
|
31
|
Burd CG and Dreyfuss G: Conserved
structures and diversity of functions of RNA-binding proteins.
Science. 265:615–621. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Siomi H, Matunis MJ, Michael WM and
Dreyfuss G: The pre-mRNA binding K protein contains a novel
evolutionary conserved motif. Nucleic Acids Res. 21:1193–1198.
1993. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
St Johnston D, Benchle D and
Nusslein-Volhard C: Staufen, a gene required to localize maternal
RNAs in the Drosophila egg. Cell. 66:51–63. 1991.PubMed/NCBI
|
|
34
|
Dubnau J, Chiang AS, Grady L, Barditch J,
Gossweiler S, McNeil J, Smith P, Buldoc F, Scott R, Certa U, Broger
C and Tully T: The staufen/pumilio pathway is involved in
Drosophila long-term memory. Curr Biol. 13:286–296. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Liu J, Hu JY, Wu F, Schwartz JH and
Schacher S: Two mRNA-binding proteins regulate the distribution of
syntaxin mRNA in Aplysia sensory neurons. J Neurosci. 26:5204–5214.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Furic L, Maher-Laporte M and
DesGroseillers L: A genome-wide approach identifies distinct but
overlapping subsets of cellular mRNAs associated with Staufen1- and
Staufen2-containing ribonucleoprotein complexes. RNA. 14:324–335.
2008. View Article : Google Scholar
|
|
37
|
Lebeau G, DesGroseillers L, Sossin W and
Lacaille JC: mRNA binding protein staufen 1-dependent regulation of
pyramidal cell spine morphology via NMDA receptor-mediated synaptic
plasticity. Mol Brain. 4:222011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yu Z, Fan D, Gui B, Shi L, Xuan C, Shan L,
Wang Q, Shang Y and Wang Y: Neurodegeneration-associated TDP-43
interacts with fragile X mental retardation protein (FMRP)/Staufen
(STAU1) and regulates SIRT1 expression in neuronal cells. J Biol
Chem. 287:22560–22572. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ravasi T, Huber T, Zavolan M, Forrest A,
Gaasterland T, Grimmond S and Hume DA; RIKEN GER Group. GSL
Members: Systematic characterization of the zinc-finger-containing
proteins in the mouse transcriptome. Genome Res. 13:1430–1442.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Brown RS: Zinc finger proteins: getting a
grip on RNA. Curr Opin Struct Biol. 15:94–98. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Xu B and Koenig RJ: An RNA-binding domain
in the thyroid hormone receptor enhances transcriptional
activation. J Biol Chem. 279:33051–33056. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Dominski Z, Erkmann JA, Yang X, Sanchez R
and Marzluff WF: A novel zinc finger protein is associated with U7
snRNP and interacts with the stem-loop binding protein in the
histone pre-mRNP to stimulate 3′-end processing. Genes Dev.
16:58–71. 2002.PubMed/NCBI
|
|
43
|
Brewer BY, Malicka J, Blackshear PJ and
Wilson GM: RNA sequence elements required for high affinity binding
by the zinc finger domain of tristetraprolin: conformational
changes coupled to the bipartite nature of AU-rich
mRNA-destabilizing motifs. J Biol Chem. 279:27870–27877. 2004.
View Article : Google Scholar
|
|
44
|
Sanduja S, Blanco FF and Dixon DA: The
roles of TTP and BRF proteins in regulated mRNA decay. Wiley
Interdiscip Rev RNA. 2:42–57. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Carballo E, Lai WS and Blackshear PJ:
Feedback inhibition of macrophage tumor necrosis factor-α
production by tristetraprolin. Science. 281:1001–1005. 1998.
|
|
46
|
Miroci H, Schob C, Kindler S,
Ölschläger-Schütt J, Fehr S, Jungenitz T, Schwarzacher SW, Bagni C
and Mohr E: Makorin ring zinc finger protein 1 (MKRN1), a novel
poly(A)-binding protein-interacting protein, stimulates translation
in nerve cells. J Biol Chem. 287:1322–1334. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Grauman PL and Marahiel MA: A superfamily
of proteins that contain the cold-shock domain. Trends Biochem Sci.
23:286–290. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Matsumoto K, Meric F and Wolffe AP:
Translational repression dependent on the interaction of the
Xenopus Y-box Protein FRGY2 with mRNA. Role of the cold shock
domain, tail domain, and selective RNA sequence recognition. J Biol
Chem. 271:22706–22712. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Nastasi T, Scaturro M, Bellafiore M,
Raimondi L, Beccari S, Cestelli A and Di Liegro I: PIPPin is a
brain-specific protein that contains a cold-shock domain and binds
specifically to H1 degrees and H3.3 mRNAs. J Biol Chem.
274:24087–24093. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Di Liegro CM, Schiera G, Proia P, Saladino
P and Di Liegro I: Identification in the rat brain of a set of
nuclear proteins interacting with H1° mRNA. Neuroscience.
229:71–76. 2013.PubMed/NCBI
|
|
51
|
Chu E and Allegra CJ: The role of
thymidylate synthase as an RNA binding protein. Bioessays.
18:191–198. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Preiss T, Chrzanowska-Lightowlers ZM and
Lightowlers RN: Glutamate dehydrogenase: an organelle-specific
mRNA-binding protein. Trends Biochem Sci. 22:2901997. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhou Y, Yi X, Stoffer JB, Bonafe N,
Gilmore-Hebert M, McAlpine J and Chambers SK: The multifunctional
protein glyceraldehyde-3-phosphate dehydrogenase is both regulated
and controls colony-stimulating factor-1 messenger RNA stability in
ovarian cancer. Mol Cancer Res. 6:1375–1384. 2008. View Article : Google Scholar
|
|
54
|
Saladino P, Di Liegro CM, Proia P, Sala A,
Schiera G, Lo Cicero A and Di Liegro I: RNA-binding activity of the
rat calmodulin-binding PEP-19 protein and of the long PEP-19
isoform. Int J Mol Med. 29:141–145. 2012.PubMed/NCBI
|
|
55
|
He L and Hannon GJ: MicroRNAs: small RNAs
with a big role in gene regulation. Nat Rev Genet. 5:522–531. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Krol J, Loedige I and Filipowicz W: The
widespread regulation of microRNA biogenesis, function and decay.
Nat Rev Genet. 11:597–610. 2010.PubMed/NCBI
|
|
57
|
Bartel B: MicroRNAs: target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Fabian MR and Sonenberg N: The mechanics
of miRNA-mediated gene silencing: a look under the hood of miRISC.
Nat Struct Mol Biol. 19:586–593. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Mendell JT: MicroRNAs: critical regulators
of development, cellular physiology and malignancy. Cell Cycle.
4:1179–1184. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Esquela-Kerscher A and Slack FJ: Oncomirs
- microRNAs with a role in cancer. Nat Rev Cancer. 6:259–269. 2006.
View Article : Google Scholar
|
|
61
|
Croce CM: Causes and consequences of
microRNA dysregulation in cancer. Nat Rev Genet. 10:704–714. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Esteller M: Non-coding RNAs in human
disease. Nat Rev Genet. 12:861–874. 2011. View Article : Google Scholar
|
|
63
|
Ciafrè S and Galardi S: MicroRNAs and
RNA-binding proteins: a complex network of interactions and
reciprocal regulations in cancer. RNA Biol. 10:935–942.
2013.PubMed/NCBI
|
|
64
|
Kosik KS and Krichevsky AM: The elegance
of the microRNAs: a neuronal perspective. Neuron. 47:779–782. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Schratt GM, Tuebing F, Nigh EA, Kane CG,
Sabatini ME, Kiebler M and Greenberg ME: A brain-specific microRNA
regulates dendritic spine development. Nature. 439:283–289. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Im HI and Kenny PJ: MicroRNAs in neuronal
function and disfunction. Trends Neurosci. 35:325–334. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Lee SR and Lykke-Andersen J: Emerging
roles for ribonucleoprotein modification and remodelling in
controlling RNA fate. Trends Cell Biol. 23:504–510. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Paquin N and Chartrand P: Local regulation
of mRNA translation: new insights from the bud. Trends Cell Biol.
18:105–111. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Struhl K and Segal E: Determinants of
nucleosome positioning. Nat Struct Mol Biol. 20:267–273. 2013.
View Article : Google Scholar
|
|
70
|
Paul S and Knott JG: Epigenetic control of
cell fate in mouse blastocysts: the role of covalent histone
modifications and chromatin remodeling. Mol Reprod Dev. Jul
24–2013.(Epub ahead of print).
|
|
71
|
Szenker E, Ray-Gallet D and Almouzni G:
The double face of the histone variant H3.3. Cell Res. 21:421–434.
2011. View Article : Google Scholar
|
|
72
|
Castiglia D, Cestelli A, Scaturro M,
Nastasi T and Di Liegro I: H1(0) and H3.3B mRNA levels in
developing rat brain. Neurochem Res. 19:1531–1537. 1994. View Article : Google Scholar
|
|
73
|
Scaturro M, Cestelli A, Castiglia D,
Nastasi T and Di Liegro I: Posttranscriptional regulation of H1
zero and H3.3B histone genes in differentiating rat cortical
neurons. Neurochem Res. 20:969–976. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Scaturro M, Nastasi T, Raimondi L,
Bellafiore M, Cestelli A and Di Liegro I: H1(0) RNA-binding
proteins specifically expressed in the rat brain. J Biol Chem.
273:22788–22791. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Castiglia D, Scaturro M, Nastasi T,
Cestelli A and Di Liegro I: PIPPin, a putative RNA-binding protein
specifically expressed in the rat brain. Biochem Biophys Res
Commun. 218:390–394. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Sala A, Scaturro M, Proia P, Schiera G,
Balistreri E, Aflalo-Rattenbach R, Créau N and Di Liegro I: Cloning
of a rat-specific long PCP4/PEP19 isoform. Int J Mol Med.
19:501–509. 2007.PubMed/NCBI
|
|
77
|
Ziai R, Pan YC, Hulmes JD, Sangameswaran L
and Morgan JI: Isolation, sequence, and developmental profile of a
brain-specific polypeptide, PEP-19. Proc Natl Acad Sci USA.
83:8420–8423. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Slemmon JR, Morgan JI, Fullerton SM, Danho
W, Hilbush BS and Wengenack TM: Camstatins are peptide antagonists
of calmodulin based upon a conserved structural motif in PEP-19,
neurogranin, and neuromodulin. J Biol Chem. 271:15911–15917. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Auld GC, Campbell DG, Morrice N and Cohen
P: Identification of calcium-regulated heat-stable protein of 24
kDa (CRHSP24) as a physiological substrate for PKB and RSK using
KESTREL. Biochem J. 389:775–783. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
St Johnston D: Moving messages: the
intracellular localization of mRNAs. Nat Rev Mol Cell Biol.
6:363–375. 2005.PubMed/NCBI
|
|
81
|
Palacios IM: How does an mRNA find its
way? Intracellular localisation of transcripts. Semin Cell Dev
Biol. 18:163–170. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
Gonsalvez GB and Long RM: Spatial
regulation of translation through RNA localization. F1000 Biol Rep.
4:162012. View
Article : Google Scholar : PubMed/NCBI
|
|
83
|
St Johnston D: The intracellular
localization of messenger RNAs. Cell. 81:161–170. 1995.PubMed/NCBI
|
|
84
|
Doyle M and Kiebler MA: Mechanisms of
dendritic mRNA transport and its role in synaptic tagging. EMBO J.
30:3540–3552. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Andreassi C and Riccio A: To localize or
not to localize: mRNA fate is in 3′UTR ends. Trends Cell Biol.
19:465–474. 2009.
|
|
86
|
Jung H, Yoon BC and Holt CE: Axonal mRNA
localization and local protein synthesis in nervous system
assembly, maintenance and repair. Nat Rev Neurosci. 13:308–324.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
87
|
Holt CE and Bullock SL: Subcellular mRNA
localization in animal cells and why it matters. Science.
326:1212–1216. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
88
|
Donnelly CJ, Fainzilber M and Twiss JL:
Subcellular communication through RNA transport and localized
protein synthesis. Traffic. 11:1498–1505. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Eberwine J, Miyashiro K, Kacharmina JE and
Job C: Local translation of classes of mRNAs that are targeted to
neuronal dendrites. Proc Natl Acad Sci USA. 98:7080–7085. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
90
|
Grooms SY, Noh KM, Regis R, Bassell GJ,
Bryan MK, Carroll RC and Zukin RS: Activity bidirectionally
regulates AMPA receptor mRNA abundance in dendrites of hippocampal
neurons. J Neurosci. 26:8339–8351. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Rook MS, Lu M and Kosik KS: CaMKIIalpha 3′
untranslated region-directed mRNA translocation in living neurons:
visualization by GFP linkage. J Neurosci. 20:6385–6393. 2000.
|
|
92
|
Tiruchinapalli DM, Oleynikov Y, Kelic S,
Shenoy SM, Hartley A, Stanton PK, Singer RH and Bassell GJ:
Activity-dependent trafficking and dynamic localization of zipcode
binding protein 1 and beta-actin mRNA in dendrites and spines of
hippocampal neurons. J Neurosci. 23:3251–3261. 2003.PubMed/NCBI
|
|
93
|
Muslimov IA, Santi E, Homel P, Perini S,
Higgins D and Tiedge H: RNA transport in dendrites: a cis-acting
targeting element is contained within neuronal BC1 RNA. J Neurosci.
17:4722–4733. 1997.PubMed/NCBI
|
|
94
|
Rozhdestvensky TS, Kopylov AM, Brosius J
and Hüttenhofer A: Neuronal BC1 RNA structure: evolutionary
conversion of a tRNA(Ala) domain into an extended stem-loop
structure. RNA. 7:722–730. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
95
|
Cristofanilli M, Iacoangeli A, Muslimov IA
and Tiedge H: Neuronal BC1 RNA: microtubule-dependent dendritic
delivery. J Mol Biol. 356:1118–1123. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Blichenberg A, Schwanke B, Rehbein M,
Garner CC, Richter D and Kindler S: Identification of a cis-acting
dendritic targeting element in MAP2 mRNAs. J Neurosci.
19:8818–8829. 1999.PubMed/NCBI
|
|
97
|
Ainger K, Avossa D, Diana AS, Barry C,
Barbarese E and Carson JH: Transport and localization elements in
myelin basic protein mRNA. J Cell Biol. 138:1077–1087. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Hoek KS, Kidd GJ, Carson JH and Smith R:
hnRNP A2 selectively binds the cytoplasmic transport sequence of
myelin basic protein mRNA. Biochemistry. 37:7021–7029. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Munro TP, Magee RJ, Kidd GJ, Carson JH,
Barbarese E, Smith LM and Smith R: Mutational analysis of a
heterogeneous nuclear ribonucleoprotein A2 response element for RNA
trafficking. J Biol Chem. 274:34389–34395. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Gao Y, Tatavarty V, Korza G, Levin MK and
Carson JH: Multiplexed dendritic targeting of alpha calcium
calmodulin-dependent protein kinase II, neurogranin, and
activity-regulated cytoskeleton-associated protein RNAs by the A2
pathway. Mol Biol Cell. 19:2311–2327. 2008. View Article : Google Scholar
|
|
101
|
Mikl M, Vendra G and Kiebler MA:
Independent localization of MAP2, CaMKIIα and β-actin RNAs in low
copy numbers. EMBO Rep. 12:1077–1084. 2011.PubMed/NCBI
|
|
102
|
Tübing F, Vendra G, Mikl M, Macchi P,
Thomas S and Kiebler MA: Dendritically localized transcripts are
sorted into distinct ribonucleoprotein particles that display fast
directional motility along dendrites of hippocampal neurons. J
Neurosci. 30:4160–4170. 2010.
|
|
103
|
Mayford M, Baranes D, Podsypanina K and
Kandel ER: The 3′-untranslated region of CaMKII alpha is a
cis-acting signal for the localization and translation of mRNA in
dendrites. Proc Natl Acad Sci USA. 93:13250–13255. 1996.
|
|
104
|
Blichenberg A, Rehbein M, Muller R, Garner
CC, Richter D and Kindler S: Identification of a cis-acting
dendritic targeting element in the mRNA encoding the alpha subunit
of Ca2+/calmodulin-dependent protein kinase II. Eur J
Neurosci. 13:1881–1888. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Huang YS, Carson JH, Barbarese E and
Richter JD: Facilitation of dendritic mRNA transport by CPEB. Genes
Dev. 17:638–653. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Mori Y, Imaizumi K, Katayama T, Yoneda T
and Tohyama M: Two cis-acting elements in the 3′ untranslated
region of alpha-CaMKII regulate its dendritic targeting. Nat
Neurosci. 3:1079–1084. 2000.
|
|
107
|
Zeitelhofer M, Macchi P and Dahm R:
Perplexing bodies: the putative roles of P-bodies in neurons. RNA
Biol. 5:244–248. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Miller LC, Blandford V, McAdam R,
Sanchez-Carbente MR, Badeaux F, DesGroseillers L and Sossin WS:
Combinations of DEAD box proteins distinguish distinct types of
RNA: protein complexes in neurons. Mol Cell Neurosci. 40:485–495.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Ma B, Savas JN, Yu MS, Culver BP, Chao MV
and Tanese N: Huntingtin mediates dendritic transport of β-actin
mRNA in rat neurons. Sci Rep. 1:1402011.
|
|
110
|
Batista PJ and Chang HY: Cytotopic
localization by long noncoding RNAs. Curr Opin Cell Biol.
25:195–199. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Lloyd RE: Regulation of stress granules
and P-bodies during RNA virus infection. Wiley Interdiscip Rev RNA.
4:317–331. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Oh JY, Kwon A, Jo A, Kim H, Goo YS, Lee JA
and Kim HK: Activity-dependent synaptic localization of processing
bodies and their role in dendritic structural plasticity. J Cell
Sci. 126:2114–2123. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Keene JD and Tenenbaum SA: Eukaryotic
mRNPs may represent posttranscriptional operons. Mol Cell.
9:1161–1167. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
114
|
Li C, Bassell GJ and Sasaki Y: Fragile X
mental retardation protein is involved in protein
synthesis-dependent collapse of growth cones induced by
semaphorin-3A. Front Neural Circuits. 3:112009.PubMed/NCBI
|
|
115
|
An JJ, Gharami K, Liao GY, Woo NH, Lau AG,
Vanevski F, Torre ER, Jones KR, Feng Y, Lu B and Xu B: Distinct
role of long 3′ UTR BDNF mRNA in spine morphology and synaptic
plasticity in hippocampal neurons. Cell. 134:175–187. 2008.
|
|
116
|
Lau AG, Irier HA, Gu J, Tian D, Ku L, Liu
G, Xia M, Fritsch B, Zheng JQ, Dingledine R, Xu B, Lu B and Feng Y:
Distinct 3′UTRs differentially regulate activity-dependent
translation of brain-derived neurotrophic factor (BDNF). Proc Natl
Acad Sci USA. 107:15945–15950. 2010.
|
|
117
|
Allen M, Bird C, Feng W, Liu G, Li W and
Perrone-Bizzozero NI: HuD promotes BDNF expression in brain neurons
via selective stabilization of the BDNF long 3′UTR mRNA. PLoS One.
8:e557182013.PubMed/NCBI
|
|
118
|
Ratti A, Fallini C, Cova L, Fantozzi R,
Calzarossa C, Zennaro E, Pascale A, Quattrone A and Silani V: A
role for the ELAV RNA-binding proteins in neural stem cells:
stabilization of Msi1 mRNA. J Cell Sci. 119:1442–1452. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
119
|
Martin KC and Ephrussi A: mRNA
localization: gene expression in the spatial dimension. Cell.
136:719–730. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Eom T, Antar LN, Singer RH and Bassell GJ:
Localization of a beta-actin messenger ribonucleoprotein complex
with zipcode-binding protein modulates the density of dendritic
filopodia and filopodial synapses. J Neurosci. 23:10433–10444.
2003.PubMed/NCBI
|
|
121
|
Perycz M, Urbanska AS, Krawczyk PS,
Parobczak K and Jaworski J: Zipcode binding protein 1 regulates the
development of dendritic arbors in hippocampal neurons. J Neurosci.
31:5271–5285. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Klein ME, Younts TJ, Castillo PE and
Jordan BA: RNA-binding protein Sam68 controls synapse number and
local β-actin mRNA metabolism in dendrites. Proc Natl Acad Sci USA.
110:3125–3130. 2013.PubMed/NCBI
|
|
123
|
Cooper MW and Smith SJ: A real time
analysis of growth cone target cell interactions during the
formation of stable contacts between hippocampal neurons in
culture. J Neurobiol. 23:814–828. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
124
|
Ziv NE and Smith SJ: Evidence for a role
of dendritic filopodia in synaptogenesis and spine formation.
Neuron. 17:91–102. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
125
|
Fiala JC, Feinberg M, Popov V and Harris
KM: Synaptogenesis via dendritic filopodia in developing
hippocampal area CA1. J Neurosci. 18:8900–8911. 1998.PubMed/NCBI
|
|
126
|
Hüttelmaier S, Zenklusen D, Lederer M,
Dictenberg J, Lorenz M, Meng X, Bassell GJ, Condeelis J and Singer
RH: Spatial regulation of beta-actin translation by Src-dependent
phosphorylation of ZBP1. Nature. 438:512–515. 2005.PubMed/NCBI
|
|
127
|
Giorgi C and Moore MJ: The nuclear nurture
and cytoplasmic nature of localized mRNPs. Semin Cell Dev Biol.
18:186–193. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Sasaki Y, Welshhans K, Wen Z, Yao J, Xu M,
Goshima Y, Zheng JQ and Bassell GJ: Phosphorylation of zipcode
binding protein 1 is required for brain-derived neurotrophic factor
signaling of local beta-actin synthesis and growth cone turning. J
Neurosci. 30:9349–9358. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Brouwer JR, Willemsen R and Oostra BA: The
FMR1 gene and fragile X-associated tremor/ataxia syndrome. Am J Med
Genet B Neuropsychiatr Genet. 150B:782–798. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Sellier C, Rau F, Liu Y, Tassone F, Hukema
RK, Gattoni R, Schneider A, Richard S, Willemsen R, Elliott DJ,
Hagerman PJ and Charlet-Berguerand N: Sam68 sequestration and
partial loss of function are associated with splicing alterations
in FXTAS patients. EMBO J. 29:1248–1261. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Gu W, Pan F, Zhang H, Bassell GJ and
Singer RH: A predominantly nuclear protein affecting cytoplasmic
localization of beta-actin mRNA in fibroblasts and neurons. J Cell
Biol. 156:41–51. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Min H, Turck CW, Nikolic JM and Black DL:
A new regulatory protein, KSRP, mediates exon inclusion through an
intronic splicing enhancer. Genes Dev. 11:1023–1036. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
133
|
Rehbein M, Kindler S, Horke S and Richter
D: Two trans-acting rat-brain proteins, MARTA1 and MARTA2, interact
specifically with the dendritic targeting element in MAP2 mRNAs.
Brain Res Mol Brain Res. 79:192–201. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
134
|
Rehbein M, Wege K, Buck F, Schweizer M,
Richter D and Kindler S: Molecular characterization of MARTA1, a
protein interacting with the dendritic targeting element of MAP2
mRNAs. J Neurochem. 82:1039–1046. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
135
|
Pan F, Huttelmaier S, Singer RH and Gu W:
ZBP2 facilitates binding of ZBP1 to beta-actin mRNA during
transcription. Mol Cell Biol. 27:8340–8351. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
136
|
White R, Gonsior C, Bauer NM,
Krämer-Albers EM, Luhmann HJ and Trotter J: Heterogeneous nuclear
ribonucleoprotein (hnRNP) F is a novel component of
oligodendroglial RNA transport granules contributing to regulation
of myelin basic protein (MBP) synthesis. J Biol Chem.
287:1742–1754. 2012. View Article : Google Scholar
|
|
137
|
Kiebler MA and Bassell GJ: Neuronal RNA
granules: movers and makers. Neuron. 51:685–690. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Kelly RB and Grote E: Protein targeting in
the neuron. Annu Rev Neurosci. 16:95–127. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
139
|
Alvarez J, Giuditta A and Koenig E:
Protein synthesis in axons and terminals: significance for
maintenance, plasticity and regulation of phenotype. With a
critique of slow transport theory. Prog Neurobiol. 62:1–62. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
140
|
Satkauskas S and Bagnard D: Local protein
synthesis in axonal growth cones: what is next? Cell Adh Migr.
1:179–184. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
141
|
Giuditta A, Chun JT, Eyman M, Cefaliello
C, Bruno AP and Crispino M: Local gene expression in axons and
nerve endings: the glia-neuron unit. Physiol Rev. 88:515–555. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Twiss JL and Fainzilber M: Ribosomes in
axons - scrounging from the neighbors? Trends Cell Biol.
19:236–243. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Perry RB and Fainzilber M: Local
translation in neuronal processes-in vivo tests of a ‘heretical
hypothesis’. Dev Neurobiol. Aug 8–2013.(Epub ahead of print).
|
|
144
|
Gumy LF, Katrukha EA, Kapitein LC and
Hoogenraad CC: New insights into mRNA trafficking in axons. Dev
Neurobiol. Aug 19–2013.(Epub ahead of print).
|
|
145
|
Willis DE, Xu M, Donnelly CJ, Tep C,
Kendall M, Erenstheyn M, English AW, Schanen NC, Kirn-Safran CB,
Yoon SO, Bassell GJ and Twiss JL: Axonal localization of transgene
mRNA in mature PNS and CNS neurons. J Neurosci. 31:14481–14487.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Donnelly CJ, Willis DE, Xu M, Tep C, Jiang
C, Yoo S, Schanen NC, Kirn-Safran CB, van Minnen J, English A, Yoon
SO, Bassell GJ and Twiss JL: Limited availability of ZBP1 restricts
axonal mRNA localization and nerve regeneration capacity. EMBO J.
30:4665–4677. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Donnelly CJ, Park M, Spillane M, Yoo S,
Pacheco A, Gomes C, Vuppalanchi D, McDonald M, Kim HH, Merianda TT,
Gallo G and Twiss JL: Axonally synthesized beta-actin and GAP-43
proteins support distinct modes of axonal growth. J Neurosci.
33:3311–3322. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Welshhans K and Bassell GJ:
Netrin-1-induced local beta-actin synthesis and growth cone
guidance requires zipcode binding protein 1. J Neurosci.
31:9800–9813. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Jirikowski GF, Sanna PP and Bloom FE: mRNA
coding for oxytocin is present in axons of the
hypothalamo-neurohypophysial tract. Proc Natl Acad Sci USA.
87:7400–7404. 1990. View Article : Google Scholar : PubMed/NCBI
|
|
150
|
Trembleau A, Morales M and Bloom FE:
Differential compartmentalization of vasopressin messenger RNA and
neuropeptide within the rat hypothalamo-neurohypophysial axonal
tracts: light and electron microscopic evidence. Neuroscience.
70:113–125. 1996. View Article : Google Scholar
|
|
151
|
Eng H, Lund K and Campenot RB: Synthesis
of beta-tubulin, actin, and other proteins in axons of sympathetic
neurons in compartmented cultures. J Neurosci. 19:1–9.
1999.PubMed/NCBI
|
|
152
|
Lee SK and Hollenbeck PJ: Organization and
translation of mRNA in sympathetic axons. J Cell Sci.
116:4467–4478. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Antar LN, Li C, Zhang H, Carroll RC and
Bassell GJ: Local functions for FMRP in axon growth cone motility
and activity-dependent regulation of filopodia and spine synapses.
Mol Cell Neurosci. 32:37–48. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
154
|
Litman P, Barg J, Rindzoonski L and
Ginzburg I: Subcellular localization of tau mRNA in differentiating
neuronal cell culture: implications for neuronal polarity. Neuron.
10:627–638. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
155
|
Wu KY, Hengst U, Cox LJ, Macosko EZ,
Jeromin A, Urquhart ER and Jaffrey SR: Local translation of RhoA
regulates growth cone collapse. Nature. 436:1020–1024. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
156
|
Bi J, Tsai NP, Lin YP, Loh HH and Wei LN:
Axonal mRNA transport and localized translational regulation of
kappa-opioid receptor in primary neurons of dorsal root ganglia.
Proc Natl Acad Sci USA. 103:19919–19924. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Willis D, Li KW, Zheng JQ, Chang JH, Smit
A, Kelly T, Merianda TT, Sylvester J, van Minnen J and Twiss JL:
Differential transport and local translation of cytoskeletal,
injury-response, and neurodegeneration protein mRNAs in axons. J
Neurosci. 25:778–791. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Cox LJ, Hengst U, Gurskaya NG, Lukyanov KA
and Jaffrey SR: Intra-axonal translation and retrograde trafficking
of CREB promotes neuronal survival. Nat Cell Biol. 10:149–159.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
159
|
Willis DE, van Niekerk EA, Sasaki Y,
Mesngon M, Merianda TT, Williams GG, Kendall M, Smith DS, Bassell
GJ and Twiss JL: Extracellular stimuli specifically regulate
localized levels of individual neuronal mRNAs. J Cell Biol.
178:965–980. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
160
|
Yoon BC, Zivraj KH and Holt CE: Local
translation and mRNA trafficking in axon pathfinding. Results Probl
Cell Differ. 48:269–288. 2009.PubMed/NCBI
|
|
161
|
Gumy LF, Yeo GS, Tung YC, Zivraj KH,
Willis D, Coppola G, Lam BY, Twiss JL, Holt CE and Fawcett JW:
Transcriptome analysis of embryonic and adult sensory axons reveals
changes in mRNA repertoire localization. RNA. 17:85–98. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Koenig E: Organized ribosome-containing
structural domains in axons. Results Probl Cell Differ. 48:173–191.
2009.PubMed/NCBI
|
|
163
|
Tcherkezian J, Brittis PA, Thomas F, Roux
PP and Flanagan JG: Transmembrane receptor DCC associates with
protein synthesis machinery and regulates translation. Cell.
141:632–644. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Court FA, Hendriks WT, Macgillavry HD,
Alvarez J and van Minnen J: Schwann cell to axon transfer of
ribosomes: toward a novel understanding of the role of glia in the
nervous system. J Neurosci. 28:11024–11029. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
165
|
Sotelo JR, Canclini L, Kun A,
Sotelo-Silveira JR, Xu L, Wallrabe H, Calliari A, Rosso G, Cal K
and Mercer JA: Myosin-Va-dependent cell-to-cell transfer of RNA
from Schwann cells to axons. PLoS One. 8:e619052013. View Article : Google Scholar : PubMed/NCBI
|
|
166
|
Kapitein LC and Hoogenraad CC: Which way
to go? Cytoskeletal organization and polarized transport in
neurons. Mol Cell Neurosci. 46:9–20. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
167
|
Franco SJ and Müller U: Shaping our minds:
stem and progenitor cell diversity in the mammalian neocortex.
Neuron. 77:19–34. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
168
|
Douen AG, Dong L, Vanance S, Munger R,
Hogan MJ, Thompson CS and Hakim AM: Regulation of nestin expression
after cortical ablation in adult rat brain. Brain Res.
1008:139–146. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
169
|
Brus M, Keller M and Lévy F: Temporal
features of adult neurogenesis: differences and similarities across
mammalian species. Front Neurosci. 7:1352013. View Article : Google Scholar : PubMed/NCBI
|
|
170
|
Kempermann G, Jessberger S, Steiner B and
Kronenberg G: Milestones of neuronal development in the adult
hippocampus. Trends Neurosci. 27:447–452. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
171
|
Pan YW, Storm DR and Xia Z: Role of adult
neurogenesis in hippocampus-dependent memory, contextual fear
extinction and remote contextual memory: new insights from ERK5 MAP
kinase. Neurobiol Learn Mem. 105:81–92. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
172
|
Ming GL, Wong ST, Henley J, Yuan XB, Song
HJ, Spitzer NC and Poo MM: Adaptation in the chemotactic guidance
of nerve growth cones. Nature. 417:411–418. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
173
|
Piper M, Salih S, Weinl C, Holt CE and
Harris WA: Endocytosis-dependent desensitization and protein
synthesis-dependent resensitization in retinal growth cone
adaptation. Nat Neurosci. 8:179–186. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
174
|
Hörnberg H and Holt C: RNA-binding
proteins and translational regulation in axons and growth cones.
Front Neurosci. 7:812013.PubMed/NCBI
|
|
175
|
Charlesworth A, Meijer HA and de Moor CH:
Specificity factors in cytoplasmic polyadenylation. Wiley
Interdiscip Rev RNA. 4:437–461. 2013. View Article : Google Scholar
|
|
176
|
Richter JD: CPEB: a life in translation.
Trends Biochem Sci. 32:279–285. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
177
|
Kundel M, Jones KJ, Shin CY and Wells DG:
Cytoplasmic polyadenylation element-binding protein regulates
neurotrophin-3-dependent beta-catenin mRNA translation in
developing hippocampal neurons. J Neurosci. 29:13630–13639. 2009.
View Article : Google Scholar
|
|
178
|
Michlewski G, Guil S, Semple CA and
Cáceres JF: Posttranscriptional regulation of miRNAs harboring
conserved terminal loops. Mol Cell. 32:383–393. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
179
|
Edbauer D, Neilson JR, Foster KA, Wang CF,
Seeburg DP, Batterton MN, Tada T, Dolan BM, Sharp PA and Sheng M:
Regulation of synaptic structure and function by FMRP-associated
microRNAs miR-125b and miR-132. Neuron. 65:373–384. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
180
|
Xu XL, Zong R, Li Z, Biswas MH, Fang Z,
Nelson DL and Gao FB: FXR1P but not FMRP regulates the levels of
mammalian brain-specific microRNA-9 and microRNA-124. J Neurosci.
31:13705–13709. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
181
|
Hengst U, Cox LJ, Macosko EZ and Jaffrey
SR: Functional and selective RNA interference in developing axons
and growth cones. J Neurosci. 26:5727–5732. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
182
|
Aschrafi A, Schwechter AD, Mameza MG,
Natera-Naranjo O, Gioio AE and Kaplan BB: microRNA-338 regulates
local cytochrome c oxidase IV mRNA levels and oxidative
phosphorylation in the axons of sympathetic neurons. J Neurosci.
28:12581–12590. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
183
|
McKee AE, Minet E, Stern C, Riahi S,
Stiles CD and Silver PA: A genome-wide in situ hybridization map of
RNA-binding proteins reveals anatomically restricted expression in
the developing mouse brain. BMC Dev Biol. 5:14–22. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
184
|
Gehman LT, Stoilov P, Maguire J, Damianov
A, Lin CH, Shiue L, Ares M Jr, Mody I and Black DL: The splicing
regulator Rbfox1 (A2BP1) controls neuronal excitation in the
mammalian brain. Nat Genet. 43:706–711. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
185
|
Hamada N, Ito H, Iwamoto I, Mizuno M,
Morishita R, Inaguma Y, Kawamoto S, Tabata H and Nagata KI:
Biochemical and morphological characterization of A2BP1 in neuronal
tissue. J Neurosci Res. 91:1303–1311. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
186
|
Kusek G, Campbell M, Doyle F, Tenenbaum
SA, Kiebler M and Temple S: Asymmetric segregation of the
double-stranded RNA binding protein Staufen2 during mammalian
neural stem cell divisions promotes lineage progression. Cell Stem
Cell. 11:505–516. 2012. View Article : Google Scholar
|
|
187
|
Vessey JP, Amadei G, Burns SE, Kiebler MA,
Kaplan DR and Miller FD: An asymmetrically localized
Staufen2-dependent RNA complex regulates maintenance of mammalian
neural stem cells. Cell Stem Cell. 11:517–528. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
188
|
Nakamura M, Okano H, Blendy JA and Montell
C: Musashi, a neural RNA-binding protein required for
Drosophila adult external sensory organ development. Neuron.
13:67–81. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
189
|
Kaneko Y, Sakakibara S, Imai T, Suzuki A,
Nakamura Y, Sawamoto K, Ogawa Y, Toyama Y, Miyata T and Okano H:
Musashi1: an evolutionally conserved marker for CNS progenitor
cells including neural stem cells. Dev Neurosci. 22:139–153. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
190
|
Maslov AY, Barone TA, Plunkett RJ and
Pruitt SC: Neural stem cell detection, characterization, and
age-related changes in the subventricular zone of mice. J Neurosci.
24:1726–1733. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
191
|
Okano H, Kawahara H, Toriya M, Nakao K,
Shibata S and Imai T: Function of RNA-binding protein Musashi-1 in
stem cells. Exp Cell Res. 306:349–356. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
192
|
Okamoto K, Nakatsukasa M, Alié A, Masuda
Y, Agata K and Funayama N: The active stem cell specific expression
of sponge Musashi homolog EflMsiA suggests its involvement in
maintaining the stem cell state. Mech Dev. 129:24–37. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
193
|
Sutherland JM, McLaughlin EA, Hime GR and
Siddall NA: The musashi family of RNA binding proteins: master
regulators of multiple stem cell populations. Adv Exp Med Biol.
786:233–245. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
194
|
Imai T, Tokunaga A, Yoshida T, Hashimoto
M, Mikoshiba K, Weinmaster G, Nakafuku M and Okano H: The neural
RNA-binding protein Musashi1 translationally regulates mammalian
numb gene expression by interacting with its mRNA. Mol Cell Biol.
21:3888–3900. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
195
|
Perez-Asensio FJ, Perpiñá U, Planas AM and
Pozas E: Interleukin-10 regulates progenitor differentiation and
modulates neurogenesis in adult brain. J Cell Sci. 126:4208–4219.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
196
|
Battelli C, Nikopoulos GN, Mitchell JG and
Verdi JM: The RNA binding protein Musashi-1 regulates neural
development through the translational repression of p21(WAF-1). Mol
Cell Neurosci. 31:85–96. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
197
|
Akamatsu W, Okano HJ, Osumi N, Inoue T,
Nakamura S, Sakakibara S, Miura M, Matsuo N, Darnell RB and Okano
H: Mammalian ELAV-like neuronal RNA-binding proteins HuB and HuC
promote neuronal development in both the central and the peripheral
nervous systems. Proc Natl Acad Sci USA. 96:9885–9890. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
198
|
Akamatsu W, Fujihara H, Mitsuhashi T, Yano
M, Shibata S, Hayakawa Y, Okano HJ, Sakakibara S, Takano H, Takano
T, Takahashi T, Noda T and Okano H: The RNA binding protein HuD
regulates neuronal cell identity and maturation. Proc Natl Acad Sci
USA. 102:4625–4630. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
199
|
Kasashima K, Terashima K, Yamamoto K,
Sakashita E and Sakamoto H: Cytoplasmic localization is required
for the mammalian ELAV-like protein HuD to induce neuronal
differentiation. Genes Cells. 4:667–683. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
200
|
Okano HJ and Darnell RB: A hierarchy of Hu
RNA binding proteins in developing and adult neurons. J Neurosci.
17:3024–3037. 1997.PubMed/NCBI
|
|
201
|
Szabo A, Dalmau J, Manley G, Rosenfeld M,
Wong E, Henson J, Posner JB and Furneaux HM: HuD, a paraneoplastic
encephalomyelitis antigen, contains RNA binding domains and is
homologous to Elav and Sex-lethal. Cell. 67:325–333. 1991.
View Article : Google Scholar : PubMed/NCBI
|
|
202
|
Darnell RB: RNA protein interaction in
neurons. Annu Rev Neurosci. 36:243–270. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
203
|
Bolognani F, Merhege MA, Twiss J and
Perrone-Bizzozero NI: Dendritic localization of the RNA-binding
protein HuD in hippocampal neurons: association with polysomes and
upregulation during contextual learning. Neurosci Lett.
371:152–157. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
204
|
Pascale A, Gusev PA, Amadio M, Dottorini
T, Govoni S, Alkon DL and Quattrone A: Increase of the RNA-binding
protein HuD and post-transcriptional upregulation of the GAP-43
gene during spatial memory. Proc Natl Acad Sci USA. 101:1217–1222.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
205
|
Clayton GH, Perez GM, Smith RL and Owens
GC: Expression of mRNA for the elav-like neural-specific RNA
binding protein, HuD, during nervous system development. Brain Res
Dev Brain Res. 109:271–280. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
206
|
Pascale A, Amadio M and Quattrone A:
Defining a neuron: neuronal ELAV proteins. Cell Mol Life Sci.
65:128–140. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
207
|
Ratti A, Fallini C, Colombrita C, Pascale
A, Laforenza U, Quattrone A and Silani V: Post-transcriptional
regulation of neuro-oncological ventral antigen 1 by the neuronal
RNA-binding proteins ELAV. J Biol Chem. 283:7531–7541. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
208
|
Ule J, Ule A, Spencer J, Williams A, Hu
JS, Cline M, Wang H, Clark T, Fraser C, Ruggiu M, Zeeberg BR, Kane
D, Weinstein JN, Blume J and Darnell RB: Nova regulates
brain-specific splicing to shape the synapse. Nat Genet.
37:844–852. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
209
|
Ule J and Darnell RB: RNA binding proteins
and the regulation of neuronal synaptic plasticity. Curr Opin
Neurobiol. 16:102–110. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
210
|
Wakamatsu Y and Weston JA: Sequential
expression and role of Hu RNA-binding proteins during neurogenesis.
Development. 124:3449–3460. 1997.PubMed/NCBI
|
|
211
|
Quattrone A, Pascale A, Nogues X, Zhao W,
Gusev P, Pacini A and Alkon DL: Posttranscriptional regulation of
gene expression in learning by the neuronal ELAV-like
mRNA-stabilizing proteins. Proc Natl Acad Sci USA. 98:11668–11673.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
212
|
Bolognani F, Qiu S, Tanner DC, Paik J,
Perrone-Bizzozero NI and Weeber EJ: Associative and spatial
learning and memory deficits in transgenic mice overexpressing the
RNA-binding protein HuD. Neurobiol Learn Mem. 87:635–643. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
213
|
Smith CL, Afroz R, Bassell GJ, Furneaux
HM, Perrone-Bizzozero NI and Burry RW: GAP-43 mRNA in growth cones
is associated with HuD and ribosomes. J Neurobiol. 61:222–235.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
214
|
Lim CS and Alkon DL: Protein kinase C
stimulates HuD-mediated mRNA stability and protein expression of
neurotrophic factors and enhances dendritic maturation of
hippocampal neurons in culture. Hippocampus. 22:2303–2319. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
215
|
Iijima T, Imai T, Kimura Y, Bernstein A,
Okano HJ, Yuzaki M and Okano H: Hzf protein regulates dendritic
localization and BDNF-induced translation of type 1 inositol 1,4,5
trisphosphate receptor mRNA. Proc Natl Acad Sci USA.
102:17190–17195. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
216
|
Iijima T, Ogura H, Takatsuki K, Kawahara
S, Wakabayashi K, Nakayama D, Fujioka M, Kimura Y, Bernstein A,
Okano HJ, Kirino Y and Okano H: Impaired motor functions in mice
lacking the RNA-binding protein Hzf. Neurosci Res. 58:183–189.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
217
|
Miller S, Yasuda M, Coats JK, Jones Y,
Martone ME and Mayford M: Disruption of dendritic translation of
CaMKIIalpha impairs stabilization of synaptic plasticity and memory
consolidation. Neuron. 36:507–519. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
218
|
Bassell GJ and Warren ST: Fragile X
syndrome: loss of local mRNA regulation alters synaptic development
and function. Neuron. 60:201–214. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
219
|
Pfeiffer BE and Huber KM: The state of
synapses in fragile X syndrome. Neuroscientist. 15:549–567. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
220
|
Sidorov MS, Auerbach BD and Bear MF:
Fragile X mental retardation protein and synaptic plasticity. Mol
Brain. 6:152013. View Article : Google Scholar : PubMed/NCBI
|
|
221
|
Westmark CJ and Malter JS: FMRP mediates
mGluR5-dependent translation of amyloid precursor protein. PLoS
Biol. 5:e522007. View Article : Google Scholar : PubMed/NCBI
|
|
222
|
Ascano M Jr, Mukherjee N, Bandaru P,
Miller JB, Nusbaum JD, Corcoran DL, Langlois C, Munschauer M,
Dewell S, Hafner M, Williams Z, Ohler U and Tuschl T: FMRP targets
distinct mRNA sequence elements to regulate protein expression.
Nature. 492:382–386. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
223
|
Dictenberg JB, Swanger SA, Antar LN,
Singer RH and Bassell GJ: A direct role for FMRP in
activity-dependent dendritic mRNA transport links filopodial-spine
morphogenesis to fragile X syndrome. Dev Cell. 14:926–939. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
224
|
Zhao MG, Toyoda H, Ko SW, Ding HK, Wu LJ
and Zhuo M: Deficits in trace fear memory and long-term
potentiation in a mouse model for fragile X syndrome. J Neurosci.
25:7385–7392. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
225
|
Dölen G, Osterweil E, Rao BS, Smith GB,
Auerbach BD, Chattarji S and Bear MF: Correction of fragile X
syndrome in mice. Neuron. 56:955–962. 2007.PubMed/NCBI
|
|
226
|
Iacoangeli A and Tiedge H: Translational
control at the synapse: role of RNA regulators. Trends Biochem Sci.
38:47–55. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
227
|
Luo Y, Shan G, Guo W, Smrt RD, Johnson EB,
Li X, Pfeiffer RL, Szulwach KE, Duan R, Barkho BZ, Li W, Liu C, Jin
P and Zhao X: Fragile x mental retardation protein regulates
proliferation and differentiation of adult neural stem/progenitor
cells. PLoS Genet. 6:e10008982010. View Article : Google Scholar : PubMed/NCBI
|
|
228
|
Guo W, Allan AM, Zong R, Zhang L, Johnson
EB, Schaller EG, Murthy AC, Goggin SL, Eisch AJ, Oostra BA, Nelson
DL, Jin P and Zhao X: Ablation of Fmrp in adult neural stem cells
disrupts hippocampus-dependent learning. Nat Med. 17:559–565. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
229
|
Krueger DD, Osterweil EK, Chen SP, Tye LD
and Bear MF: Cognitive dysfunction and prefrontal synaptic
abnormalities in a mouse model of fragile X syndrome. Proc Natl
Acad Sci USA. 108:2587–2592. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
230
|
Stein JM, Bergman W, Fang Y, Davison L,
Brensinger C, Robinson MB, Hecht NB and Abel T: Behavioral and
neurochemical alterations in mice lacking the RNA-binding protein
translin. J Neurosci. 26:2184–2196. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
231
|
Chen-Plotkin AS, Lee VM and Trojanowski
JQ: TAR DNA-binding protein 43 in neurodegenerative disease. Nat
Rev Neurol. 6:211–220. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
232
|
Kwong LK, Uryu K, Trojanowski JQ and Lee
VM: TDP-43 proteinopathies: neurodegenerative protein misfolding
diseases without amyloidosis. Neurosignals. 16:41–51. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
233
|
Aparicio-Erriu IM and Prehn JH: Molecular
mechanisms in amyotrophic lateral sclerosis: the role of
angiogenin, a secreted RNase. Front Neurosci. 6:1672012.PubMed/NCBI
|
|
234
|
Ivanov P, Emara MM, Villen J, Gygi SP and
Anderson P: Angiogenin-induced tRNA fragments inhibit translation
initiation. Mol Cell. 43:613–623. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
235
|
Tadesse H, Deschênes-Furry J, Boisvenue S
and Côté J: KH-type splicing regulatory protein interacts with
survival motor neuron protein and is misregulated in spinal
muscular atrophy. Hum Mol Genet. 17:506–524. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
236
|
Glinka M, Herrmann T, Funk N, Havlicek S,
Rossoll W, Winkler C and Sendtner M: The heterogeneous nuclear
ribonucleoprotein-R is necessary for axonal beta-actin mRNA
translocation in spinal motor neurons. Hum Mol Genet. 19:1951–1966.
2010. View Article : Google Scholar : PubMed/NCBI
|