Introduction
The incidence of cancer continues to increase
largely due to aging and the increased adoption of cancer-causing
behavior, particularly smoking and obesity. Specifically,
colorectal cancer is the third most common cancer in males and the
second in females, with over 1.2 million new cases and 608,700
deaths in 2008 (1).
Histologically, colorectal cancer incidence rates are higher in
western countries, such as New Zealand, Europe and North America
than in Asian countries. However, the incidence rates are rapidly
increasing in several countries within Eastern Asia, such as Japan,
China and Singapore (2). Such
trends are thought to reflect a change in dietary patterns
(3). For this reason, the
interest in alternative medicine for the prevention and treatment
of colorectal cancer has increased and research of the effects of
various food extracts on colorectal cancer is in progress. Among
food extracts, quercetin (3,3′,4′,5,7-pentahydroxyflavone), which
is an extracted compound from green tea and red onion, has been
reported as being able to prevent a number of types of cancer,
including colorectal cancer (4).
In tumor cells, quercetin exerts a direct pro-apoptotic effect by
regulating caspase-3, -6 and -8, and the 5′ AMP-activated protein
kinase (AMPK)/cyclooxygenase-2 (COX-2) signaling pathway. In
addition, it can block the growth of cancer cells at different
phases of the cell cycle by controlling transcription factors, such
as p53 (5–7).
Sestrin 2 is a downstream effector of p53 and is
involved in the regulation of cell viability in a variety of
cellular stresses. Sestrin 2 expression is induced upon DNA damage,
such as UV-irradiation, dependent on p53, and oxidative stress,
such as hypoxia, independent of p53. Previous studies have
indicated that sestrin 2 expression in cancer cells suppresses cell
growth and proliferation by leading to the negative control of mTOR
through AMPKα1 phosphorylation (8,9).
In addition, sestrin 2 can function as an antioxidant agent under
low concentrations of hydrogen peroxide. This leads to the
regeneration of peroxiredoxin for the reduction of hydrogen
peroxide levels and the maintenance of cell viability through the
nuclear factor erythroid-derived 2-related factor 2 (Nrf-2)/ARE
signaling pathway; however, under high concentrations of hydrogen
peroxide, sestrin 2 induces apoptosis through the p53 signaling
pathway (10–13).
AMPK and serine/threonine protein kinase,
participate in an energy-sensing cascade that responds to the
deletion of adenosine triphosphate (ATP). AMPK is activated by
various upstream factors, such as liver kinase B1 (LKB1) or
Ca(2+)/calmodulin-dependent protein kinase kinase (CaMKK),
inhibited by mTOR, and leads to apoptosis by the activation of
Tuberous sclerosis 2 (TSC2) (14–16). Previous studies have indicated
that the activation of AMPK by quercetin-generated reactive oxygen
species (ROS) induces apoptosis through the apoptosis
signal-regulating kinase 1 (ASK1)/p38 MAPK pathway in MCF-7 breast
cancer cells (17). In addition,
activated AMPK by UV radiation or hydrogen peroxide has been shown
to lead to cell death through p38 MAPK activation (18). A number of studies have shown the
involvement of AMPK upstream pathways in a variety of conditions,
such as high glucose concentrations, ischemia and calcium
ion-dependent signals, or extracellular stress; however, the
activation of AMPK by any upstream signaling pathway under
stress-induced conditions, such as hydrogen peroxide or
flavonoid-generated ROS is not yet fully understood (19–21).
In the present study, we demonstrate that quercetin
promotes the generation of intracellular ROS and induces apoptosis
by decreasing mitochondrial membrane potential through the AMPK/p38
MAPK pathway and that these effects are dependent on sestrin 2
expression. Moreover, the activation of the sestrin 2/AMPK/p38
pathway induced by the quercetin generation of ROS occurred
independently of p53.
Materials and methods
Reagent
Quercetin, N-acetylcysteine (NAC),
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT),
dichloro-dihydro-fluorescein diacetate (DCFH-DA) and
3,3-dihexyloxacarbocyanine iodide (DiOC6) were all
purchased from Sigma-Aldrich (St. Louis, MO, USA). SB203580 (p38
MAPK inhibitor) and pifithrin-α were purchased from Calbiochem (San
Diego, CA, USA). The FITC-Annexin V apoptosis detection kit was
obtained from BD Pharmingen (San Diego, CA, USA). Specific
antibodies that recognized phosphorylated (p-)AMPKα1, AMPKα1, Bax,
caspase-3, cytochrome c and β-actin are obtained from Cell
Signaling Technology (Beverly, MA, USA) and sestrin 2 was purchased
from Proteintech (Chicago, IL, USA). p-p38 MAPK was purchased from
Signalway Antibody LLC (College Park, MD, USA).
Cell culture
HCT116 and HT-29 colon cancer cells were obtained
from the American Type Culture Collection (ATCC; Rockville, MD,
USA). The cells were grown in RPMI-1640 medium (HyClone, Waltham,
MA, USA) containing 10% fetal bovine serum (HyClone) and 1%
antibiotics (100 mg/l streptomycin and 100 U/ml penicillin) at 37°C
in a 5% CO2 atmosphere. The cells were suspended by
Trypsin-EDTA (HyClone) and separated at 1.5×105/ml per
plate, every 48 h.
Detection of intracellular ROS by
fluorescence microscopy
The cells were seeded 1×105/ml in a
12-well plate with coverglasses. Following treatment for the
indicated periods of time and doses at 37°C in a 5% CO2
atmosphere, the cells were incubated with 10 μM of DCFH-DA for 30
min and fixed with 3.7% formaldehyde for 20 min. The cells were
washed with PBS twice and fluorescence was detected under a
fluorescence microscope (Carl Zeiss, Thornwood, NY, USA).
Measurement of intracellular ROS
levels
The cells were seeded 1×106/ml in 100-mm
plate and incubated for 24 h. Following incubation, the cells were
treated with the test compound for 6 h at 37°C in a 5%
CO2 atmosphere. The cells were then incubated with 40 μM
of DCFH-DA for 30 min and harvested by trypsinization, collected by
centrifugation, washed with PBS twice, and resuspended in PBS. The
fluorescence intensity was analyzed using a flow cytometer (BD
Biosciences, Frankline Lakes, NJ, USA).
Cell proliferation assay (MTT assay)
The cells were seeded at 4,000/ml each well in a
96-well plate, and incubated for 24 h. Following incubation, the
cells were treated with the test compound and then incubated at
37°C in a 5% CO2 atmosphere. After 24 h, the cells were
incubated with 20 μl MTT (5 mg/ml with PBS) solution for 1 h. The
optical densities of the solution in each well were determined
using a microplate reader (Bio-Rad Laboratories, Inc., Tokyo,
Japan) at 595 nm.
Determination of apoptosis by Annexin
V/PI staining
The cells were seeded at 1×106/ml in
100-mm plate and incubated for 24 h. Following incubation, the
cells were treated with the test compound for 24 h at 37°C in a 5%
CO2 atmosphere. Total cells were harvested by
trypsinization, collected by centrifugation, washed with PBS, and
resuspended in binding buffer. Cells were stained with Annexin V
and PI for 15 min. Fluorescence intensity were analyzed using a
flow cytometer (BD Biosciences).
Determination of apoptosis by Hoechst
33342 staining
The cells were seeded at 1×104/ml in a
12-well plate with coverglasses. Following treatment at the
indicated doses, the cells were incubated with 10 μM Hoechst 33342
for 30 min and fixed with 3.5% formaldehyde for 20 min. The cells
were then washed twice with PBS, and fluorescence was measured
using a fluorescence microscope (Carl Zeiss).
Measurement of mitochondrial membrane
potential
The cells were seeded at 1×106/ml in a
100-mm plate and incubated for 24 h. Following incubation, the
cells treated with the test compound for 24 h at 37°C in a 5%
CO2 atmosphere. Total cells were harvested by
trypsinization, collected by centrifugation, washed with PBS, and
fixed with 70% ethanol. Cell were incubated with 100 ng/ml of
DiOC6 for 15 min at room temperature before being
analyzed under a flow cytometer (BD Biosciences).
Mitochondrial and cytosolic
fractions
We used the Mitochondria/Cytosol Fractionation kit
(Abcam plc, Cambridge, UK). The cells were seeded at
1×106/ml in 100-mm plate and incubated for 24 h.
Following incubation, the cells were treated with the test compound
for 24 h at 37°C in a 5% CO2 atmosphere. Total cells
were harvested by trypsinization, collected by centrifugation,
washed with PBS, and homogenized in ice-cold cytosol extraction
buffer mix containing DTT and protease inhibitor using a
homogenizer. The homogenates were centrifuged at 3,000 rpm for 10
min at 4°C and supernatants were collected. The supernatants were
centrifuged at 13,000 rpm for 30 min at 4°C and the collected
supernatants for cytosolic proteins and pellets were resuspended
with ice-cold mitochondrial extraction buffer containing DTT and
protease inhibitor for mitochondrial proteins.
Transient transfection with small
interfering RNA (siRNA)
siRNA was purchased from Dharmacon (Chicago, IL,
USA). For transient transfection, the cells were seeded
5×103/ml on a 6-well plate with antibiotics-free medium.
Following incubation overnight, targeting siRNA was transfected
using DharmaFECT1 transfection reagent (Dharmacon) according to the
manufacturer’s instructions. Following incubation for 72 h, the
cells were treated with quercetin for the indicated periods of
time.
Western blot analysis
The cells were seeded at 1×105/ml in a
6-well plate and incubated for 24 h. Following incubation, the
cells were treated with the test compound for 6 h at 37°C in a 5%
CO2 atmosphere. The cells were then rinsed twice with
ice-cold PBS and scraped with lysis buffer (50 mM Tris-HCl pH 8.0,
150 mM NaCl, 1% NP-40, 0.5% sodium deoxycholate, 1 mM PMSF) and
subjected to western blot analysis. The primary antibody was then
added following by overnight incubation at 4°C; following the
addition of the secondary antibody, the cells were reacted for 75
min at room temperature with gentle agitation.
Statistical analysis
Cell viability was statistically analyzed using an
unpaired t-test (SPSS Inc.; Chicago, IL, USA). A value of P<0.05
was considered to indicate a statistically significant
difference.
Results
Quercetin generates intracellular ROS
production in HCT116 colon cancer cells
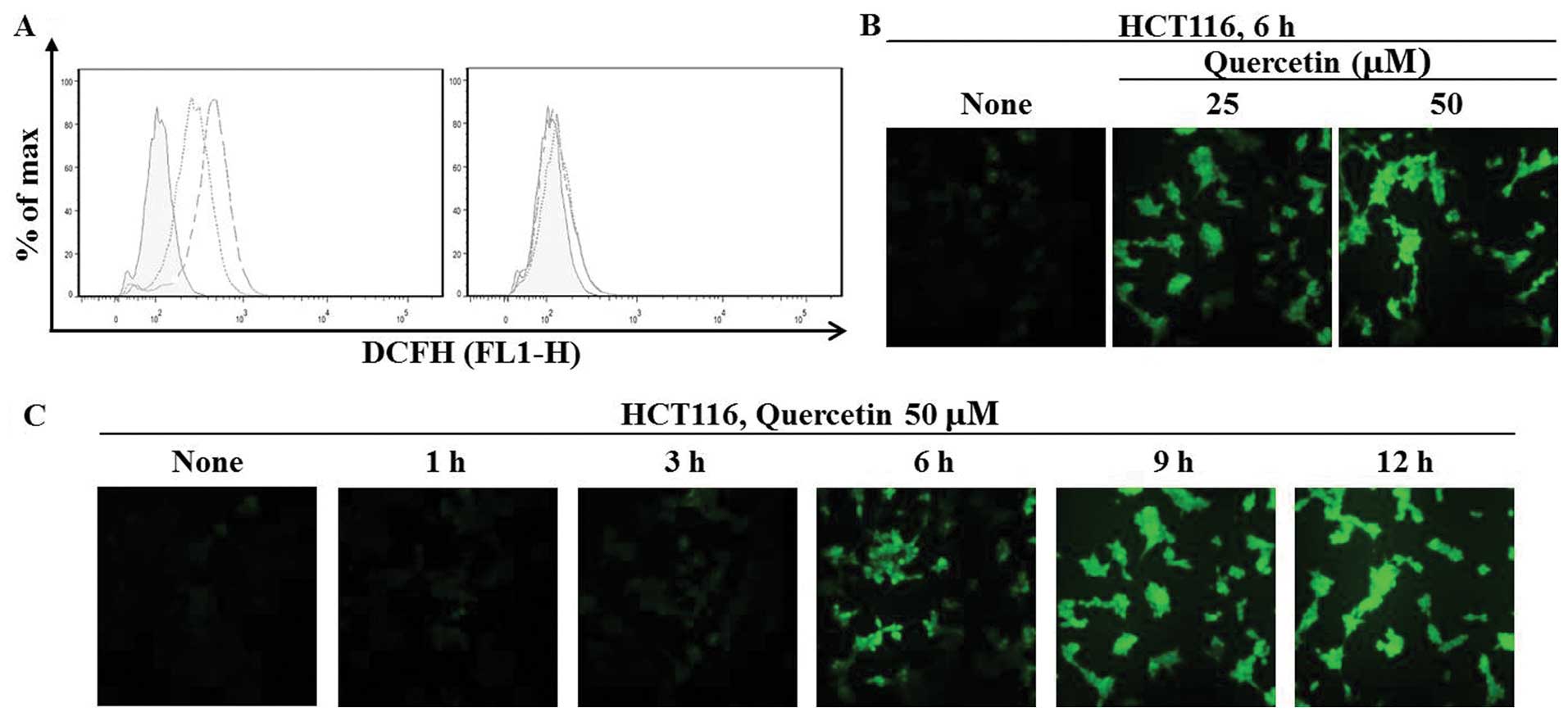
To examine whether quercetin promotes the generation
of ROS in HCT116 colon cancer cells, we measured the intracellular
ROS levels following treatment of the cells with quercetin (25 μM
or 50 μM) for 6 h. As shown in Fig.
1A, quercetin increased ROS levels at the indicate
concentrations (left panel). These effects were completely blocked
by combined treatment with NAC, a ROS scavenger (right panel). We
also observed intracellular ROS levels under a fluorescence
microscope following staining with DCFH-DA. The quercetin-induced
the generation of ROS continuously in a dose and time-dependent
manner (Fig. 1B and C).
Quercetin suppresses cell proliferation
and induces apoptosis
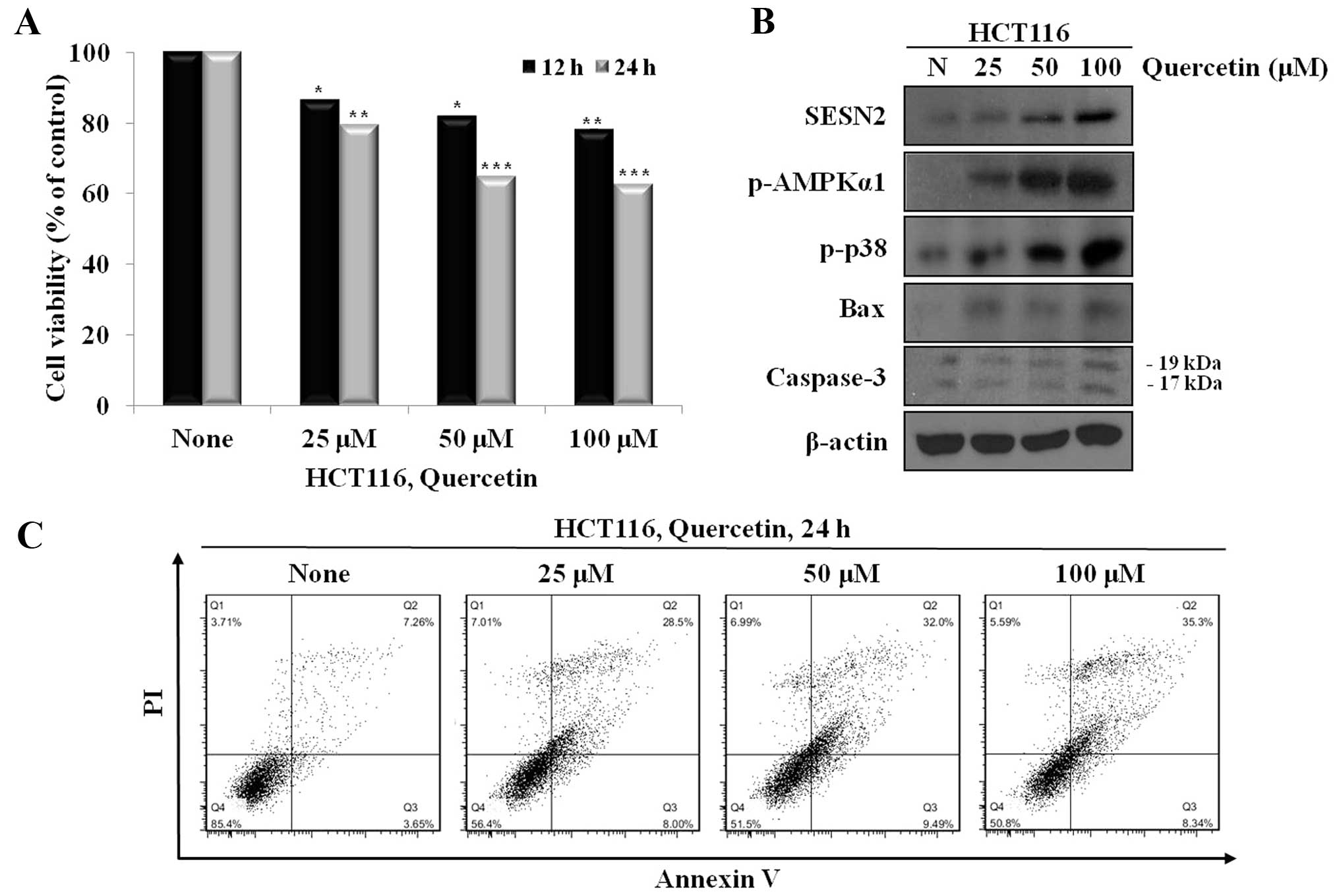
We investigated the anti-proliferative and apoptotic
effects of quercetin through the increase in intracellular ROS. For
this reason, we treated the cells with quercetin (25, 50 and 100
μM) for 24 h, and the viability and apoptosis of the cells were
then examined. The cells treated with quercetin showed a decrease
in viability and an increase in the number of Annexin V-positive
cells in a dose-dependent manner (Fig. 2A and C).
Quercetin regulates the expression of
sestrin 2 and AMPK, and p38 activation
To examine the mechanisms through which quercetin
induces apoptosis, we first examined the mechanisms through which
quercetin increases sestrin 2 expression and activates AMPKα1 and
p38. We analyzed the changes in the levels of sestrin 2, p-AMPKα1
and p-p38, as well as apoptosis-related proteins, such as Bax and
caspase-3 following treatment with quercetin at different
concentrations by western blot analysis. Our results revealed that
quercetin markedly increased the expression of sestrin 2 and
activated AMPK and p38 in a dose-dependent manner. In addition, we
also observed the increased expression of Bax and the cleavage of
caspase-3 (Fig. 2B).
Quercetin modulates the expression of
sestrin 2 and AMPK, and the activation of p38 through the
generation of intracellular ROS
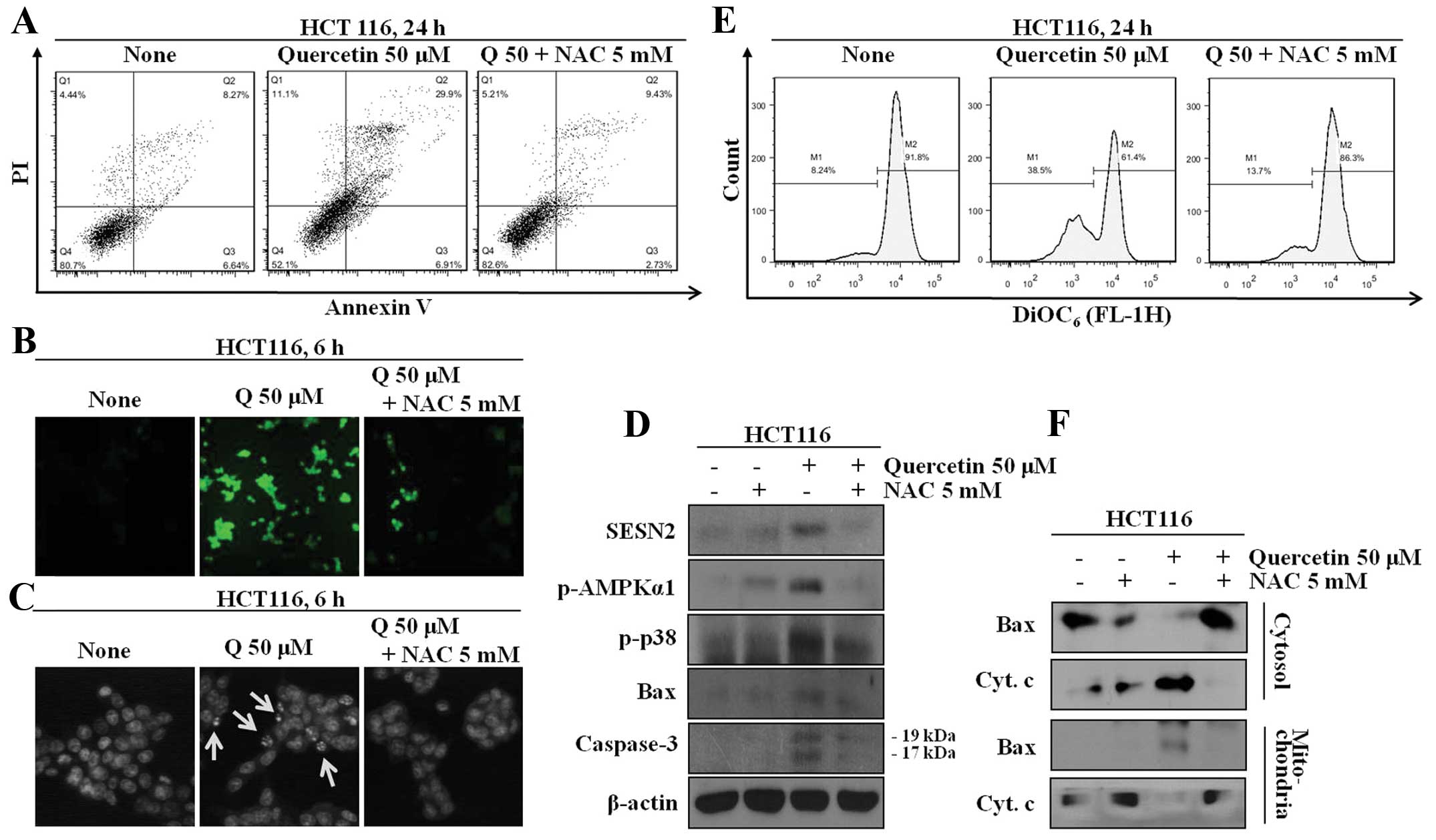
To determine whether the quercetin-induced increase
in the expression of sestrin-2 and AMPK and the induction of
apoptosis are involved in the increase in intracellular ROS levels,
the cells wer co-treated with NAC and an quercetin; the proteins
levels and the number of and Annexin V-positive cells were then
determined. The cells co-treated with quercetin and NAC displayed
decreased expression levels of sestrin 2, AMPK and p38
phosphorylation. The quercetin-treated group displayed increased
apoptotic cell death through the regulation of mitochondrial
membrane potential, leading to the secretion of cytochrome
c, which is a marker protein for apoptosis, from the
mitochondria to the cytosol (Fig.
3).
Sestrin 2 is an important element for the
induction of apoptosis and the activation of the AMPK/p38/BAX
signaling pathway
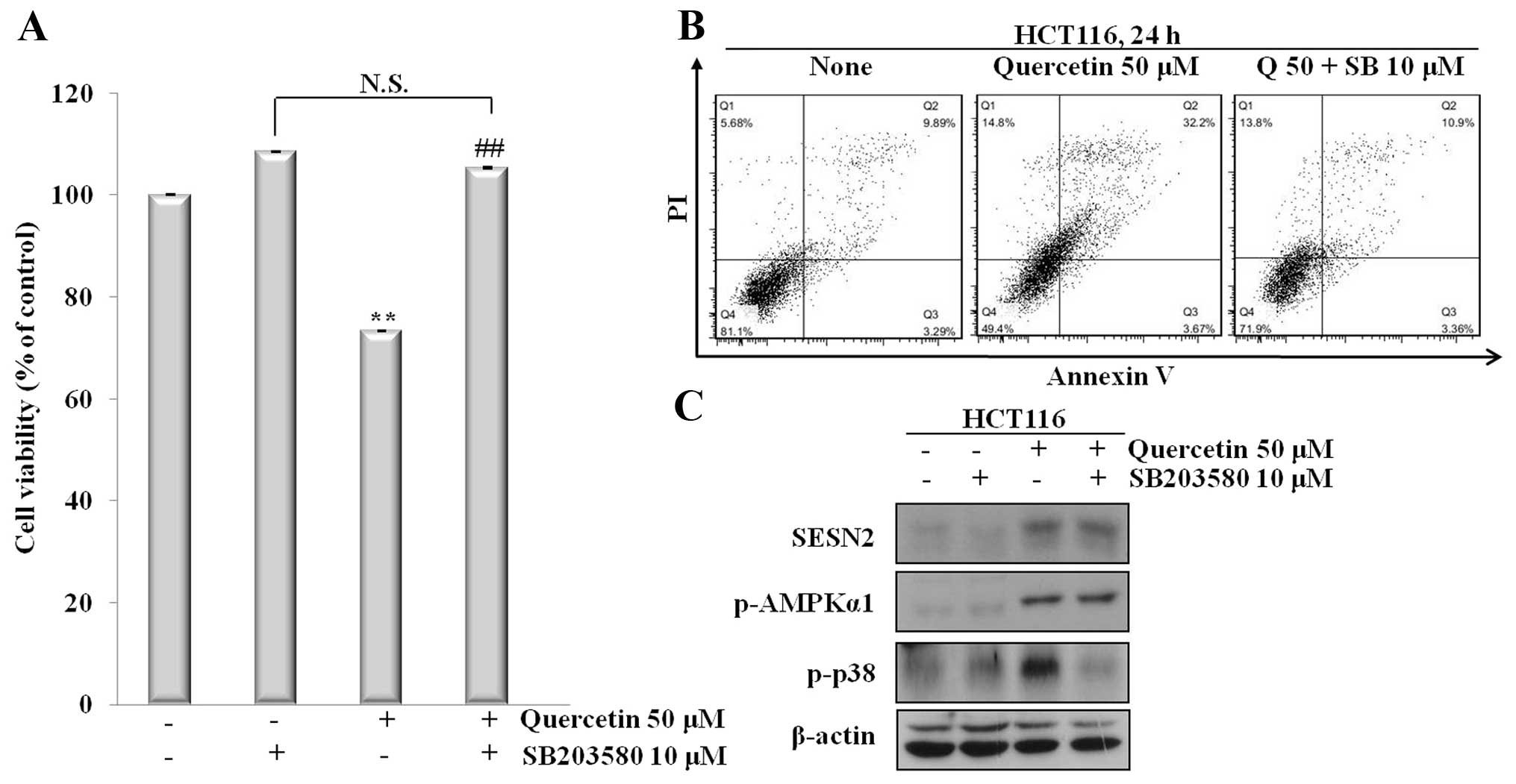
To determine whether the quercetin-induced apoptosis
was dependent on sestrin 2 expression, and whether the activation
of the AMPK/p38 signaling pathway was involved, we co-treated the
cells with quercetin and SB203580, a p38 inhibitor. The cells were
then analyzed for apoptotic cell death by Annexin V/PI staining and
the proteins levels were determined by western blot analysis.
Co-treatment with the inhibitor led to reduced cell death; however,
the cells treated with quercetin displayed increased apoptotic cell
death. In addition, co-treatment with quercetin and SB203580
induced an increase in the expression of sestrin 2 and AMPK
phosphorylation (Fig. 4). Of
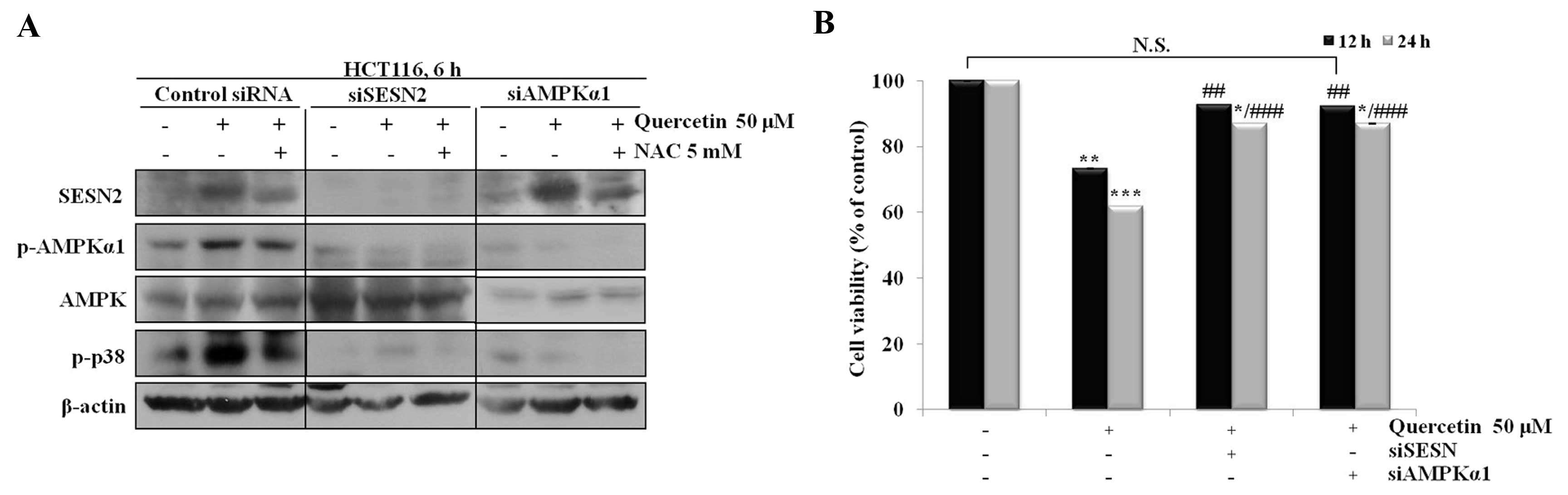
note, quercetin did not regulate the AMPK/p38 signaling pathway and
did not reduce cell viability when sestrin 2 was silenced using
siRNA (Fig. 5).
Sestrin 2 is expressed and induces
apoptosis in p53-negative cells
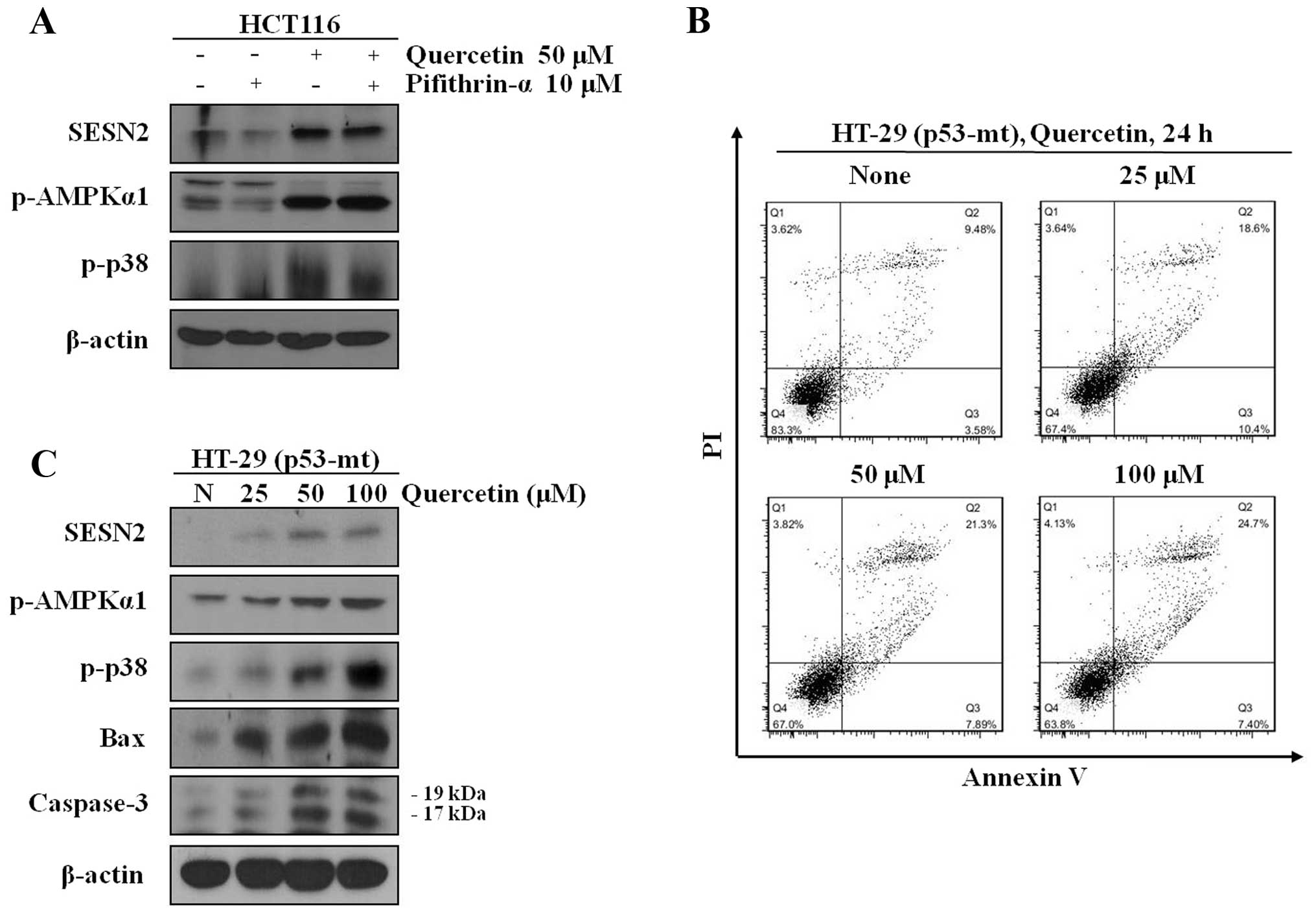
To examine the expression of sestrin 2 independent
of p53, we co-treated the cells with quercetin and pifithrin-α, a
p53-dependent transactivity inhibitor. Co-treatment with quercetin
and pifithrin-α led to an increase in the expression of sestrin 2,
AMPK and p38 phosphorylation. In addition, our results revealed
that sestrin 2 expression levels, as well as those of AMPK and p38
phosphorylation were increased in the HT-29, which are p53 mutant
cells. Moreover, quercetin induced apoptotic cell death in the
HT-29 cells (Fig. 6).
Discussion
The incidence of colorectal cancer has increased due
to changes in dietary patterns. For this reason, there has been an
increased interest in the effects of food extracts on the
prevention and treatment of colorectal cancer. These foods extracts
induce apoptosis and cell cycle arrest through the regulatkion of
intracellular protein signals in cancer cells, leading to abnormal
cell proliferation. Various foods extracts have been used in
chemotheraphy experiments. This is particularly the case with
quercetin, which is a polyphenolic compound extracted from red
onion and green tea, and is known to have diverse pharmacological
activities, including anticancer, anti-inflammatory and
anti-proliferative activities (4,22).
Recently, a study found that quercetin generates intracellular ROS
and induces apoptosis through controlling the AMPK/ASK1/p38 pathway
in MCF-7 breast cancer cells (17). Moreover, in a previous study, when
HT-29 colon cancer cells were treated with quercetin, apoptosis was
induced through the regulation of the AMPK/COX-2 pathway (6). It has been suggested that quercetin
induces apoptosis through the activation of AMPK. However, the
activation of AMPK by any upstream factors following treatment with
quercetin is not clear. According to certain studies, the
transcription of p53 induces AMPK phosphorylation; however, these
results cannot explain the phosphorylation of AMPK in p53 mutant
cells (23,24). However, as previously
demonstrated, a specific type of stress, such as oxidative stress
or DNA damage stimulates sestrin 2 transcription in cancer cells,
and this induces the phosphorylation of AMPK through the
interaction with AMPK (8,11). According to these studies, the
inhibition of mTOR activity and cell cycle arrest does not occur
through AMPK phosphorylation when sestrin 2 transcription is
suppressed. Thus, we hypothesized that the quercetin-induced
generation of intracellular ROS and the induction of apoptosis
through AMPK activation are dependent on sestrin 2 expression. Our
results revealed that quercetin suppressed proliferation and
induced apoptotic cell death by increasing the expression of
sestrin 2. Moreover, quercetin, by increasing intracellular ROS
production, can transcript sestrin 2 directly, and the upstream
function in the AMPK/p38 signaling pathway was confirmed.
Importantly, it is known that approximately 50% of
cancer cells are p53 mutant (25). Thus, a p53-independent manner for
the induction of apoptosis is very important in cancer prevention
studies. Our results revealed that not only does quercetin regulate
the AMPK/p38 signaling pathway by increasing sestrin 2 expression,
but it induces the apoptosis of HT-29 colon cancer cells, which are
p53 mutant cells.
In this study, we demonstrate that quercetin induces
apoptosis through the activation of the ROS/AMPK/p38 pathway.
During this process, the increased expression of sestrin 2 induced
by quercetin was found to be crucial. The silencing of sestrin 2
silencing using siRNA resulted in the deactivation of AMPK and p38
and did not reduced cell viability following treatment with
quercetin. Furthermore, quercetin induced the generation of
intracellular ROS and induced apoptosis by regulating the sestrin
2/AMPK/p38 pathway in p53 mutant cells. From these results, it can
be suggested that the quercetin-induced apoptosis is carried out
through the sestrin 2/AMPK/p38 signaling pathway, and that sestrin
2 is an important regulator of the AMPK/p38 pathway in a
p53-independent manner.
Acknowledgements
This study was supported by the Korea Research
Foundation Grant (KRF-2010-0021402) and the Hannam University Fund
2013.
References
|
1
|
Jemal A, Bray F, Center MM, et al: Global
cancer statistics. CA Cancer J Clin. 61:69–90. 2011. View Article : Google Scholar
|
|
2
|
Center MM, Jemal A and Ward E:
International trends in colorectal cancer incidence rates. Cancer
Epidemiol Biomarkers Prev. 18:1688–1694. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Giovannucci E: Modifiable risk factors for
colon cancer. Gastroenterol Clin North Am. 31:925–943. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Gibellini L, Pinti M, Nasi M, et al:
Quercetin and cancer chemoprevention. Evid Based Complement
Alternat Med. 2011:5913562011.PubMed/NCBI
|
|
5
|
Chien SY, Wu YC, Chung JG, et al:
Quercetin-induced apoptosis acts through mitochondrial- and
capase-2-dependent pathways in human breast cancer MDA-MB-231
cells. Hum Exp Toxicol. 28:493–503. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Lee YK, Park SY, Kim YM, et al: AMP
kinase/cyclooxygenase-2 pathway regulates proliferation and
apoptosis of cancer cells treated with quercetin. Exp Mol Med.
41:201–207. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Tanigawa S, Fujii M and Hou DX:
Stabilization of p53 is involved in quercetin-induced cell cycle
arrest and apoptosis in HepG2 cells. Biosci Biotechnol Biochem.
72:797–804. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Budanov AV and Karin M: p53 target genes
Sestrin1 and Sestrin2 connect genotoxic stress and mTor signaling.
Cell. 134:451–460. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Sanli T, Linher-Melville K, Tsakiridis T,
et al: Sestrin2 modulates AMPK subunit expression and its response
to ionizing radiation in breast cancer cells. Plos One.
7:e320352012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Budanov AV, Lee JH and Karin M: Stressin’
Sestrins take an aging fight. EMBO Mol Med. 2:388–400. 2010.
|
|
11
|
Budanov AV, Shoshani T, Faerman A, et al:
Identification of a novel stress-responsive gene Hi95 involved in
regulation of cell viability. Oncogene. 21:6017–6031. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Shin BY, Jin SH, Cho IJ, et al: Nrf2-ARE
pathway regulates induction of Sestrin-2 expression. Free Radic
Biol Med. 53:834–841. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Sablina AA, Budanov AV, Ilyinskaya GV, et
al: The antioxidant function of the p53 tumor suppressor. Nat Med.
11:1306–1313. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Abbott MJ, Edelman AM and Turcotte LP:
CaMKK is an upstream signal of AMP-activated protein kinase in
regulation of substrate metabolism in contracting skeletal muscle.
Am J Physiol Regul Integr Comp Physiol. 297:1724–1732. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Alexander A and Walker CL: The role of
LKB1 and AMPK in cellular responses to stress and damage. FEBS
Lett. 585:952–957. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shaw RJ, Kosmatka M, Bardeesy N, et al:
The tumor suppressor LKB1 kinase directly activates AMP-activated
kinase and regulates apoptosis in response to energy stress. Proc
Natl Acad Sci USA. 9:3329–3335. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Lee YK, Hwang JT, Kwon DY, et al:
Induction of apoptosis by quercetin is mediated through
AMPKα1/ASK1/p38 pathway. Cancer Lett. 292:228–236. 2010.
|
|
18
|
Cao C, Lu S, Kivlin R, et al:
AMP-activated protein kinase contribute to UV− and H2O2− induced
apoptosis inhuman keratinocytes. J Biol Chem. 283:28897–28908.
2008.
|
|
19
|
Capano M and Crompton M: Bax translocates
to mitochondria of heart cells during simulated ischaemia:
involvement of AMP-activated and p38 mitogen-activated protein
kinases. Biochem J. 395:57–64. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Park S, Scheffler TL, Rossie SS, et al:
AMPK activity is regulated by calcium-mediated protein phosphatase
2A activity. Cell Calcium. 53:217–223. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jones RG, Plas DR, Kubek S, et al:
AMP-activated protein kinase induces a p53-dependent metabolic
checkpoint. Mol Cell. 29:283–293. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mouria M, Gukovskaya AS, Jung Y, et al:
Food-derived polyphenols inhibit pancreatic cancer growth through
mitochondrial Cytochrome c release and apoptosis. Int J Cancer.
98:761–769. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Feng Z, Zhang H, Levine AJ, et al: The
coordinate regulation of the p53 and mTOR pathways in cells. Proc
Natl Acad Sci USA. 102:8204–8209. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lee SO, Andey T, Jin UH, et al: The
nuclear receptor TR3 regulates mTORC1 signaling in lung cancer
cells expressing wild-type p53. Oncogene. 31:3265–3276. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Olivier M, Hollstein M and Hainaut P: TP53
mutation in human cancers: origins, consequences, and clinical use.
Cold Spring Harb Perspect Biol. 2:a0010082010. View Article : Google Scholar : PubMed/NCBI
|