Introduction
Cell proliferation, neuronal migration and cortical
organization represent three important stages of brain maturation
(1). Cortical development is
critical for brain maturation as cortical malformations are
increasingly recognized as causes of severe epileptic syndrome,
neuropsychological disorders and mental retardation (2,3).
Moreover, the incorrect positioning of cortical neurons following
cortical cell migration often leads to cortical dysplasia.
Therefore, the identification of regulatory factors responsible for
brain development and maturation is important for understanding the
pathogenesis of cortical abnormalities.
Lis1 is a brain-specific gene that encodes
for the non-catalytic subunit of platelet-activating factor
acetylhydrolase isoform 1B (PAFAH1B), which inactivates
platelet-activating factor (PAF) (4,5).
Lis1 is known to regulate cell proliferation and migration
during brain development through its interaction with proteins,
such as dynein (6). Subjects with
Miller-Dieker syndrome (MDS) or isolated lissencephaly sequence
(ILS) have a hemizygous deletion or mutation of the Lis1
gene (7,8). ILS and MDS often result from
haploinsufficiency at human chromosome 17p13.3, a chromosomal
region that includes the Lis1 gene. The disruption of
Lis1 in patients with ILS and MDS (9) suggests that mutations within
Lis1 are responsible for defective neuronal migration.
microRNAs (miRNAs or miRs) are a class of small,
non-coding, regulatory RNA molecules (10). Over the past decade, research has
identified important regulatory roles for miRNAs in cell
development, differentiation, proliferation, apoptosis and
metabolism, as well as in the pathogenesis of several diseases
(11). Approximately 70% of all
known miRNAs are expressed in the mammalian brain, and the levels
of many miRNAs are dramatically altered during brain development
(12). However, the roles of
miRNAs in the regulation of mammalian brain development are still
poorly defined (13).
Further knowledge of the molecular mechanisms
underlying cortical neuronal migration may provide insight into
improved therapeutic options for the treatment of malformations of
cortical development. In this study, we examined miRNA expression
profiles in immature rats with liquid nitrogen lesion-induced focal
cortical dysplasia. Our aim was to identify the miRNAs that
modulate cortical neuronal migration. We identified and
characterized miR-139-5p, indicating that the loss of miR-139-5p
regulates cortical neuronal migration through the modulation of
Lis1 expression.
Materials and methods
miRNA microarray analysis
RNA labeling and hybridization to miRNA microarray
chips were performed as previously described (14). Whole brain tissues from immature
Sprague-Dawley (20–80 days) rats were pooled and total RNA was
extracted using TRIzol (Invitrogen, Shanghai, China). Briefly, 50
mg of total RNA were purified using the mirVana miRNA isolation kit
(Ambion, Austin, TX, USA) resulting in a small, enriched RNA
fraction. Purified RNA was labeled with Cy3 and hybridization was
carried out using a miRNA microarray chip (CapitalBio Corp.,
Beijing, China) containing 381 probes in triplicate.
Quantitative reverse
transcription-polymerase chain reaction (qRT-PCR) of
miR-139-5p
We performed miRNA qRT-PCR as previously described
(15). Briefly, rat brain RNA (1
μg) was reverse transcribed with a stem-loop reverse transcriptase
primer and quantitative PCR (qPCR) was then performed. The program
was initially 2 min at 95°C, followed by 40 cycles of 30 sec at
95°C, and 60 sec at 60°C. The primers used for miR-139-5p qRT-PCR
were as follows: stem-loop RT primer,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTG AGAGACACGT-3′; and the qPCR
primers: miR-139-5p forward, 5′-ACACTCCAGCTGGGTCTACAGTGCAC-3′ and
reverse, 5′-TGGTGTCGTGGAGTCG-3; and U6 forward, 5′-GCTTC
GGCAGCACATATACTAAAAT-3′ and reverse, 5′-CGCTT
CACGAATTTGCGTGTCAT-3′.
Analysis of miR-139-5p predicted
targets
The prediction of miR-139-5p targets was performed
using the following algorithms: PicTar (http://pictar.mdc-berlin.de/), TargetScan, (http://www.targetscan.org/vert_50/) and miRanda
(http://www.ebi.ac.uk/enright-srv/microcosm/cgi-bin/targets/v5/mirna.pl).
Cell culture and transfection
PC12 cells were maintained in DMEM high glucose
medium, supplemented with 10% fetal bovine serum (FBS) (both from
Gibco, Carlsbad, CA, USA) and 5% horse serum. The cells were
cultured in a humidified incubator at 37°C with 5% CO2.
The PC12 cells were transfected with 50 nM of either a
non-targeting small RNA oligonucleotide (GenePharma Co., Ltd.,
Shanghai, China) as a negative control (control), or miR-139-5p
mimics (stably enhanced miR-139-5p oligonucleotide; GenePharma Co.,
Ltd.) using Lipofectamine™ 2000 (Invitrogen, Carlsbad, CA, USA), as
previously described (16). The
transfected cells were harvested after 48 h, RNA was extracted from
the cells using TRIzol reagent (Invitrogen, Shanghai, China), and
proteins were extracted using lysis buffer [20 mM Tris (pH 7.4), 1
mM EDTA, 1% Triton X-100, protease inhibitors] (GE Healthcare Life
Sciences, Shanghai, China), as previously described (17). The subsequent qRT-PCR and western
blot analysis results were obtained from three separate
transfections.
qRT-PCR of Lis1
The mRNA qRT-PCR analyses were conducted as
previously described (18).
Briefly, RNA (1 μg) obtained from the rat brains or PC12 cells was
reverse transcribed using oligo(dT)18 followed by qPCR. The program
was initially run for 2 min at 95°C, followed by 40 cycles of 30
sec at 95°C, and 60 sec at 60°C. The primers used for Lis1 qRT-PCR
were as follows: Lis1 forward, 5′-TGCCCAAGACTACTCAACCC-3′ and
reverse, 5′-GCACCCTGTGACGAAAGC-3′; and 18S RNA, forward,
5′-AGCAACTGCGCCTGAAAC-3′ and reverse, 5′-CCCTGTCCCGCTCAACTA-3′.
Western blot analysis for Lis1
The cells were rinsed once with phosphate-buffered
saline (PBS) then lysed in lysis buffer [50 mmol/l Tris-HCl (pH
7.5), 5 mmol/l EDTA, 1% (V/V) Triton X-100 and 0.15 mol/l NaCl] on
ice for 10 min. Insoluble components were removed by centrifugation
(12,000 rpm for 5 min), and the protein concentration was measured,
as previously described (19).
Cellular proteins (100 μg) were adjusted to a total concentration
of 5 μg protein/μl. After boiling for 5 min in loading buffer,
proteins were separated by 8% Tris-glycine gels for Lis1. Western
blot analysis was performed using mouse anti-rat Lis1 antibody
(Abcam Biotech, Cambridge, MA, USA) and goat anti-mouse HRP-linked
secondary antibody. Immunocomplexes were visualized using the
LumiGLO® chemiluminescent detection kit (Cell Signaling
Technology, Boston, MA, USA) according to the manufacturer’s
instructions.
Luciferase targeting assay and Transwell
assay
We cloned 400 bp of the Lis1 3′ untranslated region
(3′UTR) containing the 7-bp target site for miR-139-5p into the
SpeI/HindIII sites of a luciferase gene in the
pMIR-REPORT luciferase vector (Ambion, Shanghai, China). PCR
analyses were performed using rTaq polymerase (Takara Bio Inc.,
Japan). The PC12 cells (5×104) were co-transfected with
150 ng of pMIR-REPORT, Lis1-3′UTR plasmid and 25 nM of either a
stably-enhanced non-targeting small RNA oligonucleotide as a
negative control (control), or miR-139-5p mimics (both from
GenePharma Co., Ltd.) using Lipofectamine™ 2000 (Invitrogen,
Shanghai, China), as previously described (20,21). Cells transfected with pre-scramble
miRNA or or anti-scramble miRNA were also used as negative
controls. The transfected cells were harvested after 24 h and then
assayed using a Dual-Luciferase Reporter assay system (Promega,
Madison, WI, USA). The results were obtained from three separate
experiments with each one conducted in triplicate.
A Transwell migration assay was performed as
previously described (22). The
cells (2×105) were seeded onto a 35-mm dish one day
prior to transfection. The transfection protocol was conducted
according to the cell culture and transfection protocols previously
described (23). After 24 h of
transfection, 5 ng/ml of Lis1 protein were added to the cell
culture medium. The PC12 and HCN-2 cells that had migrated onto the
membrane were fixed with methanol and stained with crystal violet
after 24 h. Images of randomly selected fields of the fixed cells
were acquired and the cells were counted. Experiments were repeated
in triplicate.
Rat model of focal cortical
dysplasia
Thirty-three days after birth, 30 Sprague-Dawley
rats were obtained from the Laboratory Animal Center of Nanjing
University (Nanjing, China). All animals were housed in a specific
pathogen-free (SPF) facility and received human care according to
the Chinese legal requirements. The experimental animal procedures
were approved by the Nanjing Medical University Institutional
Animal Care and Use Committee. All experiments were performed
according to the guidelines of the European Community Council.
The rat model of liquid nitrogen lesion-induced
focal cortical dysplasia was established as previously described
(24). Twenty Sprague-Dawley rats
with cortical dysplasia were randomly divided into two groups: the
model group and the miR-139-5p administration group. In the model
group, brain tissue was obtained on days 0, 20, 60 and 80. In the
miR-139-5p administration group, miR-139-5p was injected into the
rat brains using a rat brain locator at a 5 mg/kg dose on days 40
and 50, and brain tissue was obtained on day 60. The control
(healthy; n=10) group was administered the same volume of PBS. On
day 60, brain tissue was rapidly removed and prepared for
hematoxylin and eosin (H&E) and Nissl staining.
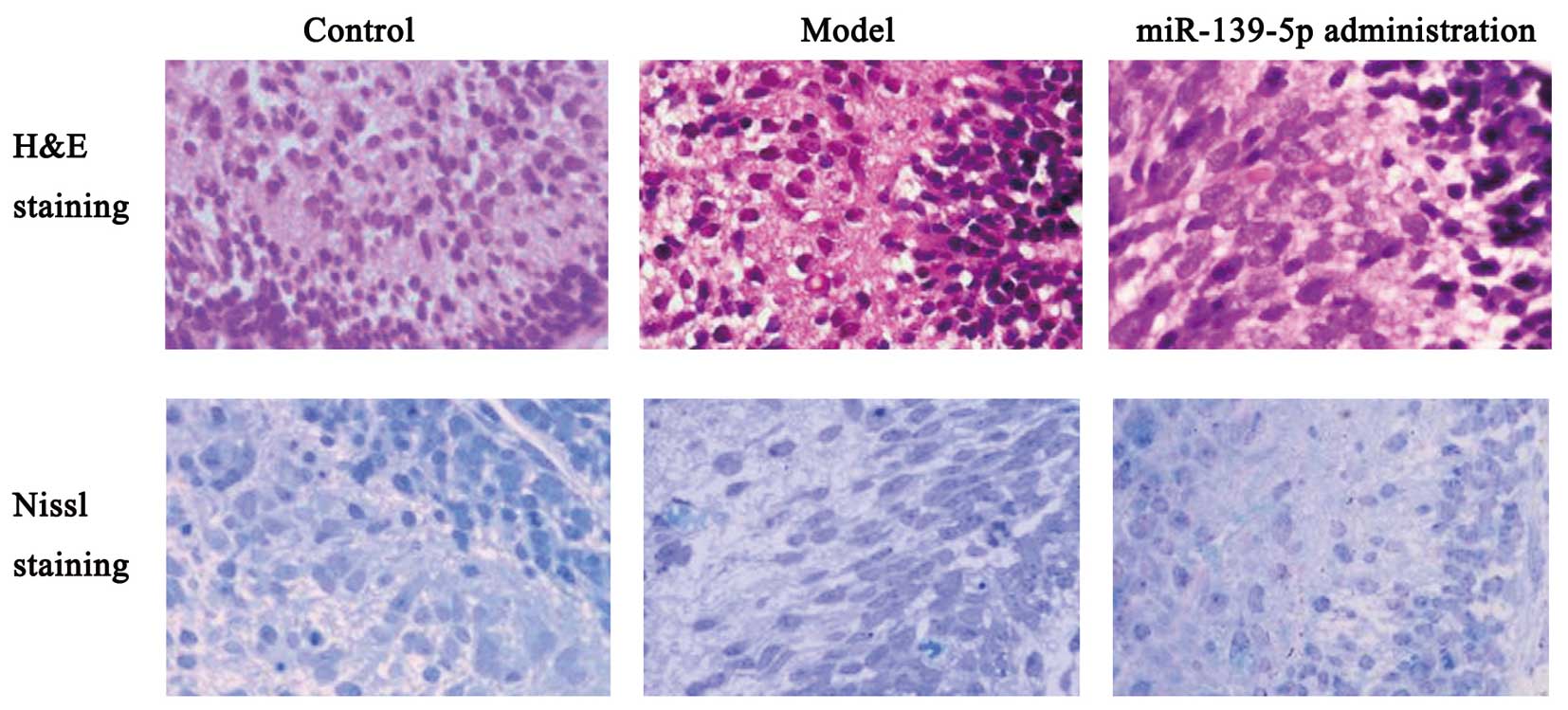
H&E and Nissl staining
The rat brain tissues were rapidly fixed in 10%
formaldehyde and embedded in paraffin. Sections (4-μm-thick) were
mounted for H&E and Nissl staining. Brain tissue morphological
characteristics and differences between the three experimental
groups were observed under a microscope (Nikon, Tokyo, Japan).
Statistical analysis
All experiments were performed in triplicate.
Hierarchical cluster analysis was carried out using Gene Cluster
software (Stanford University, Stanford, CA, USA). Comparisons
between groups were made by a Student’s t-test. A P-value <0.05
was considered to indicate a statistically significant difference,
as previously described (25).
Results
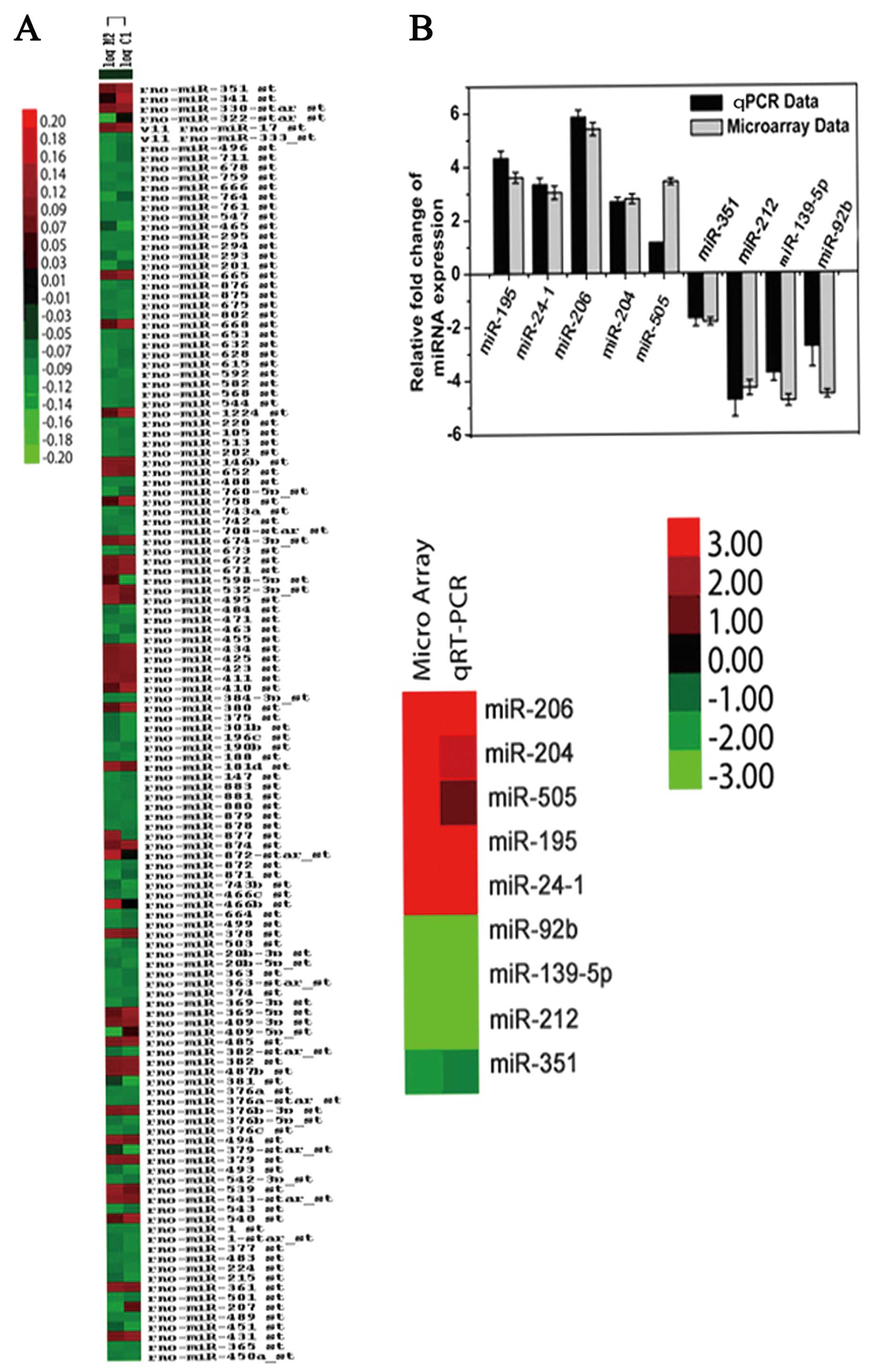
Cluster analysis for microarray data and
qRT-PCR validation
We examined the expression profiles of miRNAs in
immature rats with liquid nitrogen lesion-induced focal cortical
dysplasia. The dendrogram generated by cluster analysis revealed
the separation of the model of dysplasia from the control samples
on the basis of miRNA profiling (Fig.
1A). The relative changes in miRNA expression as shown by the
qRT-PCR data were in agreement with the microarray data (Fig. 1B). These results indicate a
downregulation in miR-139-5p expression during rat brain
development.
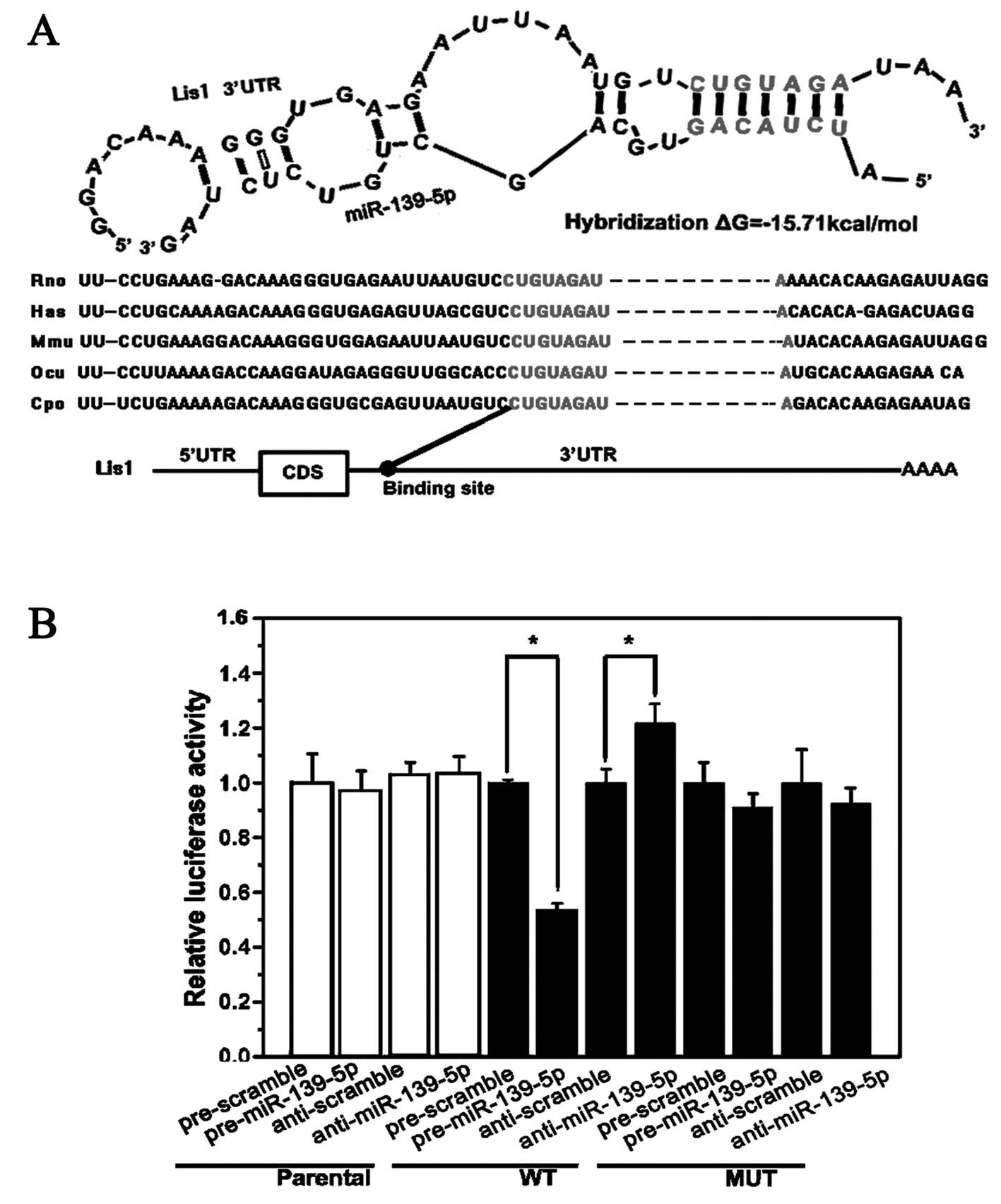
miR-139-5p targets Lis1 mRNA as shown by
luciferase assay
Multiple algorithms (TargetScan, miRanda and PicTar)
for predicting the putative targets of miR-139-5p identified
several potential targets, such as Lis1, Capn8,
Gmfb, Mapk1, Dclk1, Vim, Mgst1,
Gnb1, Klf15 and Cdkn1b. Lis1 was selected for
further examination due to its established role in cell
proliferation and migration, as well as its role in brain
development (26). Bioinformatic
analysis for the target site of miR-139-5p in the Lis1 3′UTR
revealed that mature miR-139-5p shares the same sequence in rats,
mice and humans (Fig. 2A). There
is one 7 bp target site in the Lis1 3′UTR which is conserved
in mammals. Wild-type or mutant Lis1 3′UTR was constructed
in the luciferase reporter plasmid for conducting luciferase
targeting assays (Fig. 2B) as it
contains the 7-bp target site for miR-139-5p and the mutant 7-bp
site. The luciferase activity of the wild-type plasmid was
suppressed by approximately 42% at 24 h in the PC12 cells
transfected with miR-139-5p mimics, while the activity of the
mutant plasmid was not suppressed (Fig. 2B).
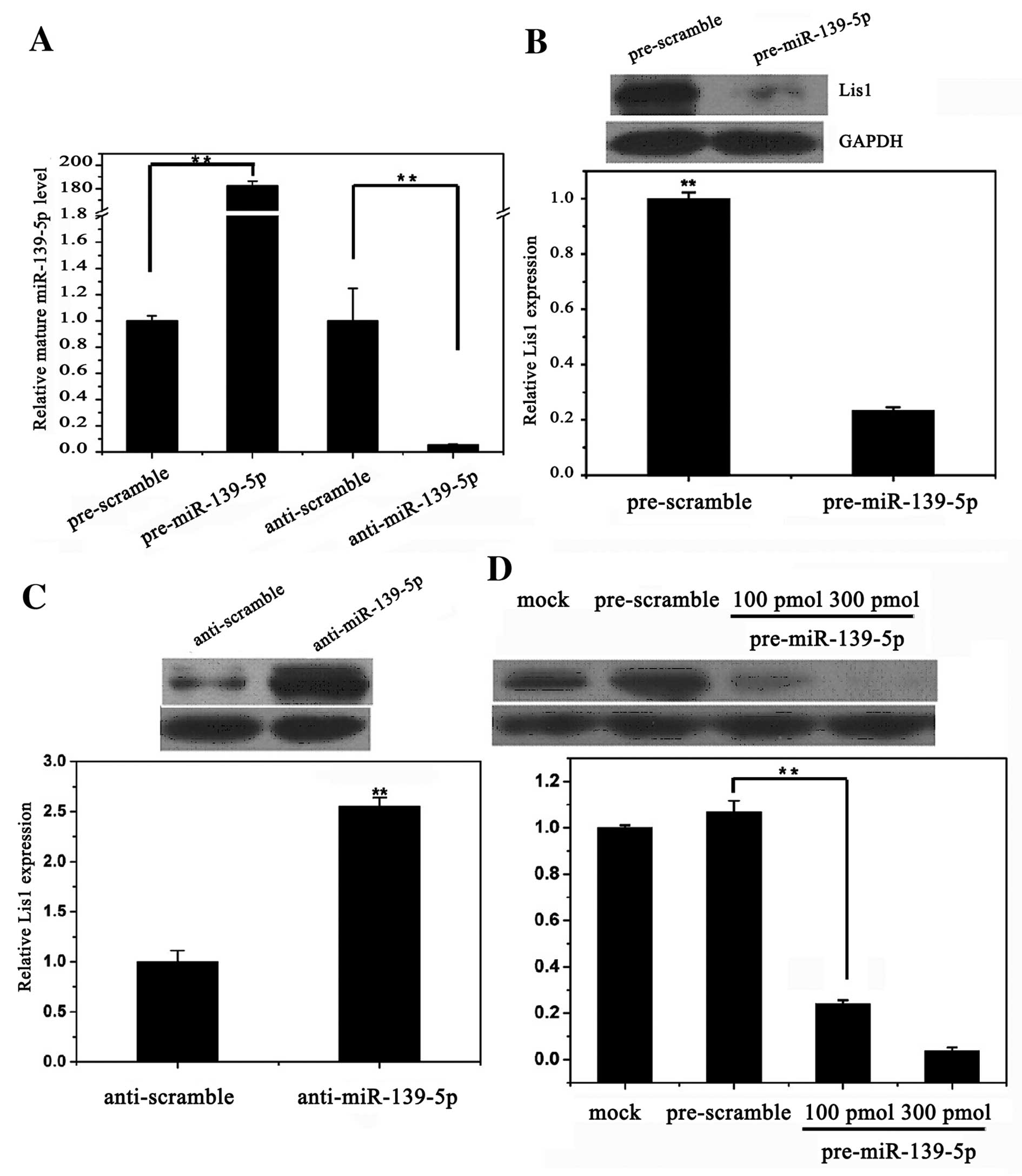
Correlation between the expression of
miR-139-5p and Lis1 in PC12 cells
We confirmed that miR-139-5p was successfully
transfected into the PC12 cells (Fig.
3A). There was an inhibitory effect of miR-139-5p on the
expression of Lis1 in the PC12 cells transfected with
pre-miR-139-5p after 24 h (Fig.
3B). Transfection with miR-139-5p inhibited the protein
expression of Lis1 by approximately 60 and 90% by using 100 and 300
pmol pre-miR-139-5p, respectively (Fig. 3D). By contrast, there was a
promotional effect of anti-miR-139-5p on the expression of Lis1 in
the PC12 cells transfected with anti-miR-139-5p (Fig. 3C).
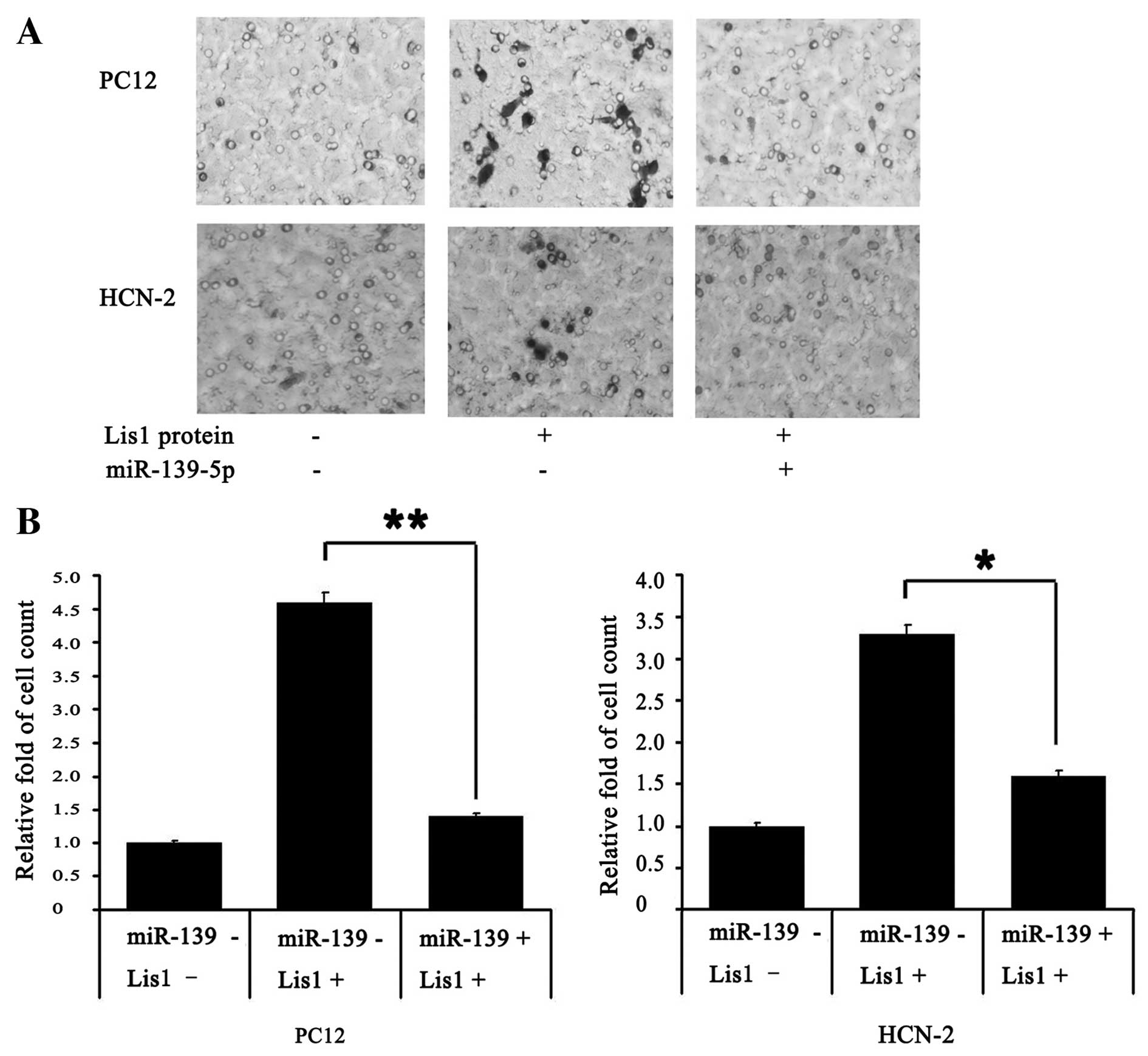
miR-139-5p inhibits cell migration
Transwell assays revealed that miR-139-5p
significantly inhibited the migration of PC12 and HCN-2 cells
treated with Lis1 protein (Fig.
4A). The migration ability of the PC12 and HCN-2 cells was
enhanced only upon the addition of Lis1 protein; this enhanced
migration was attenuated by transfection with miR-139-5p.
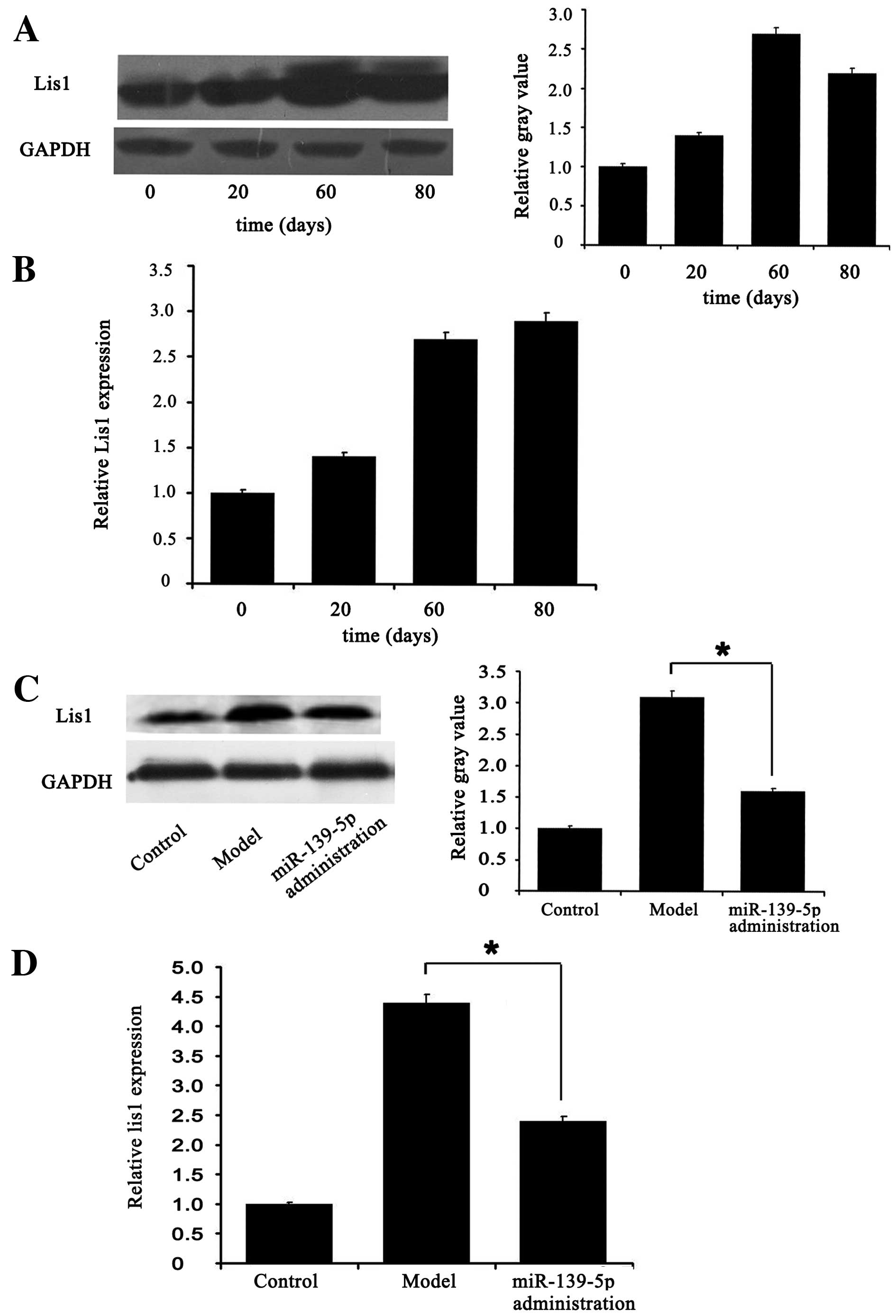
miR-139-5p modulates rat brain
development by targeting Lis1
qRT-PCR (Fig. 5B)
and western blot analysis (Fig.
5A) indicated that the mRNA and protein levels of Lis1
were higher at days 20–80 after the rat model of dysplasia was
established compared with day 0 (healthy control group). Sixty days
after the rat models were established, miR-139-5p was administered,
and this led to the marked downregulation of Lis1 mRNA
(Fig. 5D) and protein levels
(Fig. 5C). Additionally, it was
found that miR-139-5p altered cell morphology in the rat brain, as
indicated by H&E and Nissl staining (Fig. 6).
Discussion
Neuronal migration has been studied extensively for
over 30 years in diverse mammalian species from mice to humans
(27). Abnormal neuronal
migration often leads to cortical dysplasia (28–30). Lis1 is known to regulate
cell proliferation and migration during brain development (31,32) and its expression is known to be
disrupted in patients with ILS and MDS, suggesting that a mutation
of Lis1 leads to cortical dysplasia (30). The regulatory function of miRNAs
in various developmental, migration and apoptotic pathways of
diverse organisms is known. Therefore, the investigation of miRNAs
that may regulate Lis1 expression could potentially lead to
the development of novel therapeutic methods for the treatment of
patients with cortical dysplasia.
In this study, we profiled the expression of 391
miRNAs in the brains of immature Sprague-Dawley (20–80 days) rats
by miRNA microarray to identify the miRNAs responsible for cortical
neuronal migration. Among the miRNAs found with significant changes
in expression, we selected miR-139-5p to clarify its function in
the brain. The screening standard required that the miRNA appear in
all databases and have a 2 to 10-fold upregulation in expression,
although multiple genes are predicted by several algorithms
(TargetScan, miRanda and PicTar) as potential miR-139-5p targets.
The microarray results revealed that the expression of miR-139-5p
was decreased in the brains of Sprague-Dawley rats (on days 20–80),
and that Lis1 is a target of miR-139-5p.
The regulatory function of miRNA in various
developmental, differentiation, proliferation, migration and
apoptotic pathways of diverse organisms is known (33–35). As expected, we found that
miR-139-5p plays a role in rat brain development. Furthermore,
miR-139-5p expression was decreased in Sprague-Dawley rats (on days
20–80) with focal cortical dysplasia induced by liquid nitrogen
lesions. Due to the significant change in miR-139-5p expression
during rat brain development, our data suggest that Lis1 is
a target for miR-139-5p. As Lis1 is associated with neuronal
migration, we speculate that miR-139-5p regulates rat cortical
neuronal migration by modulating the expression of certain target
genes. These targets, as predicted by multiple algorithms (36), include Lis1, Capn8,
Gmfb, Mapk1, Dclk1, Vim, Mgst1,
Gnb1, Klf15 and Cdkn1b (37).
Studies have previously demonstrated the upregulated
expression of Lis1 by proteomic analysis of a freeze-lesion
model of focal cortical dysplasia (38,39). Furthermore, Lis1 is a key
gene in brain development due to its role in cell proliferation and
migration (40,41). Notably, in our study, Lis1
expression was markedly upregulated at day 20 and its expression
was maintained at these high levels until day 80 (Fig. 5A and B). This is likely an
important reflection of the critical role of Lis1 in
promoting cell proliferation and migration. Cell migration assays
revealed that miR-139-5p significantly inhibited the migration of
PC12 and HCN-2 cells treated with or without Lis1 protein. At the
same time, miR-139-5p was administered to the rats with focal
cortical dysplasia; miR-139-5p administration markedly decreased
the expression of Lis1. In addition, the injured cortex of
these rat models showed a certain degree of recovery. Furthermore,
we confirmed that miR-139-5p targets Lis1 and inhibits its
expression, as confirmed by luciferase assay.
It can be concluded that the upregulation of Lis1 in
the rat brain is at least partially caused by a reduction in
miR-139-5p. As the upregulation of Lis1 can promote the occurrence
of certain developmental events in the rat brain, miR-139-5p
modulates rat brain development thorugh the regulation of Lis1
expression. miR-139-5p may also contribute to rat brain development
by affecting the expression of several other putative targets
(30,42,43). Futher studies are required to
validate other predicted targets involved in brain development. The
data presented in our study suggest an important role of miR-139-5p
in regulating cell migration, thus offering a novel target for the
development of therapeutic agents against focal cortical
dysplasia.
Acknowledgements
This study was supported by the Nanjing Medical
Science and technique Development Foundation (ZKX07019) and by the
Nanjing City Science and Technology Project (201106018).
Abbreviations:
|
UTR
|
untranslated region
|
|
qRT-PCR
|
quantitative reverse
transcription-polymerase chain reaction
|
|
FBS
|
fetal bovine serum
|
|
SPF
|
specific pathogen-free
|
|
PBS
|
phosphate-buffered saline
|
References
|
1
|
Barkovich AJ, Kuzniecky RI, Jackson GD,
Guerrini R and Dobyns WB: A developmental and genetic
classification for malformations of cortical development.
Neurology. 65:1873–1887. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Andiman E, Haynes L, Trachtenberg L, et
al: The cerebral cortex overlying periventricular leukomalacia:
analysis of pyramidal neurons. Brain Pathol. 20:803–814. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Barkovich J, Millen J and Dobyns B: A
developmental and genetic classification for midbrain-hindbrain
malformations. Brain. 132:3199–3230. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lo Nigro C, Chong CS, Smith AC, Dobyns WB,
Carrozzo R and Ledbetter DH: Point mutations and an intragenic
deletion in LIS1, the lissencephaly causative gene in isolated
lissencephaly sequence and Miller-Dieker syndrome. Hum Mol Genet.
6:157–164. 1997.PubMed/NCBI
|
|
5
|
Tsai JW, Bremner KH and Vallee RB: Dual
subcellular roles for LIS1 and dynein in radial neuronal migration
in live brain tissue. Nat Neurosci. 10:970–979. 2007. View Article : Google Scholar
|
|
6
|
Yamada M, Toba S, Yoshida Y, et al: LIS1
and NDEL1 coordinate the plus-end-directed transport of cytoplasmic
dynein. EMBO J. 27:2471–2483. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bi W, Sapir T, Shchelochkov OA, et al:
Increased LIS1 expression affects human and mouse brain
development. Nat Genet. 41:168–177. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Haverfield V, Whited J, Petras S, Dobyns B
and Das S: Intragenic deletions and duplications of the LIS1 and
DCX genes: a major disease-causing mechanism in lissencephaly and
subcortical band heterotopia. Eur J Hum Genet. 17:911–918. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wynshaw-Boris A: Lissencephaly and LIS1:
insights into the molecular mechanisms of neuronal migration and
development. Clin Genet. 72:296–304. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Carthew W: Gene regulation by microRNAs.
Curr Opin Genet Dev. 16:203–208. 2006. View Article : Google Scholar
|
|
11
|
Bushati N and Cohen M: microRNA functions.
Annu Rev Cell Dev Biol. 23:175–205. 2007. View Article : Google Scholar
|
|
12
|
Sayed D and Abdellatif M: MicroRNAs in
development and disease. Physiol Rev. 91:827–887. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Qureshi IA and Mehler MF: Emerging roles
of non-coding RNAs in brain evolution, development, plasticity and
disease. Nat Rev Neurosci. 13:528–541. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Castoldi M, Schmidt S, Benes V, et al: A
sensitive array for microRNA expression profiling (miChip) based on
locked nucleic acids (LNA). RNA. 12:913–929. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Meng F, Henson R, Wehbe-Janek H, et al:
MicroRNA-21 regulates expression of the PTEN tumor suppressor gene
in human hepatocellular cancer. Gastroenterology. 133:647–658.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang J and Ruan K: miR-200c affects the
mRNA expression of E-cadherin by regulating the mRNA level of TCF8
during post-natal epididymal development in juvenile rats. Acta
Biochim Biophys Sin (Shanghai). 42:628–634. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Nakanishi N, Nakagawa Y, Tokushige N, et
al: The up-regulation of microRNA-335 is associated with lipid
metabolism in liver and white adipose tissue of genetically obese
mice. Biochem Biophys Res Commun. 385:492–496. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wu AJ, Hua H, Munson SH and McDevitt HO:
Tumor necrosis factor-alpha regulation of
CD4+CD25+T cell levels in NOD mice. Proc Natl
Acad Sci USA. 99:12287–12292. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shenouda SK and Alahari SK: MicroRNA
function in cancer: oncogene or a tumor suppressor? Cancer
Metastasis Rev. 28:369–378. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Norman L and Sarnow P: Modulation of
hepatitis C virus RNA abundance and the isoprenoid biosynthesis
pathway by microRNA miR-122 involves distinct mechanisms. J Virol.
84:666–670. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hui AB, Lenarduzzi M, Krushel T, et al:
Comprehensive MicroRNA profiling for head and neck squamous cell
carcinomas. Clin Cancer Res. 16:1129–1139. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wong M, Yam P, Ching P, et al: Rho
GTPase-activating protein deleted in liver cancer suppresses cell
proliferation and invasion in hepatocellular carcinoma. Cancer Res.
65:8861–8868. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gitlin L, Karelsky S and Andino R: Short
interfering RNA confers intracellular antiviral immunity in human
cells. Nature. 418:430–434. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Takase I, Shigeto H, Suzuki SO, Kikuchi H,
Ohyagi Y and Kira J: Prenatal freeze lesioning produces
epileptogenic focal cortical dysplasia. Epilepsia. 49:997–1010.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Calin GA, Sevignani C, Dumitru CD, et al:
Human microRNA genes are frequently located at fragile sites and
genomic regions involved in cancers. Proc Natl Acad Sci USA.
101:2999–3004. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Yingling J1, Youn YH, Darling D, et al:
Neuroepithelial stem cell proliferation requires LIS1 for precise
spindle orientation and symmetric division. Cell. 132:474–486.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wegiel J, Kuchna I, Nowicki K, et al: The
neuropathology of autism: defects of neurogenesis and neuronal
migration, and dysplastic changes. Acta Neuropathol. 119:755–770.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Manent JB, Wang Y, Chang Y, Paramasivam M
and LoTurco JJ: Dcx reexpression reduces subcortical band
heterotopia and seizure threshold in an animal model of neuronal
migration disorder. Nat Med. 15:84–90. 2009. View Article : Google Scholar
|
|
29
|
Manzini MC and Walsh CA: What disorders of
cortical development tell us about the cortex: one plus one does
not always make two. Curr Opin Genet Dev. 21:333–339. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Guerrini R and Parrini E: Neuronal
migration disorders. Neurobiol Dis. 38:154–166. 2010. View Article : Google Scholar
|
|
31
|
Walser M, Hansén A, Svensson A, et al:
Peripheral administration of bovine GH regulates the expression of
cerebrocortical beta-globin, GABAB receptor 1, and the
Lissencephaly-1 protein (LIS-1) in adult hypophysectomized rats.
Growth Horm IGF Res. 21:16–24. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Cappello S, Monzo P and Vallee RB: NudC is
required for interkinetic nuclear migration and neuronal migration
during neocortical development. Dev Biol. 357:326–335. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Schepeler T, Reinert JT, Ostenfeld MS, et
al: Diagnostic and prognostic microRNAs in stage II colon cancer.
Cancer Res. 68:6416–6424. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Zhang C: Novel functions for small RNA
molecules. Curr Opin Mol Ther. 11:641–651. 2009.PubMed/NCBI
|
|
35
|
Winter J, Jung S, Keller S, Gregory RI and
Diederichs S: Many roads to maturity: microRNA biogenesis pathways
and their regulation. Nat Cell Biol. 11:228–234. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Goswami S, Angkasekwinai P, Shan M, et al:
Divergent functions for airway epithelial matrix metalloproteinase
7 and retinoic acid in experimental asthma. Nat Immunol.
10:496–503. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Davalos V and Esteller M: MicroRNAs and
cancer epigenetics: a macrorevolution. Curr Opin Oncol. 22:35–45.
2010. View Article : Google Scholar
|
|
38
|
Verrotti A, Spalice A, Ursitti F, et al:
New trends in neuronal migration disorders. Eur J Paediatr Neurol.
14:1–12. 2010. View Article : Google Scholar
|
|
39
|
Huang YJ, Zheng G, Lu XP, Lu HY and Mo XM:
Proteomics of epilepsy induced by focal disorder of cortical
development. Chin J Neuromed. 9:670–673. 2010.
|
|
40
|
Sand M, Sand D, Altmeyer P and Bechara FG:
MicroRNA in non-melanoma skin cancer. Cancer Biomark. 11:253–257.
2012.PubMed/NCBI
|
|
41
|
Hippenmeyer S, Youn YH, Moon HM, et al:
Genetic mosaic dissection of Lis1 and Ndel1 in neuronal migration.
Neuron. 68:695–709. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Saugstad JA: MicroRNAs as effectors of
brain function with roles in ischemia and injury, neuroprotection,
and neurodegeneration. J Cereb Blood Flow Metab. 30:1564–1576.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Shen K, Liang Q, Xu K, et al: MiR-139
inhibits invasion and metastasis of colorectal cancer by targeting
the type I insulin-like growth factor receptor. Biochem Pharmacol.
84:320–330. 2012. View Article : Google Scholar : PubMed/NCBI
|