Introduction
Gastric cancer is responsible for almost one million
deaths worldwide per year. Although the age-adjusted mortality of
gastric cancer has decreased over the last few decades, gastric
cancer remains the second leading cause of cancer-related
mortatlity (1). Gastric cancer is
an asymptomatic disease during the early stages, and therefore, is
often detected at a late stage. The 5-year survival rate of gastric
cancer is only 20–30% (2). For
these reasons, intensive research is required, in order to devise
new approaches to its diagnosis and treatment. Apoptosis is a key
factor that contributes to the antitumor activity of
chemotherapeutic drugs and therefore, plays a key role in the
search for new approaches for the treatment of cancer. In addition,
as cell numbers can be regulated by factors that influence cell
survival, proliferation and differentiation, apoptosis is a
gene-directed program that has had profound implications for our
understanding of developmental biology and tissue homeostasis
(3). Most anticancer agents now
in use were developed using empirical screens designed to identify
agents that selectively kill tumor cells (4). It is now well-established that
anticancer agents induce apoptosis, and that the disruption of
apoptotic programs can reduce treatment sensitivity (4). Anticancer agents induce apoptosis in
normal tissues, as well as in tumors. In fact, many of the
pathologists who identified apoptosis in tumors realized that
apoptotic cell death was induced in a subset of normal tissues
(e.g., bone marrow and the intestines), and it was suggested that
the process may contribute to the ‘toxicity’ associated with
chemotherapy (3). Although there
is much more to learn, our current understanding of apoptosis
provides new avenues for cancer diagnostics, prognosis and therapy.
It seems likely that rational strategies to manipulate cell suicide
programs will produce novel therapies that are less toxic and
mutagenic than current treatment regimens (3).
Quercetin (3,3′,4′,5,7-pentahydroxyflavone) is a
bioflavonoid, and is one of the most widely distributed dietary
polyphenolics; for example, it is found in vegetables, fruits, tea
and wine (5). Research into the
anticarcinogenic potential of flavonoids with animal and cellular
model systems supports a protective role against the progression of
colorectal cancer (6–9). Apart from other flavonoids,
quercetin has been proposed to have anti-atherogenic,
anti-inflammatory, antihypertensive, and anticancer effects
(3). Quercetin has been reported
to protect against important diseases, including lung and colon
cancers (10–12). In addition, quercetin has been
shown to inhibit human breast cancer cell proliferation and to
induce apoptosis through the regulation of Bcl-2 and Bax (13).
However, the effects of quercetin on human gastric
adenocarcinoma cells have not yet been fully elucidated. Therefore,
in this study, we investigated the effects of quercetin in human
gastric adenocarcinoma cells and the mechanisms responsible for its
activities.
Materials and methods
Materials
Quercetin was purchased from Alexis Biochemicals
(Vinci Biochem, Florence, Italy) and PD98059 (ERK inhibitor),
SB203580 (p38 kinase inhibitor), and SP600125 (JNK inhibitor) were
purchased from Tocris Bioscience (Bristol, UK). All other agents
were purchased from Sigma-Aldrich (St. Louis, MO, USA).
Cells
The AGS cell line (a human gastric adenocarcinoma
cell line) was established at the Cancer Research Center, College
of Medicine, Seoul National University, Seoul, Korea, and
propagated in RPMI-1640 medium (Gibco-BRL, Carlsbad, CA, USA)
supplemented with 10% heat-inactivated fetal bovine serum
containing 20 μg/ml penicillin and streptomycin in a 5%
CO2 atmosphere at 37°C.
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay
Cell viability was assessed by MTT assay. The AGS or
human embryo kidney (HEK)293 cells were seeded into each well of
12-well culture plates and then cultured in RPMI-1640 supplemented
with 10% heat-inactivated fetal bovine serum containing 20 μg/ml
penicillin and streptomycin for 72 h. MTT solution [100 μl; 5 mg/ml
in phosphate-buffered saline (PBS)] was then added to each well,
and the plates were incubated at 37°C for 4 h. After removing the
supernatant and shaking with 200 μl of dimethyl sulfoxide (Jersey
Lab Supply, Livingston, NJ, USA) for 30 min, and absorbances were
measured at 570 nm. All experiments were repeated at least 3
times.
Flow cytometric analysis
In order to investigate the effects of treatment on
the cell cycle of the AGS cells, flow cytometric analysis was
performed with propidium iodine (PI) staining, as previously
described (14,15). The cells (1×106) were
placed in an Eppendorf tube, and 700 μl of ice-cold fixation buffer
(ethyl alcohol) was slowly added with vortexing. The tubes were
sealed with parafilm, incubated at 4°C overnight, spun for 3 min at
106 × g at 4°C, and the supernatants were aspirated and discarded.
Cell pellets were resuspended in 200 μl of PI staining solution [2
μl of PI (5 mg/ml) and 2 μl of RNase in a total of 196 μl of PBS]
and centrifuged at 2,0817 × g for 5 sec. After being kept for 30
min in the dark at room temperature, the samples were analyzed
using a fluorescence-activated cell sorter (FACScan;
Becton-Dickinson, Mountain View, CA, USA) at λ=488 nm using
Cell-Quest software (Becton-Dickinson). The DNA content
distribution of normally growing AGS cells is characterized by 2
peaks, representing the G1/G0 and G2/M phases. The G1/G0 phase is
composed of cells in the resting state of the cell cycle and has
the greatest diploid DNA content, whereas the DNA content in the
G2/M phase is more than diploid. Cells in the sub-G1 phase have the
least DNA content, which is termed hypodiploid (a hypoploid DNA
content represents DNA fragmentation) (16).
Assessment of mitochondrial membrane
depolarization
Mitochondrial membrane depolarization was evaluated
using the JC-1 fluorescence probe according to the manufacturer’s
instructions (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA).
The AGS cells were labeled with 2 μM JC-1 for 30 min at 37°C and
then analyzed by flow cytometry at excitation and emission
wavelengths of 530/30 or 585/42 nm and 488 nm, respectively. The
cells without red fluorescence were regarded to manifest
mitochondrial membrane depolarization.
Caspase assay
Caspase-3 kits (Cellular Activity Assay kit PLUS)
were purchased from Biomol International (Plymouth Meeting, PA,
USA). After the treatments, the cells were centrifuged (10,000 × g,
4°C, 10 min), washed with PBS, resuspended in ice-cold cell lysis
buffer, incubated on ice for 10 min, centrifuged at 10,000 × g
(4°C, 10 min), and the supernatants were removed. The supernatants
(10 μl) were then incubated with 50 μl of substrate (400-μM
Ac-DEVD-pNA) in 40 μl of assay buffer at 37°C. Absorbance was read
at 405 nm at several time points. The pNA concentration in the
samples was determined using a standard pNA concentration vs.
absorbance plot. The pan-caspase inhibitor, zVAD-fmk (Calbiochem,
La Jolla, CA, USA), was used to validate the assay method.
Patch-clamp experiments
Whole-cell configuration patch-clamp experiments
were performed at room temperature (22–25°C). The AGS cells were
transferred to a small chamber on an inverted microscope stage
(IX70; Olympus, Tokyo, Japan), and constantly perfused with a
solution containing (mmol/l) KCl 2.8, NaCl 145, CaCl2 2,
glucose 10, MgCl2 1.2 and HEPES 10 (adjusted to pH 7.4
with NaOH). The pipette solution contained (mmol/l) Cs-glutamate
145, NaCl 8, Cs-2-bis(2-aminophenoxy)-ethane-N,N,N′,N′-tetraacetic
acid 10 and HEPES-CsOH 10 (adjusted to pH 7.2 with CsOH). An
Axopatch I-D (Axon Instruments, Foster City, CA, USA) was used to
amplify the membrane currents and potentials. pClamp software
version 9.2 and Digidata 1322A (Axon Instruments) were used for
data acquisition and to apply command pulses. The results were
analyzed using pClamp and Origin software (Microcal Origin version
6.0).
Transient receptor potential melastatin
(TRPM)7 expression in HEK293 cells
The HEK293 cells were transfected with Flag-murine
LTRPC7/pCDNA4-TO construct and grown on glass coverslips in
Dulbecco’s modified Eagle’s medium supplemented with 10% fetal
bovine serum, blasticidin (5 μg/ml) and zeocin (0.4 mg/ml). TRPM7
(LTRPC7) expression was induced by the addition of 1 μg/ml
tetracycline to the culture medium. Whole-cell patch-clamp
experiments were performed at 21–25°C using cells grown on glass
coverslips.
Sodium dodecyl sulfate-polyacrylamide gel
electrophoresis and western blot analysis
Western blot analysis was performed using AGS cell
lysates. Proteins were separated by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis by using 8%
polyacrylamide gels, transferred onto polyvinylidene difluoride
membranes, and quantified by incubating the membranes with the
indicated antibodies. Antibodies against poly(ADP-ribose)
polymerase (PARP), p-p38, p-JNK, p-ERK, p38, ERK and β-actin were
purchased from Cell Signaling Technology (Danvers, MA, USA).
Antibodies against JNK and GAPDH were obtained from Santa Cruz
Biotechnology. The data are representative of at least 3
independent experiments.
Statistical analysis
Data are expressed as the means ± SEM. The Student’s
t-test was used to determine the significances of the differences.
Statistical significance was set at P<0.05.
Results
Quercetin induces AGS cell death
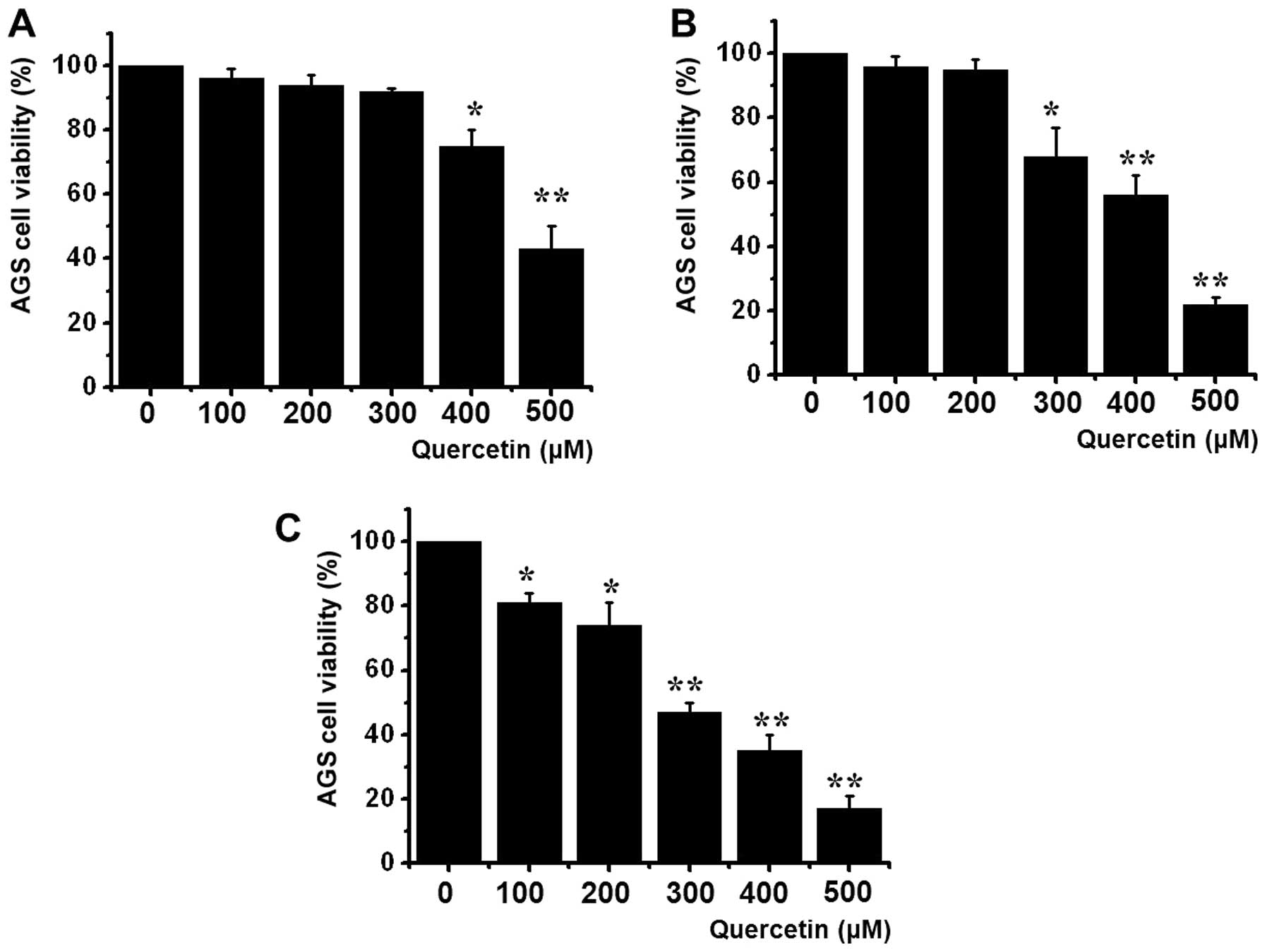
To ascertain whether quercetin kills AGS cells, we
performed MTT assays. Viable cell populations were gradually
reduced by quercetin in a concentratio- and a time-dependent manner
at 24, 48 and 72 h with an IC50 value of 267 μM
(Fig. 1).
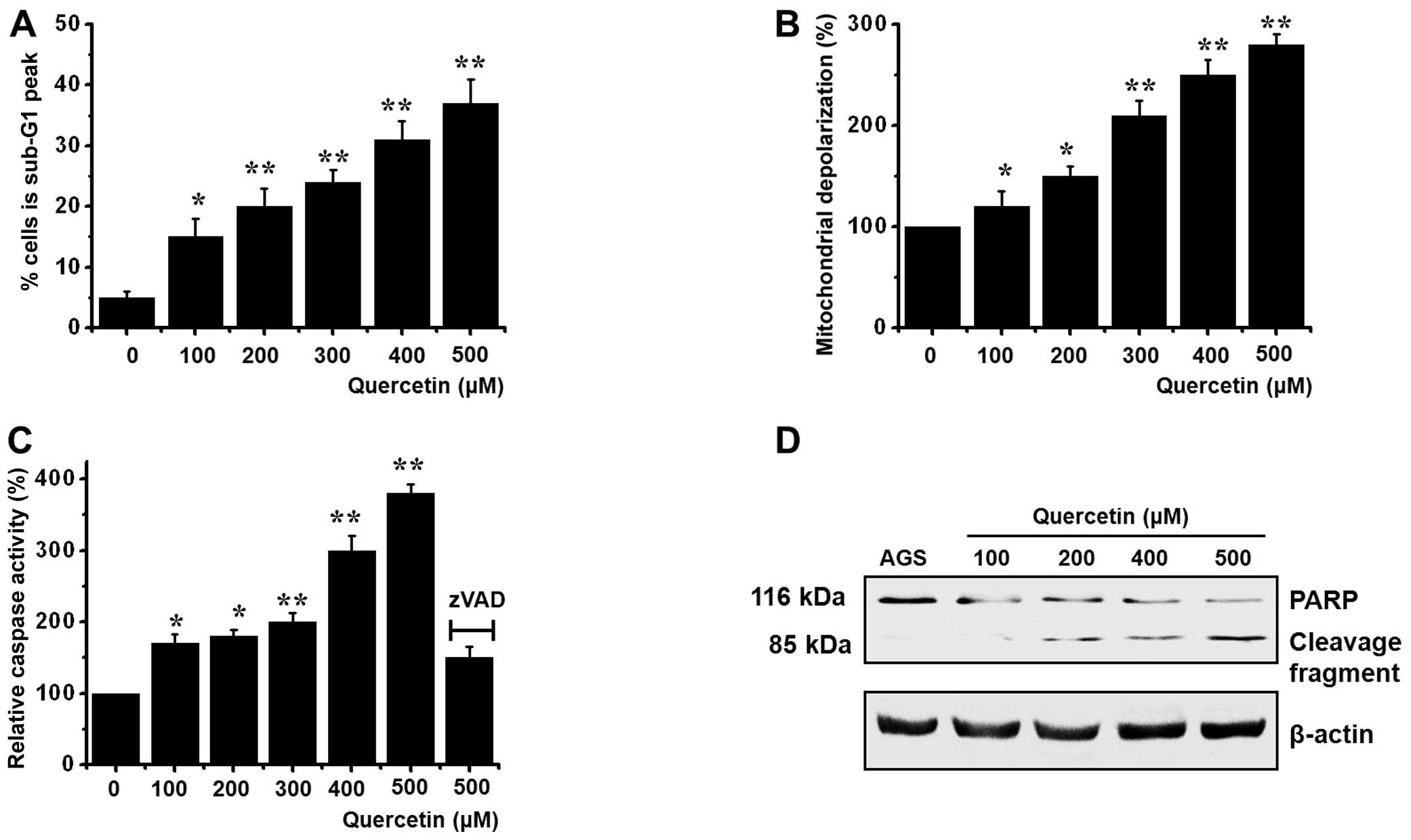
Quercetin induces apoptosis of AGS
cells
To determine whether the AGS-induced cell death was
due to apoptosis, we conducted sub-G1 analysis, as previously
described (17,18). Briefly, the cells were incubated
with quercetin and stained with a fluorescent DNA stain (PI).
Endogenous endonucleases in apoptotic cells cleave DNA into
endonucleosomal fragments. The loss of DNA is detected by FACS
analysis, as a reduction of nuclear staining in apoptotic cells,
which results in a novel (sub-G1) fluorescence peak to the left of
the regular fluorescence peak. Flow cytometric analysis revealed
that the percentage of cells in the sub-G1 phase markedly increased
in a dose-dependent manner in the cells treated with quercetin
(Fig. 2A). In addition, quercetin
elevated mitochondrial membrane depolarization, an early event of
intrinsic apoptotic signaling (Fig.
2B). Thus, our findings suggest that quercetin induces
apoptosis through intrinsic apoptotic mechanism(s). In addition,
using a synthetic substrate, we examined caspase-3 activity
(caspase-3 activation is a hallmark of apoptotic cell death) in the
AGS cells. Quercetin increased the activity of caspase-3, which was
suppressed by zVAD-fmk (a pan-caspase inhibitor) (Fig. 2C). We also examined changes in
caspase-3 activity by western blot analysis for its natural
substrate, PARP, which has been shown to function as a cellular
target of caspase-3 and other caspases. During apoptosis, PARP is
proteolytically cleaved from its 116-kDa-intact form into 85 and 25
kDa fragments. Following incubation with quercetin for 72 h, the
AGS cells showed increased PARP cleavage (Fig. 2D).
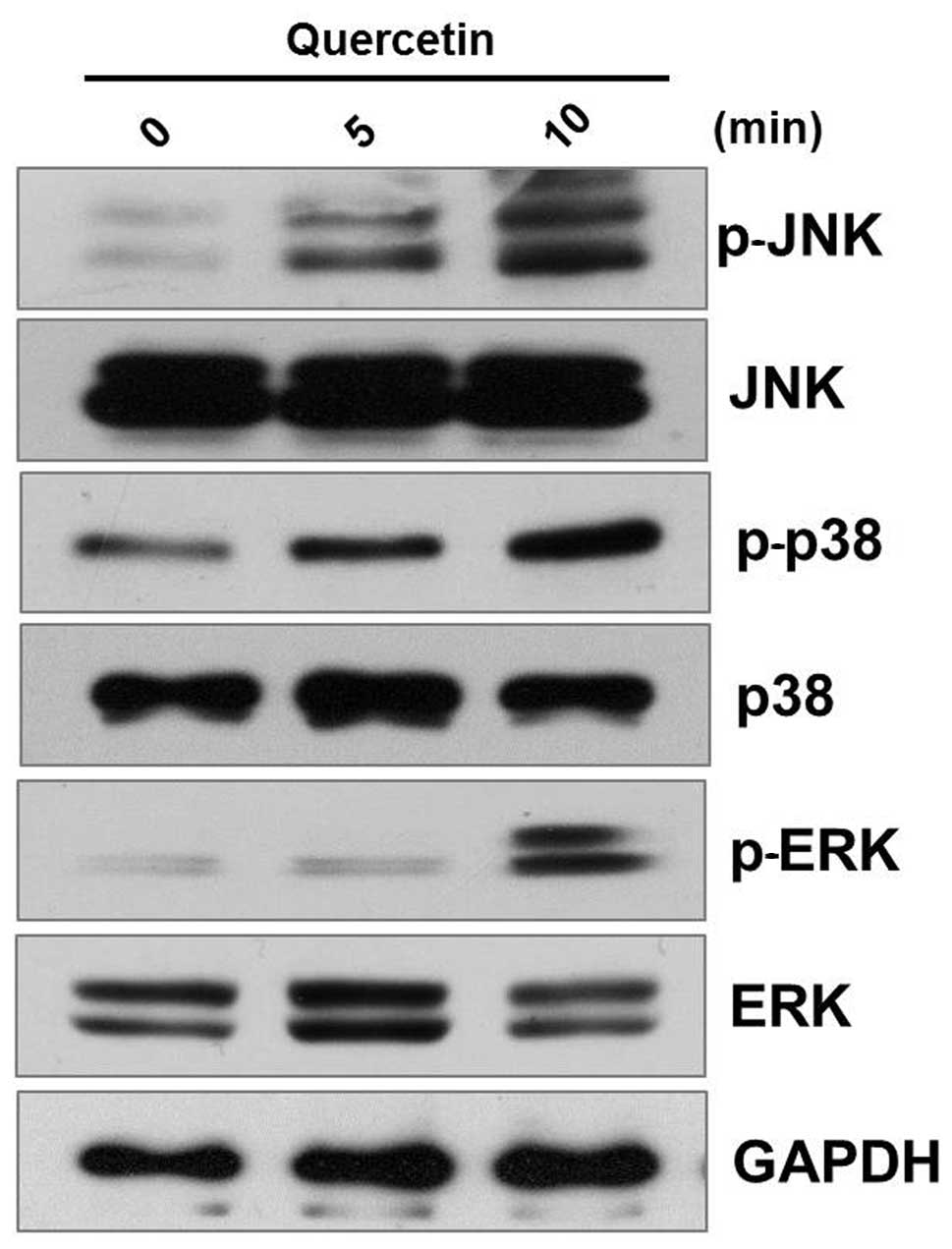
Involvement of mitogen-activated protein
kinases (MAPKs) in quercetin-induced apoptosis of AGS cells
To investigate the signaling pathway of
quercetin-induced apoptosis in AGS cells, we assessed the effects
of quercetin on MAPKs, as they play critical roles in the
apoptosis-related signaling pathway. As shown in Fig. 3, quercetin markedly increased the
phosphorylation levels of p38, JNK and ERK (Fig. 3). As the role of MAPK signaling in
the quercetin induced apoptosis of AGS cells has been little
characterized in this study (Fig.
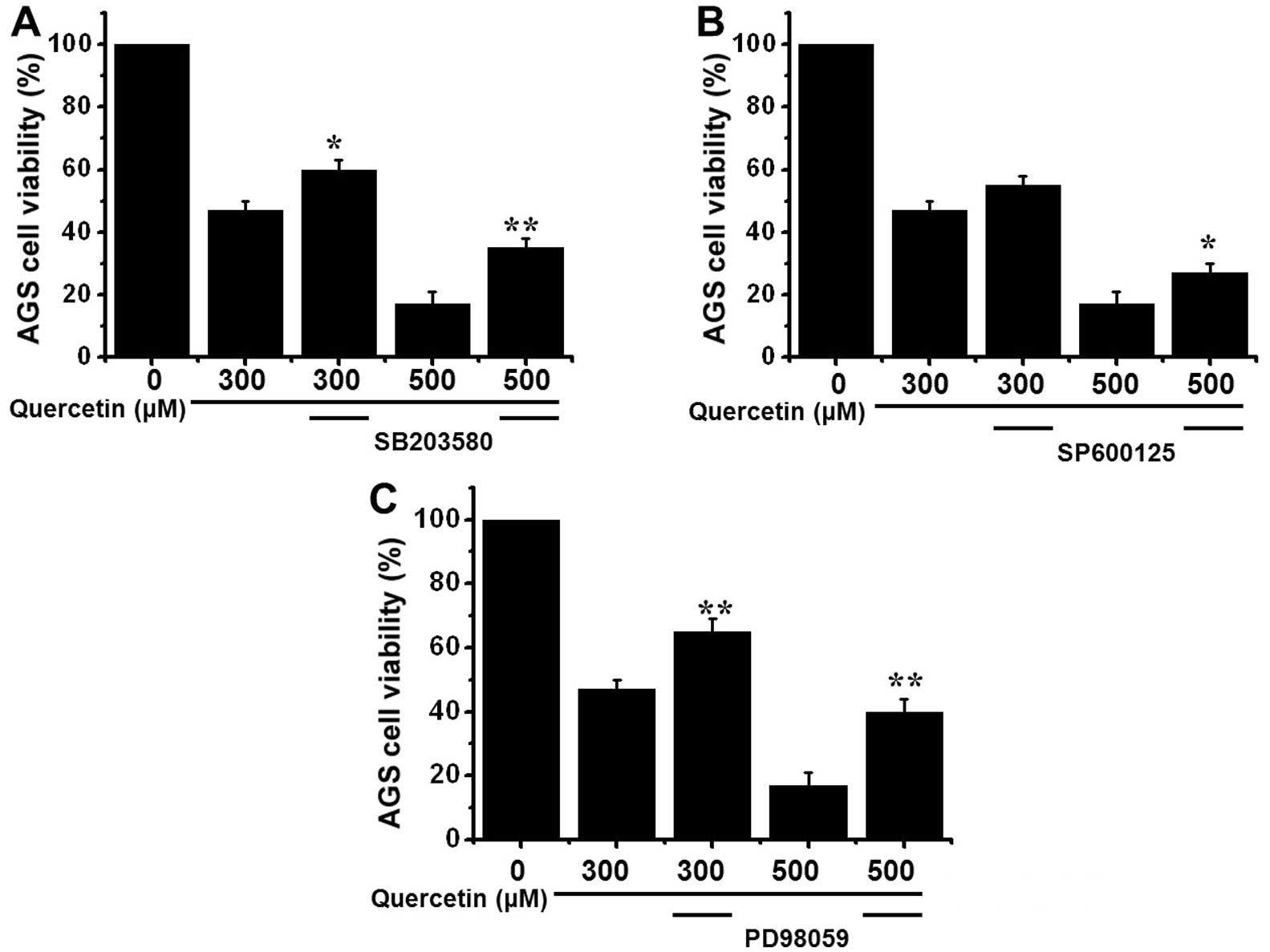
3), we performed MTT assays using MAPK inhibitors to assess the
effects of MAPK signaling on quercetin signaling. MTT assays
revealed that exposure to quercetin with SB203580 (Fig. 4A), SP600125 (Fig. 4B) and PD98059 (Fig. 4C) resulted in an increase in the
viable cell populations.
Effects of quercetin in TRPM7 currents on
AGS cells and HEK293 cells overexpressing TRPM7
In a previous study, we found that AGS cells express
the TRPM7 channel, which is known to be essential for cell survival
and has been suggested as a pharmacological target for the
treatment of gastric cancer (19). Therefore, in this study, we
investigated whether quercetin influences TRPM7 currents in AGS
cells. To confirm the effects of quercetin on TRPM7 currents, we
investigated the effects of quercetin in AGS cells using
patch-clamp techniques. First, to examine the effects of internal
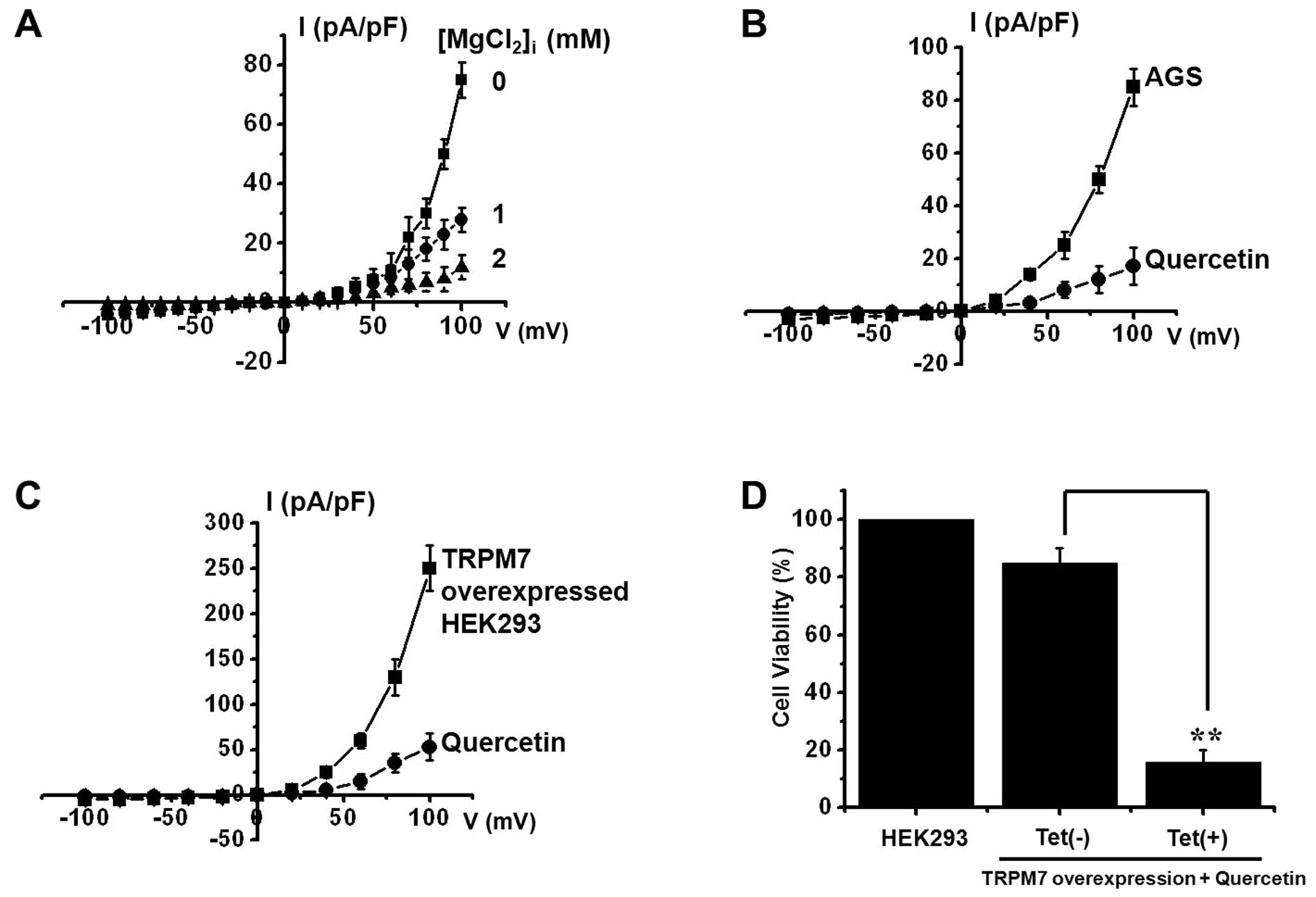
Mg2+ on TRPM7-like currents, Mg2+ was added
to the pipette solution at different concentrations from 0 to 2 mM.
Current amplitudes were indeed larger at the lower concentration,
and the presence of 2 mM Mg2+ in the pipette solution
significantly inhibited the current (n=4; Fig. 5A). Whole cell voltage-clamp
recordings were then taken to investigate the effects of quercetin
on TRPM7-like currents in AGS cells. A voltage ramp from +100 to
−100 mV evoked small inward currents at negative potentials,
whereas larger outward currents were evoked at positive potentials,
showing outwardly rectifying cation currents (n=6; Fig. 5B). However, in the presence of 500
μM quercetin, the amplitudes of these currents were inhibited
outwardly by 79.3±1.3% and inwardly by 89.2±1.4% (n=6; Fig. 5B). Furthermore, similar results
were obtained withthe HEK293 cells overexpressing TRPM7 channels
(Fig. 5C). To provide additional
evidence supporting the positive contribution made by TRPM7
channels to quercetin toxicity, we investigated the effects of
different TRPM7 channel levels on quercetin-mediated cell death.
For this purpose, we used HEK293 cells with inducible TRPM7 channel
expression (20,21). In the absence of induced TRPM7
channel expression [TRPM7(−) cells, Tet(−)], quercetin did not
induce HEK293 cell death, as shown by MTT assay (n=6; Fig. 5D). However, when TRPM7 channel
overexpression was induced by the addition of tetracycline
[TRPM7(+) cells, Tet(+)], quercetin markedly induced more HEK293
cell death, indicating that TRPM7 channel levels are positively
associated with quercetin-induced cell death.
Discussion
Quercetin is a polyphenolic flavonoid that is found
in fruits and vegetables, such as, apples, blueberries, broccoli,
grapes, leeks, lettuce, onions and tomatoes. The mechanisms
underlying the cancer-preventive effects of quercetin and other
flavonoids are believed to be associated with antioxidant effects,
the inhibition of enzymes that activate carcinogens, the
modification of signal transduction pathways, and interactions with
receptors and/or proteins (22,23). However, the effects of quercetin
on AGS human adenocarcinoma cells have not yet been fully
elucidated. Quercetin has been shown to exert antiproliferative
effects on the parental EPG85-257P cell line and its
daunorubicin-resistant variant, EPG85-257RDB, mainly through the
induction of apoptosis (24).
Quercetin has also been shown to induce the apoptosis of BGC-823
cells and decrease the Bcl-2/Bax ratio with the increased
expression of caspase-3 (25). As
previously demonstrated, quercetin induced pronounced pro-apoptotic
effects by activating the mitochondrial pathway in AGS and MKN28
gastric cancer cells. Treatment with quercetin induced the
appearance of autophagic vacuoles, the formation of acidic
vesicular organelles, the conversion of LC3-I to LC3-II, the
recruitment of LC3-II to the autophagosomes, as well as the
activation of autophagy genes, suggesting that quercetin initiates
autophagic progression in gastric cancer cells (26). However, to our knowledge, there
are no studies available on the association of MAPK and TRP
channels in quercetin-induced apoptosis in gastric cancer
cells.
Certain studies have suggested that MAPK activation
plays a role in the apoptosis of cancer cells (27,28). PKI 166 induced apoptosis through
the activation of the p53, MAPK and caspase pathway in epidermoid
carcinoma (27). In addition,
isoorientin induced mitochondrial-mediated apoptosis through MAPK
signaling pathways in human hepatoblastoma cancer cells (28). Thus, in this study, we
investigated whether MAPKs play a role in quercetin-induced
apoptosis in AGS cells. The 3 principal ubiquitously expressed
MAPKs are p38 MAPK, JNK and ERK and JNK (29), and in the present study,
inhibitors of p38, JNK and ERK inhibited AGS cell apoptosis induced
by quercetin.
TRP channels were first cloned from the
Drosophila species, and constitute a superfamily of proteins
that encode a diverse group of Ca2+-permeable
non-selective cation channels (NSCCs) (30). Based on their amino acid
sequences, the TRP superfamily can be divided into 7 subfamilies:
the TRPC (‘Canonical’), TRPM (‘Melastatin’), TRPV (‘Vanilloid’),
TRPP (‘Polycystin’), TRPML (‘Mucolipin’), TRPA (‘Ankyrin’) and the
TRPN (‘NOMPC’) families (31).
TRPM7, a member of the TRPM ion channel subfamily, is a widely
expressed bifunctional protein with both ion channel and α-kinase
domains (20,21). In a previous study [Kim et
al (19)], we reported that
human gastric adenocarcinoma cells express TRPM7 channels and that
this channel is essential for cell survival. It was also suggested
that this channel be viewed as a potential target for the
pharmacological treatment of gastric cancer.
The TRPM7 channel is widely expressed in various
tissues, including those of the brain, spleen, kidneys, lungs,
liver and heart (20,32). Furthermore, its expression has
also been reported in head and neck carcinoma (21), retinoblastoma (33), breast cancer (34,35) and gastric cancer (19). The activation of TRPM7 channels in
human head and neck squamous carcinoma cell lines enhances
proliferation (21), whereas
TRPM7 knockdown by siRNA and the blockade of TRPM7 channels by
Gd3+ or 2-APB, which exerts broad inhibitory effects on
TRP superfamilies, including TRPM7, suppresses proliferation. The
proliferation of human retinoblastoma cells is promoted by
increasing spontaneous Ca2+ influx (33). Furthermore, the blockade of cation
channel by Gd3+, La3+, LOE908, or 2-APB
inhibits spontaneous Ca2+ influx and decreases the
proliferation of retinoblastoma cells. TRPM7 channel-related
proliferation is also associated with MCF-7 human breast cancer
cells (35) and AGS human gastric
adenocarcinoma cells (19). As
previously demonstrated, the proliferation of both cells was
prevented by TRPM7 protein knockout by siRNA transfection and by
blocking TRPM7 channels with La3+ or 2-APB. Thus, TRPM7
channels may play an important role in valuable diagnostic and/or
prognostic markers, as well as targets for pharmaceutical
intervention and more in depth investigations are required to
elucidate the role TRPM7 channels in cancer (21).
In the present study, we found that quercetin
inhibited TRPM7-like currents in AGS cells (Fig. 5B). To investigate the the effects
of quercetin on TRPM7 channels, we overexpressed TRPM7 channels in
HEK293 cells. It was found that quercetin inhibited the TRPM7
currents (Fig. 5C), and that its
inhibitory effects were similar to those in the AGS cells.
Therefore, we hypothesized that TRPM7 is involved in the apoptotic
activity of quercetin in AGS cells.
TRPM7 has also been suggested to play a central role
in: cellular Mg2+ homeostasis (36), central nervous system ischemic
injury (37), skeletogenesis in
zebrafish (38), defecation
rhythm in Caenorhabditis elegans (39), cholinergic vesicle fusion with the
plasma membrane (40),
phosphoinositide-3-kinase signaling in lymphocytes (41), and in osteoblast proliferation
(42).
Apoptotic pathways are induced by a variety of
signals, and a certain mechanism consistently implicated in
apoptosis is the activation of a series of cytosolic proteases,
that is, the caspases (43).
Caspases are synthesized as inactive pro-enzymes that are processed
in cells undergoing apoptosis by self-proteolysis and/or cleavage
by another protein. Functionally, active caspases form a
proteolytic cascade, which is capable of cleaving and activating
specific substrates, including PARP (an enzyme involved in DNA
repair and genomic maintenance). Cleavage events of this type may
result in important alterations to normal homeostatic cellular
processes (44). Furthermore,
PARP has been shown to be a cellular target of caspase-3 and of
other caspases. In our study, After 72 h of incubation with
quercetin, PARP became proteolytically cleaved in AGS cells from
its 116-kDa intact form into 85-kDa fragments (Fig. 2D), and caspase-3 activity
increased accordingly (Fig.
2C).
In addition, we found that quercetin inhibited cell
viability and that MAPK signaling pathways were involved in AGS
cell apoptosis (Figs. 3 and
4). Furthermore, TRPM7 currents
in AGS cells and in HEK293 cells which overexpress TRPM7 were
inhibited by quercetin (Fig. 5B and
C). Furthermore, in TRPM7-overexpressing HEK293 cells,
quercetin induced cell death at a greater rate, suggesting that the
upregulation of TRPM7 channels increases quercetin-induced cell
death. These results suggest that quercetin plays an important
pathophysiological role in AGS cells through MAPK signaling and
TRPM7 channels. Accordingly, we suggest that quercetin be viewed as
a potential drug for the pharmacological treatment of gastric
cancer.
Acknowledgements
This study was supported by the Basic Science
Research Program of the Korean National Research Foundation (NRF)
funded by the Ministry of Education, Science and Technology (Grant
no. 2010-0021347).
References
|
1
|
Ferlay J, Shin HR, Bray F, Forman D,
Mathers C and Parkin DM: Estimates of worldwide burden of cancer in
2008: GLOBOCAN 2008. Int J Cancer. 127:2893–2917. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bosman FT, Carneiro F, Hruban RH and
Theise ND: WHO Classification of Tumours of the Digestive System.
3. 4th edition. IARC Press; Geneva: 2010
|
|
3
|
Lowe SW and Lin AW: Apoptosis in cancer.
Carcinogenesis. 21:485–495. 2000. View Article : Google Scholar
|
|
4
|
Schmitt CA and Lowe SW: Apoptosis and
therapy. J Pathol. 187:127–137. 1999. View Article : Google Scholar
|
|
5
|
Ishizawa K, Yoshizumi M, Kawai Y, Terao J,
Kihira Y, Ikeda Y, Tomita S, Minakuchi K, Tsuchiya K and Tamaki T:
Pharmacology in health food: metabolism of quercetin in vivo and
its protective effect against arteriosclerosis. J Pharmacol Sci.
115:466–470. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Williams RJ, Spencer JPE and Rice-Evans C:
Flavonoids: antioxidants or signaling molecules? Free Rad Biol Med.
36:838–849. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Daskiewicz JB, Depeint F, Viornery L,
Bayet C, Comte-Sarrazin G, Comte G, Gee JM, Johnson IT, Ndjoko K,
Hostettmann K and Barron D: Effects of flavonoids on cell
proliferation and caspase activation in a human colonic cell line
HT29: an SAR study. J Med Chem. 48:2790–2804. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Surh YJ, Kundu JK, Na HK and Lee JS:
Redox-sensitive transcription factors as prime targets for
chemoprevention with anti-inflammatory and antioxidative
phytochemicals. J Nutr. 135:S2993–S3001. 2005.PubMed/NCBI
|
|
9
|
Kyle JA, Sharp L, Little J, Duthie GG and
McNeill G: Dietary flavonoid intake and colorectal cancer: a
case-control study. Br J Nutr. 103:429–436. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Stefani ED, Boffetta P, Deneo-Pellegrini
H, Mendilaharsu M, Carzoglio JC, Ronco A and Olivera L: Dietary
antioxidants and lung cancer risk: a case-control study in Uruguay.
Nutr Cancer. 34:100–110. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
van Erk MJ, Roepman P, van der Lende TR,
Stierum RH, Aarts JM, van Bladeren PJ and van Ommen B: Integrated
assessment by multiple gene expression analysis of quercetin
bioactivity on anticancer-related mechanisms in colon cancer cells
in vitro. Eur J Nutr. 44:143–156. 2005.PubMed/NCBI
|
|
12
|
Dihal AA, de Boer VC, van der Woude H,
Tilburgs C, Bruijntjes JP, Alink GM, Rietjens IM, Woutersen RA and
Stierum RH: Quercetin, but not its glycosidated conjugate rutin,
inhibits azoxymethane-induced colorectal carcinogenesis in F344
rats. J Nutr. 136:2862–2867. 2006.PubMed/NCBI
|
|
13
|
Duo J, Ying GG, Wang GW and Zhang L:
Quercetin inhibits human breast cancer cell proliferation and
induces apoptosis via Bcl-2 and Bax regulation. Mol Med Rep.
5:1453–1456. 2012.PubMed/NCBI
|
|
14
|
Schönherr R: Clinical relevance of ion
channels for diagnosis and therapy of cancer. J Membr Biol.
205:175–184. 2005.PubMed/NCBI
|
|
15
|
Lehen’kyi V, Shapovalov G, Skryma R and
Prevarskaya N: Ion channels and transporters in cancer. 5 Ion
channels in control of cancer and cell apoptosis. Am J Physiol Cell
Physiol. 301:C1281–C1289. 2011.PubMed/NCBI
|
|
16
|
Wang BJ, Won SJ, Yu ZR and Su CL: Free
radical scavenging and apoptotic effects of Cordyceps
sinensis fractionated by supercritical carbon dioxide. Food
Chem Toxicol. 43:543–552. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Hotz MA, Gong J, Traganos F and
Darzynkiewicz Z: Flow cytometric detection of apoptosis: comparison
of the assays of in situ DNA degradation and chromatin changes.
Cytometry. 15:237–244. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Vermes I, Haanen C and Reutelingsperger C:
Flow cytometry of apoptotic cell death. J Immunol Methods.
243:167–190. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim BJ, Park EJ, Lee JH, Jeon JH, Kim SJ
and So I: Suppression of transient receptor potential melastatin 7
channel induces cell death in gastric cancer. Cancer Sci.
99:2502–2509. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Nadler MJ, Hermosura MC, Inabe K, Perraud
AL, Zhu Q, Stokes AJ, Kurosaki T, Kinet JP, Penner R, Scharenberg
AM and Fleig A: LTRPC7 is a Mg.ATP-regulated divalent cation
channel required for cell viability. Nature. 411:590–595. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jiang J, Li MH, Inoue K, Chu XP, Seeds J
and Xiong ZG: Transient receptor potential melastatin 7-like
current in human head and neck carcinoma cells: role in cell
proliferation. Cancer Res. 67:10929–10938. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Daker M, Ahmad M and Khoo AS:
Quercetin-induced inhibition and synergistic activity with
cisplatin - a chemotherapeutic strategy for nasopharyngeal
carcinoma cells. Cancer Cell Int. 12:342012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Murakami A, Ashida H and Terao J:
Multitargeted cancer prevention by quercetin. Cancer Lett.
269:315–325. 2008. View Article : Google Scholar
|
|
24
|
Borska S, Chmielewska M, Wysocka T,
Drag-Zalesinska M, Zabel M and Dziegiel P: In vitro effect of
quercetin on human gastric carcinoma: targeting cancer cells death
and MDR. Food Chem Toxicol. 50:3375–3383. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Wang P, Zhang K, Zhang Q, Mei J, Chen CJ,
Feng ZZ and Yu DH: Effects of quercetin on the apoptosis of the
human gastric carcinoma cells. Toxicol In Vitro. 26:221–228. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang K, Liu R, Li J, Mao J, Lei Y, Wu J,
Zeng J, Zhang T, Wu H, Chen L, Huang C and Wei Y: Quercetin induces
protective autophagy in gastric cancer cells: involvement of
Akt-mTOR- and hypoxia-induced factor 1α-mediated signaling.
Autophagy. 7:966–978. 2011.PubMed/NCBI
|
|
27
|
Das S, Dey KK, Bharti R, MaitiChoudhury S,
Maiti S and Mandal M: PKI 166 induced redox signalling and
apoptosis through activation of p53, MAP kinase and caspase pathway
in epidermoid carcinoma. J Exp Ther Oncol. 10:139–153. 2012.
|
|
28
|
Yuan L, Wang J, Xiao H, Wu W, Wang Y and
Liu X: MAPK signaling pathways regulate mitochondrial-mediated
apoptosis induced by isoorientin in human hepatoblastoma cancer
cells. Food Chem Toxicol. 53:62–68. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Pearson G, Robinson F, Beers Gibson T, Xu
BE, Karandikar M, Berman K and Cobb MH: Mitogen-activated protein
(MAP) kinase pathways: regulation and physiological functions.
Endocr Rev. 22:153–183. 2001.PubMed/NCBI
|
|
30
|
Clapham DE: TRP channels as cellular
sensors. Nature. 426:517–524. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Pedersen SF, Owsianik G and Nilius B: TRP
channels: an overview. Cell Calcium. 38:233–252. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Runnels LW, Yue L and Clapham DE:
TRP-PLIK, a bifunctional protein with kinase and ion channel
activities. Science. 291:1043–1047. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Hanano T, Hara Y, Shi J, Morita H,
Umebayashi C, Mori E, Sumimoto H, Ito Y, Mori Y and Inoue R:
Involvement of TRPM7 in cell growth as a spontaneously activated
Ca2+ entry pathway in human retinoblastoma cells. J
Pharmacol Sci. 95:403–419. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Dhennin-Duthille I, Gautier M, Faouzi M,
Guilbert A, Brevet M, Vaudry D, Ahidouch A, Sevestre H and
Ouadid-Ahidouch H: High expression of transient receptor potential
channels in human breast cancer epithelial cells and tissues:
correlation with pathological parameters. Cell Physiol Biochem.
28:813–822. 2011. View Article : Google Scholar
|
|
35
|
Guilbert A, Gautier M, Dhennin-Duthille I,
Haren N, Sevestre H and Ouadid-Ahidouch H: Evidence that TRPM7 is
required for breast cancer cell proliferation. Am J Physiol Cell
Physiol. 297:C493–C502. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Schmitz C, Perraud AL, Johnson CO, Inabe
K, Smith MK, Penner R, Kurosaki T, Fleig A and Scharenberg AM:
Regulation of vertebrate cellular Mg2+ homeostasis by
TRPM7. Cell. 114:191–200. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Aarts M, Iihara K, Wei WL, Xiong ZG,
Arundine M, Cerwinski W, MacDonald JF and Tymianski M: A key role
for TRPM7 channels in anoxic neuronal death. Cell. 115:863–877.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Elizondo MR, Arduini BL, Paulsen J,
MacDonald EL, Sabel JL, Henion PD, Cornell RA and Parichy DM:
Defective skeletogenesis with kidney stone formation in dwarf
zebrafish mutant for trpm7. Curr Biol. 15:667–671. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Vriens J, Owsianik G, Voets T, Droogmans G
and Nilius B: Invertebrate TRP proteins as functional models for
mammalian channels. Pflugers Arch. 449:213–226. 2004.PubMed/NCBI
|
|
40
|
Brauchi S, Krapivinsky G, Krapivinsky L
and Clapham DE: TRPM7 facilitates cholinergic vesicle fusion with
the plasma membrane. Proc Natl Acad Sci USA. 105:8304–8308. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Sahni J and Scharenberg AM: TRPM7 ion
channels are required for sustained phosphoinositide 3-kinase
signaling in lymphocytes. Cell Metab. 8:84–93. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Abed E and Moreau R: Importance of
melastatin-like transient receptor potential 7 and cations
(magnesium, calcium) in human osteoblast-like cell proliferation.
Cell Prolif. 40:849–865. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Patel T, Gores GJ and Kaufmann SH: The
role of proteases during apoptosis. FASEB J. 10:587–597. 1996.
|
|
44
|
Lin C, Holland RE Jr, Donofrio JC, McCoy
MH, Tudor LR and Chambers TM: Caspase activation in equine
influenza virus induced apoptotic cell death. Vet Microbiol.
84:357–365. 2002. View Article : Google Scholar : PubMed/NCBI
|