Introduction
Increasingly, ischemic heart diseases such as
myocardial infarction (MI) are important causes of morbidity and
mortality worldwide (1). Due to
the loss of cardiomyocytes and formation of scar tissue after MI,
cardiac function deteriorates and a series of fatal complications
inevitably occur. Current treatment strategies based on stem cell
transplantation for ischemic heart diseases are emerging,
particularly those utilizing adult stem cells from heart tissues,
termed cardiac stem cells (CSCs) (2–6).
Phase I clinical trials such as SCIPIO (NCT00474461), ALCADIA
(NCT00981006) and CADUCEUS (NCT00893360) involving heart-derived
cells have been conducted (4).
The feasibility, safety and effectiveness of injection of
autologous heart-derived cells were assessed in these clinical
trials with encouraging preliminary results, as evidenced by the
reduction in myocardial scar mass or improvement in the left
ventricular ejection fraction following cell treatment (7,8).
CSCs comprise a cluster of small cells located in a
specific niche of the heart. Some subtypes of CSCs have been
reported based on cell surface markers and cardiac location
(9–12). In 2003, Beltrami et al
(9) first reported that
Lin−/c-Kit+ CSCs were detected in the adult
rat heart, as located in the atria, apex and base-midregion of the
ventricle. C-Kit+ CSCs are multipotent stem cells that
can differentiate into myocardiocytes, smooth muscle cells and
vascular epithelia cells under certain conditions. Findings of
recent studies showed that c-Kit+ CSC transplantation
improved the performance of heart tissue injured through coronary
artery ligation (13,14). The results of the SCIPIO clinical
trial also showed that transplantation of c-Kit+ CSCs
enhanced the ejection fraction in vivo (7). Ellison et al reported that
c-Kit+ CSCs are necessary and sufficient for functional
cardiac regeneration and repair following myocardial damage
(15). These reports highlight
the viability and effectiveness of c-Kit+ CSC
transplantation in myocardial regeneration.
Myocardium in peri-infarcted zones is in a state of
stress post-MI, thus, several cardioprotective molecules including,
but not limited to, PI3K, hypoxia-induced factor 1 (HIF1), NOTCH1
and stromal cell-derived factor (SDF), are upregulated (16–20). Previous results indicated that
stem cell factor (SCF), a powerful stem cell chemokine, is
upregulated in the cardiomyocytes of peri-infarcted zones (21), thus activating the chemokine
signaling of the SCF/c-Kit axis. In this manner, c-Kit+
CSCs are migrated towards injured areas to fulfill critical roles
in the process of myocardial regeneration. Endogenous
c-Kit+ CSCs are located mainly in the niche of the
atria, while most MI lesions clinically occur within the left
ventricular because of left anterior descending (LAD) coronary
artery disorders. Consequently, there is a large barrier that the
chemoactivated c-Kit+ CSCs in atria must navigate when
migrating towards injured zones within the left ventricular
post-MI. Further knowledge regarding the mechanisms involved in the
migration of activated c-Kit+ CSCs post-MI would
therefore strengthen the evidence for CSCs transplantation in the
treatment of MI.
PI3K/AKT signaling is known to be an important
signal transduction cascade involved in cancer cell survival,
apoptosis and motility (3). This
type of signaling is crucial in stem cell biology. Activation of
the PI3K/AKT pathway is crucial for VEGF-mediated c-Kit+
CSC migration in vitro and in vivo (22), and enhances cellular engraftment
post-MI (23–25). However, the role of the PI3K/AKT
pathway in SCF/c-Kit signaling-mediated CSC migration remains
elusive. In the present investigation, we aimed to explore the
crosstalk of SCF/c-Kit and PI3K signaling in the migration of
c-Kit+ CSCs. Our results indicated that SCF-mediated
c-Kit+ CSCs migration occurs at least partly via the
activation of PI3K/AKT/matrix metalloproteinase (MMP)-2/-9
signaling.
Materials and methods
Isolation and culture of CSCs from adult
rat hearts
CSCs were isolated by magnet-activated cell sorting
(MACS) from the hearts of male Sprague-Dawley rats as described
previously (13,21). Briefly, the heart was excised and
the aorta was rapidly cannulated, followed by perfusion with
Ca2+-free Tyrode solution for 10 min and then digestion
with 0.5 mg/ml collagenase (Sigma, St. Louis, MO, USA) and 0.05
mg/ml trypsin (Difco, Kansas, MO, USA) at 37°C for 30 min. The
heart tissue was sectioned and the resulting cell suspension was
filtered with a strainer (Becton-Dickson, Franklin Lakes, NJ, USA).
Cells were then incubated with a rabbit anti-c-Kit antibody (1:50;
Santa Cruz Biotechnology, Inc., Texas, USA) and separated using
immunomagnetic microbeads (Miltenyi Biotech, Bergish Gladbach,
Germany). CSCs were then cultured in Dulbecco’s modified Eagle’s
medium/Ham’s Nutrient Mixture F12 (1:1) (DMEM/F12) (Sigma-Aldrich)
containing 15% fetal bovine serum (FBS) (Gibco, Carlsbad, CA, USA),
10 ng/ml basic fibroblast growth factor (bFGF), 20 ng/ml epidermal
growth factor (EGF) (both from Sigma-Aldrich) and 2.5 μ/ml
erythropoietin (EPO) (BioLegend, San Diego, CA, USA) at 37°C. After
28 days of culture, confluent CSCs were passaged.
RNA isolation and quantitative RT-PCR
(RT-qPCR)
Total RNA was extracted with TRIzol reagent
(Invitrogen Life Technologies, Carlsbad, CA, USA). The total RNA (1
μg) was used as a template to generate cDNA by oligo(dT18) using
the Fermentas RT System (cat. no. K1622; Thermo Fisher Scientific,
Inc., Guangzhou, China). Primer pairs (5′-3′) used for PCR were
synthesized by Sangon Biotech Co., Ltd. (Shanghai, China) and are
shown in Table I. PCR products
were separated by 1.5% agarose gel electrophoresis and visualized
under UV using a gel documentation system (Bio-Rad, Hercules, CA,
USA). RT-qPCR was conducted using the LightCycler480 instrument
[Roche (China) Ltd., Shanghai, China] in a final volume of 20 μl,
which included 10 μl SYBR-Green I PCR Master mix (Toyobo Co., Ltd.,
Osaka, Japan), 0.4 μl forward primer (10 μM), 0.4 μl reverse primer
(10 μM), 2 μl cDNA and 7.2 μl dH2O. PCR amplification
was performed using the following protocol: 95°C for 1 min, then 40
cycles of 95°C for 15 sec, and finally 60°C for 1 min. The relative
abundance of target gene mRNAs was determined from the CT values
and plotted as the fold change compared with the control groups.
For the PCR analysis, the transcription levels of GAPDH served as a
loading control.
 | Table IPrimers used for PCR. |
Table I
Primers used for PCR.
| Genes (ID) |
Semi-quantitative
PCR sequences (5′→3′) |
|---|
| C-Kit
(D12524.1) | F:
ttggcaaagaagacaacgac
R: gcacagacaccactgggata |
| GATA-4
(NM_144730.1) | F:
gcagaaacaacaaagggaaat
R: gggagaaacagcgtaaatga |
| Nkx2.5
(NM_053651.1) | F:
ttccagaaccgccgctacaag
R: ccgacgccaaagttcacgaag |
| Troponin I
(NM_017184.1) | F:
gggagactggaggaagaacg
R: gagggaacaacaacagcaaaa |
| vwF
(NM_053889.1) | F:
cctacggcttgcacgattca
R: ccacttcctcttccgacttac |
| Tagln
(NM_031549) | F:
gaggactgtaatggctttgg
R: gccttccctttctaactgat |
| GAPDH
(BC059110) | F:
cagtgccagcctcgtctcat
R: ggggccatccacagtcttc |
|
| Genes (ID) |
Quantitative
PCR sequences (5′→3′) |
|
| MMP-2
(NM_031054) | F:
ggaagcatcaaatcggactg
R: caccctcttaaatctgaaatcacc |
| MMP-9
(NM_031055) | F:
cccacttactttggaaacg
R: gaagatgaatggaaatacgc |
| GAPDH
(BC059110) | F:
cccatctatgagggttacgc
R: tttaatgtcacgcacgatttc |
Western blot analysis
Total proteins were extracted using a NP-40 protein
extraction kit (cat. no. P0013F), and quantified using an Enhanced
BCA Protein Assay kit (cat. no. P0010) (both from Beyotime
Institute of Biotechnology, Haimen, China). Protein was transferred
onto PVDF membranes by electrophoretic transfer following
electrophoretic separation by SDS-PAGE. The membranes were probed
with primary antibodies against MMP-2, MMP-9, AKT, p-AKT (1:1,000;
Cell Signaling Technology, Beverly, MA, USA), and GAPDH (1:1,000;
Santa Cruz Biotechnology) in TBST plus 5% skimmed milk overnight at
4°C. After three washes with TBST, the membranes were incubated
with horseradish peroxidase-conjugated secondary antibodies for 1 h
at room temperature. Bands were visualized using enhanced
chemiluminescence reagents (Thermo Fisher Scientific, Inc.,
Rockford, IL, USA) and analyzed with a Bio-Rad VersaDoc™ 5000 MP
system (Life Science Research, Hercules, CA, USA). GAPDH and AKT
were used as loading controls.
Gelatin zymography of MMP enzyme
activity
MMP-2 and MMP-9 activity was measured by SDS-PAGE
under non-reducing conditions using gels containing 1% gelatin
(Mini-PROTEAN II system; Bio-Rad), and electrophoresis was carried
out at 4°C. After washing with 2% Triton X-100 to remove the SDS,
the gels were incubated in 37°C with buffer containing 50 mM Tris
(pH 7.5), 5 mmol/CaCl2 and 1 mmol/l ZnCl2 for
18 h. MMP activity was visualized by staining with Coomassie Blue
R-250 (Bio-Rad).
Transwell migration assay
Chemotaxis experiments were performed using a
24-well Transwell chemotaxis chamber technique (Millipore,
Billerica, MA, USA) as previously described (21). Briefly, DMEM (600 μl) alone or
medium containing 5, 10, 20, 30 and 50 ng/ml recombinant rat SCF
(PeproTech, Rocky Hill, NJ, USA) was placed in the lower chamber. A
total of 1×105 CSCs in 200 μl of medium were seeded into
the upper chamber (pore size, 8 μm). For the inhibition experiment,
CSCs were preincubated with a c-Kit blocking antibody or a PI3K/AKT
inhibitor, LY294002 (100 nM), for 30 min prior to seeding. The
chamber was then incubated for 12 h at 37°C in a humidified
atmosphere with 5% CO2. The membrane was removed and its
upper surface was wiped away with a cotton swab to remove the
unmigrated CSCs. The membrane (Millipore) was then fixed in neutral
formalin for 10 min at room temperature and then stained with 0.1%
crystal violet for 5 min. The number of CSCs that had migrated to
the lower surface of the membrane was counted in 10 random
high-power fields (HPFs) under a light microscope (Nikon Eclipse
80i; Nikon Instruments, Inc., Melville, NY, USA). A chemotactic
index (CI) was calculated to express stimulated migration: CI =
stimulated migration (CSCs number per HPF)/random migration (CSCs
number per HPF). Each assay was performed in triplicate.
Statistical analysis
Statistical analysis was performed using PRISM
Software® (GraphPad, La Jolla, CA, USA). The data are
presented as means ± SD. For analysis of differences between two
groups, Student’s t-tests were performed. For multiple groups,
ANOVA was carried out followed by the Student-Newman-Keuls method.
P<0.05 was considered statistically significant.
Results
Characteristics and identification of
c-Kit+ cells from adult rat hearts
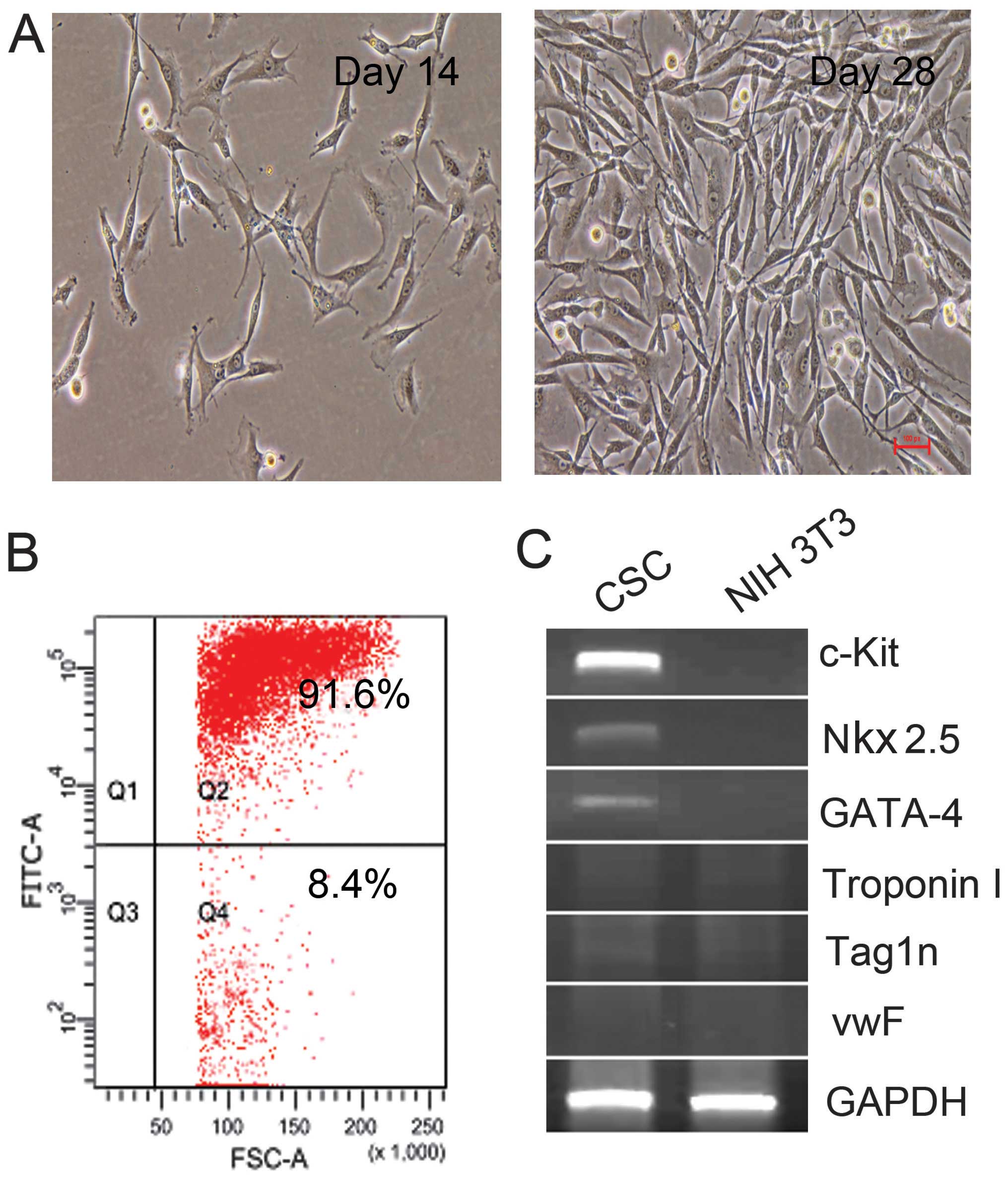
Using immunomagnetic microbeads, c-Kit+
cells were isolated and collected from adult rat hearts. Under
light microscopy, the freshly isolated c-Kit+ CSCs
appeared as small, round and phase-bright cells, and were suspended
in the medium. Cells turned into polygonal and spindle-like shape 7
days later, and cells reached confluence at 4 weeks. The purity of
c-Kit+ CSCs was 91.6% as determined by flow cytometry
(Fig. 1B). The expression of
c-Kit was detected by RT-qPCR. As shown in Fig. 1C, c-Kit mRNA was detectable in
isolated CSCs, while as a negative control, NIH 3T3 cells did not
express c-Kit mRNA. Furthermore, Nkx2.5 and GATA-4 mRNA were
detectable in the isolated cells, while markers for myocardium
(Troponin I), epithelial cells (vwF) and smooth muscle cells
(Tag1n) were negative. These results confirmed that the isolated
cells were c-Kit+ CSCs.
Effects of SCF on CSC migration in
vitro
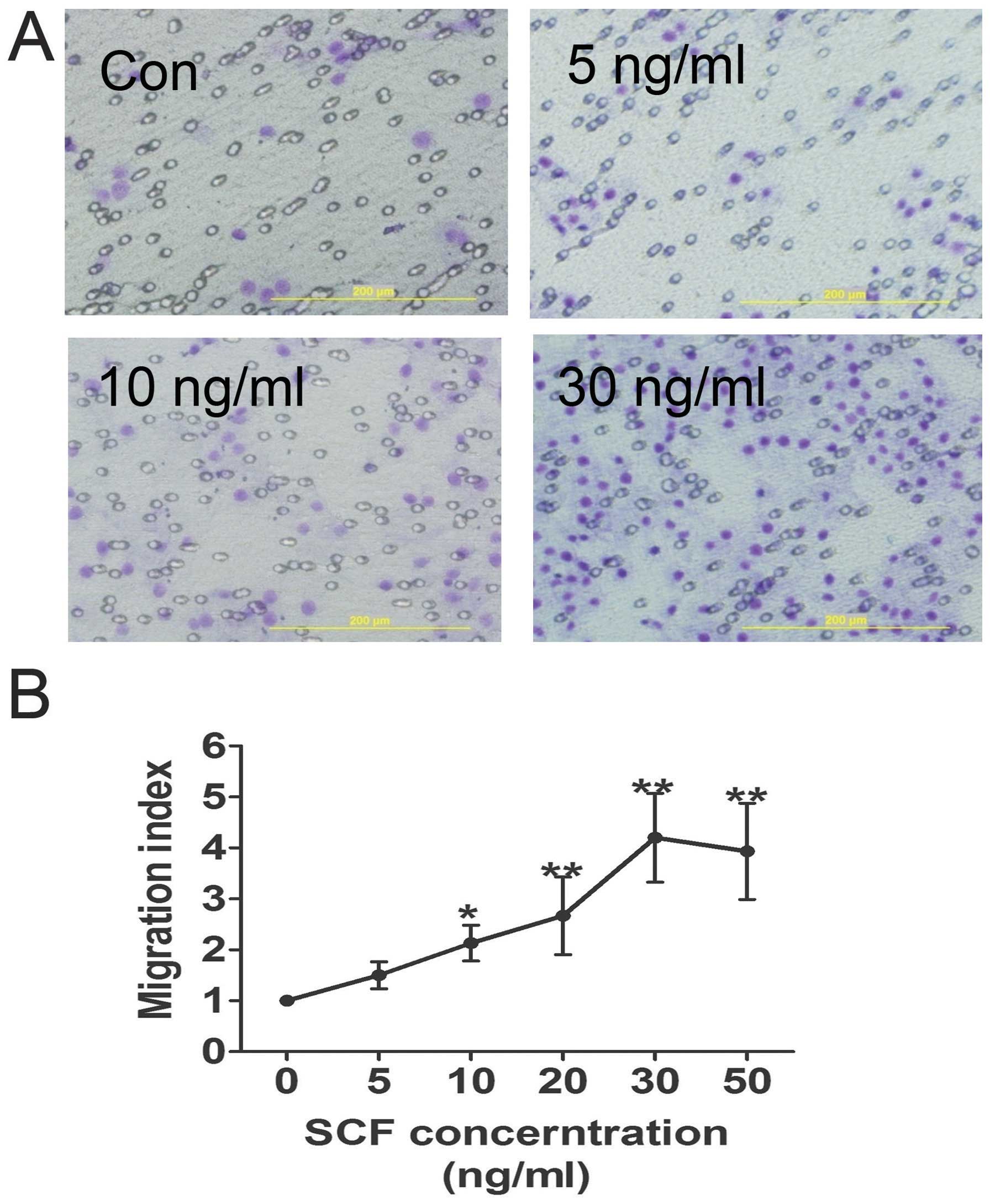
A Transwell-based migration assay was established to
quantitatively evaluate CSC migration in vitro. As shown in
Fig. 2, compared with the control
group the average number of migrated CSCs increased significantly
in the conditioned medium groups with increasing SCF concentration,
which reached a peak at 30 ng/ml. Furthermore, SCF-induced CSC
migration was inhibited by pre-treatment of the CSCs with SCF
antibody, as well as LY294002 (Fig.
4C).
SCF-mediated MMP-2/-9 expression and
enzymatic activity
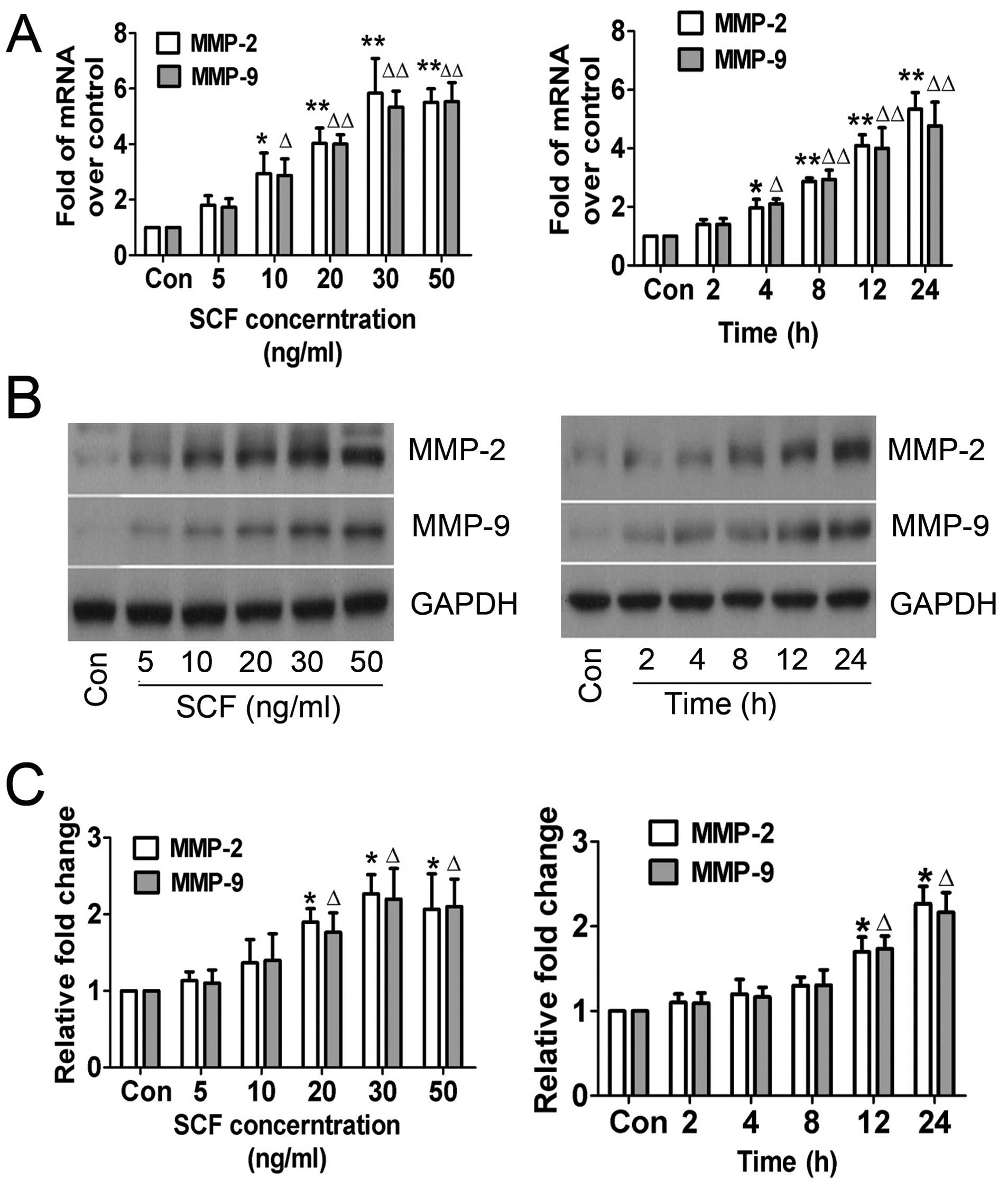
The role of SCF in MMP-2 and MMP-9 expression in
CSCs in vitro was examined. Gelatin zymography assay was
conducted based on the theory that MMP-2 and MMP-9 degrades
gelatin. The results of the RT-qPCR and western blot analysis
revealed that MMP-2 and MMP-9 expression in CSCs was not regulated
when treated with higher concentrations of SCF for longer periods
of time, which was confirmed by the enzymatic activity results
(Fig. 3). The trend of increasing
MMP-2 and MMP-9 indicated that SCF mediated their expression and
enzymatic activity in a concentration- and time-dependent
manner.
PI3K/AKT is involved in SCF-mediated
MMP-2/-9 expression
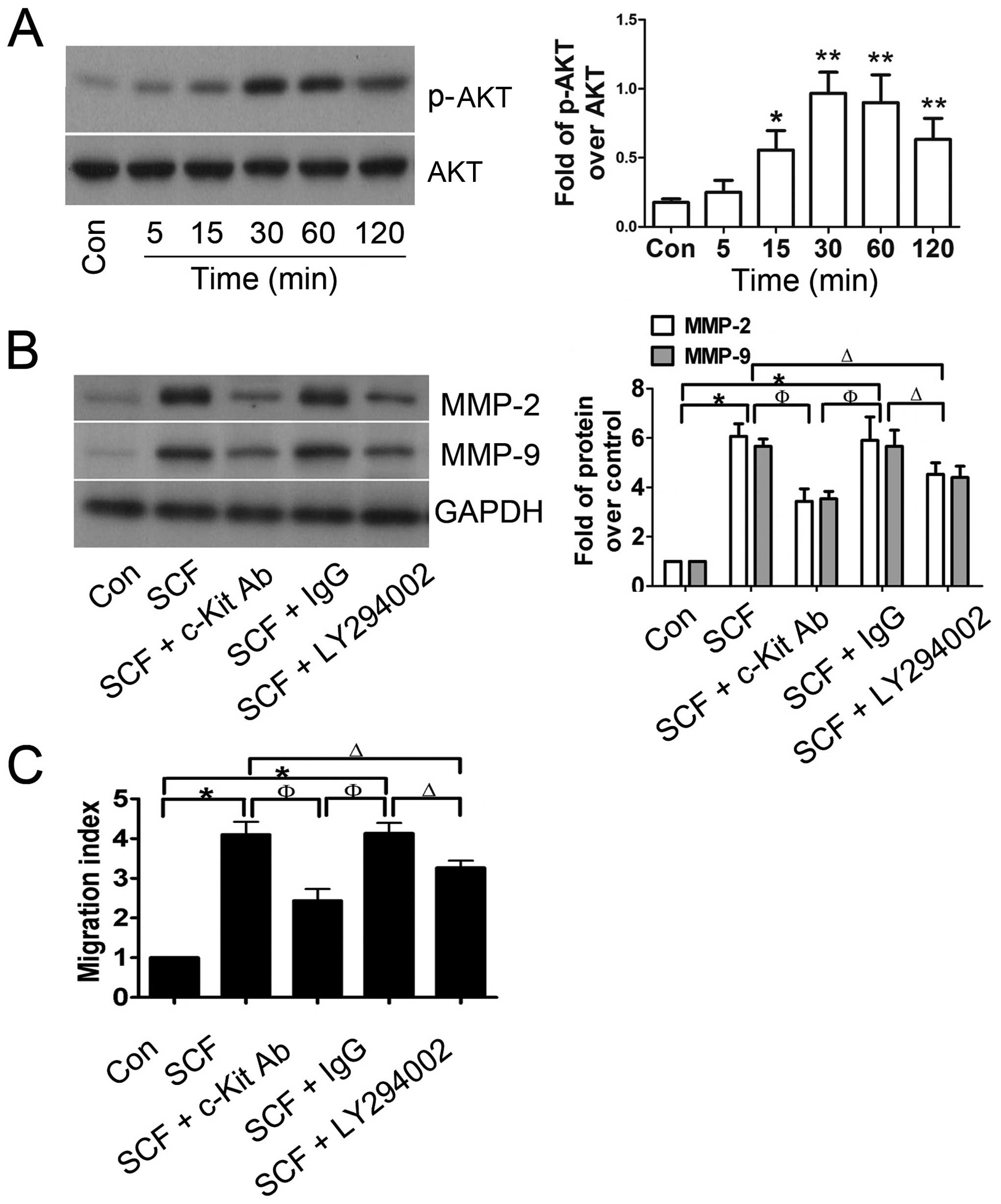
To explore whether SCF-induced CSC migration was
associated with the PI3K/AKT pathway, western blot analysis was
performed to detect the expression of total AKT and phospho-AKT
protein. As shown in Fig. 4A,
total AKT protein expression was not markedly altered, however, the
levels of phospho-AKT were significantly increased in SCF-treated
CSCs and reached a peak at 30 min after incubation with 30 ng/ml
SCF. Furthermore, CSCs were preincubated with c-Kit blocking
antibody or LY294002, resulting in significant suppression of the
SCF-mediated upregulation of MMP-2 and MMP-9 (Fig. 4B), which was matched with the
migration index of the CSCs (Fig.
4C). These results suggested that the PI3K/AKT pathway was
involved in SCF-induced CSC migration.
Discussion
Enzymatic digestion and the tissue expansion method
are standard approaches in the isolation of c-Kit+ CSCs.
Choi et al reported that the enzymatic digestion method is
more effective in isolating human c-Kit+ CSCs compared
with the tissue expansion method (26). Using the enzymatic digestion
method, the stem cell marker, c-Kit, is efficiently preserved and
the isolated CSCs proliferate much better than cells isolated
through the tissue expansion method. He et al confirmed that
MACS following enzymatic digestion is a simple but cost-effective
approach that can be used to obtain sufficient numbers of
stably-expressed c-Kit+ CSCs (27). Furthermore, the differentiation
potential of c-Kit+ CSCs is preserved after long-term
culture for 40 passages (28). In
the present investigation, we performed the enzymatic digestion
method plus MACS to isolate rat c-Kit+ CSCs, as
previously described (13,21).
As a result, the small, round c-Kit+ CSCs were
successfully obtained from adult rat hearts. After 4 weeks of
culture, cells reached confluence. These c-Kit+ CSCs
were positive for Nkx2.5 and GATA-4, the markers for early
cardiomyocyte commitment, which were in concordance with the
results of Choi et al (26). However, c-Kit+ CSCs
derived from newborn hearts were negative for transcription factors
and cytoplasmic proteins specific to cardiomycte (Troponin I),
smooth muscle cell (Tag1n) and endothelial cell (vwF) and
hematopoietic cells (29). Thus,
Nkx2.5+/GATA-4+/c-Kit+ CSCs from
adult rats may present a relatively late stage of cell
differentiation.
Activation of SCF/c-Kit signaling plays a crucial
role in a variety of cell biological functions, such as the
regulation of cell differentiation and proliferation, cell
apoptotic resistance, and mediation of cell migration through the
activation of downstream signaling molecules (30). Accumulating data have indicated
that the activation of SCF/c-Kit signaling plays a crucial role in
mediating stem cell migration and homing. Lutz et al
demonstrated that local injection of SCF improves myocardial homing
of systemically delivered c-Kit+ bone marrow-derived
stem cells (31), while Kuang
et al (21) showed that
SCF/c-Kit signaling mediated c-Kit+ CSC migration via
the activation of the downstream p38 cascade. On the other hand,
hyperglycemia impairs c-Kit+ CSC migration via a
reduction in the activity of ERK1/2 and p38 (32), and hyperhomocysteinemia inhibits
the homing of CSCs to peri-infarcted areas post-MI in rats with an
associated mechanism that may be due to the inhibition of NF-κB
(33). Consistent with the
above-mentioned reports, in the present investigation we found that
c-Kit+ CSC chemotaxis was promoted by exogenous SCF,
with its CI reaching a peak at a concentration of 30 ng/ml.
However, the migratory potential of c-Kit+ CSCs promoted
by exogenous SCF may be significantly abrogated by SCF antibodies.
In future studies, the mechanism of SCF/c-Kit signaling-mediated
CSC migration should be investigated.
Considering PI3K/AKT signaling is important for cell
survival, apoptosis and motility (3), we suggest that PI3K/AKT singling is
involved in the process of SCF/c-Kit signaling-mediated CSC
migration. The current results are consistent with our hypothesis
in that administration of the PI3K/AKT inhibitor LY294002 clearly
suppressed SCF-mediated CSC migration. Our results suggest that
there is a connection between SCF/c-Kit and PI3K/AKT signaling. A
previous report indicated that activation of the PI3K/AKT pathway
is crucial for VEGF-mediated c-Kit+ CSC migration in
vitro and in vivo (22). Furthermore, PI3K/AKT signaling is
important for infused stem cell survival post-MI (23–25). Thus, this pathway plays a clear
role in the biology of c-Kit+ CSCs, and deserves
additional attention in the field of MI cellular therapy based on
stem cell transplantation.
Cell migration is a complex and elaborately
regulated process. Several factors such as adhesion strength and
the type of substratum [including extracellular matrix (ECM)
ligands], external migratory signals and cues, mechanical
pliability, dimensionality, and the organization of the cellular
cytoskeleton determine the specifics of cellular motility (34,35). Among these, the complex
interactions of cells with ECM are crucial in mediating and
regulating cell migration (36).
As a critical factor mediating the interactions of cells with the
ECM, the abnormal expression of MMPs is thought to be an important
determinant in cell migration, and there is considerable evidence
that this occurs in cancerous cells. For instance, the upregulation
of MMP-2 and MMP-9 collagenases is associated with the invasion and
metastasis of cervical uterine neoplasm (37). Consistent with the results
obtained in cancer, the upregulation of MMP-2 and MMP-9 also
contributes to the migratory/invasive behavior of human mesenchymal
stem cells (hMSC) (38).
Furthermore, activation of PI3K/AKT signaling is sufficient to
induce MMP-2 and MMP-9 expression in erythropoietin-treated mouse
brain endothelial cells (39).
Therefore, we conducted experiments to investigate whether a high
expression of MMP-2 and MMP-9 contributed to SCF-enhanced CSC
motility. As expected, treatment of the c-Kit+ CSCs with
exogenous SCF effectively activated PI3K/AKT signaling and led to
the upregulation of MMP-2 and MMP-9 expression and activity in a
concentration- and time-dependent manner. We observed that c-Kit
blocking antibody or LY294002 significantly attenuated SCF-induced
MMP-2 and MMP-9 expression, which was accompanied by reduced CSC CI
values. Thus, it seems that activation of the
SCF/c-Kit/PI3K/AKT/MMP-2/-9 pathway is critical for CSC migration,
which is consistent with the rationale that hepatocyte growth
factor (HGF) increases the function of MMP-2 and MMP-9 in CSCs,
thus facilitating their migration (40).
Taken together, our current study provides evidence
that SCF partially mediated c-Kit+ CSC migration via the
activation of PI3K/AKTMMP-2/-9 signaling. A correlation between
SCF/c-Kit and PI3K/AKT signaling was identified, which explains
some of the diverse biological activity of CSCs. In conclusion,
these findings may assist in the progression of translational
medicine that utilizes CSCs in the repair of injured heart
tissue.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China ( nos. 81000073, 81160020 and
81170121), the Natural Science Foundation of Hainan Province (nos.
310043, 811197 and 812204), the Key Program of Science and
Technology of Hainan Province (ZDXM20100045), the Key Project of
Chinese Ministry of Education (212137) and HJHZ2013-06, and the
Technology Innovation Project of Department of Education of
Guangdong Province (2013KJCX0088).
References
|
1
|
World Health Organization. The top 10
causes of death. http://who.int/mediacentre/factsheets/fs310/en/.
Accessed July, 2013
|
|
2
|
Koudstaal S, Jansen Of, Lorkeers SJ, et
al: Concise review: heart regeneration and the role of cardiac stem
cells. Stem Cells Transl Med. 2:434–443. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hou J, Wang L, Jiang J, et al: Cardiac
stem cells and their roles in myocardial infarction. Stem Cell Rev.
9:326–338. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Karantalis V, Balkan W, Schulman IH,
Hatzistergos KE and Hare JM: Cell-based therapy for prevention and
reversal of myocardial remodeling. Am J Physiol Heart Circ Physiol.
303:H256–H270. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Bolli R, Tang XL, Sanganalmath SK, et al:
Intracoronary delivery of autologous cardiac stem cells improves
cardiac function in a porcine model of chronic ischemic
cardiomyopathy. Circulation. 128:122–131. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Ptaszek LM, Mansour M, Ruskin JN and Chien
KR: Towards regenerative therapy for cardiac disease. Lancet.
379:933–942. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Bolli R, Chugh AR, D’Amario D, et al:
Cardiac stem cells in patients with ischaemic cardiomyopathy
(SCIPIO): initial results of a randomised phase 1 trial. Lancet.
378:1847–1857. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Makkar RR, Smith RR, Cheng K, et al:
Intracoronary cardiosphere-derived cells for heart regeneration
after myocardial infarction (CADUCEUS): a prospective, randomised
phase 1 trial. Lancet. 379:895–904. 2012. View Article : Google Scholar
|
|
9
|
Beltrami AP, Barlucchi L, Torella D, et
al: Adult cardiac stem cells are multipotent and support myocardial
regeneration. Cell. 114:763–776. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Oh H, Bradfute SB, Gallardo TD, et al:
Cardiac progenitor cells from adult myocardium: homing,
differentiation, and fusion after infarction. Proc Natl Acad Sci
USA. 100:12313–12318. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Laugwitz KL, Moretti A, Lam J, et al:
Postnatal isl1+ cardioblasts enter fully differentiated
cardiomyocyte lineages. Nature. 433:647–653. 2005.
|
|
12
|
Winter EM, Grauss RW, Hogers B, et al:
Preservation of left ventricular function and attenuation of
remodeling after transplantation of human epicardium-derived cells
into the infarcted mouse heart. Circulation. 116:917–927. 2007.
View Article : Google Scholar
|
|
13
|
Guo J, Jie W, Kuang D, et al:
Ischaemia/reperfusion induced cardiac stem cell homing to the
injured myocardium by stimulating stem cell factor expression via
NF-kappaB pathway. Int J Exp Pathol. 90:355–364. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Williams AR, Hatzistergos KE, Addicott B,
et al: Enhanced effect of combining human cardiac stem cells and
bone marrow mesenchymal stem cells to reduce infarct size and to
restore cardiac function after myocardial infarction. Circulation.
127:213–223. 2013. View Article : Google Scholar
|
|
15
|
Ellison GM, Vicinanza C, Smith AJ, et al:
Adult c-kit(pos) cardiac stem cells are necessary and sufficient
for functional cardiac regeneration and repair. Cell. 154:827–842.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cui G, Shan L, Hung M, et al: A novel
Danshensu derivative confers cardioprotection via PI3K/Akt and Nrf2
pathways. Int J Cardiol. 168:1349–1359. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tekin D, Dursun AD and Xi L: Hypoxia
inducible factor 1 (HIF-1) and cardioprotection. Acta Pharmacol
Sin. 31:1085–1094. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gude NA, Emmanuel G, Wu W, et al:
Activation of Notch- mediated protective signaling in the
myocardium. Circ Res. 102:1025–1035. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Saxena A, Fish JE, White MD, et al:
Stromal cell-derived factor-1alpha is cardioprotective after
myocardial infarction. Circulation. 117:2224–2231. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang X, Chen Y, Kuang D, et al: The
cardioprotective effects of erythropoietin in myocardial ischemic
injury via upregulation of SDF-1 by JAK2/STAT3. Int J Cardiol.
156:320–322. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Kuang D, Zhao X, Xiao G, et al: Stem cell
factor/c-kit signaling mediated cardiac stem cell migration via
activation of p38 MAPK. Basic Res Cardiol. 103:265–273. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Tang J, Wang J, Kong X, et al: Vascular
endothelial growth factor promotes cardiac stem cell migration via
the PI3K/Akt pathway. Exp Cell Res. 315:3521–3531. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lovell MJ, Yasin M, Lee KL, et al: Bone
marrow mononuclear cells reduce myocardial reperfusion injury by
activating the PI3K/Akt survival pathway. Atherosclerosis.
213:67–76. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lu G, Haider HK, Jiang S and Ashraf M:
Sca-1+ stem cell survival and engraftment in the infarcted heart:
dual role for preconditioning-induced connexin-43. Circulation.
119:2587–2596. 2009.
|
|
25
|
Zhang Z, Li S, Cui M, et al: Rosuvastatin
enhances the therapeutic efficacy of adipose-derived mesenchymal
stem cells for myocardial infarction via PI3K/Akt and MEK/ERK
pathways. Basic Res Cardiol. 108:3332013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Choi SH, Jung SY, Suh W, Baek SH and Kwon
SM: Establishment of isolation and expansion protocols for human
cardiac C-kit-positive progenitor cells for stem cell therapy.
Transplant Proc. 45:420–426. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
He JQ, Vu DM, Hunt G, Chugh A, Bhatnagar A
and Bolli R: Human cardiac stem cells isolated from atrial
appendages stably express c-kit. PLoS One. 6:e277192011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Miyamoto S, Kawaguchi N, Ellison GM,
Matsuoka R, Shin’oka T and Kurosawa H: Characterization of
long-term cultured c-kit+cardiac stem cells derived from
adult rat hearts. Stem Cells Dev. 19:105–116. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Urbanek K, Cabral-da-Silva MC, Ide-Iwata
N, et al: Inhibition of notch1-dependent cardiomyogenesis leads to
a dilated myopathy in the neonatal heart. Circ Res. 107:429–441.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Liang J, Wu YL, Chen BJ, Zhang W, Tanaka Y
and Sugiyama H: The C-kit receptor-mediated signal transduction and
tumor-related diseases. Int J Biol Sci. 9:435–443. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Lutz M, Rosenberg M, Kiessling F, et al:
Local injection of stem cell factor (SCF) improves myocardial
homing of systemically delivered c-kit+bone
marrow-derived stem cells. Cardiovasc Res. 77:143–150. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
She T, Wang X, Gan Y, et al: Hyperglycemia
suppresses cardiac stem cell homing to peri-infarcted myocardium
via regulation of ERK1/2 and p38 MAPK activities. Int J Mol Med.
30:1313–1320. 2012.PubMed/NCBI
|
|
33
|
Wan J, Deng Y, Guo J, et al:
Hyperhomocysteinemia inhibited cardiac stem cell homing into the
peri-infarcted area post myocardial infarction in rats. Exp Mol
Pathol. 91:411–418. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Petrie RJ, Doyle AD and Yamada KM: Random
versus directionally persistent cell migration. Nat Rev Mol Cell
Biol. 10:538–549. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Van Haastert PJ and Devreotes PN:
Chemotaxis: signalling the way forward. Nat Rev Mol Cell Biol.
5:626–634. 2004.PubMed/NCBI
|
|
36
|
Berrier AL and Yamada KM: Cell-matrix
adhesion. J Cell Physiol. 213:565–573. 2007. View Article : Google Scholar
|
|
37
|
Libra M, Scalisi A, Vella N, et al:
Uterine cervical carcinoma: role of matrix metalloproteinases
(Review). Int J Oncol. 34:897–903. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Neth P, Ries C, Karow M, Egea V, Ilmer M
and Jochum M: The Wnt signal transduction pathway in stem cells and
cancer cells: influence on cellular invasion. Stem Cell Rev.
3:18–29. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Wang L, Zhang ZG, Zhang RL, et al: Matrix
metalloproteinase 2 (MMP2) and MMP9 secreted by
erythropoietin-activated endothelial cells promote neural
progenitor cell migration. J Neurosci. 26:5996–6003. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Urbanek K, Rota M, Cascapera S, et al:
Cardiac stem cells possess growth factor-receptor systems that
after activation regenerate the infarcted myocardium, improving
ventricular function and long-term survival. Circ Res. 97:663–673.
2005. View Article : Google Scholar
|