Atherosclerosis is a systemic disease affecting
multiple regions in the arterial wall, which is the primary cause
of myocardial infarction, stroke and other peripheral vascular
diseases. It is known to be involved in ongoing inflammatory
responses and processes (1), and
inflammation plays a central role in all phases of atherosclerotic
development (2). Thus,
atherosclerosis is also a progressive inflammatory disease of the
arterial wall (3), which was
formerly regarded as a lipid storage disease. In addition to
inflammation, the deposition of cholesterol in the arterial wall
plays an important role in the pathogenesis of atherosclerosis
(4). The condition causes
progressive smooth muscle cell (SMC) proliferation and migration
that contributes to vascular stenosis (5). Diabetes, obesity and dyslipidemia
are primary risk factors for the development of atherosclerosis
(6), which are also
epidemiologically linked to the increased susceptibility to a
various types of cancer (7). The
association between obesity and breast cancer, for example, is well
established (8). Atherosclerotic
plaques grow by the accumulation of inflammatory cells and lipid
substances, which is associated with the production of reactive
oxygen species (ROS). It has been demonstrated that ROS can
interact with cellular DNA to cause DNA strand breaks and/or base
modifications (9). These
oxidative modifications to DNA may cause a variety of mutations.
One of the prominent features of dyslipidemia is the enhanced
production of oxidized low-density lipoprotein (ox-LDL), which has
been implicated in the key steps of the proliferation of vascular
cells and atherogenesis (10). In
vascular cells, the internalization of ox-LDL has been shown to
trigger signaling events resulting in the overproduction of ROS,
inflammation and proliferation (11).
The cellular composition of an atherosclerotic
lesion is an important determinant of lesion stability (12). Therefore, the modulation of lesion
composition is important in order to reduce the risk of
atherosclerotic plaque rupture. Cell proliferation and cell death
are key processes in regulating the cellular composition of
atherosclerotic lesions (13).
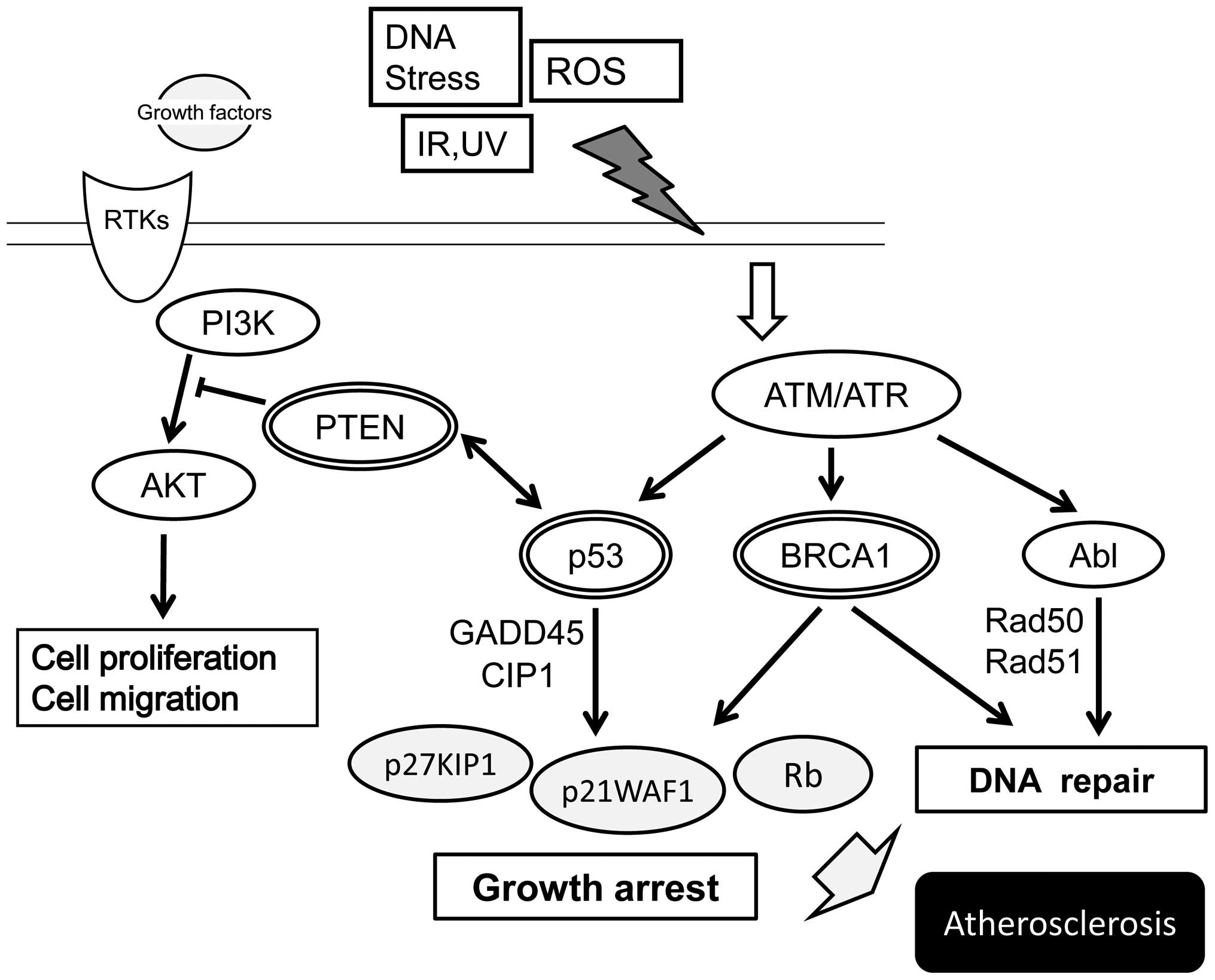
Tumor suppressor molecules play a pivotal role in regulating both
cell proliferation and cell death in a number of cell types,
although different genes also play important roles in the
initiation and modulation of atherosclerotic disease through
different mechanisms, either through up- or downregulation
(14) (Fig. 1). Consequently, tumor suppressor
genes involved in regulating cell proliferation and cell death may
play an important role in the progression of atherosclerotic
lesions, coinciding with changes in cellular composition. For
example, the deletion of the tumor suppressor gene, p53, an
essential molecule in cell proliferation, DNA repair and apoptosis,
strongly exacerbates atherosclerosis (15). In addition, the p53 downstream
target, p21WAF1, an inhibitor of cell cycle progression through the
inactivation of cyclin-cyclin-dependent kinase (CDK) complexes
during the G1 phase of the cell cycle, has pro-atherogenic
functions (16). These data
indicate the important role of tumor suppressor molecules in
controlling atherogenesis. In this review, the association between
atherosclerosis and certain tumor suppressor molecules is
summarized with focus on the pathogenesis of atherosclerosis, which
would essentially facilitate more effective treatments for a better
prognosis.
The proliferation of vascular smooth muscle cells
(VSMCs) contributes to a variety of pathological states including
atherosclerosis (17). Cell
invasion of the extracellular matrix is essential for the cross
tissue migration of VSMCs in atherosclerosis (18). The tumor suppressor, p53, has
gained attentiondue to its additional function as a suppressor of
cell migration and invasion. p53 also facilitates the apoptosis of
VSMCs and is involved in the ox-LDL-induced apoptosis of
macrophages (19). As much of the
genotoxic stress response flows through the p53 pathway, p53 is
considered to play an important role in the effects of genotoxic
stress. Furthermore, DNA damage disorders caused by mutations in
genotoxic stress-response genes are characterized by
atherosclerosis (Fig. 1). p53 is
ubiquitously expressed in all cell types as an inactive
transcription factor which undergoes activation in response to
various types of cellular stress. Evidence has implicated p53 as a
regulator of pathological vascular remodeling (20). The effects of p53 are mediated
through different downstream effectors and targets. Among these,
the CDK inhibitor, p21WAF1, is a key mediator of p53 action, which
may be involved in monocyte and dendritic cell differentiation
(21). There is substantial
evidence that p21WAF1 also participates in the protective effects
of p53 on atherosclerosis (22).
Generally, p21WAF1 is considered as a potential therapeutic gene
that may be used to treat or prevent atherosclerosis. p21WAF1
regulates cell-cycle progression, senescence and differentiation in
injured blood vessels, which may function to prevent atherogenesis
by regulating the redox balance, which leads to the inhibition of
adhesion molecules (23). The CDK
inhibitor, p27KIP1, also regulates cell proliferation, vascular
remodeling and inhibits atherosclerosis. While the expression of
tissue factors is a key initiator of the coagulation cascade
associated with atherosclerosis, p27KIP1 inhibits tissue factor
expression at the transcriptional level (24). In addition, a single nucleotide
polymorphism (838C>A) in the p27KIP1 gene is associated with a
commonly encountered genetic variant associated with therapeutic
cardiovascular interventions (25). Furthermore, the inactivation of
p27KIP1 has been shown to exacerbate atherosclerosis in a mouse
model (26). Minocycline has been
shown to reduce plaque size and stenosis in diet-induced
atherosclerosis through increased p27KIP1 expression in a mouse
model (27).
The residue-specific phosphorylation profile of the
retinoblastoma tumor suppressor (Rb) appears to differ between the
internal mammary artery and coronary artery (28). The differential profile of Rb
phosphorylation may be a consequence of variances in the content of
the CDK2 and CDK4 phosphorylation inhibitor, p15INK4.
siRNA-mediated CDK2 knockdown modifies the profile of Rb
phosphorylation in VSMCs of the coronary artery, as well as the
proliferative response of these cells to mitogenic stimulation
(28,29). The intrinsic functional and
protein composition specificity of the VSMC population in the
coronary artery may contribute to the increased incidence of
atherosclerosis in the arteries. In fact, foam cell formation is
induced through increased Rb phosphorylation (30). Apolipoprotein E (ApoE)-deficient
mice lacking macrophage Rb display accelerated atherosclerosis
coinciding with increased macrophage proliferation, suggesting that
macrophage Rb is a suppressive factor in the progression of
atherosclerosis by reducing macrophage cell proliferation (31). The phosphatase and tensin homolog
deleted on chromosome 10 (PTEN)/AKT signaling pathway has also been
implicated in the pathogenesis of vascular diseases. PTEN is a
dual-specificity phosphatase that has been shown to inhibit VSMC
proliferation and migration (32). Genetic research has demonstrted
that the PTEN gene is critical to the pathological development of
atherosclerosis (33). In
addition, elevated PTEN expression and concomitant AKT inactivation
have been observed in the endothelium of atherosclerotic arteries
(34). The PTEN pathway may be
important in the regulation of the inflammatory response in VSMCs.
The tumor suppressor breast cancer 1, early onset (BRCA1) gene
implicated in the development of breast and ovarian cancers exerts
multiple effects on DNA repair and affords resistance against
cellular stress responses (35),
which is basally expressed in endothelial cells. BRCA1 may be a
gatekeeper of inflammation-induced endothelial cell function
(36,37). Whereas BRCA1 silencing exaggerates
inflammation-induced endothelial cell apoptosis, BRCA1
overexpression protects cells againts this. BRCA1 overexpression
intensely attenuates the production of ROS, upregulating
endothelial nitric oxide synthase and vascular endothelial growth
factor expression. BRCA1 expression is attenuated in the plaque
region of human atherosclerotic arteries. It has been shown that
single nucleotide polymorphisms (SNPs) in the BRCA1-associated
protein (BRAP) gene are associasted with the risk of myocardial
infarction in a large cohort (38).
Again, cell proliferation and cell death are
important processes in regulating macrophage and VSMC numbers in
the atherosclerotic lesion, which may directly influence lesion
stability. Atherosclerosis is initiated by the subendothelial
accumulation of cholesterol-engorged macrophages (39). Atherosclerotic plaques contain
β-Gal-positive vascular endothelial cells (VECs) and VSMCs exhibit
the morphological features of senescence. It has also been reported
that interleukin-1β (IL-1β) is expressed in senescent cells located
in human atherosclerotic lesions, suggesting that senescent cells
may promote inflammation in lesions (40). Tumor suppressor molecules regulate
diverse cellular activities, including DNA damage repair, cell
proliferation, cell differentiation, cell migration, cellular
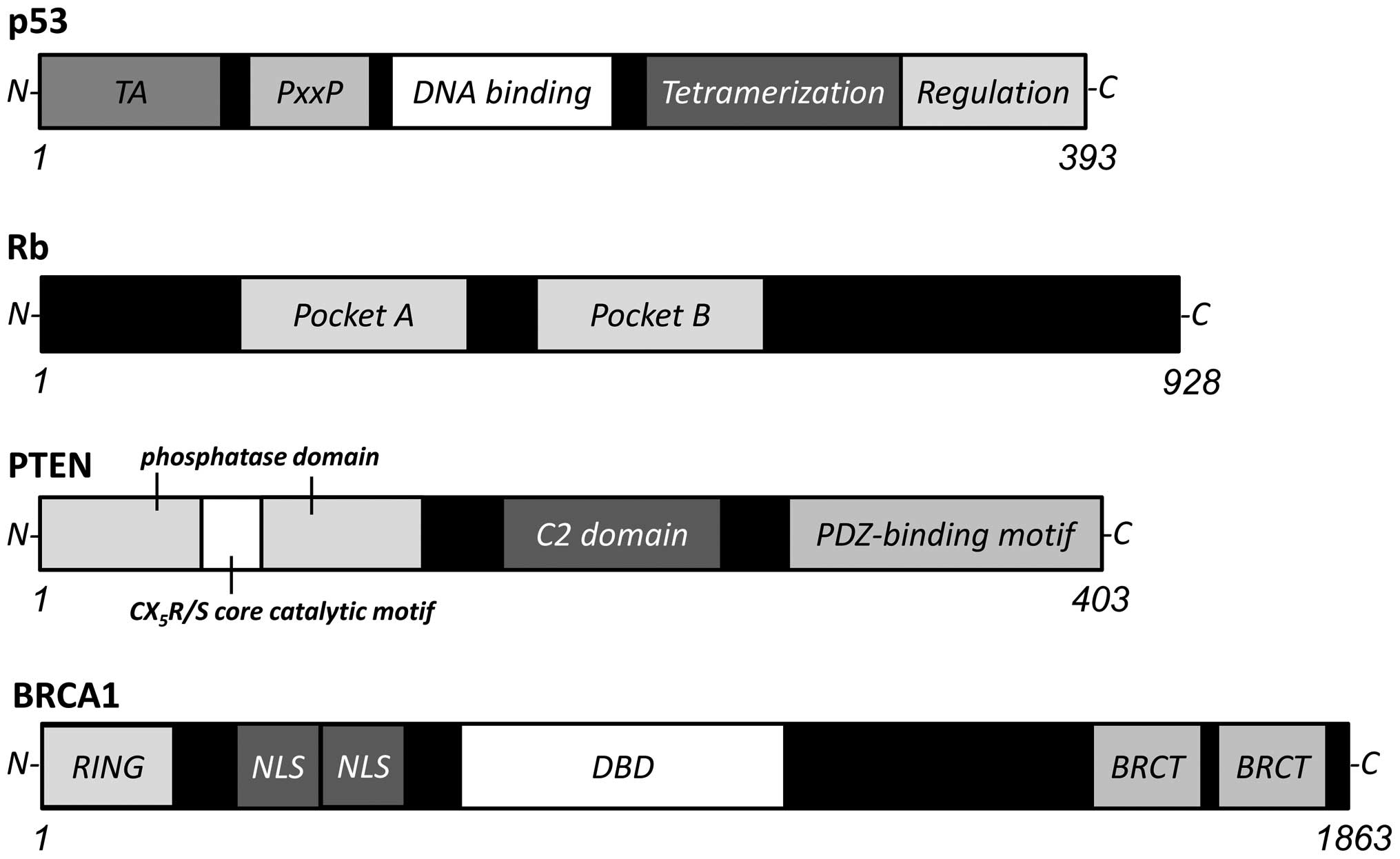
senescence and programmed cell death (Fig. 1). An important tumor suppressor is
the p53 tumor suppressor. Other examples of tumor suppressors,
include Rb, PTEN, p21WAF1, p27KIP1 and BRCA1 (Fig. 2) (18). Various cell proliferation- and
apoptosis-signal transduction pathways are built on complex
networks between oncogenes and tumor suppressor genes, such as p53
and its downstream factors. For example, the tumor suppressor p53
regulates the expression of various genes and plays an important
role in cell proliferation and in the modulation of signal
transduction pathways (41). The
accumulation of p53 in cells following DNA damage leads to cell
cycle arrest and the induction of apoptosis. In addition, p53 is
involved in the repair of damaged DNA and thus prevents the
accumulation of mutations and suppresses tumor development
(42). There are two types of p53
genes, the wild-type p53 gene and the mutant p53 gene (43). Oncogenic p53 mutations usually
confer the mutant protein with a dominant-negative activity over
the remaining wild-type gene. Many mutant p53 forms acquire
dominant-negative activities, and sometimes gain oncogenic
properties (43). These
activities of p53 are also regulated by post-translational
modification (44). The
phosphorylation and acetylation state, subcellular localization and
interaction with other cellular proteins are likely to influence
the function of p53 (45). In
cells facing oxidative stress and DNA-damage, p53 dissociates from
its ubiquitin ligase mouse double minute 2 homolog (MDM2) (46), through various post-translational
modifications which promote its stabilization.
Functional studies have linked p53 with
proliferative vascular disease. Arterial p53 inactivation following
human cytomegalovirus infection may contribute to coronary
restenosis (47). In addition,
p53-null mice are susceptible to atherosclerosis (48). However, the role of p53 in VSMC
proliferation and apoptosis in atherosclerosis is somewhat
controversial. Cell-specific p53 deficiency worsens the progression
of atheroma in animal models of diet-induced atherosclerosis
(49). The induction of p53
during ischemia has been shown to contribute to tissue damage
through the activation of apoptosis. By contrast, the temporary
inhibition of p53 function may be beneficial to the prevention of
injury to diverse organs (50) or
in the treatment of myocardial infarction (51). p53 may act to promote cell death
or survival depending on the cell type, gene expression profile,
protein activity and the type of stress stimuli (52). In addition to p53, a number of
cell cycle regulators modulate Rb function through its
phosphorylation status. Hypo-phosphorylated wild-type Rb is tightly
bound to the nuclear matrix and seems to be critical in the
inhibition of cellular proliferation (53). By contrast, hyper-phosphorylation
is a physiological mechanism of the inactivation of Rb. Active
complexes of G1 cyclins and CDKs inactivate Rb through its
phosphorylation (54), while
p21WAF1 and p27KIP1 inhibit CDKs. As reversible phosphorylation
plays a fundamental role in regulating intracellular signaling,
dysregulation of the mechanisms that regulate phosphorylation may
also play a role in the initiation and maintenance of
atherosclerosis.
The adenoviral vector-mediated delivery of the
p21WAF1 gene to the vessel wall has been shown to protect arteries
against the development of intimal hyperplasia (55), which also protects against
restenosis in ApoE-deficient mice by reducing VSMC proliferation
and macrophage infiltration (56). However, it has been shown that
p21WAF1 may also be a proatherogenic molecule. The inactivation of
p21WAF1 appears to protect against atherosclerosis, inhibiting
lesion growth and promoting its stability. Therapies that target
p21WAF1 for inactivation in the appropriate situation may offer
protection against atherosclerosis (57). PTEN upregulation induces
endothelial dysfunction by attenuating the availability and
signaling of multiple angiogenic pathways in VSMCs (34,58). However, adenovirus-mediated PTEN
overexpression inhibits the formation of vascular obstructive
lesions induced by mechanical injury. BRCA1 overexpression develops
less aortic plaque lesions, exhibits reduced macrophage
infiltration, and generates less ROS (36,59). Aortic segments from
BRCA1(−/−) mice have demonstrated more
inflammation-associated apoptosis and impaired endothelial function
(36,59). Mainly, the above-mentioned tumor
suppressor molecules may be regulators of systemic lipid
homeostasis and of the development of atherosclerosis.
Potential therapeutic strategies exploit the
observation that defects in critical processes required for
maintaining cellular homeostasis produce a metabolic situation
characterized by atherosclerosis (48,60). It has been reported that the free
cholesterol loading of macrophages induces apoptosis in
atherosclerosis and that apoptosis decreases after lipid lowering
(61). In addition, the
nutritional control of gene regulation guides the transformation of
VSMCs into foam cells in atherosclerosis. Actually, several gene
transcriptions are regulated by dietary polyunsaturated fatty acids
(62). Apoptosis in VSMCs can be
promoted by the deregulation of tumor suppressor molecules.
Furthermore, VSMCs in atherosclerotic plaques may be lost through
the apoptosis, increasing the risk of thrombosis. As p53 deficiency
leads to a substantial doubling of atherosclerotic lesion size, it
may be speculated that diets that stimulate p53 expression in
macrophages may lead to a reduction in atherosclerosis (63). Therefore, it would be important to
define appropriate strategies to achieve benefits from diet to
control the expression of tumor suppressor molecules.
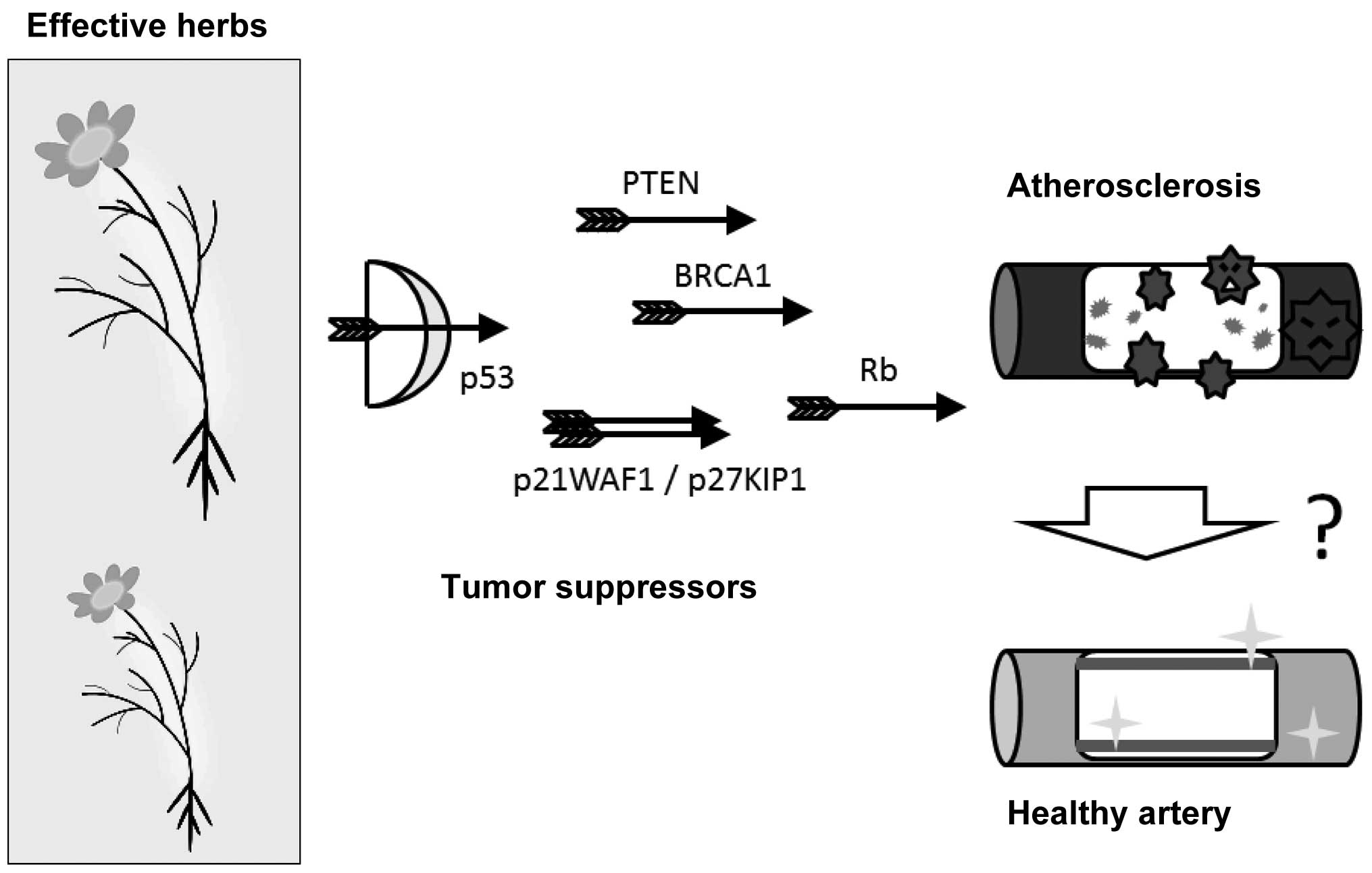
It will be a challenge to seek out how to use these
medicinal herbs for the correction in critical processes required
for maintaining cellular homeostasis linked to a metabolic
situation characterized by atherosclerosis (Fig. 3).
Atherosclerosis is likely to dominate clinical
practice for decades. The information presented herein may provide
further insight into the molecular mechanisms underlying the
clinical use of herbs as a therapy for atherosclerosis. The
identification of target molecules relevant for atherosclerosis
allows screening for natural products capable of modulating
targets. This may also represent the basis for the development of
rational dietary treatment of other diseases. Future studies are
required to demonstrate whether tumor suppressors and/or their
downstream targets can be used to modulate the cellular composition
of plaques, thereby enhancing plaque stability. Atherosclerosis is
caused by a number of factors, including oxidative stress which
increases sphingomyelinase and ceramides correlated with the level
of ox-LDL (83). During
atherogenesis, ROS are formed, resulting in oxidative DNA damage
which is followed by increased DNA repair activity so that initial
damage is effectively repaired. Tumor suppressors are key molecules
at this point. Further studies are required to elucidate the
potential roles of damaged DNA in the pathogenesis of
atherosclerosis. Further understanding of the local determinants of
the phenotype of endothelial cells in the lesion and how they
interact with atherosclerosis risk factors may lead to a notable
improvement in therapies.
This study was supported by Grants-in-Aid from the
Ministry of Education, Culture, Sports, Science and Technology in
Japan.
|
1
|
Libby P: Inflammation in atherosclerosis.
Nature. 420:868–874. 2002. View Article : Google Scholar
|
|
2
|
Spagnoli LG, Bonanno E, Sangiorgi G and
Mauriello A: Role of inflammation in atherosclerosis. J Nucl Med.
48:1800–1815. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Curtiss LK and Tobias PS: The toll of
Toll-like receptors, especially toll-like receptor 2, on murine
atherosclerosis. Curr Drug Targets. 8:1230–1238. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Cicchi R, Matthäus C, Meyer T, Lattermann
A, Dietzek B, Brehm BR, Popp J and Pavone FS: Characterization of
collagen and cholesterol deposition in atherosclerotic arterial
tissue using non-linear microscopy. J Biophotonics. 7:135–143.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Pakala R, Rha SW, Kuchulakanti PK, Cheneau
E, Baffour R and Waksman R: Peroxisome proliferator-activated
receptor gamma; Its role in atherosclerosis and restenosis.
Cardiovasc Radiat Med. 5:44–48. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Bays HE: Adiposopathy, diabetes mellitus,
and primary prevention of atherosclerotic coronary artery disease:
treating ‘sick fat’ through improving fat function with
antidiabetes therapies. Am J Cardiol. 110:B4–B12. 2012.PubMed/NCBI
|
|
7
|
Blüher M: Adipose tissue dysfunction in
obesity. Exp Clin Endocrinol Diabetes. 117:241–250. 2009.
|
|
8
|
Stoll BA: Western nutrition and the
insulin resistance syndrome: a link to breast cancer. Eur J Clin
Nutr. 53:83–87. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kligerman AD, Malik SI and Campbell JA:
Cytogenetic insights into DNA damage and repair of lesions induced
by a monomethylated trivalent arsenical. Mutat Res. 695:2–8. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lin FY, Lin YW, Huang CY, Chang YJ, Tsao
NW, Chang NC, Ou KL, Chen TL, Shih CM and Chen YH: GroEL1, a heat
shock protein 60 of Chlamydia pneumoniae, induces lectin-like
oxidized low-density lipoprotein receptor 1 expression in
endothelial cells and enhances atherogenesis in
hypercholesterolemic rabbits. J Immunol. 186:4405–4414. 2011.
View Article : Google Scholar
|
|
11
|
Dai Y, Mercanti F, Dai D, Wang X, Ding Z,
Pothineni NV and Mehta JL: LOX-1, a bridge between GLP-1R and
mitochondrial ROS generation in human vascular smooth muscle cells.
Biochem Biophys Res Commun. 437:62–66. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Dickhout JG, Basseri S and Austin RC:
Macrophage function and its impact on atherosclerotic lesion
composition, progression, and stability: the good, the bad, and the
ugly. Arterioscler Thromb Vasc Biol. 28:1413–1415. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Edlin RS, Tsai S, Yamanouchi D, Wang C,
Liu B and Kent KC: Characterization of primary and restenotic
atherosclerotic plaque from the superficial femoral artery:
potential role of Smad3 in regulation of SMC proliferation. J Vasc
Surg. 49:1289–1295. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Humphries SE and Ordovas JM: Genetics and
atherosclerosis: broadening the horizon. Atherosclerosis.
154:517–519. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Mercer J and Bennett M: The role of p53 in
atherosclerosis. Cell Cycle. 5:1907–1909. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Khanna AK: Enhanced susceptibility of
cyclin kinase inhibitor p21 knockout mice to high fat diet induced
atherosclerosis. J Biomed Sci. 15:16–66. 2009.PubMed/NCBI
|
|
17
|
Heusch G, Libby P, Gersh B, Yellon D, Böhm
M, Lopaschuk G and Opie L: Cardiovascular remodelling in coronary
artery disease and heart failure. Lancet. 383:1933–1943. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Mak AS: p53 in cell invasion, podosomes,
and invadopodia. Cell Adh Migr. Feb 18–2014.(Epub ahead of
print).
|
|
19
|
Ishii N, Matsumura T, Kinoshita H,
Motoshima H, Kojima K, Tsutsumi A, Kawasaki S, Yano M, Senokuchi T,
Asano T, Nishikawa T and Araki E: Activation of AMP-activated
protein kinase suppresses oxidized low-density lipoprotein-induced
macrophage proliferation. J Biol Chem. 284:34561–34569. 2009.
View Article : Google Scholar
|
|
20
|
Anderson JL, Ashwell CM, Smith SC, Shine
R, Smith EC and Taylor RL Jr: Atherosclerosis-susceptible and
atherosclerosis-resistant pigeon aortic cells express different
genes in vivo. Poult Sci. 92:2668–2680. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Liu WM, Scott KA, Thompson M and Dalgleish
AG: Dendritic cell phenotype can be improved by certain
chemotherapies and is associated with alterations to
p21(waf1/cip1.). Cancer Immunol Immunother. 62:1553–1561. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kim HJ, Yoo EK, Kim JY, Choi YK, Lee HJ,
Kim JK, Jeoung NH, Lee KU, Park IS, Min BH, Park KG, Lee CH, Aronow
BJ, Sata M and Lee IK: Protective role of clusterin/apolipoprotein
J against neointimal hyperplasia via antiproliferative effect on
vascular smooth muscle cells and cytoprotective effect on
endothelial cells. Arterioscler Thromb Vasc Biol. 29:1558–1564.
2009. View Article : Google Scholar
|
|
23
|
Obikane H, Abiko Y, Ueno H, Kusumi Y,
Esumi M and Mitsumata M: Effect of endothelial cell proliferation
on atherogenesis: a role of p21(Sdi/Cip/Waf1) in monocyte adhesion
to endothelial cells. Atherosclerosis. 212:116–122. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Breitenstein A, Akhmedov A, Camici GG,
Lüscher TF and Tanner FC: p27(Kip1) inhibits tissue factor
expression. Biochem Biophys Res Commun. 439:559–563. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Conte MS, Owens CD, Belkin M, Creager MA,
Edwards KL, Gasper WJ, Kenagy RD, LeBoeuf RC, Sobel M and Clowes A:
A single nucleotide polymorphism in the p27(Kip1) gene is
associated with primary patency of lower extremity vein bypass
grafts. J Vasc Surg. 57:1179–1185. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Díez-Juan A, Pérez P, Aracil M, Sancho D,
Bernad A, Sánchez-Madrid F and Andrés V: Selective inactivation of
p27(Kip1) in hematopoietic progenitor cells increases neointimal
macrophage proliferation and accelerates atherosclerosis. Blood.
103:158–161. 2004.
|
|
27
|
Shahzad K, Thati M, Wang H, Kashif M,
Wolter J, Ranjan S, He T, Zhou Q, Blessing E, Bierhaus A, Nawroth
PP and Isermann B: Minocycline reduces plaque size in diet induced
atherosclerosis via p27(Kip1). Atherosclerosis. 219:74–83. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Lange M, Fujikawa T, Koulova A, Kang S,
Griffin MJ, Lassaletta AD, Erat A, Tobiasch E, Bianchi C, Elmadhun
N, Sellke FW and Usheva A: Arterial territory-specific
phosphorylated retinoblastoma protein species and CDK2 promote
differences in the vascular smooth muscle cell response to
mitogens. Cell Cycle. 13:315–323. 2014. View Article : Google Scholar
|
|
29
|
Faber AC and Chiles TC: Inhibition of
cyclin-dependent kinase-2 induces apoptosis in human diffuse large
B-cell lymphomas. Cell Cycle. 6:2982–2989. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Ma KL, Liu J, Wang CX, Ni J, Zhang Y, Wu
Y, Lv LL, Ruan XZ and Liu BC: Activation of mTOR modulates SREBP-2
to induce foam cell formation through increased retinoblastoma
protein phosphorylation. Cardiovasc Res. 100:450–460. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Boesten LS, Zadelaar AS, van Nieuwkoop A,
Hu L, Jonkers J, van de Water B, Gijbels MJ, van der Made I, de
Winther MP, Havekes LM and van Vlijmen BJ: Macrophage
retinoblastoma deficiency leads to enhanced atherosclerosis
development in ApoE-deficient mice. FASEB J. 20:953–955. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chen WJ, Lin KH, Lai YJ, Yang SH and Pang
JH: Protective effect of propylthiouracil independent of its
hypothyroid effect on atherogenesis in cholesterol-fed rabbits:
PTEN induction and inhibition of vascular smooth muscle cell
proliferation and migration. Circulation. 110:1313–1319. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Yuan M, Wang X, Zhan Q, Duan X, Yang Q and
Xia J: Association of PTEN genetic polymorphisms with
atherosclerotic cerebral infarction in the Han Chinese population.
J Clin Neurosci. 19:1641–1645. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kuo HM, Lin CY, Lam HC, Lin PR, Chan HH,
Tseng JC, Sun CK, Hsu TF, Wu CC, Yang CY, Hsu CM and Tai MH: PTEN
overexpression attenuates angiogenic processes of endothelial cells
by blockade of endothelin-1/endothelin B receptor signaling.
Atherosclerosis. 221:341–139. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Bae I, Fan S, Meng Q, Rih JK, Kim HJ, Kang
HJ, Xu J, Goldberg ID, Jaiswal AK and Rosen EM: BRCA1 induces
antioxidant gene expression and resistance to oxidative stress.
Cancer Res. 64:7893–7909. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Singh KK, Shukla PC, Quan A, Al-Omran M,
Lovren F, Pan Y, Brezden-Masley C, Ingram AJ, Stanford WL, Teoh H
and Verma S: BRCA1 is a novel target to improve endothelial
dysfunction and retard atherosclerosis. J Thorac Cardiovasc Surg.
146:949–960. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lovren F, Pan Y, Quan A, Singh KK, Khan R,
Gupta N, Brezden-Masley C, Teoh H, Wheatcroft MD, Al-Omran M and
Verma S: BRCA1 shields vascular smooth muscle cells from oxidative
stress. J Thorac Cardiovasc Surg. 147:1946–1955. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Ozaki K, Sato H, Inoue K, Tsunoda T,
Sakata Y, Mizuno H, Lin TH, Miyamoto Y, Aoki A, Onouchi Y, Sheu SH,
Ikegawa S, Odashiro K, Nobuyoshi M, Juo SH, Hori M, Nakamura Y and
Tanaka T: SNPs in BRAP associated with risk of myocardial
infarction in Asian populations. Nat Genet. 41:329–333. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Jerome WG: Lysosomes, cholesterol and
atherosclerosis. Clin Lipidol. 5:853–865. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Suzuki E, Takahashi M, Oba S and
Nishimatsu H: Oncogene- and oxidative stress-induced cellular
senescence shows distinct expression patterns of proinflammatory
cytokines in vascular endothelial cells. ScientificWorldJournal.
2013:7547352013. View Article : Google Scholar
|
|
41
|
Boominathan L: The guardians of the genome
(p53, TA-p73, and TA-p63) are regulators of tumor suppressor miRNAs
network. Cancer Metastasis Rev. 29:613–639. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
O’Neil N and Rose A: DNA repair. WormBook.
1–12. 2006.
|
|
43
|
Muller PA and Vousden KH: 53 mutations in
cancer. Nat Cell Biol. 15:2–8. 2013. View Article : Google Scholar
|
|
44
|
Conforti F, Sayan AE, Sreekumar R and
Sayan BS: Regulation of p73 activity by post-translational
modifications. Cell Death Dis. 3:e2852012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Appella E and Anderson CW:
Post-translational modifications and activation of p53 by genotoxic
stresses. Eur J Biochem. 268:2764–2772. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Hock AK and Vousden KH: The role of
ubiquitin modification in the regulation of p53. Biochim Biophys
Acta. 1843:137–149. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Mattila KJ, Valtonen VV, Nieminen MS and
Asikainen S: Role of infection as a risk factor for
atherosclerosis, myocardial infarction, and stroke. Clin Infect
Dis. 26:719–734. 1998. View
Article : Google Scholar : PubMed/NCBI
|
|
48
|
Razani B, Feng C and Semenkovich CF: p53
is required for chloroquine-induced atheroprotection but not
insulin sensitization. J Lipid Res. 51:1738–1746. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Martinet W, Knaapen MW, De Meyer GR,
Herman AG and Kockx MM: Oxidative DNA damage and repair in
experimental atherosclerosis are reversed by dietary lipid
lowering. Circ Res. 88:733–739. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Georgiev P, Dahm F, Graf R and Clavien PA:
Blocking the path to death: anti-apoptotic molecules in
ischemia/reperfusion injury of the liver. Curr Pharm Des.
12:2911–2921. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Matsusaka H, Ide T, Matsushima S, Ikeuchi
M, Kubota T, Sunagawa K, Kinugawa S and Tsutsui H: Targeted
deletion of p53 prevents cardiac rupture after myocardial
infarction in mice. Cardiovasc Res. 70:457–465. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Kim YC, Kitaura H, Taira T, Iguchi-Ariga
SM and Ariga H: Oxidation of DJ-1-dependent cell transformation
through direct binding of DJ-1 to PTEN. Int J Oncol. 35:1331–1341.
2009.PubMed/NCBI
|
|
53
|
Markiewicz E, Ledran M and Hutchison CJ:
Remodelling of the nuclear lamina and nucleoskeleton is required
for skeletal muscle differentiation in vitro. J Cell Sci.
118:409–420. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Wikman H and Kettunen E: Regulation of the
G1/S phase of the cell cycle and alterations in the RB pathway in
human lung cancer. Expert Rev Anticancer Ther. 6:515–530. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Smith RC, Branellec D, Gorski DH, Guo K,
Perlman H, Dedieu JF, Pastore C, Mahfoudi A, Denèfle P, Isner JM
and Walsh K: p21CIP1-mediated inhibition of cell proliferation by
overexpression of the gax homeodomain gene. Genes Dev.
11:1674–1689. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Condorelli G, Aycock JK, Frati G and
Napoli C: Mutated p21/WAF/CIP transgene overexpression reduces
smooth muscle cell proliferation, macrophage deposition,
oxidation-sensitive mechanisms, and restenosis in
hypercholesterolemic apolipoprotein E knockout mice. FASEB J.
15:2162–2170. 2001. View Article : Google Scholar
|
|
57
|
Merched AJ and Chan L: Absence of
p21Waf1/Cip1/Sdi1 modulates macrophage differentiation and
inflammatory response and protects against atherosclerosis.
Circulation. 110:3830–3841. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Brader S and Eccles SA: Phosphoinositide
3-kinase signalling pathways in tumor progression, invasion and
angiogenesis. Tumori. 90:2–8. 2004.PubMed/NCBI
|
|
59
|
Musumeci M, Maccari S, Corritore E,
Massimi A, Stati T, Marano G and Catalano L: Signaling
pathway-focused gene expression profiling in pressure overloaded
hearts. Ann Ist Super Sanita. 47:290–295. 2011.PubMed/NCBI
|
|
60
|
Ouimet M: Autophagy in obesity and
atherosclerosis: Interrelationships between cholesterol
homeostasis, lipoprotein metabolism and autophagy in macrophages
and other systems. Biochim Biophys Acta. 1831:1124–1133. 2013.
View Article : Google Scholar
|
|
61
|
Yao PM and Tabas I: Free cholesterol
loading of macrophages induces apoptosis involving the fas pathway.
J Biol Chem. 275:23807–23813. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Waters SM, Coyne GS, Kenny DA and Morris
DG: Effect of dietary n-3 polyunsaturated fatty acids on
transcription factor regulation in the bovine endometrium. Mol Biol
Rep. 41:2745–2755. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Merched AJ, Williams E and Chan L:
Macrophage-specific p53 expression plays a crucial role in
atherosclerosis development and plaque remodeling. Arterioscler
Thromb Vasc Biol. 23:1608–1614. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Lee SJ, Park K, Ha SD, Kim WJ and Moon SK:
Gleditsia sinensis thorn extract inhibits human colon cancer
cells: the role of ERK1/2, G2/M-phase cell cycle arrest and p53
expression. Phytother Res. 24:1870–1876. 2010. View Article : Google Scholar
|
|
65
|
Lee SJ, Park SS, Kim WJ and Moon SK:
Gleditsia sinensis thorn extract inhibits proliferation and
TNF-α-induced MMP-9 expression in vascular smooth muscle cells. Am
J Chin Med. 40:373–386. 2012. View Article : Google Scholar
|
|
66
|
Lu Y, Li CS and Dong Q: Chinese herb
related molecules of cancer-cell-apoptosis: a minireview of
progress between Kanglaite injection and related genes. J Exp Clin
Cancer Res. 27:312008. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Li B, Zhao J, Wang CZ, et al: Ginsenoside
Rh2 induces apoptosis and paraptosis-like cell death in colorectal
cancer cells through activation of p53. Cancer Lett. 301:185–192.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Gali-Muhtasib H, Diab-Assaf M, Boltze C,
et al: Thymoquinone extracted from black seed triggers apoptotic
cell death in human colorectal cancer cells via a p53-dependent
mechanism. Int J Oncol. 25:857–866. 2004.PubMed/NCBI
|
|
69
|
Lee SJ, Kim HM, Cho YH, et al: Aqueous
extract of Magnolia officinalis mediates proliferative
capacity, p21WAF1 expression and TNF-α-induced NF-κB activity in
human urinary bladder cancer 5637 cells; involvement of p38 MAP
kinase. Oncol Rep. 18:729–736. 2007.
|
|
70
|
Dong LH, Wen JK, Miao SB, et al: Baicalin
inhibits PDGF-BB-stimulated vascular smooth muscle cell
proliferation through suppressing PDGFRβ-ERK signaling and increase
in p27 accumulation and prevents injury-induced neointimal
hyperplasia. Cell Res. 20:1252–1262. 2010.PubMed/NCBI
|
|
71
|
Way TD, Lee JC, Kuo DH, et al: Inhibition
of epidermal growth factor receptor signaling by Saussurea
involucrata, a rare traditional Chinese medicinal herb, in
human hormone-resistant prostate cancer PC-3 cells. J Agric Food
Chem. 58:3356–3365. 2010.PubMed/NCBI
|
|
72
|
Hahm ER and Singh SV: Honokiol causes
G0–G1 phase cell cycle arrest in human prostate cancer cells in
association with suppression of retinoblastoma protein
level/phosphorylation and inhibition of E2F1 transcriptional
activity. Mol Cancer Ther. 6:2686–2695. 2007.
|
|
73
|
Tao R, Lu L, Zhang R, Hu J, Ni J and Shen
W: Triptolide inhibits rat vascular smooth muscle cell
proliferation and cell cycle progression via attenuation of ERK1/2
and Rb phosphorylation. Exp Mol Pathol. 90:137–142. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Shan BE, Zeki K, Sugiura T, Yoshida Y and
Yamashita U: Chinese medicinal herb, Acanthopanax
gracilistylus, extract induces cell cycle arrest of human tumor
cells in vitro. Jpn J Cancer Res. 91:383–389. 2000.
|
|
75
|
Xiao XY, Hao M, Yang XY, et al:
Licochalcone A inhibits growth of gastric cancer cells by arresting
cell cycle progression and inducing apoptosis. Cancer Lett.
302:69–75. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Kametani S, Oikawa T, Kojima-Yuasa A, et
al: Mechanism of growth inhibitory effect of cape aloe extract in
ehrlich ascites tumor cells. J Nutr Sci Vitaminol (Tokyo).
53:540–546. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
77
|
Yang P, Cartwright C, Chan D, Vijjeswarapu
M, Ding J and Newman RA: Zyflamend-mediated inhibition of human
prostate cancer PC3 cell proliferation: effects on 12-LOX and Rb
protein phosphorylation. Cancer Biol Ther. 6:228–236. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Liu H, Zang C, Emde A, et al: Anti-tumor
effect of honokiol alone and in combination with other anti-cancer
agents in breast cancer. Eur J Pharmacol. 591:43–51. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Yang JY, Della-Fera MA, Rayalam S and
Baile CA: Enhanced effects of xanthohumol plus honokiol on
apoptosis in 3T3-L1 adipocytes. Obesity (Silver Spring).
16:1232–1238. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
Roy S, Yu Y, Padhye SB, Sarkar FH and
Majumdar AP: Difluorinated-curcumin (CDF) restores PTEN expression
in colon cancer cells by down-regulating miR-21. PLoS One.
8:e685432013. View Article : Google Scholar : PubMed/NCBI
|
|
81
|
Yoshida H, Okumura N, Kitagishi Y,
Nishimura Y and Matsuda S: Ethanol extract of rosemary repressed
PTEN expression in K562 culture cells. Int J Appl Boil Pharm
Technol. 2:316–322. 2011.
|
|
82
|
Bosviel R, Dumollard E, Déchelotte P,
Bignon YJ and Bernard-Gallon D: Can soy phytoestrogens decrease DNA
methylation in BRCA1 and BRCA2 oncosuppressor genes in breast
cancer? OMICS. 16:235–244. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Kobayashi K, Nagata E, Sasaki K,
Harada-Shiba M, Kojo S and Kikuzaki H: Increase in secretory
sphingomyelinase activity and specific ceramides in the aorta of
apolipoprotein E knockout mice during aging. Biol Pharm Bull.
36:1192–1196. 2013. View Article : Google Scholar : PubMed/NCBI
|