Introduction
Various diseases, including hereditary hepatic
metabolic disorders, hepatic viral infection, autoimmune disorder,
as well as chemical agents, such as alcohol can lead to hepatic
injury. As a result of alcohol addiction, long-term alcohol
consumption becomes one of the leading causes of chronic hepatic
disease (1). It has been reported
that the risk of developing hepatitis and hepatic fibrosis
increases significantly if an individual ingests alcohol everyday
(60–80 g for males and 20 g for females) in over an extented period
of time (2). As the prequel to
cirrhosis, hepatic fibrosis is characterized by excessive
extracellular matrix (ECM) generation, which impairs normal hepatic
function (3). There are positive
correlations between hepatic fibrosis and hepatic cirrhosis/hepatic
cellular carcinoma (HCC), which are the leading causes of mortality
in patients with liver diseases (4).
In response to hepatic damage, hepatic stellate
cells (HSCs), which take charge of hepatic ECM secretion and
deposition, are often activated (5). The activation of HSCs is
characterized by the transformation of HSCs from a quiescent to a
proliferative and profibrogenic state (6). It is believed that activated HSCs
are the main source of collagen and tissue inhibitors of
metalloproteinase (TIMPs) during hepatic fibrosis (7). Although multiple factors may
participate in HSC activation, including growth factors,
inflammatory factors and free radicals, evidence strongly suggests
an association with the transforming growth factor-β (TGF-β)
signaling pathway (8). The
TGF-β/Smad signaling pathway is activated by TGF-β binding to its
receptors located on the cell membrane. The downstream proteins,
namely Smad2 and Smad3, are activated by phosphorylation (9), which further promotes the
transcription of genes conducting ECM protein synthesis (10). Additionally, in this progress,
another Smad protein, Smad7, acts as the inhibitor of the
TGF-β/Smad signaling pathway through its ubiquitin-proteasome
degradation activity (11).
Since ancient times, herbal medicine has been used
in China and India. Some effective ingredients extracted from
herbs, such as baicalein, emodin, ginsenoside and curcumin have
become hot research topics due to their efficacy against diseases,
such as inflammatory diseases, cancer, viral infections, ischemia
and chronic hepatic disease (12–15). Curcumin, also known as
1,7-bis(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione, is an
important phytochemical extracted from a herbal plant known as
turmeric (Curcuma longa Linn.). Curcumin possesses a variety
of biological properties, including antioxidant, anti-carcinogenic
and anti-inflammatory properties (16). The hepatoprotective effects of
curcumin against hepatic fibrosis have previously been demonstrated
(17). Furthermore, it has been
demonstrated that the mechanisms of action of curcumin in
inhibiting hepatic fibrosis correlate with the inhibition of the
TGF-β/Smad signaling pathway (18). However, to the best of our
knowledge, the majority of studies on the anti-fibrotic effects of
curcumin have been based on carbon tetrachloride-induced hepatic
fibrosis, which does not mimic the toxicity of alcohol
specifically. Moreover, there are limited studies available that
have focused on the effects of curcumin on HSCs incubated in
alcoholic conditions; these cells may be one of the critical
mechanisms through which curcumin exerts its effects against
hepatic fibrosis.
In the present study, we assessed and investigated
the capacity and possible mechanisms of action of curcumin in
alleviating alcohol-induced hepatic fibrosis in rats. Additionally,
an in vitro investigation was also conducted to evaluate the
effects of curcumin on the alcohol-induced activation of rat HSCs.
Curcumin may be considered as a possible therapeutic target in
alcohol-induced hepatic fibrosis.
Materials and methods
Animal grouping and treatment
Thirty male Sprague-Dawley (SD) rats (6 weeks old;
mean body weight, 205±25 g) purchased from the Animal Experimental
Center of Xi’an Jiaotong University, Xi’an, China were used in this
study. The animals were raised in separate polypropylene cages for
1 week prior to treatment under controlled conditions (artificial
12/12 h-cycle; mean temperature of 22±1°C; humidity at 65±4%). The
animal experimental procedures were carried out strictly in
accordance with the Guide for the Care and Use of Laboratory
Animals of the National Institutes of Health (NIH) and approved by
the Medical Animal Research Ethics Committee of Xi’an Jiaotong
University. The rats were divided into 3 groups evenly and
randomly, including a control group (Ctrl, n=10), alcohol group
(Alc, n=10) and alcohol + curcumin group (Alc + Cur, n=10). Hepatic
fibrosis was induced by the oral administration of alcohol as
described in a previous study (19). Rats in the Alc group received an
oral administration of ethanol solution [30% v/v, intragastric
(i.g.), 10 ml/kg/day] for 4 weeks; rats in the Alc + Cur group
received an oral administration of ethanol solution (30% v/v, i.g.,
10 ml/kg/day) for 4 weeks and then curcumin solution [suspended in
phosphate-buffered saline (PBS), i.g., 200 mg/kg, 300 mg/kg body
weight) for 12 weeks (20).
Assessment of hepatic function
Whole blood sample from rats were collected by
femoral artery puncture when the rats were sacrificed by an
overdose of chloral hydrate by intraperitoneal injection (10% v/v).
The samples were delivered to the Clinical Laboratory at the The
First Affiliated Hospital of Medical College, Xi’an Jiaotong
University. The activities of alkaline phosphatase (ALP), aspartate
aminotransferase (AST) and alanine aminotransferase (ALT) in serum
were detected using an enzyme coupling rate method.
Histopathological and immunohistochemical
evaluation
Immediately after harvesting, hepatic tissue was
fixed by neutral buffered formalin (10% v/v, pH 7.4), embedded in
paraffin and then sectioned at a thickness of 4 μm. Tissue slices
were stained with fibrosis-specific staining, Sirius red staining,
in accordance with the protocols described in previous studies
(21,22). The prepared sections were also
subjected to immunohistochemical staining by incubation with
antibodies against α-smooth muscle actin (α-SMA) and collagen-I
(Coll-I) (both from Abcam, Cambridge, UK). After images were
acquired, the quantification of the positively stained areas
expressed as optical density (OD) was performed using ImageJ
software (version 1.43b; NIH, Bethesda, MD, USA).
Cell culture and treatments
HSCs were isolated from the livers of the SD rats
(body weight, 101±7 g). The isolation procedures were implemented
following previously described protocols (23). The purity of the isolated HSCs was
evaluated by immunohistochemistry using antibody against desmin
(Abcam) and intrinsic vitamin A autofluorescence under a
phase-contrast microscope (Model CH20; Olympus, Tokyo, Japan). Cell
viability was evaluated by trypan blue exclusion assay. Cell purity
and viability exceeded 90%. The cells were maintained in culture
flasks (Corning, Inc., Corning, NY, USA) containing minimum
essential medium (MEM) supplemented with 5% fetal calf serum (FCS)
(both from Gibco, Carlsbad, CA, USA), 1% non-essential amino acid
(NEA; HyClone, Logan, UT, USA) and penicillin-streptomycin (100
U/ml; Invitrogen, Carlsbad, CA, USA) in a humidified atmosphere of
5% CO2 at 37°C. After being washed with sterilized PBS,
the cells were subcultured after trypsinization twice per week. The
effects of curcumin on the HSCs were investigated by pre-treatment
with curcumin at serial concentrations (0, 10, 20, 30, 40, 50 and
60 μmol/l). Equal amounts of cells were assigned to the Ctrl, Alc
and Alc + Cur groups. Cells in the Alc + Cur group were treated
with ethanol (50 mmol/l; Sigma-Aldrich (St. Louis, MO, USA) for 24
h and then treated with curcumin (50 mmol/l; Sigma-Aldrich) for a
further 24 h; cells in the Alc group were only treated with ethanol
for 24 h. The cells were then cultured in a humidified atmosphere
of 5% CO2 at 37°C for 24 h.
Cell viability assay
A colorimetric
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay was used to assess the cell viability in the present study.
Briefly, cells from each group planted in a 96-well culture plate
(Corning, Inc.) for 24 h were washed with sterilized PBS and then
incubated with MTT (5 mg/ml; Sigma-Aldrich) for 4 h. After being
washed with sterilized PBS, the cells were then dissolved in
dimethyl sulfoxide (DMSO; Sigma-Aldrich). The absorbance value at
450 nm was read using a 96-well plate reader (Bio-Rad, Hercules,
CA, USA). The ratio of the number of viable cells in the
experimental wells to the control wells was used to express the
results of cell viability.
Assessment of cell apoptosis
Cell apoptosis was evaluated by flow cytometry.
Cells at a density of 5×103 cells/well in each group
were collected for the analysis of apoptosis by V-FITC and
propidium iodide (PI) (both from Santa Cruz Biotechnology, Inc.,
Santa Cruz, CA, USA) double staining using a flow cytometer
(FACSCalibur; BD Biosciences, Franklin Lakes, NJ, USA) in
accordance with the protocol described in a previous study
(24).
Real-time reverse
transcription-polymerase chain reaction (PCR)
From both rat hepatic tissue and cultured HSCs,
total RNA was extracted using the RNAfast 200 kit (Fastagen Biotech
Co., Ltd., Shanghai, China) then reverse transcription was
performed using the PrimeScript RT reagent kit (Takara Bio, Inc.,
Shiga, Japan). SYBR Premix Ex Taq™ II (Takara Bio, Inc.) was used
to perform real-time PCR and the results were then detected using a
PRISM 7500 real-time PCR detection system (Applied Biosystems,
Foster City, CA, USA). Oligonucleotide primers for α-SMA,
collagen-I, TGF-β1, fibronectin, Smad7 and glyceraldehyde
3-phosphate dehydrogenase (GAPDH) were synthesized by Takara Bio,
Inc. Relative mRNA expression levels were calculated by Δcycle
threshold (ΔCt = CtTarget-CtGAPDH) and GAPDH
was introduced as an internal reference gene.
Western blot analysis
Both the rat hepatic tissue and cultured HSCs were
homogenized using lysis buffer (40 mmol/l Tric-HCl, 150 mmol/l KCl,
1 mmol/l EDTA, 100 mmol/l NaVO3, 1% Triton X-100) with
PMSF (1 mmol/l; Santa Cruz Biotechnology, Inc.). After
concentration detection using the BCA protein assay kit (Santa Cruz
Biotechnology, Inc.), 50 μg protein sample were separated by
vertical sodium dodecyl sulfate-polyacrylamide gel (10 or 8%)
electrophoresis and then transferred onto PVDF membranes which were
subsequently blocked in defatted milk [5%, dissolved in
Tris-buffered saline with Tween-20 (TBST) buffer] to block
non-specific binding. Immunoblots were then detected using specific
antibodies against collagen-I, fibronectin, α-SMA, TGF-β1 and
proliferating cell nuclear antigen (PCNA) (all purchased from
Abcam), Smad3, phospho-Smad3, Smad7 (all from Cell Signaling
Technology, Danvers, MA, USA), glucose-regulated protein-78
(GRP-78), CCAAT/enhancer-binding protein homologous protein (CHOP)
(both from Abcam) and GAPDH (Santa Cruz Biotechnology, Inc.). After
the unbounded antibodies were washed with TBST-Tween-20 (0.02%),
the membranes were incubated with antibody conjugated to HRP (Santa
Cruz Biotechnology, Inc.) for 2 h at 37°C. Finally, the immunoblots
were visualized and determined using ECL western blotting detection
reagent (Amersham Pharmacia Biotech, Piscataway, NJ, USA). ImageJ
software (version 1.43b; NIH) was then used to determine and
quantify the density of the bands.
Statistical analysis
Data collected in this study are expressed as the
means ± standard deviation (SD). All statistic analyses were
performed using SPSS software (version 15.0; SPSS Inc., Chicago,
IL, USA). Differences in parameters among the groups were analyzed
by one-way analysis of variance followed by the Nweman-Keuls
post-hoc test. Differences were considered statistically
significant with P-values <0.05.
Results
Curcumin alleviates alcohol-induced
hepatic functional damage and fibrosis in vivo
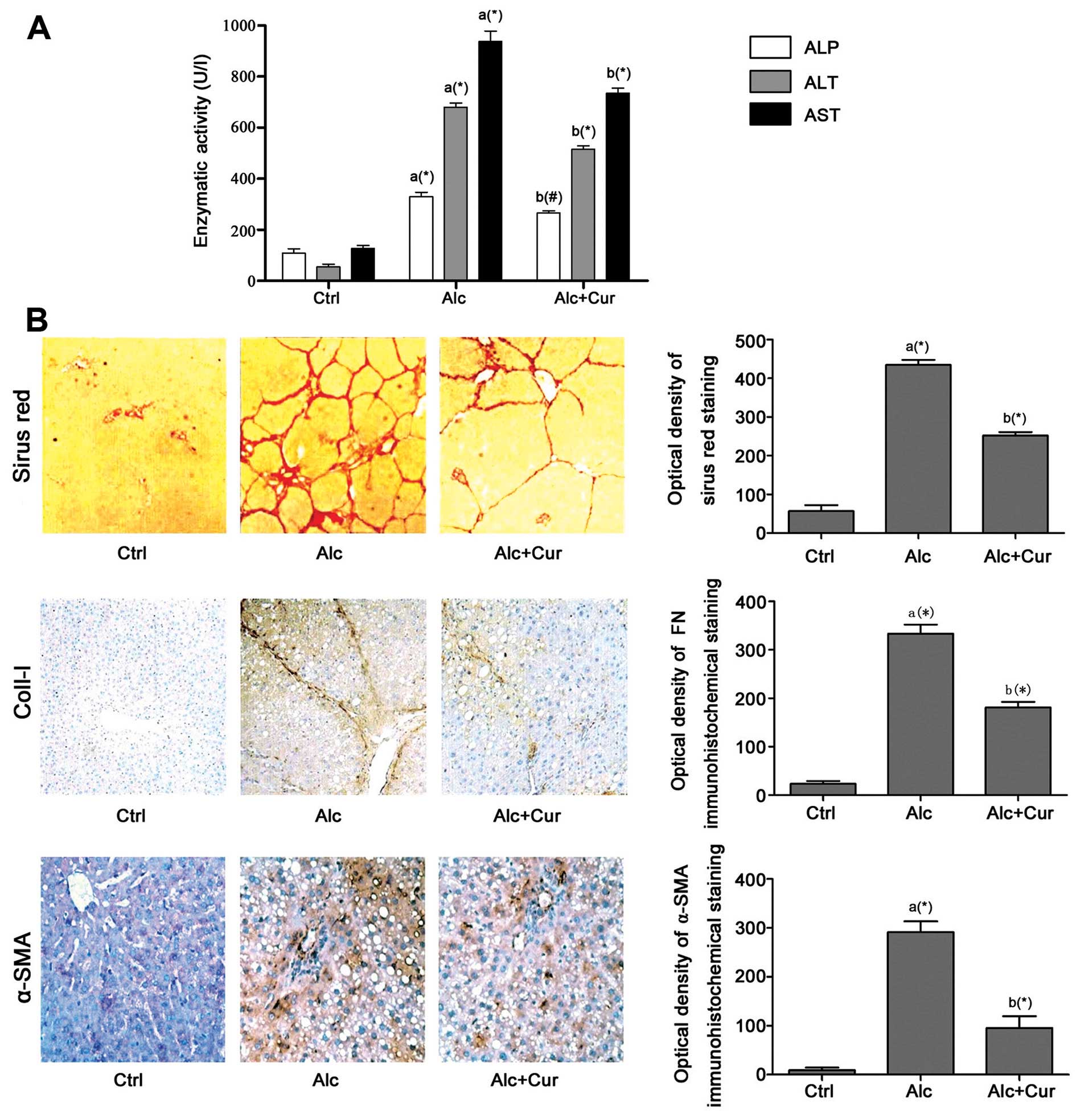
The administration of alcohol for 4 weeks
significantly impaired hepatic function, which was evidenced by the
marked increase in serum ALP, ALT and AST levels in the Alc group
compared with the Ctrl group (Fig.
1). By contrast, treatment with curcumin (Alc + Cur group)
resulted in a marked decrease in the serum levels of AST and ALT
compared with the Alc group. The therapeutic effects of curcumin
against hepatic fibrosis were then examined by histopathological
and immunohistochemical staining. Sirius red staining was used to
stain hepatic collagen fiber. The optical densities of the
positively stained areas in Sirius red staining, and collagen-I
staining increased significantly in the Alc group compared with the
Ctrl group. However, following treatment with curcumin, the
densities of the positively stained areas in collagen-I and Sirius
red staining decreased significantly in the Alc + Cur group
compared with the Alc group. Moreover, stimulation with alcohol
induced a significant increase in the density of α-SMA-positive
staining, which suggests the activation of the HSCs in the Alc
group compared with the Ctrl group. The inhibitory effects of
curcumin against HSC activation were demonstrated by the marked
decrease in the density of α-SMA-positive staining in the Alc + Cur
group.
Curcumin inhibits HSC activation by
suppressing HSC proliferation and inducing apoptosis through the
enhancement of endoplasmic reticulum (ER) stress in vitro
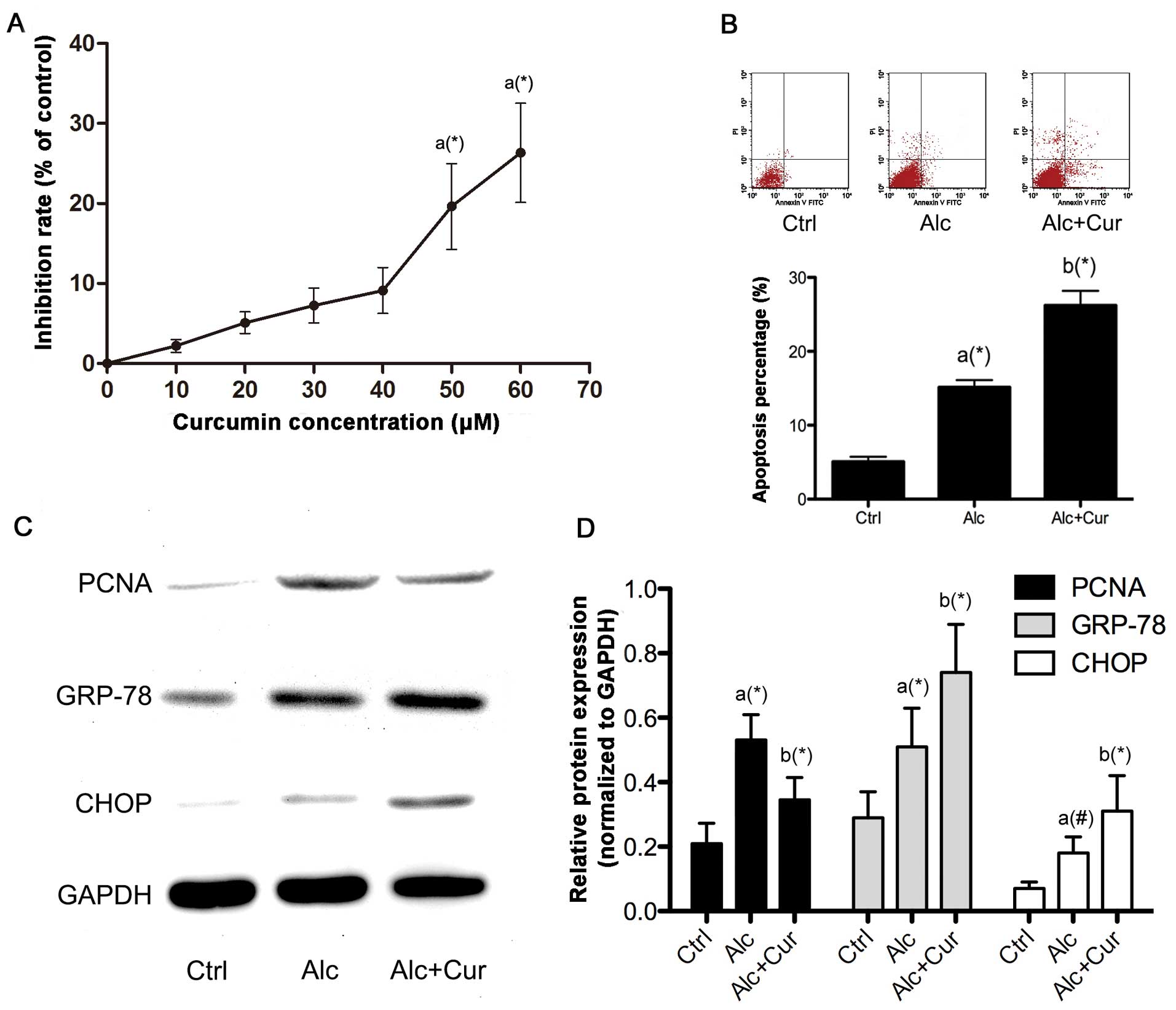
The proliferation of the HSCs was promoted by
incubation with alcohol as evidenced by the observation of the
increased expression of PCNA (Fig.
2). Following pre-treatment with serial concentrations of
curcumin, the proliferation of the alcohol-stimulated HSCs was
inhibited, as evidenced by the decrease in cell viability and the
decreased PCNA expression. The results from MTT cell viability
assay indicated that treatment with curcumin at a concentration of
50 μM markedly inhibited the proliferation of the
alcohol-stimulated HSCs. Furthermore, treatment with curcumin
markedly increased the apoptosis of the alcohol-stimulated HSCs
through ER stress signaling, which was evidenced by the elevated
apoptotic rate detected by flow cytometry and the increased
expression of GRP-78 and CHOP.
Curcumin deactivates HSCs and impaires
the expression of collagen-I and fibronectin in alcohol-stimulated
HSCs in vitro
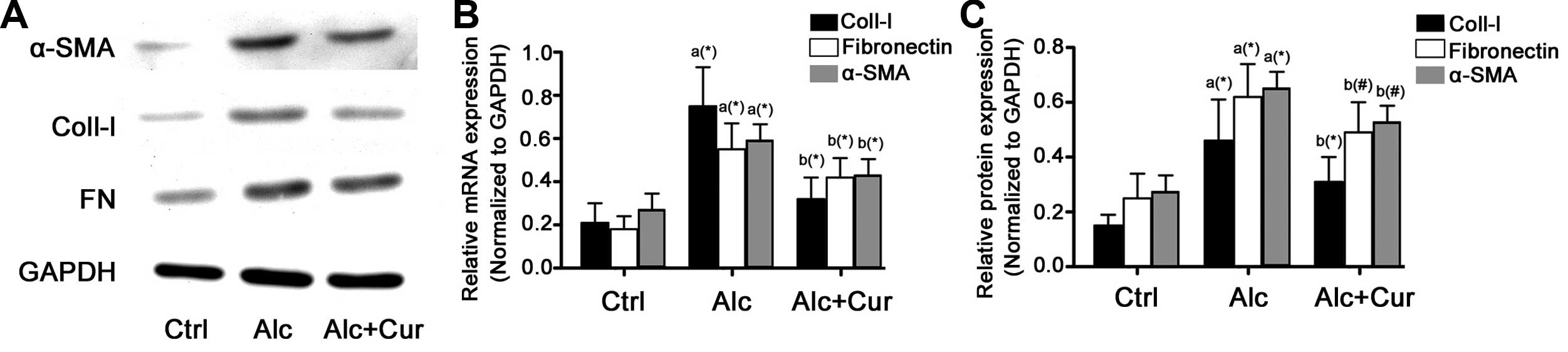
The expression of α-SMA, collagen-I and fibronectin
at both the transcriptional and translational levels markedly
increased following incubation of the cells with alcohol (Fig. 3). However, following the
administration of curcumin, the mRNA and protein expression of
α-SMA, collagen-I and fibronectin markedly decreased.
Curcumin deactivates the TGF-β signaling
pathway
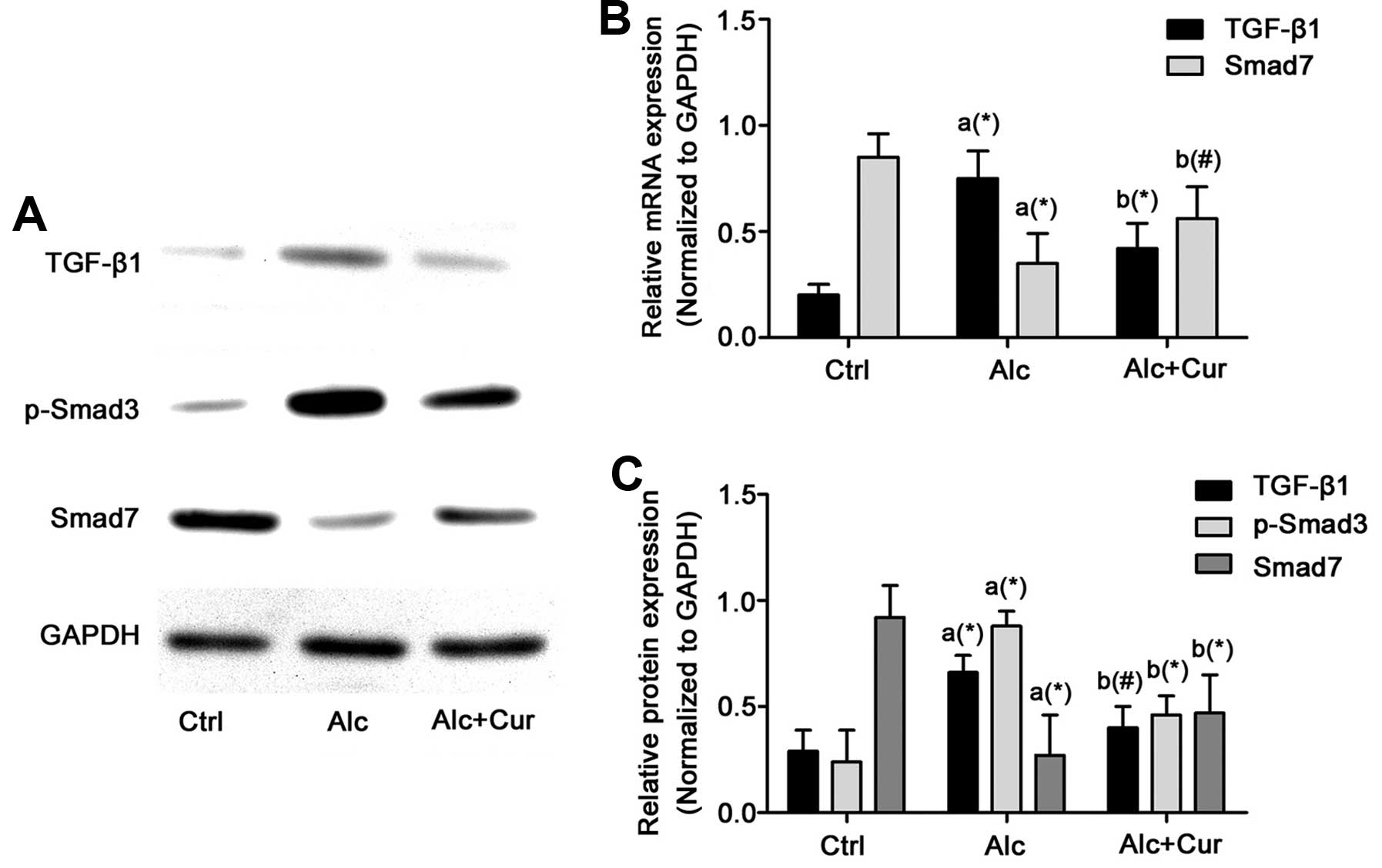
The protein expression of TGF-β and phosphorylated
Smad3 markedly increased, while that of Smad7 markedly decreased
following the administration of alcohol in the HSCs (Fig. 4). However, following treatment
with cucumin (Alc + Cur group), the TGF-β signaling pathway was
markedly inhibited, which was reflected by the increased expression
of Smad7 and the decreased expression of TGF-β and phosphorylated
Smad3 compared with the Alc group. Compared with the Alc group,
Smad7 expression increased, while that of TGF-β and phosphorylated
Smad3 markedly decreased in the isolated HSCs treated with
curcumin.
Discussion
As one of the hepatotoxic agents, alcohol abuse
results in liver damage, which eventually leads to progressive and
chronic hepatic fibrosis, a process through which excessive
collagen- rich ECM is generated to replace normal hepatic tissue
(15). Hepatic fibrosis
progresses and develops into hepatic cirrhosis, whose association
with hepatic failure, hepatocellular carcinoma and portal
hypertension has been confirmed (25). The progression of hepatic fibrosis
into cirrhosis may be reversible during the early stages (26), thus preventing chronic liver
disease.
Curcumin, also referred to as diferuloylmethane, is
extracted from the rhizome of Curcuma longa Linn. which as
long been used in Chinese and Indian traditional medicine (27). Curcumin has attracted the
attention of researchers due to its potential therapeutic effects
in the treatment of carcinomas, inflammation, asthma, diabetes and
hepatic failure (28). Studies
have reported that curcumin attenuates the development of liver
fibrosis in various animal models (29,30). In the present study, we
demonstrate the hepatoprotective effects of curcumin in a rat model
of hepatic fibrosis induced by the oral administration of alcohol.
We demonstrate that curcumin alleviates the alcohol-induced loss of
hepatic function. Furthermore, our results demonstrated that
curcumin reduced collagen generation in hepatic tissue, which was
stimulated following the administration of alcohol. These results
from our in vivo study suggest and confirm that curcumin
mitigates alcohol-induced hepatic fibrosis.
It is believed that HSCs are the major source of the
components of the ECM, including collagen, fibronectin and other
matrix proteins contributing to the genesis and development of
hepatic fibrosis (31). HSCs in a
quiescent state are activated by hepatic injury which is induced by
a number of agents, including alcohol. In such circumstances,
activated HSCs are characterized by proliferation, contractility
and fibrogenesis, which are marked by the increased expression of
α-SMA, according to both in vivo and in vitro studies
(32,33). In this study, the observation of
the decreased hepatic α-SMA expression following treatment with
curcumin in rats administered alcohol suggests that the
deactivation of HSCs plays an important role in the
hepatoprotective effects of curcumin against fibrosis.
Thus, the deactivation and inhibition of HSCs has
become a critical and potential therapeutic target in the treatment
of hepatic fibrosis. The ER is a cell organelle conducting protein
folding and handling calcium homeostasis (34). ER stress occurs in response to
harmful stimuli. Prolonged and overwhelming ER stress activates the
downstream apoptotic pathway through the activation of
pro-apoptotic molecules, such as CHOP (35). Our data revealed that the
proliferation of HSCs was stimulated following incubation of the
cells with alcohol, as evidenced by the increased expression of
PCNA and increased cell viability (MTT assay). We also demonstrated
that curcumin induced the apoptosis of the alcohol-stimulated HSCs,
which resulted in the inhibition of proliferation. The expression
of GRP-78, a molecular marker of ER stress (36), and CHOP increased significantly
following treatment with curcumin in the alcohol-stimulated HSCs.
These results suggest that curcumin inhibits the proliferation and
induces the apoptosis of alcohol-stimulated HSCs by activating ER
stress.
It is believed that TGF-β is among the most potent
cytokines that initiate the generation of myofibrobalsts (37). Studies have demonstrated that the
activation of the TGF-β/Smad signaling pathway is responsible for
the progression of hepatic fibrosis following liver injury
(38). TGF-β stimulates its
downstream Smad proteins through tansmembrane receptors, and the
transcriptions of several genes that encode ECM components, such as
collagen and fibronectin are then activated (39). In this study, an in vitro
experiment was implemented to investigate the effects of curcumin
on the TGF-β/Smad signaling pathway in an animal model of
alcohol-induced hepatic fibrosis. As expected, this signaling
pathway was activated following the administration of alcohol, but
was then inhibited following treatment with curcumin. Furthermore,
TGF-β/Smad signaling seems indispensible in transforming HSCs from
a quiescent state to an active one (40). TGF-β induces the phosphorylatoin
and nuclear translocation of Smad2 and Smad3 in HSCs (41). Additionally, it has been
demonstrated that the activation of Smad3 is more important than
that of Smad2 in the morphological and functional maturation of
fibrogenic cells (39). It has
been suggested that Smad7, another Smad protein, acts as an
inhibitor of TGF-β/Smad signaling by abrogating the effects of
TGF-β1 (42). In this study, the
downregulation of Smad7 with the upregulation of TGF-β1 and
phosphorylated Smad3 was observed in the alcohol-stimulated HSCs,
suggesting that the HSCs were activated by alcohol. By contrast,
following the administration of curcumin, the TGF-β/Smad signaling
pathway was inhibited in the alcohol-stimulated HSCs, as
characterized by the upregulation of Smad7 and the downregulation
of TGF-β1 and phosphorylated Smad3 expression, resulting in the
impairment of collagen-I and fibronectin synthesis and secretion in
the activated HSCs.
In conlcusion, the data from the present study
indicate that curcumin exerts hepatoprotective effects against the
development of alcoloho-induced hepatic fribosis by inhibiting the
proliferation and promoting the apoptosis of HSCs. These effects
are mediated through the activation of ER stress and the
suppression of the TGF-β/Smad signaling pathway.
References
|
1
|
Williams R: Global challenges in liver
disease. Hepatology. 44:521–526. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mandayam S, Jamal MM and Morgan TR:
Epidemiology of alcoholic liver disease. Semin Liver Dis.
24:217–232. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Poli G: Pathogenesis of liver fibrosis:
role of oxidative stress. Mol Aspects Med. 21:49–98. 2000.
View Article : Google Scholar
|
|
4
|
Brenner DA: Molecular pathogenesis of
liver fibrosis. Trans Am Clin Climatol Assoc. 120:361–368.
2009.PubMed/NCBI
|
|
5
|
Senoo H: Structure and function of hepatic
stellate cells. Med Electron Microsc. 37:3–15. 2004. View Article : Google Scholar
|
|
6
|
Gao B and Bataller R: Alcoholic liver
disease: pathogenesis and new therapeutic targets.
Gastroenterology. 141:1572–1585. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Siegmund SV, Dooley S and Brenner DA:
Molecular mechanisms of alcohol-induced hepatic fibrosis. Dig Dis.
23:264–274. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Dooley S, Streckert M, Delvoux B and
Gressner AM: Expression of Smads during in vitro
transdifferentiation of hepatic stellate cells to myofibroblasts.
Biochem Biophys Res Commun. 283:554–562. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Derynck R and Zhang YE: Smad-dependent and
Smad-independent pathways in TGF-beta family signalling. Nature.
425:577–584. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Shi Y and Massague J: Mechanisms of
TGF-beta signaling from cell membrane to the nucleus. Cell.
113:685–700. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kavsak P, Rasmussen RK, Causing CG, et al:
Smad7 binds to Smurf2 to form an E3 ubiquitin ligase that targets
the TGF beta receptor for degradation. Mol Cell. 6:1365–1375. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen K, Zhang S, Ji Y, et al: Baicalein
inhibits the invasion and metastatic capabilities of hepatocellular
carcinoma cells via down-regulation of the ERK pathway. PLoS One.
8:e729272013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Wu L, Cai B, Zheng S, Liu X, Cai H and Li
H: Effect of emodin on endoplasmic reticulum stress in rats with
severe acute pancreatitis. Inflammation. 36:1020–1029. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Geng J, Peng W, Huang Y, Fan H and Li S:
Ginsenoside-Rg1 from Panax notoginseng prevents hepatic fibrosis
induced by thioacetamide in rats. Eur J Pharmacol. 634:162–169.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
O’Connell MA and Rushworth SA: Curcumin:
potential for hepatic fibrosis therapy? Br J Pharmacol.
153:403–405. 2008.
|
|
16
|
Noorafshan A and Ashkani-Esfahani S: A
review of therapeutic effects of curcumin. Curr Pharm Des.
19:2032–2046. 2013.PubMed/NCBI
|
|
17
|
Tu CT, Yao QY, Xu BL, Wang JY, Zhou CH and
Zhang SC: Protective effects of curcumin against hepatic fibrosis
induced by carbon tetrachloride: modulation of high-mobility group
box 1, Toll-like receptor 4 and 2 expression. Food Chem Toxicol.
50:3343–3351. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yao QY, Xu BL, Wang JY, Liu HC, Zhang SC
and Tu CT: Inhibition by curcumin of multiple sites of the
transforming growth factor-beta1 signalling pathway ameliorates the
progression of liver fibrosis induced by carbon tetrachloride in
rats. BMC Complement Altern Med. 12:1562012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim JM, Kim HG, Han JM, et al: The herbal
formula CGX ameliorates the expression of vascular endothelial
growth factor in alcoholic liver fibrosis. J Ethnopharmacol.
150:892–900. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Bruck R, Ashkenazi M, Weiss S, et al:
Prevention of liver cirrhosis in rats by curcumin. Liver Int.
27:373–383. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Wang Y, Cheng M, Zhang B, Nie F and Jiang
H: Dietary supplementation of blueberry juice enhances hepatic
expression of metallothionein and attenuates liver fibrosis in
rats. PLoS One. 8:e586592013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Traber PG, Chou H, Zomer E, et al:
Regression of fibrosis and reversal of cirrhosis in rats by
galectin inhibitors in thioacetamide-induced liver disease. PLoS
One. 8:e753612013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Su LJ, Chang CC, Yang CH, et al:
Graptopetalum paraguayense ameliorates chemical-induced rat hepatic
fibrosis in vivo and inactivates stellate cells and Kupffer cells
in vitro. PLoS One. 8:e539882013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu ZW, Zhu HT, Chen KL, et al: Protein
kinase RNA-like endoplasmic reticulum kinase (PERK) signaling
pathway plays a major role in reactive oxygen species
(ROS)-mediated endoplasmic reticulum stress-induced apoptosis in
diabetic cardiomyopathy. Cardiovasc Diabetol. 12:1582013.
View Article : Google Scholar
|
|
25
|
Bataller R and Brenner DA: Liver fibrosis.
J Clin Invest. 115:209–218. 2005. View
Article : Google Scholar
|
|
26
|
Lee UE and Friedman SL: Mechanisms of
hepatic fibrogenesis. Best Pract Res Clin Gastroenterol.
25:195–206. 2011. View Article : Google Scholar
|
|
27
|
Padhye S, Chavan D, Pandey S, Deshpande J,
Swamy KV and Sarkar FH: Perspectives on chemopreventive and
therapeutic potential of curcumin analogs in medicinal chemistry.
Mini Rev Med Chem. 10:372–387. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sharma RA, Gescher AJ and Steward WP:
Curcumin: the story so far. Eur J Cancer. 41:1955–1968. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fu Y, Zheng S, Lin J, Ryerse J and Chen A:
Curcumin protects the rat liver from CCl4-caused injury and
fibrogenesis by attenuating oxidative stress and suppressing
inflammation. Mol Pharmacol. 73:399–409. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Rivera-Espinoza Y and Muriel P:
Pharmacological actions of curcumin in liver diseases or damage.
Liver Int. 29:1457–1466. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Sun X, He Y, Ma TT, Huang C, Zhang L and
Li J: Participation of miR-200a in TGF-β1-mediated hepatic stellate
cell activation. Mol Cell Biochem. 388:11–23. 2014.
|
|
32
|
Kodama T, Takehara T, Hikita H, et al:
Increases in p53 expression induce CTGF synthesis by mouse and
human hepatocytes and result in liver fibrosis in mice. J Clin
Invest. 121:3343–3356. 2011. View
Article : Google Scholar
|
|
33
|
Friedman SL: Evolving challenges in
hepatic fibrosis. Nat Rev Gastroenterol Hepatol. 7:425–436. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ron D and Walter P: Signal integration in
the endoplasmic reticulum unfolded protein response. Nat Rev Mol
Cell Biol. 8:519–529. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
35
|
Rao RV, Ellerby HM and Bredesen DE:
Coupling endoplasmic reticulum stress to the cell death program.
Cell Death Differ. 11:372–380. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Lee AS: The ER chaperone and signaling
regulator GRP78/BiP as a monitor of endoplasmic reticulum stress.
Methods. 35:373–381. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Roberts AB, Tian F, Byfield SD, et al:
Smad3 is key to TGF-beta-mediated epithelial-to-mesenchymal
transition, fibrosis, tumor suppression and metastasis. Cytokine
Growth Factor Rev. 17:19–27. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Leask A and Abraham DJ: TGF-beta signaling
and the fibrotic response. FASEB J. 18:816–827. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bissell DM, Roulot D and George J:
Transforming growth factor beta and the liver. Hepatology.
34:859–867. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Schnabl B, Kweon YO, Frederick JP, Wang
XF, Rippe RA and Brenner DA: The role of Smad3 in mediating mouse
hepatic stellate cell activation. Hepatology. 34:89–100. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Liu C, Gaca MD, Swenson ES, Vellucci VF,
Reiss M and Wells RG: Smads 2 and 3 are differentially activated by
transforming growth factor-beta (TGF-beta) in quiescent and
activated hepatic stellate cells. Constitutive nuclear localization
of Smads in activated cells is TGF-beta-independent. J Biol Chem.
278:11721–11728. 2003. View Article : Google Scholar
|
|
42
|
Friedman SL: Mechanisms of hepatic
fibrogenesis. Gastroenterology. 134:1655–1669. 2008. View Article : Google Scholar : PubMed/NCBI
|