Introduction
ErbB2 is a member of the epidermal growth factor
receptor (EGFR, also known as ErbB) family of receptor tyrosine
kinases. There are 4 members in the ErbB family, including ErbB1-4.
Endogenous ligands and the homo- or hetero-dimerization of ErbB
receptors stimulate its tyrosine kinase activity, and regulate
cellular proliferation and survival (1–3).
Unlike other members of the ErbB family, ErbB2 is an orphan
receptor without an endogenous ligand. However, the overexpression
of ErbB2 is clinically associated with approximately 30% of ovarian
cancers, breast cancers (4), and
has been shown to correlate with the metastasis, therapeutic
resistance and poor prognosis of cancer (5–7).
In a phage display study, EC1, an artificial peptide, was found to
bind the extracellular domain of ErbB2 in living cells and
fresh-frozen human breast cancer specimens (8). Moreover, biotin-conjugated EC1 and
the recombinant protein, EC1-eGFP, retained the affinity for ErbB2,
and were selectively internalized into ErbB2-overexpressing cancer
cells (8–10). Recently, divalent and multivalent
forms of the EC1-Fc ligand in liposomes have been reported to
improve the affinity for ErbB2 and enhance internalization
(11). Thus, the EC1 peptide is a
potential artificial ligand for targeting ErbB2.
On the other hand, bioluminescence has been
extensively used as a relatively simple, cost-effective and
extremely sensitive imaging system in intact cells and living
animals (12). The most common
luciferases for bioluminescence imaging include firefly luciferase
(FLuc), Renilla luciferase (RLuc) and Gaussia
luciferase (GLuc). Each luciferase has distinct properties in the
application of bioluminescence imaging. FLuc (62 kDa) catalyzes the
oxidation of luciferin to yield bioluminescence in the presence of
O2, magnesium and adenosine triphosphate (ATP) (13). RLuc (36 kDa) and GLuc (19.9 kDa)
catalyze the oxidative decarboxylation of coelenterazine (CTZ) to
emit light independent of ATP. However, RLuc has a lower quantum
yield than FLuc, and also less enzymatic efficiency (14,15). GLuc yields approximately 200-
(in vivo) to 1,000-fold (in vitro) more
bioluminescence in mammalian cells than FLuc and RLuc (16). The small molecular size (19.9
kDa), independence from ATP and strong emission render GLuc much
more suitable for bioluminescence imaging (17,18).
Liposomes are mainly composed of phospholipids and
cholesterols which are also the natural components in cells
(19). The hydrophobic
interactions lead to the self-assembly of lipid bilayers in water
and form phospholipid vesicles (20). The concentric bilayers can be used
to incorporate water-soluble materials, such as chemical compounds,
proteins and siRNA in the aqueous core of the liposome (21–23). The addition of a conjugate of
polyethylene glycol (PEG) linked to a lipid anchor to the liposomal
formulation significantly prolongs the liposome circulation time,
which greatly benefits the application of liposomes in vivo
(24). Liposomes are nanoscale in
size; thus, they are also considered as nanoscale drug delivery
systems (DDS) or nanocapsules (25). In recent years, multifunctional
liposomes have been developed to meet the requirements of different
DDS by altering lipid composition, size and lipid surface
modification (25,26). In our previous studies, the EGFR
antibody-conjugated liposomes were constructed for the selective
delivery of sodium borocaptate and bioluminescence imaging in
EGFR-overexpressing glioma (18,22). In the present study, the
EC1-GLuc-liposome was constructed to combine bioluminescence
imaging and targeted DDS for ErbB2-overexpressing metastatic
ovarian carcinoma.
Ovarian carcinoma is one of the most common causes
of mortality from gynaecological malignancies. Ovarian carcinoma is
highly metastatic, and patients mainly succumb to the disease due
to metastasis (27). Unlike other
solid tumors, the metastasis of ovarian carcinoma is mostly induced
by direct dissemination into the peritoneal cavity, rather than
circulatory or lymphatic metastasis (28). Due to the complexity of the
peritoneal cavity, it is difficult to clean-up the metastatic foci
using currently available therapeutic strategies, such as surgery,
radiotherapy and chemotherapy. Therefore, novel therapeutic
strategies involving targeted molecular imaging and DDS are
urgently required in order to improve the therapeutic efficacy in
metastatic ovarian carcinoma. In this study, a novel
ErbB2-targeting bioluminescence protein was generated by fusing EC1
with GLuc, and was conjugated to a liposome to construct the
EC1-GLuc-liposome. MCF7 (ErbB2-negative) and SKOv3
(ErbB2-overexpressing) cells were employed to evaluate the efficacy
of the EC1-GLuc-liposome in ErbB2-targeted imaging and drug
delivery. It was found that the ErbB2-targeted bioluminescence
imaging and selective delivery of HPTS by the EC1-GLuc-liposome was
effective not only in the SKOv3 cells in vitro, but also in
metastatic ovarian tumors in vivo. Thus, the present study
suggests the EC1-GLuc-liposome to be a novel strategy for combining
bioluminescence imaging and drug delivery for monitoring and
treating ErbB2-overexpressing metastatic ovarian carcinoma.
Materials and methods
Cell culture and detection of ErbB2
expression
The MCF7 and SKOv3 cell lines were obtained from the
American Type Culture Collection (ATCC, Manassas, VA, USA). The 2
cell lines were cultured in Dulbecco’s modified Eagle’s medium
(DMEM) medium supplemented with 10% fetal bovine serum (FBS) and
100 μg/ml penicillin-streptomycin (P/S). The medium, FBS and P/S
were from Invitrogen (Carlsbad, CA, USA). All cultures were
maintained in a humidified incubator at 37°C with an atmosphere
containing 5% CO2.
The expression of ErbB2 in the MCF7 and SKOv3 cells
was examined by immunoblot analsyis. Briefly, cells on 35 mm dishes
were washed once with phosphate-buffered saline (PBS), and scraped
in 1X SDS sample buffer. Cell lysates were subjected to 6%
SDS-PAGE, and immunoblotted with rabbit anti-ErbB2 antibody
(1:1,000; Cell Signaling Technology, Danvers, MA, USA) overnight at
4°C. HRP-conjugated secondary antibody (Sigma-Aldrich, St. Louis,
MO, USA) was used at 1:2,000. Immunoblotting signals were detected
using the VersaDoc 5000 Imaging system (Bio-Rad, Hercules, CA, USA)
with an enhanced chemiluminescence detection kit (Amersham
Biosciences, Pittsburgh, PA, USA).
Plasmid construction
The pGLuc plasmid was purchased from LUX
Biotechnology Ltd. (Scotland, UK). The GLuc gene was amplified from
the plasmid using a sense primer (5′-GGATCCG
AAACCAACTGAAAACAATGAAG-3′) and an antisense primer
(5′-GTCGACATCACCACCGGCACCCTTTAT-3′). The PCR product was ligated
into the pCR2.1 TOPO vector (Invitrogen) for amplification, and
recloned in the pET52b(+) vector (Invitrogen) to construct
GLuc-pET52b(+). The following sense and antisense oligonucleotides
with appropriate resistance enzyme sites for EC1 peptide were
prepared: 5′-GGGTGGAC
TGGCTGGTGCCTGAATCCAGAAGAATCTACTTGGGG
ATTCTGTACTGGATCTTTCGGTGGAGGTAGTTCAG-3′ (sense, underline indicates
SamI and BamHI sites), 5′-GATCCT
GAACTACCTCCACCGAAAGATCCAGTACAGAATCCCC
AAGTAGATTCTTCTGGATTCAGGCACCAGCCAGTCCA CCC-3′ (antisense, underline
indicates BamHI and SamI sites). The oligonucleotides
for EC1 were modified by phosphorylation at 5′. The sense and
antisense oligonucleotides (10 pmol each) were co-incubated at 95°C
for 10 min, and annealed at room temperature to form
double-stranded DNA. The double-stranded DNA for EC1 was ligated to
GLuc-pET52b treated with the appropriate resistance enzymes to
construct EC1-GLuc-pET52b. The sequences of the constructed
plasmids were confirmed using an ABI 3100 sequencer (Applied
Biosystems, Bedford. MA, USA).
Expression and purification of
recombinant proteins
Recombinant protein expression and purification was
carried out as previously described (10). Briefly, E. coli BL21 (DE3)
transformed with the plasmid for GLuc or EC1-GLuc was grown in a LB
medium containing 100 μg/ml ampicillin at 37°C. When the
OD600 reached 0.6, the expression of the recombinant
proteins was induced by the addition of 0.2 mM isopropyl
1-thio-β-D-galactopyranoside at 25°C overnight. The
GLuc-6His and EC1-GLuc-6His proteins were purified from the
supernatant using ProBond Nickel-chelating resin (Invitrogen). The
recombinant proteins were subjected to 15% SDS-PAGE, and the
expression was confirmed by Coomassie brilliant blue staining and
western blot analysis with rabbit anti-GLuc serum (1:1,000;
Nanolight Technology, Pinetop, AZ, USA). The proteins were finally
dialyzed against PBS (pH 7.4) at 4°C for 24 h, and the
concentrations were determined using a Biadford protein assay kit
(Bio-Rad). Aliquots of protein were stored at −80°C prior to
use.
Construction of GLuc-liposome and
EC1-GLuc-liposome
Liposomes composed of
DOPC:DOPG:DOGS-NTA-Ni:CH:DSPE-PEG2000 (molar ratio,
3:3:1:4:0.1) were prepared as previously described with a slight
modification (18,22). Briefly, 100 μM of lipid were
dissolved in 2 ml of a chloroform/diethyl ether mixture (1:1 v/v).
The ratio of organic to aqueous phase was 2:1. The mixture was
added to a rotary evaporator to form a lipid gel at 50°C under
reduced pressure. Subsequently, 2 ml of 35 mM HPTS in PBS was mixed
with the lipid gel. The mixture was vortexed and sonicated. To
obtain liposomes with homogeneous size, the liposome emulsion was
extruded through a polycarbonate membrane 100 nm in pore size using
an extruder device at 60°C. The mean diameter and zeta potential of
the prepared liposomes were determined using an electrophoretic
light scattering spectrophotometer (ELS-8000; Photal, Osaka,
Japan). The unencapsulated free HPTS was removed using a PD-10
desalting column (Amersham Biosciences). To construct
luciferase-conjugated liposomes, the nickel-chelated liposome was
incubated with GLuc-His or EC1-GLuc-His at a molar ratio of 20:1
overnight at room temperature at a low rotating speed. Free
GLuc-His or EC1-GLuc-His was removed with a sepharose CL-4B column
and the eluted GLuc or EC1-GLuc conjugated liposomes were
concentrated. The lipid in each fraction was analyzed by the DAOS
method using a Phospholipid C reagent kit (Wako Pure Chemical
Industiries, Ltd., Osaka, Japan). The recombinant protein
conjugated on the liposome in each fraction was detected by western
blot analayis with rabbit anti-GLuc serum or a Biadford protein
assay kit.
Preparation of CTZ
The substrate for GLuc was prepared as previously
described (10,12). Briefly, CTZ (Nanolight Technology)
was dissolved in acidified methanol [1 drop of concentrated HCl
(12.4 N) in 10 ml of methanol] to a concentration of 5 mg/ml.
Aliquots of 100 μl were stored at −80°C. For luminescence imaging,
the aliquots of CTZ (5 mg/ml) were diluted with PBS (containing 5
mM NaCl, pH 7.2) for greater light output and more stability.
Bioluminescence imaging and selective
delivery of HPTS into ErbB2-overexpressing cells in vitro by the
EC1-GLuc-liposome
For bioluminescence imaging in vitro, the
MCF7 and SKOv3 cells were cultured on 35-mm glass-bottomed dishes
for 24–48 h. To improve the adherence of MCF7 and SKOv3 cells, all
glass-bottomed dishes were pre-coated with laminin (Roche
Diagnostics, Indianapolis, IN, USA). The MCF7 and SKOv3 cells were
incubated with 1 mM GLuc-liposome or EC1-GLuc-liposome. After 2 h
of incubation, the cells were washed 3 times with DMEM.
Bioluminescence was detected with an Olympus Luminoview LV 200
bioluminescence microscope (Olympus, Tokyo, Japan) immediately
after the addition of 1 μg/ml CTZ to the serum-free DMEM. Images
were acquired and analyzed using Metamorph software (Molecular
Devices, Sunnyvale, CA, USA).
To examine the internalization of the
luciferase-conjugated liposomes in cells, the MCF7 and SKOv3 cells
were incubated with 1 mM of the liposomes. In a blocking
experiment, the SKOv3 cells were treated with anti-ErbB2 antibody
against the extracellular domain of ErbB2 (chicken, 1.0 μg/ml;
Abcam, Cambridge, MA, USA) 1 h prior to incubation with
EC1-GLuc-liposome. After 2 h of incubation, the cells were washed
with PBS 3 times and trypsinized. For western blot analysis, cell
lysate was prepared and subjected to 15% SDS-PAGE. The
internalization of the luciferase-conjugated liposomes was analyzed
by immunoblotting with rabbit anti-GLuc serum. β-actin was used as
an endogenous control. To detect the delivery of HPTS, the cells
were fixed with 4% paraformaldehyde (PFA) for 10 min, and washed
with PBS twice. The fluorescence signal of HPTS in the cells was
observed using a confocal laser microscope (Olympus).
Preparation of tumor-bearing mice
All animal handling was performed in accordance with
the Animal Research Committee guidelines of Nanchang University,
Nanchang, China. The cultured SKOv3 cells were washed twice with
PBS, typsinized and harvested by centrifugation. The SKOv3 cells
(5.0×106) suspended in DMEM were intraperitoneally
implanted in anesthetized nude mice (Balb/c Slc-nu/nu, female, 6–8
weeks old) under aseptic conditions. After cell implantation, the
mice with tumors which had developed for 3 weeks were used for
in vivo experiments. During tumor development, breath and
behavioral activity were monitored twice each day to evaluate pain
in the mice. The experiments were immediately terminated if the
mice presented with significant pain symptoms. All mice were
euthanized by an intraperitoneal injection of 15 mg/100 g
pentobarbital solution after the experiments.
The expression of ErbB2 in the SKOv3 tumor
xenografts was confirmed by immunofluorescence (IF) staining.
Paraffin-embedded slices of tumors were prepared at a thickness of
5 μm. First, histological observations of the xenografted tumors
were made using hematoxylin and eosin (H&E) staining. IF
staining was performed with anti-ErbB2 antibody (1:50; Cell
Signaling Technology). The secondary antibody was Cy3-conjugated
anti-rabbit IgG (1:100; Invitrogen, Molecular Probes). The nucleus
was counterstained with 0.1 μg/ml of Hoechst 33248 (Sigma-Aldrich)
for 5 min. Fluorescence signals were observed using a confocal
laser microscope (FV500; Olympus).
Bioluminescence imaging and delivery of
HPTS into metastatic SKOv3 tumors in vivo by the
EC1-GLuc-liposome
The mice bearing metastatic SKOv3 tumors were
administrated intravenously 300 μl of luciferase-conjugated
liposomes (40 mM) via the tail vein. After 8 h, the mice were
imaged under anesthesia by intraperitoneally injecting 200 μl of
CTZ solution (5 mg/kg) using a cooled CCD camera (IVIS; Xenogen,
Alameda, CA, USA) as previously described (18). In addition, a laparotomy was
performed to directly image the SKOv3 tumors using the cooled CCD
camera. The bioluminescence intensity of the selected region over
the tumor was recorded as maximum photons
sec/cm2/steradian. To detect the delivery of HPTS in
vivo by the EC1-GLuc-liposome, the mice were euthanized by an
intraperitoneal injection of 15 mg/100 g pentobarbital solution
soon after bioluminescence imaging. The metastatic SKOv3 tumors
were removed for preparation of 10-μm tissue section. The
fluorescence signal of HPTS was observed using a confocal laser
microscope (Olympus).
Results
Construction and characterization of the
EC1-GLuc-liposome
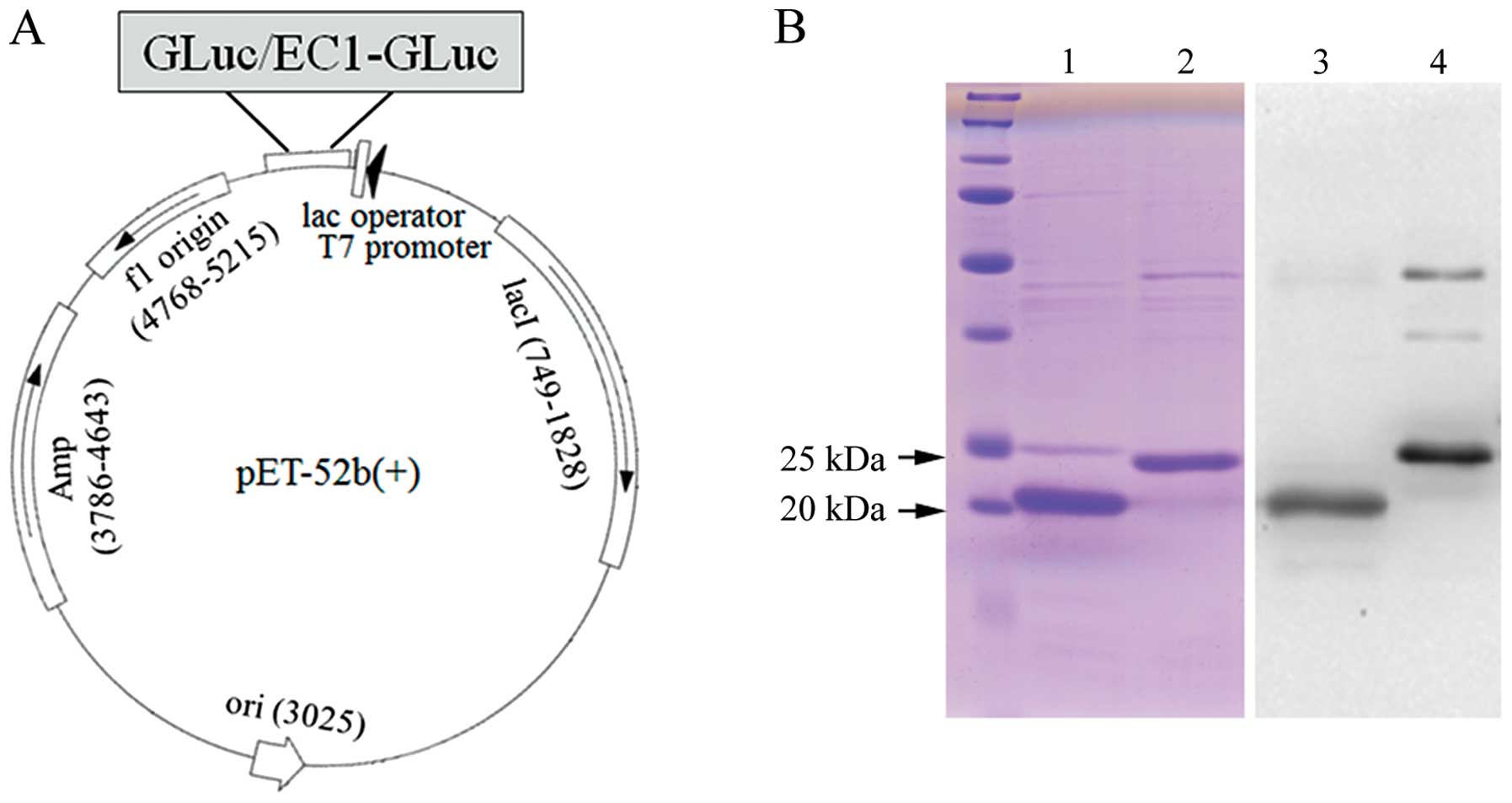
Firstly, GLuc and EC1-GLuc were subcloned into the
pET-52b(+) vector for the expression of recombinant proteins
(Fig. 1A). The recombinant
GLuc-His and EC1-GLuc-His were expressed in E. coli BL21 and
purified using a column of ProBond Nickel-chelating resin.
Following purification, the recombinant proteins were subjected to
15% SDS-PAGE and confirmed with Coomassie brilliant blue staining
and western blot analysis (Fig.
1B). The results reveaked that GLuc-His and EC1-GLuc-His were
highly purified. The molecular sizes of GLuc-His and EC1-GLuc-His
were approximately 20 and 23 kDa, respectively. On the other hand,
the molar ratio of DOPC:DOPG:DOGS-NTA-Ni:CH:DSPE-PEG2000
in 3:3:1:4:0.1 was used as the nickel-liposome formulation
(Table I). A cell-impermeable
fluorescence dye (HPTS) was encapsulated in the liposomes as
previously described (18). The
recombinant proteins were conjugated to the liposomes through their
affinity with nickel to construct the GLuc-liposome and
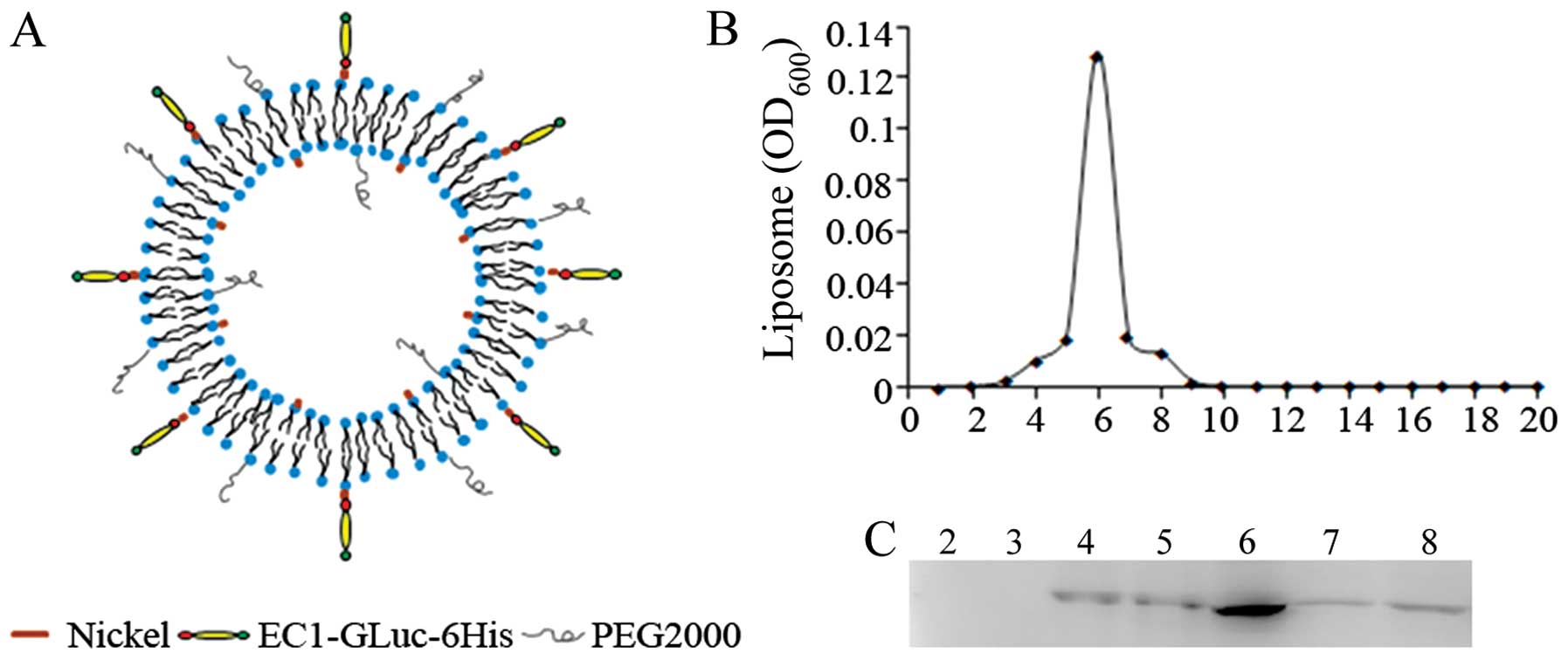
EC1-GLuc-liposome (Fig. 2A). The
GLuc-liposome was used as the control in the experiments. After
construction of the luciferase-conjugated liposomes, the diameter
and zeta potential of the liposomes were measured (Table I). The diameter of the liposomes
was 106.37 to 117.58 nm. The zeta potential was −30.46 to −27.52
mV. To examine the conjugation of EC1-GLuc-His to the
nickel-liposome, the positions of the recombinant protein and
liposome were compared after their separation with Sepharose CL-4B.
The liposome mostly appeared in fraction 6 (Fig. 2B). Western blot analysis indicated
that EC1-GLuc-His was also most abundant in fraction 6 (Fig. 2C). These results suggest that
EC1-GLuc-His was effectively conjugated to the surface of the
liposome through its affinity with nickel.
 | Table IComposition and properties of
liposomes. |
Table I
Composition and properties of
liposomes.
| Lipid composition
(mmol) | DOPC | 30 |
| DOPG | 30 |
| Chol | 40 |
| DOGS-NTA-Ni | 10 |
|
DSPE-PEG2000 | 1 |
| Total (mmol) | 111 |
| Lipid
properties | zeta potential
(mV) | −30.46 to
27.52 |
| Size (nm) | 106.37 to
117.58 |
ErbB2-targeting and delivery of HPTS by
the EC1-GLuc-liposome in vitro
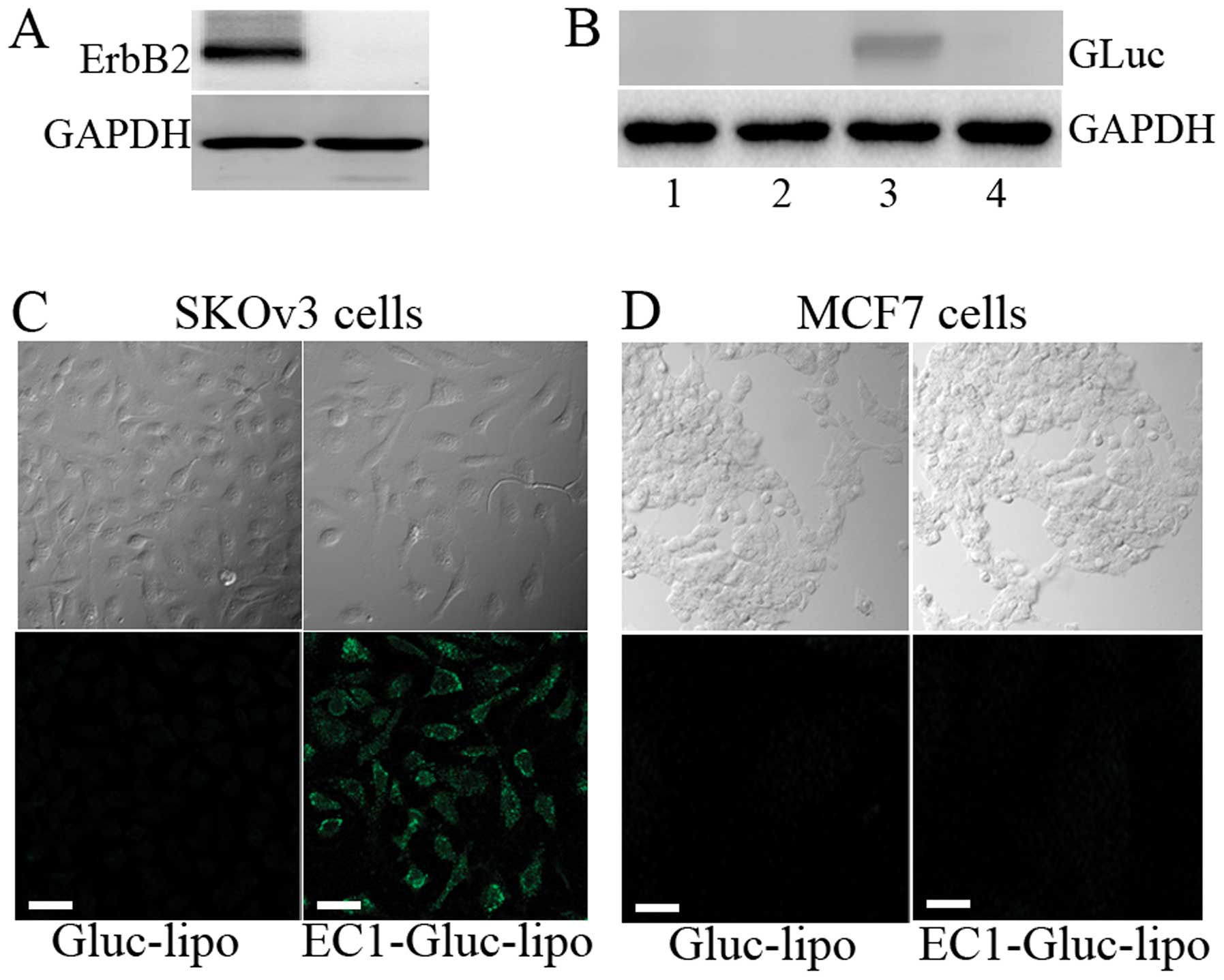
Two human carcinoma cell lines, MCF7 and SKOv3, were
used for the in vitro experiments. Firstly, the expression
level of ErbB2 in the 2 cell lines was examined by western blot
analysis. The overexpression of ErbB2 was observed in the SKOv3
cells, while the expression of ErbB2 was undetectable in the MCF7
cells (Fig. 3A). Moreover,
western blot analysis revealed that the internalization of the
recombinant protein was only observed in the EC1-GLuc-liposome
treated SKOv3 cells. The internalization was mostly undetectable
when the extracellular domain of ErbB2 was blocked by anti-ErbB2
antibody (Fig. 3B). The selective
delivery of HPTS was also observed in the SKOv3 cells treated with
the EC1-GLuc-liposome, but not in the MCF7 cells (Fig. 3C). These results suggest that the
EC1-GLuc-liposome retains its ErbB2-targeting and is effective for
DDS in vitro.
The EC1-GLuc-liposome targets ErbB2 for
bioluminescence imaging in vitro
To detect the ErbB2-targeted bioluminescence, the
MCF7 and SKOv3 cells were incubated with 1 mM GLuc-liposome or
EC1-GLuc-liposome. After 2 h of incubation, the cells were washed 3
times with DMEM. The bioluminescent signal was detected using an
Olympus Luminoview LV 200 bioluminescence microscope immediately
after the addition of 1 μg/ml CTZ in serum-free DMEM. A strong
signal was detected in ErbB2-overexpressing SKOv3 cells incubated
with the EC1-GLuc-liposome (Fig.
4). By contrast, no significant bioluminescence was observed in
the SKOv3 cells incubated with the GLuc-liposome and the MCF7 cells
incubated with both liposomes. This indicates that the
EC1-GLuc-liposome effectively targets ErbB2-overexpressing cells
for bioluminescence imaging in vitro.
Bioluminescence imaging and delivery of
HPTS into metastatic ovarian carcinoma tumors by the
EC1-GLuc-liposome in vivo
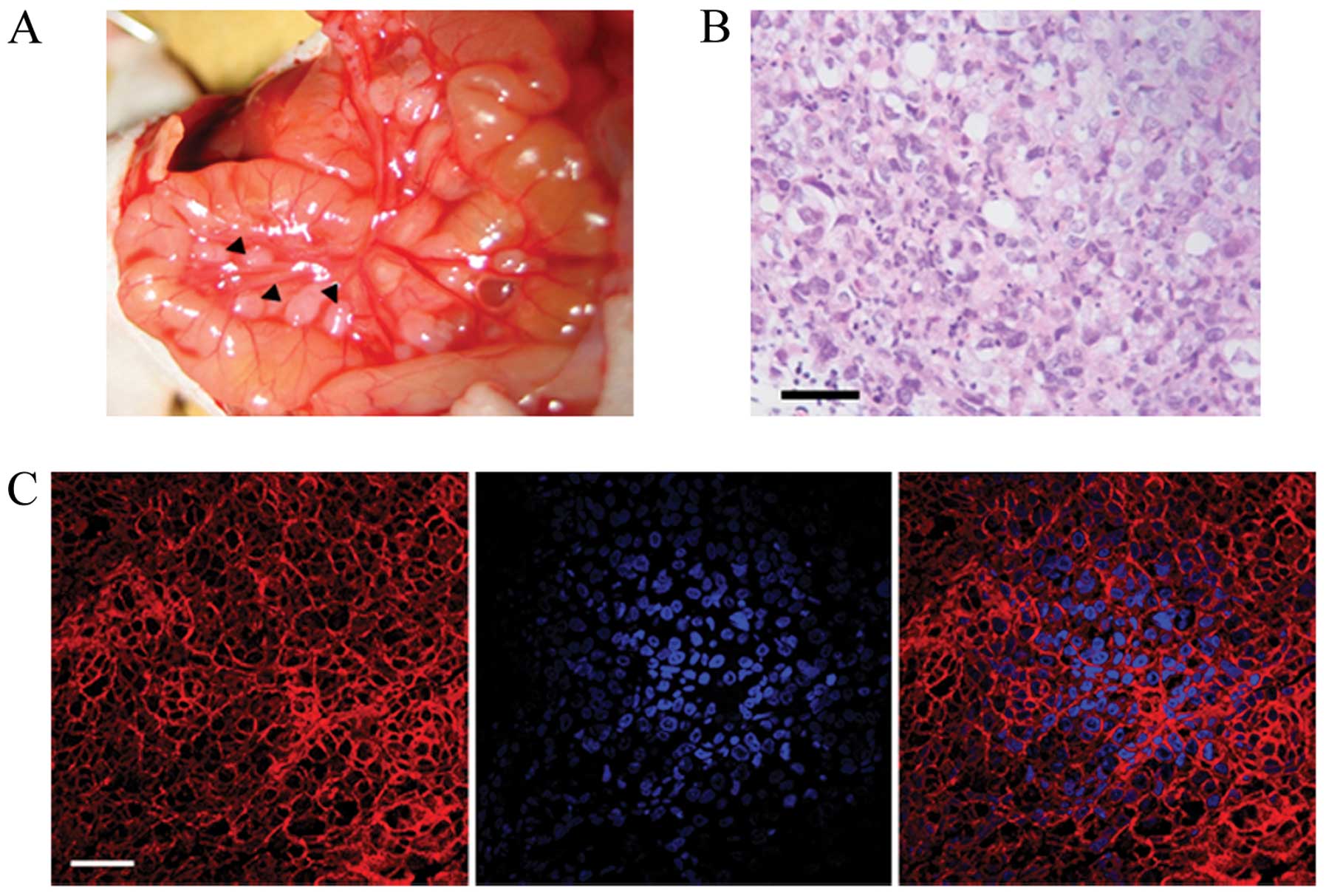
For in vivo experiments, we established
metastatic ovarian carcinoma tumor-bearing mice by the
intraperitoneal implantation of 5×106 SKOv3 cells.
Following tumor development for 3 weeks, the metastatic ovarian
tumors were clearly visible by laparotomy (Fig. 5A). Histological observations of
the tumors were carried out using H&E staining (Fig. 5B). In addition, the expression of
ErbB2 in the metastatic SKOv3 tumors was also confirmed by IF
staining (Fig. 5C). To evaluate
the ErbB2-targeted imaging of the EC1-GLuc-liposome in vivo,
metastatic SKOv3 tumor-bearing mice were injected with the
GLuc-liposome or EC1-GLuc-liposome via the tail vein. Eight hours
later, bioluminescence images were acquired after the
administration of CTZ. A strong bioluminescent signal was detected
in the mice injected with the EC1-GLuc-liposome, while it was
undetectable in the mice injected with the GLuc-liposome (Fig. 6A). Moreover, direct
bioluminescence imaging was achieved after laparotomy in the
metastatic SKOv3 tumor-bearing mice injected with the
EC1-GLuc-liposome (Fig. 6B). To
investigate the selective delivery of HPTS, the metastatic SKOv3
tumors were removed for section soon after bioluminescence imaging.
A strong fluorescence signal of HPTS was observed in the
EC1-GLuc-liposome-treated mice (Fig.
6C). However, in the GLuc-liposome-treated mice, no significant
fluorescence was detected. These results suggest that the
EC1-GLuc-liposome is effective for ErbB2-targeted bioluminescence
imaging and targeted DDS for metastatic ovarian carcinoma in
vivo.
Discussion
The overexpression of certain cancer-specific
surface molecules and gene mutations has been identified in cancer
cells. Among these, the ErbB/EGFR family is one of the best known.
On the other hand, some cancer-specific surface molecules have been
successfully applied in targeted cancer therapy and imaging
(30,31). In our previous study, we
constructed a ErbB2-targeting protein by fusing the EC1 peptide
with GLuc and the p53C peptide. The EC1-GLuc-p53C exerts its
function for ErbB2-targeted bioluminescence and cancer therapy
in vitro. However, the function of EC1-GLuc-p53C in
vivo was very poor due to the very low accumulation in tumors
when intravenously injected (10). In the present study,
EC1-GLuc-liposome was constructed by conjugation EC1-GLuc-His to a
nickel-chelating liposome. In vitro experiments indicated
that the EC1-GLuc-liposome was selectively internalized and
delivered HPTS into ErbB2-overexpressing SKOv3 cells (Fig. 3B and C). ErbB2-targeted
bioluminescence imaging was carried out successfully in SKOv3 cells
(Fig. 4). In addition, the
bioluminescence imaging and selective delivery of HPTS into
metastatic SKOv3 tumors in vivo were also achieved using the
EC1-GLuc-liposome (Fig. 6). Thus,
the in vitro and in vivo experiments in the present
study demonstrated that the novel target liposome was effective for
ErbB2-targeted bioluminescence imaging and drug delivery.
In our previous studies, the immunoliposomes were
constructed by conjugating anti-EGFR antibody to liposomes using
the antibody affinity motif of protein A (ZZ) as an adaptor for
targeted DDS and the imaging of glioblastoma (18,22). These immunoliposomes effectively
targeted EGFR-overexpressing glioblastoma cells for the delivery of
borocaptate and bioluminescence imaging in vitro and in
vivo. However, the immunoliposome contains 3 main components:
the liposome, the ZZ protein and EGFR antibody. In the present
study, the target liposome only contained only 2 components: the
liposome and a small protein, EC1-GLuc. The decrease in components
may facilitate the construction of the target liposome and may also
decrease the size of the liposome. The diameter of the previous
immunoliposome was approximately 130 nm (22), but the diameter of the
EC1-GLuc-liposome was decreased to 117 nm (Table I). The smaller size may improve
the drug delivery efficience of liposomes into targeted cells
(32). In addition, it has been
reported that divalent or multivalent forms of the EC1 peptide
fused-Fc-liposome selectively target ErbB2-overexpressing breast
cancer cells and are efficiently internalized into cells (11). Consistent with the previous
studies, thye EC1-GLuc-liposome retains its affinity with ErbB2,
and is effectively internalized into ErbB2-overexpressing SKOv3
cells for bioluminescence imaging and drug delivery.
Ovarian carcinoma is one of the common
gynaecological tumors with a high metastatic rate (27,29). The metastatic foci of ovarian
carcinoma in the peritoneal cavity are very difficult to be
clean-up with conventional surgery, radiotherapy and chemotherapy
(33). As a result, patients with
ovarian carcinoma mainly succumb to the disease due to metastasis.
Therefore, a reliable approach for visualizing metastatic ovarian
tumors and a targeted DDS are urgently required in order improve
the therapeutic effects. In the present study, the
EC1-GLuc-liposome selectively targeted ErbB2 for bioluminescence
imaging and drug delivery for metastatic ovarian carcinoma in
vitro and in vivo. Therefore, the EC1-GLuc-liposome may
not only service as a surgical navigation to clean-up metastatic
ovarian carcinoma by bioluminescence imaging, but may also provide
a molecular targeting DDS to improve the chemotherapy and
radiotherapy in the future.
The combination of a diagnostic test and a
therapeutic entity is termed theranostics (34,35). In recent years, theranostics has
developed rapidly along with selective targeting strategies. The
targeting moieties include proteins (mainly antibodies and their
fragments), peptides and some small molecules (folate and flavin
mononucleotide) (35). Peptides
are attractive targeting molecules due to their small size, low
immunogenicity and ease of manufacture at low costs. In the present
study, we used the EC1 peptide as a targeting moiety. The
EC1-GLuc-liposome effectivly targeted ErbB2-overexpressing ovarian
cancer cells for bioluminescence imaging in vitro and in
vivo, and also proved to be effective in targeted DDS. Thus,
our multifunctional EC1-GLuc-liposome may prove to be a promising
theranostic reagent for ErbB2-overexpressing metastatic ovarian
carcinoma in the future, although the development of this
technology is still at the early stage and requires some
optimization.
Acknowledgements
This study was supported by grants from the National
Natural Science Foundation of China (31360241 and 81472371), the
Postgraduate Student Foundation for New Teacher from the Ministry
of Education of China (20123601120001) and the Foundation from
Department of Education of Jiangxi Province (GJJ13162).
References
|
1
|
Tzahar E, Waterman H, Chen X, Levkowitz G,
Karunagaran D, Lavi S, Ratzkin BJ and Yarden Y: A hierarchical
network of interreceptor interactions determines signal
transduction by Neu differentiation factor/neuregulin and epidermal
growth factor. Mol Cell Biol. 16:5276–5287. 1996.
|
|
2
|
Graus-Porta D, Beerli RR, Daly JM and
Hynes NE: ErbB-2, the preferred heterodimerization partner of all
ErbB receptors, is a mediator of lateral signaling. EMBO J.
16:1647–1655. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Yarden Y and Sliwkowski MX: Untangling the
ErbB signalling network. Nat Rev Mol Cell Biol. 2:127–137. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Slamon DJ, Godolphin W, Jones LA, et al:
Studies of the HER-2/neu proto-oncogene in human breast and ovarian
cancer. Science. 244:707–712. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Berchuck A, Kamel A, Whitaker R, et al:
Overexpression of HER-2/neu is associated with poor survival in
advanced epithelial ovarian cancer. Cancer Res. 50:4087–4091.
1990.PubMed/NCBI
|
|
6
|
Slamon DJ, Clark GM, Wong SG, Levin WJ,
Ullrich A and McGuire WL: Human breast cancer: correlation of
relapse and survival with amplification of the HER-2/neu oncogene.
Science. 235:177–182. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kim R, Tanabe K, Uchida Y, Osaki A and
Toge T: The role of HER-2 oncoprotein in drug-sensitivity in breast
cancer. Oncol Rep. 9:3–9. 2002.PubMed/NCBI
|
|
8
|
Pero SC, Shukla GS, Armstrong AL, Peterson
D, Fuller SP, Godin K, Kingsley-Richards SL, Weaver DL, Bond J and
Krag DN: Identification of a small peptide that inhibits the
phosphorylation of ErbB2 and proliferation of ErbB2 overexpressing
breast cancer cells. Int J Cancer. 111:951–960. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hashizume T, Fukuda T, Nagaoka T, Tada H,
Yamada H, Watanabe K, Salomon DS and Seno M: Cell type dependent
endocytic internalization of ErbB2 with an artificial peptide
ligand that binds to ErbB2. Cell Biol Int. 32:814–826. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Han XJ, Sun LF, Nishiyama Y, Feng B,
Michiue H, Seno M, Matsui H and Tomizawa K: Theranostic protein
targeting ErbB2 for bioluminescence imaging and therapy for cancer.
PLoS One. 8:e752882013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Vaidyanath A, Hashizume T, Nagaoka T,
Takeyasu N, Satoh H, Chen L, Wang J, Kasai T, Kudoh T, Satoh A, Fu
L and Seno M: Enhanced internalization of ErbB2 in SK-BR-3 cells
with multivalent forms of an artificial ligand. J Cell Mol Med.
15:2525–2538. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tannous BA: Gaussia luciferase
reporter assay for monitoring biological processes in culture and
in vivo. Nat Protoc. 4:582–591. 2009. View Article : Google Scholar
|
|
13
|
Lembert N and Idahl LA: Regulatory effects
of ATP and luciferin on firefly luciferase activity. Biochem J.
305:929–933. 1995.PubMed/NCBI
|
|
14
|
Matthews JC, Hori K and Cormier MJ:
Substrate and substrate analogue binding properties of
Renilla luciferase. Biochemistry. 16:5217–5220. 1977.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lorenz WW, McCann RO, Longiaru M and
Cormier MJ: Isolation and expression of a cDNA encoding
Renilla reniformis luciferase. Proc Natl Acad Sci USA.
88:4438–4442. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tannous BA, Kim DE, Fernandez JL,
Weissleder R and Breakefield XO: Codon-optimized Gaussia
luciferase cDNA for mammalian gene expression in culture and in
vivo. Mol Ther. 11:435–443. 2005.PubMed/NCBI
|
|
17
|
Venisnik KM, Olafsen T, Gambhir SS and Wu
AM: Fusion of Gaussia luciferase to an engineered
anti-carcinoembryonic antigen (CEA) antibody for in vivo optical
imaging. Mol Imaging Biol. 9:267–277. 2007.PubMed/NCBI
|
|
18
|
Feng B, Tomizawa K, Michiue H, Han XJ,
Miyatake S and Matsui H: Development of a bifunctional
immunoliposome system for combined drug delivery and imaging in
vivo. Biomaterials. 31:4139–4145. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Hyodo K, Yamamoto E, Suzuki T, Kikuchi H,
Asano M and Ishihara H: Development of liposomal anticancer drugs.
Biol Pharm Bull. 36:703–707. 2013. View Article : Google Scholar
|
|
20
|
Oude Blenke E, Mastrobattista E and
Schiffelers RM: Strategies for triggered drug release from tumor
targeted liposomes. Expert Opin Drug Deliv. 10:1399–1410.
2013.PubMed/NCBI
|
|
21
|
Yamada T, Iwasaki Y, Tada H, Iwabuki H,
Chuah MK, VandenDriessche T, Fukuda H, Kondo A, Ueda M, Seno M,
Tanizawa K and Kuroda S: Nanoparticles for the delivery of genes
and drugs to human hepatocytes. Nat Biotechnol. 21:885–890. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Feng B, Tomizawa K, Michiue H, Miyatake S,
Han XJ, Fujimura A, Seno M, Kirihata M and Matsui H: Delivery of
sodium borocaptate to glioma cells using immunoliposome conjugated
with anti-EGFR antibodies by ZZ-His. Biomaterials. 30:1746–1755.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tsutsui Y, Tomizawa K, Nagita M, Michiue
H, Nishiki T, Ohmori I, Seno M and Matsui H: Development of
bionanocapsules targeting brain tumors. J Control Release.
122:159–164. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Woodle MC and Lasic DD: Sterically
stabilized liposomes. Biochim Biophys Acta. 1113:171–199. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Duncan R: Polymer conjugates as anticancer
nanomedicines. Nat Rev Cancer. 6:688–701. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Jiang W, Kim BY, Rutka JT and Chan WC:
Advances and challenges of nanotechnology-based drug delivery
systems. Expert Opin Drug Deliv. 4:621–633. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jemal A, Murray T, Samuels A, Ghafoor A,
Ward E and Thun MJ: Cancer statistics, 2003. CA Cancer J Clin.
53:5–26. 2003. View Article : Google Scholar
|
|
28
|
Rose PG, Piver MS, Tsukada Y and Lau TS:
Metastatic patterns in histologic variants of ovarian cancer. An
autopsy study. Cancer. 64:1508–1513. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jiang Y, Berk M, Singh LS, Tan H, Yin L,
Powell CT and Xu Y: KiSS1 suppresses metastasis in human ovarian
cancer via inhibition of protein kinase C alpha. Clin Exp
Metastasis. 22:369–376. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Richter M and Zhang H: Receptor-targeted
cancer therapy. DNA Cell Biol. 24:271–282. 2005. View Article : Google Scholar
|
|
31
|
Haberkorn U, Markert A, Eisenhut M, Mier W
and Altmann A: Development of molecular techniques for imaging and
treatment of tumors. Q J Nucl Med Mol Imaging. 55:655–670.
2011.PubMed/NCBI
|
|
32
|
Jiang W, Kim BY, Rutka JT and Chan WC:
Nanoparticle-mediated cellular response is size-dependent. Nat
Nanotechnol. 3:145–150. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kapp KS, Kapp DS, Poschauko J,
Stücklschweiger GF, Hackl A, Pickel H, Petru E and Winter R: The
prognostic significance of peritoneal seeding and size of
postsurgical residual in patients with stage III epithelial ovarian
cancer treated with surgery, chemotherapy, and high-dose
radiotherapy. Gynecol Oncol. 74:400–407. 1999. View Article : Google Scholar
|
|
34
|
Del Vecchio S, Zannetti A, Fonti R, Pace L
and Salvatore M: Nuclear imaging in cancer theranostics. Q J Nucl
Med Mol Imaging. 51:152–163. 2007.
|
|
35
|
Yu MK, Park J and Jon S: Targeting
strategies for multifunctional nanoparticles in cancer imaging and
therapy. Theranostics. 2:3–44. 2012. View Article : Google Scholar : PubMed/NCBI
|