Introduction
Malignant tumors are characterized by uncontrolled
growth, invasion and metastasis. The elevated expression and
activation of oncogenes are strongly associated with the initiation
and progression of cancer. Ras, one of the well-characterized
proto-oncogenes, controls multiple intracellular signaling networks
(1). Aberrant Ras activation is
highly associated with cancer initiation, as well as progression,
and its presence has been shown in 20–30% of all human tumors,
particularly in colon and pancreatic cancer (2,3).
Radiation therapy is an important therapeutic
strategy in cancer treatment. Minimizing injury to the surrounding
normal tissue and organs, while increasing the anticancer effects
remains an unresolved issue in the field of radiation therapy. Ras
has been studied and linked to radioresistance. A number of studies
have reported that the inhibition of Ras activation by anti-Ras
antibody or Ras inhibitors can enhance radiation-induced cell death
in human tumor cell lines (4,5).
The involvement of the phosphoinositide 3-kinase (PI3K)/protein
kinase B (Akt) signaling pathway in Ras-induced radioresistance
suggests that the inhibition of PI3K/Akt activity can greatly
enhance the radiosensitivity of Ras-transformed cells (6). Everolimus is an inhibitor of
mammalian target of rapamycin (mTOR) and a derivative of rapamycin
with immunosuppressiveand anticancer properties (7,8).
The anticancer effects of everolimus are associated with the
inhibition of the PI3K/AKT/mTOR pathway (8,9).
Thus, everolimus may act as a radiosensitizer in Ras-transformed
cells. The present study aimed to investigate the radiosensitizing
effects of everolimus and to elucidate the molecular mechanisms
underlying everolimus-mediated radiosensitization.
Materials and methods
Chemicals and antibodies
Everolimus was obtained from Novartis (Basel,
Switzerland) and dissolved in dimethyl sulfoxide (DMSO) as a
concentrated stock solution. DMSO was used as the vehicle solution.
As an internal control, medium only was used. DMSO and anti-actin
antibody were from Sigma-Aldrich (St. Louis, MO, USA). Antibodies
against Ras, phospho-eukaryotic initiation factor 4E-binding
protein 1 (p-4EBP1), 4EBP1, phospho-ribosomal protein S6 kinase 1
(p-S6K1), S6K1 and phospho-Akt (p-Akt; Ser-473) were from Upstate
Biotechnology (Lake Placid, NY, USA). Antibodies against
phospho-mTOR (p-mTOR), mTOR, Akt and eukaryotic translation
initiation factor 4E (eIF4E) were from BD Biosciences (San Diego,
CA, USA). Beclin 1 small interfering RNA (siRNA) was from Thermo
Fisher Scientific (Carlsbad, CA, USA).
Cell culture
RK3E/tv-a cells were obtained from the laboratory of
Shu-Ling Fu, who established a non-transformed rat kidney
epithelial RK3E cell line that constitutively expresses tv-a
(receptor for subgroup A avian leukosis virus, ALV) for the
delivery of foreign genes via avian retroviral infection. The
RK3E/tv-a cells were infected with retrovirus carrying the
oncogene, H-ras, or the control gene puromycin N-acetyltransferase
(puro) as previously described (10). Ras-expressing cells, but not
puro-expressing cells, underwent malignant transformation in
vitro and tumorigenesis in vivo. These cells were
maintained as previously described (10).
MTT assays
3-(4,5-Dimethyl-2-thiazolyl)-2,5-diphenyl-2H
tetrazolium bromide (MTT) assays were carried out to measure the
cytotoxic and radiosensitizing effects of everolimus.
Ras-transformed cells (5×103) were seeded into 96-well
plates, incubated overnight, treated with everolimus and/or
radiation and cultured for 4 days. The medium was removed and the
cells were incubated with MTT solution (0.5 mg/ml, 4 h, overnight,
5% CO2 incubator, 37°C) and lysed with solubilization
solution [12.5% sodium dodecyl sulfate (SDS), 45% dimethylformamide
(DMF)]. The absorbance of converted dye was measured at 570 nm
using a microplate reader. (Multiskan RC, Labsystems, Stockholm,
Sweden). The absorbance of the untreated cells measured at 570 nm
was defined as 100%. The relative viability was calculated by
dividing the A570 nm of the treated cells by that of the untreated
cells.
siRNA-mediated knockdown of Beclin 1
The Ras-transformed cells (4×105) were
seeded into a 6-cm dish, cultured overnight, and transfected at 40%
confluency with the indicated specific constructs (Beclin 1 siRNA,
or non-targeted control siRNA) for 24 h according to the
manufacturer’s instructions (Thermo Scientific Dharmacon Inc.,
Lafayette, CO, USA). These sequences were 5′-GGACAGUUUGGCACAAUCA-3′
(951–969). For each transfection, 2 solutions (one of 3 μl of siRNA
diluted in 100 μl transfection medium and another of 3 μl of
transfection reagents diluted in 100 μl transfection medium) were
combined at room temperature for 20 min and added to each well, and
the wells were incubated for 24 h. The transfection efficiency was
determined at 24 h by western blot analysis. MTT assays were used
to measure the effects of everolimus-mediated
radiosensitization.
Annexin V staining
An Annexin V-FITC kit was used according to the
manufacturer’s instructions (R&D Systems Inc., Minneapolis, MN,
USA). In brief, the cells (105) were grown on 6-well
plates, treated with the vehicle (DMSO) or everolimus (10 nM) with
or without 6 Gy radiation, cultured for 96 h and stained with
Annexin V-FITC and propidium iodide (PI) (5 μg/ml). The stained
cells were analyzed using a flow cytometer (FACScalibur;
Becton-Dickinson, San Jose, CA, USA).
Terminal deoxynucleotidyl
transferase-mediated dUTP nick-end labeling (TUNEL) assay
A (TUNEL) assay was performed to measure cell death
according to the manufacturer’s instructions (Roche, Mannheim,
Germany). The cells (105) were seeded into 6-well
plates, incubated overnight and subsequently treated with the
vehicle, everolimus and/or radiation under various conditions.
Following treatment, the floating and adherent cells were collected
and fixed with 4% paraformaldehyde in PBS, permeablized with 0.1%
Triton X-100, stained with TUNEL reaction mixture for 60 min and
analyzed by flow cytometry using a FACScan flow cytometer
(Becton-Dickinson).
Cell cycle analysis
Cells (105) were grown on 6-well plates,
treated under various conditions, incubated for 96 h at 37°C, then
trypsinized, washed, fixed in 80% cold ethanol for 30 min, stained
with 50 μg/ml propium iodide in PBS containing 50 μg/ml of RNase A
for 30 min, and immediately analyzed by flow cytometry using a
FACScan flow cytometer.
Western blot analysis
The cells (106) were seeded into 100-mm
plates, incubated overnight, treated with the vehicle, everolimus
or radiation under various conditions, and collected by scraping
and centrifugation at 3,000 rpm for 5 min. The cell pellets were
then lysed in radioimmune precipitation assay buffer (RIPA) in the
presence of general protease inhibitors. Total protein (50 μg of
lysate proteins) was analyzed by 10% sodium dodecyl sulfate
polyacrylamide-gel electrophoresis (SDS-PAGE) and transferred onto
PVDF membranes. After blocking with 5% non-fat milk/TBS-Tween, the
membranes were incubated with primary antibodies, including
anti-p-Akt, anti-Akt, anti-p-mTOR, anti-mTOR, anti-phospho-S6K
(p-S6K), anti-S6K, anti-p-4EBP1, anti-4EBP1, anti-eIF4E and
anti-LC3 antibodies overnight at 4°C, reacted with horseradish
peroxidase (HRP)-conjugated secondary antibodies for 1 h, treated
with Immobilon™ Western Chemiluminescent HRP Substrate (ECL) and
exposed to X-ray film to detect the signals.
Statistical analysis
Data are expressed as the means ± SD of 3independent
experiments. Statistical analyses were performed using the unpaired
two-tailed Student’s t-test to compare 2 groups of data. A value of
P<0.05 was considered to indicate a statistically signficant
difference.
Results
Everolimus-mediated radiosensitization of
Ras-transformed cells in vitro
We first evaluated the cytotoxicity of everolimus to
Ras-transformed cells by MTT assay. The cytotoxic effects were
found to increase with the higher everolimus concentrations. The
survival fraction of the cells treated with everolimus was (10, 30
and 50 nM) was 0.91±0.03, 0.84±0.04 and 0.8±0.02, respectively.
Thus, everolimus (30 nM; 96 h of incubation) was used in the
subsequent radiosensitivity experiments. As shown in Fig. 1, treatment with everolimus (30 nM)
sensitized the Ras-transformed cells to radiation (6 Gy). The
survival fractions of the irradiated cells in the medium, vehicle
and everolimus solution were 0.45±0.04, 0.42±0.04 and 0.12±0.04,
respectively.
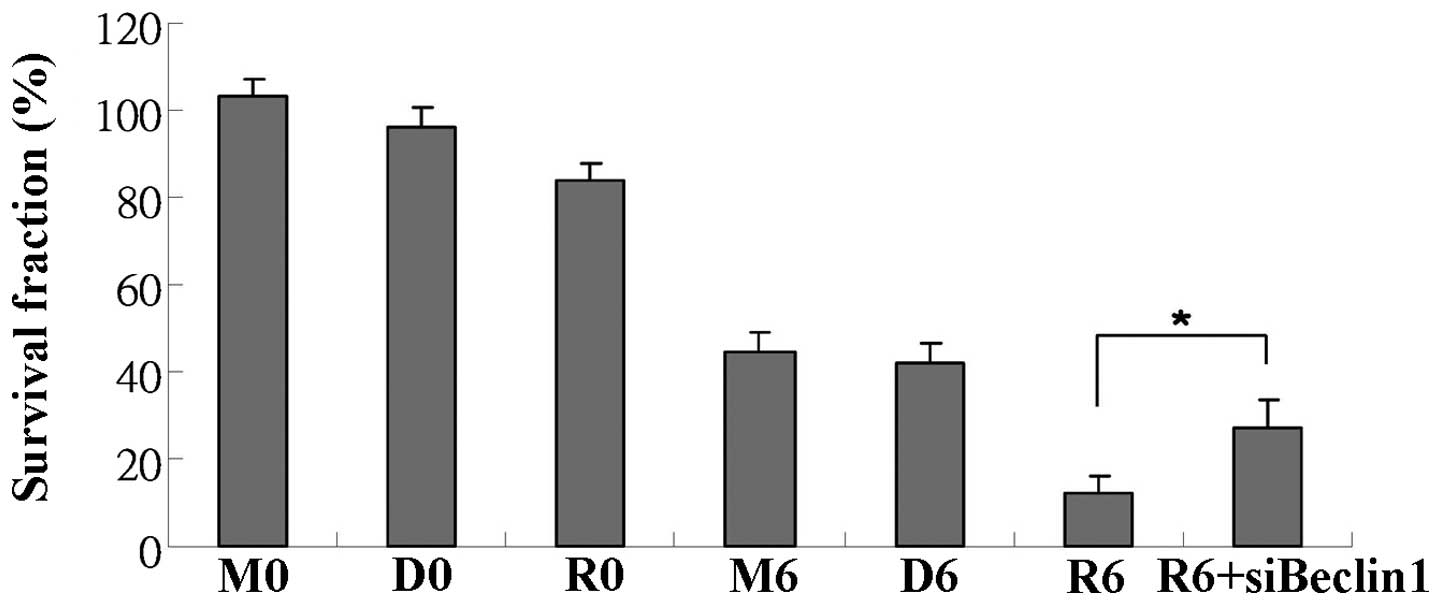 | Figure 1Everolimus sensitizes Ras-transformed
cells to radiation in vitro. MTT assay was used to determine
the radiosensitizing effects of everolimus. The cell survival
fractions were calculated, and the calculated cell survival
fractions following 6 Gy of radiation and treatment with medium
only, vehicle (DMSO) or everolimus solution (stock diluted
appropriately in medium) were 0.45±0.04, 0.42±0.04 and 0.12±0.04,
respectively. In addition, the radiosensitizating effects of
everolimus were reversed following transfection with Beclin 1
siRNA. *P<0.05. M0: medium, 0 Gy; M6: medium, 6 Gy;
D0: 0.1% DMSO, 0 Gy; D6: 0.1% DMSO, 6 Gy; R0: everolimus (30 nM), 0
Gy; R6: everolimus (30 nM), 6 Gy radiation. |
Everolimus does not promote
radiation-induced apoptosis
We wished to determine whether everolimus, with or
without radiation, induces apoptosis. Thus, we performed Annexin V
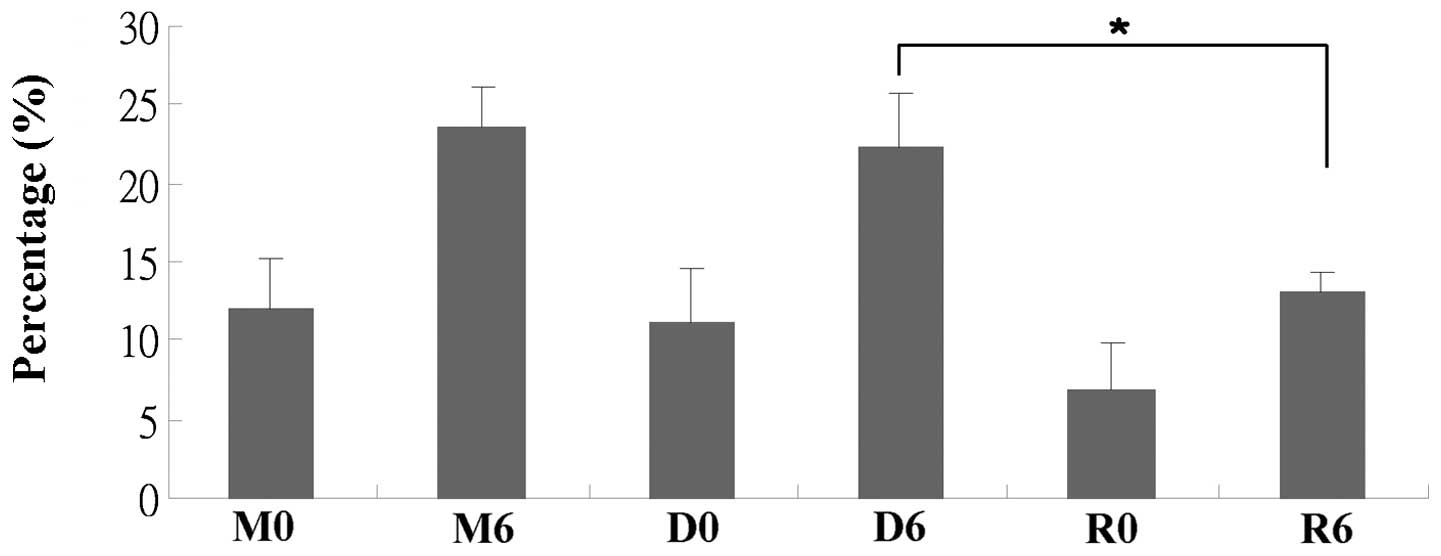
staining and TUNEL assay. The percentage of Annexin V-stained cells
was increased following radiation, but not following treatment with
everolimus (Fig. 2). Consistent
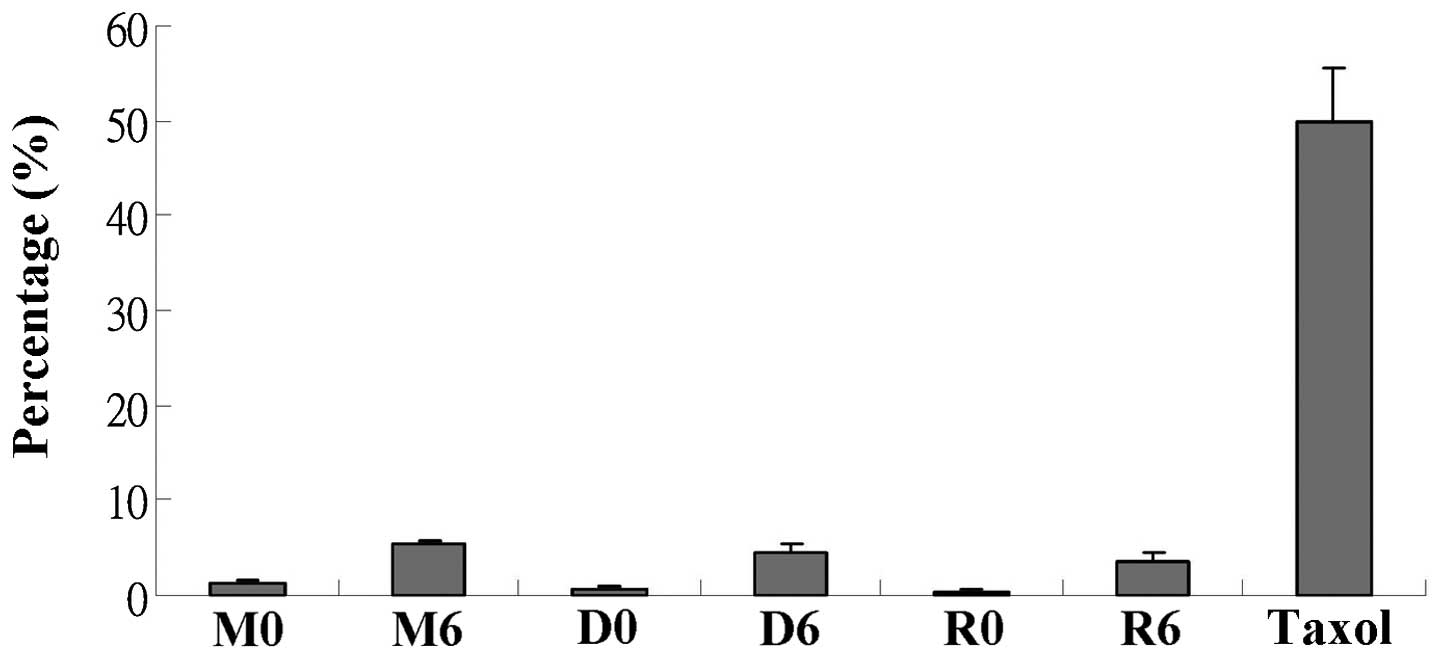
with the above observations, DNA fragmentation (measured by TUNEL
assay) was mainly caused by radiation (Fig. 3). By contrast, combination
treatment with everolimus and radiation decreased the percentage of
dead/apoptotic cells. Taken together, these data indicate that
apoptosis is mainly induced by radiation and not is promoted by
everolimus. The decreased percentage of apoptotic cell death
suggests that everolimus-mediated radiosensitization leading to
cell death may occur through a mechanism other than apoptosis.
Radiation affects cell cycle
distribution
We wished to determine whether everolimus, with or
without radiation, affects cell cycle distribution. Thus, we used
PI staining followed by flow cytometry using a FACScan flow
cytometer. As shown in Table I,
radiation significantly increased the size of the sub-G1
and G2/M fractions (P<0.05), and radiation in
combination with everolimus (compared to radiation alone) only
slightly decreased the size of the sub-G1 fraction.
 | Table ICell cycle distribution under various
treatment conditions. |
Table I
Cell cycle distribution under various
treatment conditions.
| Phase | M0 | M6 | D0 | D6 | R0 | R6 |
|---|
|
Sub-G1 | 2.9±0.8 | 7.1±2.0 | 3.1±0.3 | 7.8±4.1 | 2.2±1.4 | 5.8±2.9 |
|
G0/G1 | 64.9±6.4 | 47.5±6.1 | 66.0±4.8 | 48.1±6.0 | 62.0±3.7 | 49.4±6.3 |
| S | 14.3±2.3 | 15.2±1.8 | 13.3±0.9 | 14.5±1.2 | 13.9±0.7 | 13.9±1.4 |
| G2/M | 18.1±4.6 | 30.5±3.1 | 18.1±4.6 | 30.0±2.6 | 22.3±2.9 | 31.3±2.9 |
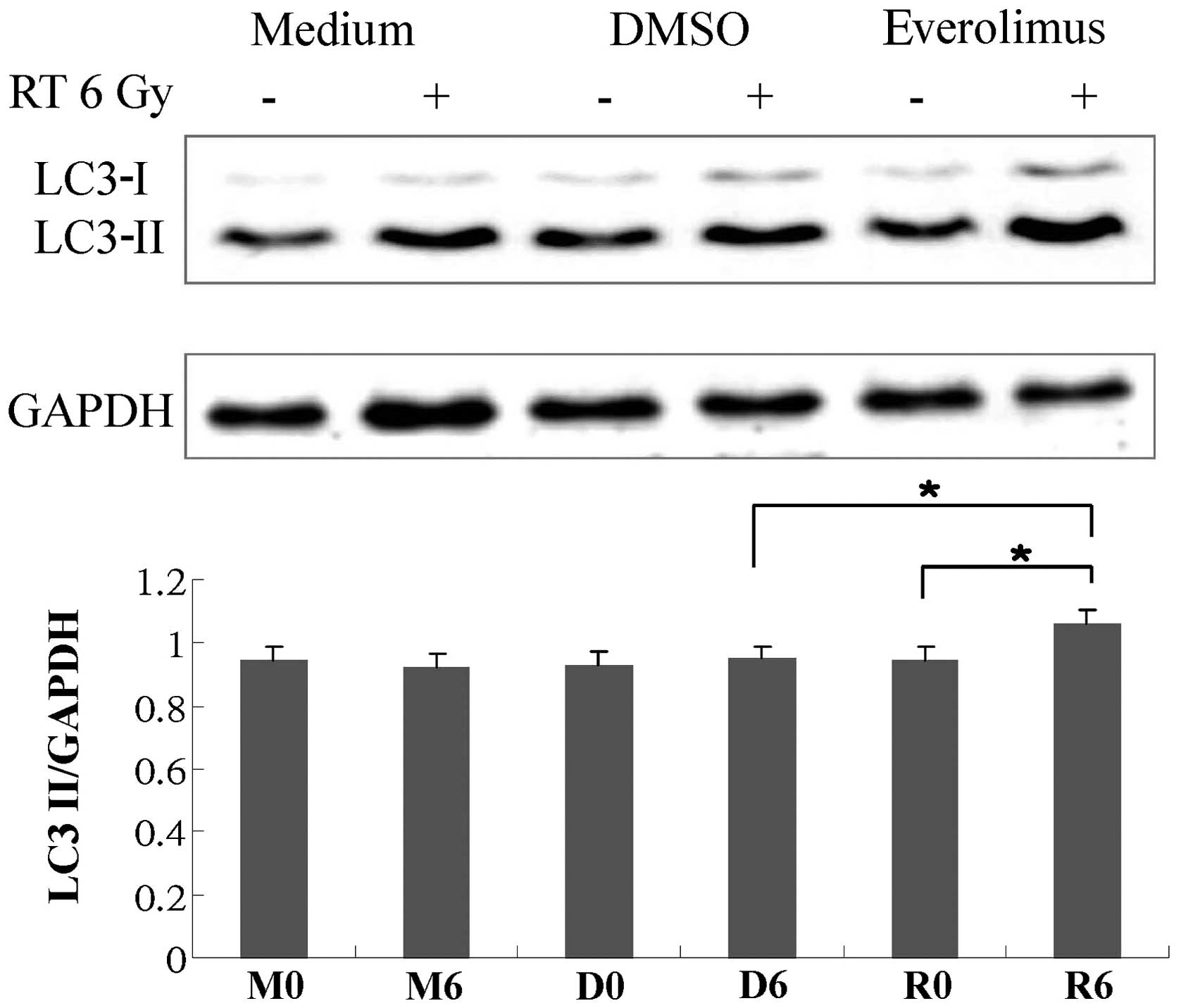
LC3-II expression increases following
treatment with everolimus in combination with radiation and the
radiosensitizing effects of everolimus are reversed by transfection
with Beclin 1 siRNA
We wished to determine whether everolimus, with or
without radiation, induces autophagy. Thus, we performed western
blot analysis. Fig. 4 shows the
protein expression of LC3-II under various conditions. Everolimus
in combination with radiation significantly increased LC3-II
expression. We then wished to determine whether the
everolimus-mediated radiosensitizing effects were dependent on
autophagy. Thus, we transfected the cells with Beclin 1 siRNA. As
shown in Fig. 1, the
radiosensitization effects of everolimus were partially reversed
following transfection with Beclin 1 siRNA.
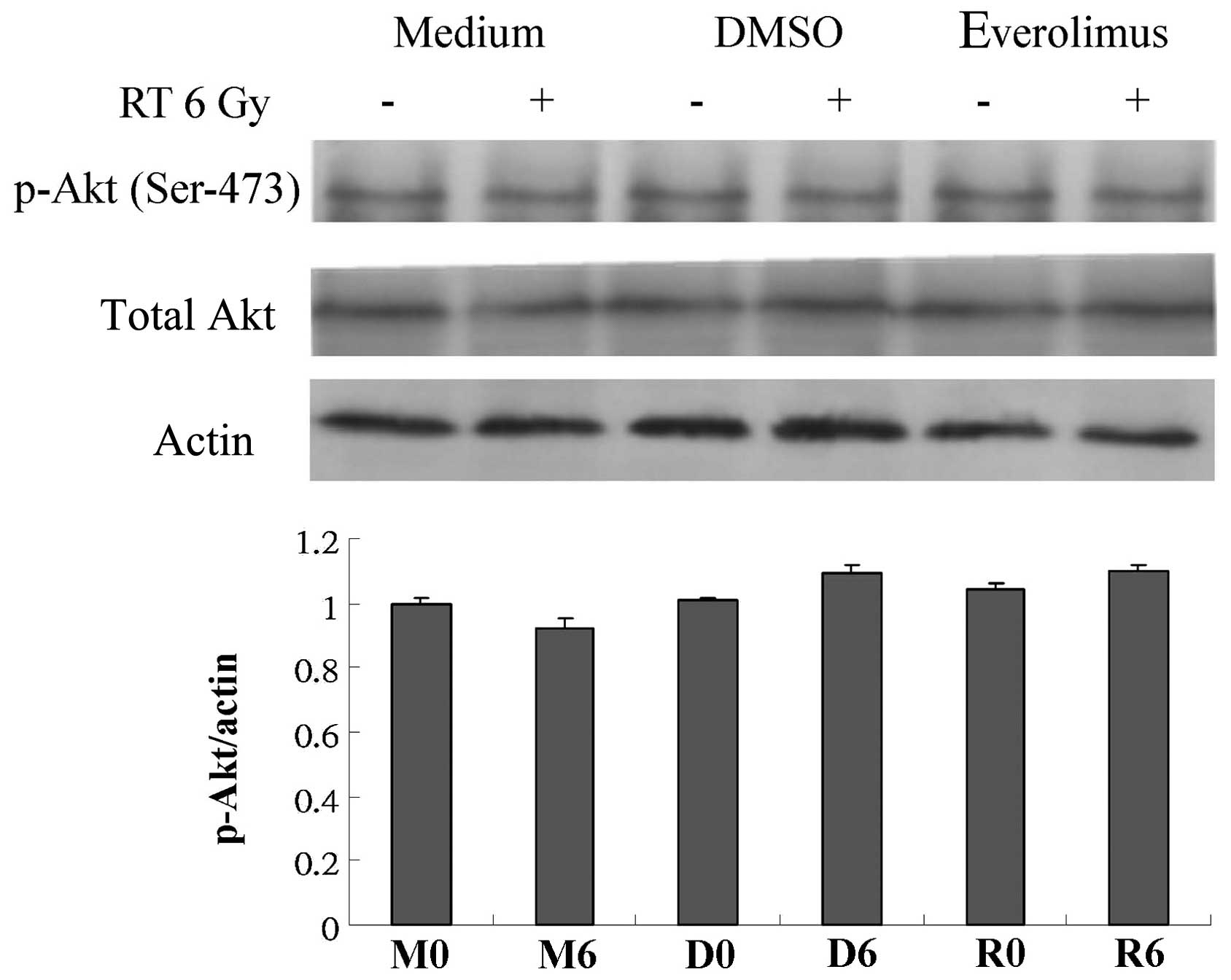
Akt protein expression is not altered by
everolimus or radiation
Since the PI3K/Akt pathway is a target of
everolimus, the possible association of the radiosensitizing
activity of everolimus with the PI3K/Akt signaling pathway was
assessed. Western blot analysis revealed that the expression or
activation of Akt protein was not significantly altered by
everolimus or radiation (Fig.
5).
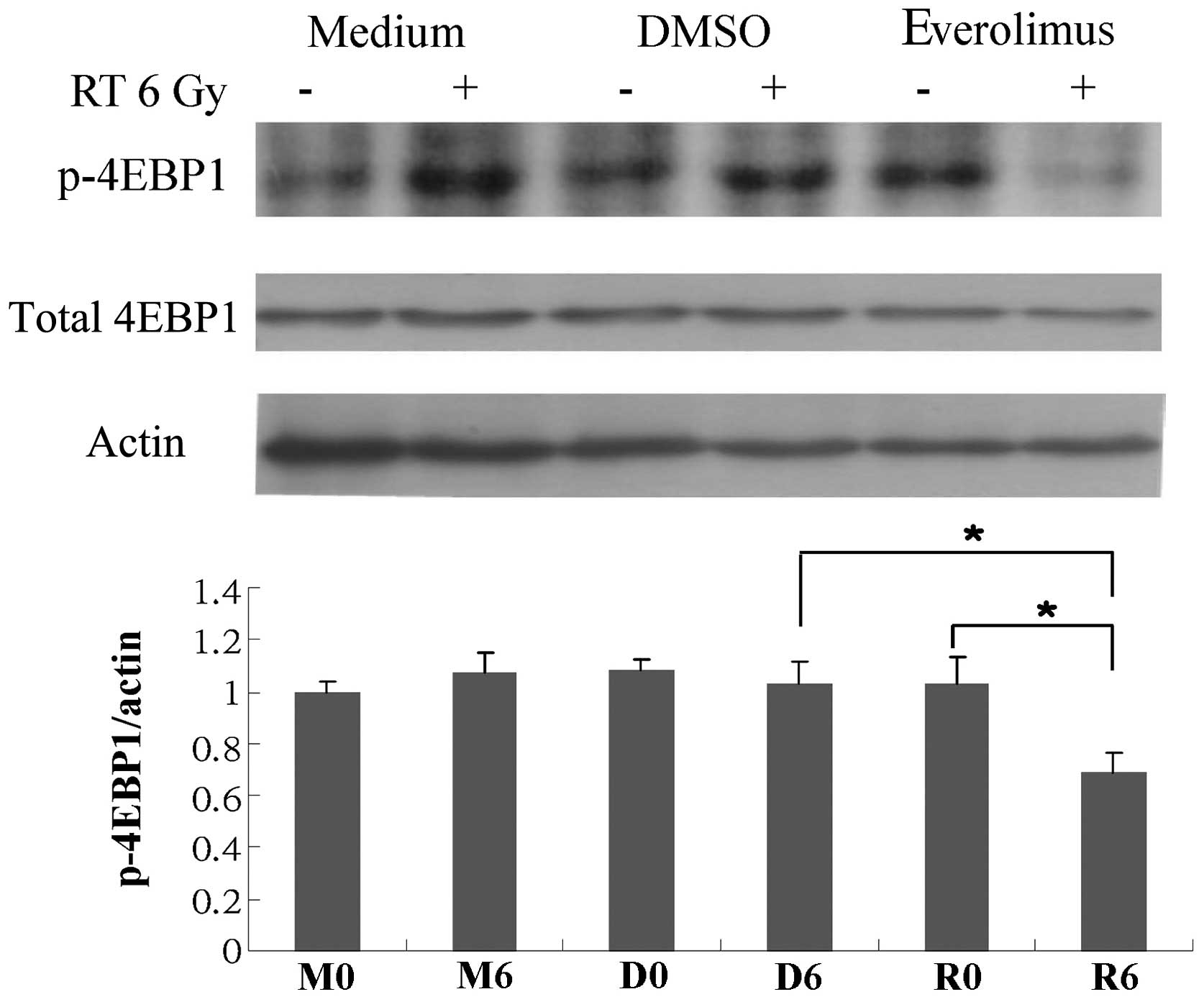
Phosphorylation of 4EBP1 is suppressed
following treatment with everolimus in combination with
radiation
We wished to determine whether everolimus, with or
without radiation, regulates the PI3K/AKT/mTOR pathway. Thus, we
performed western blot analysis in order to determine the protein
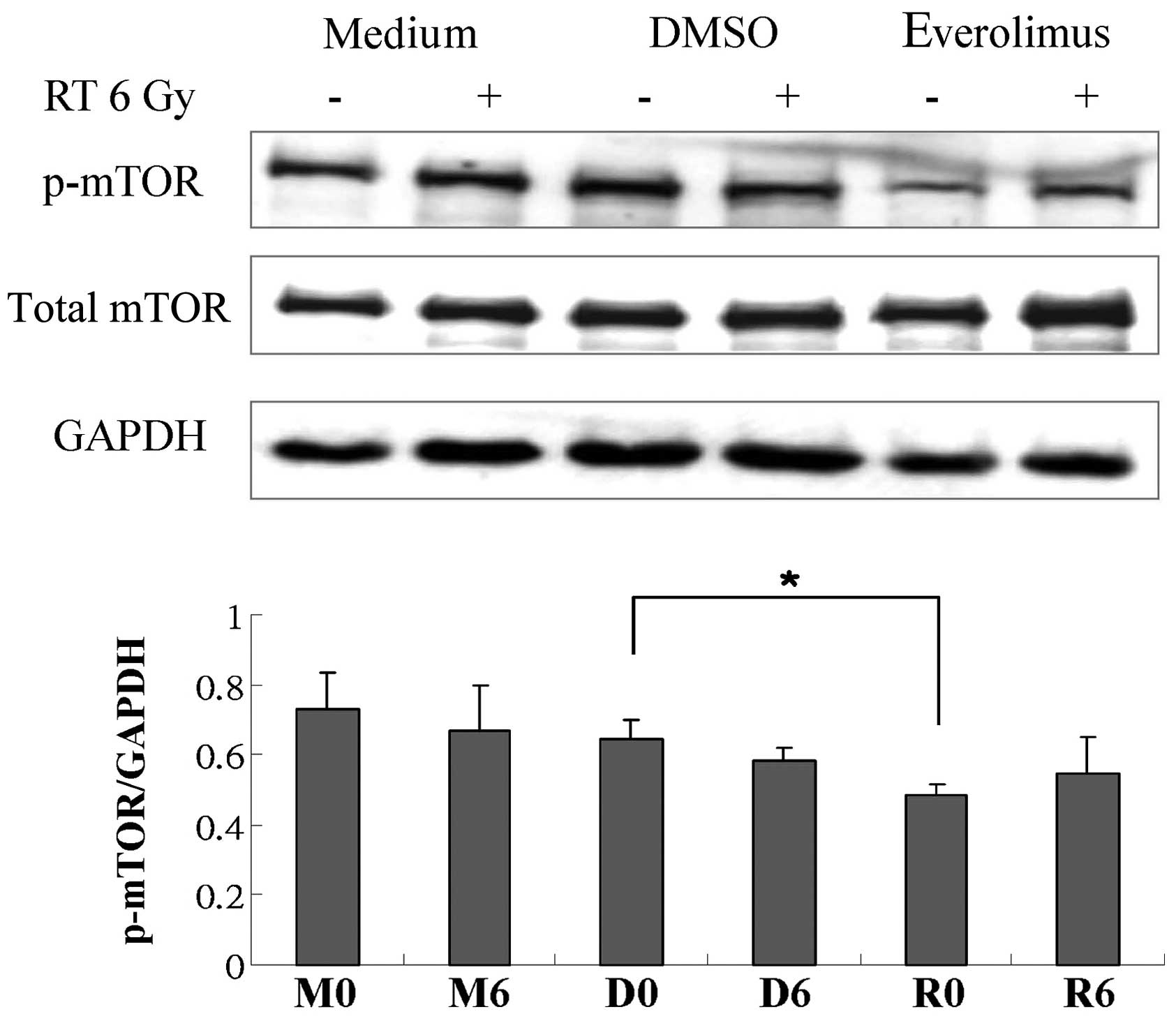
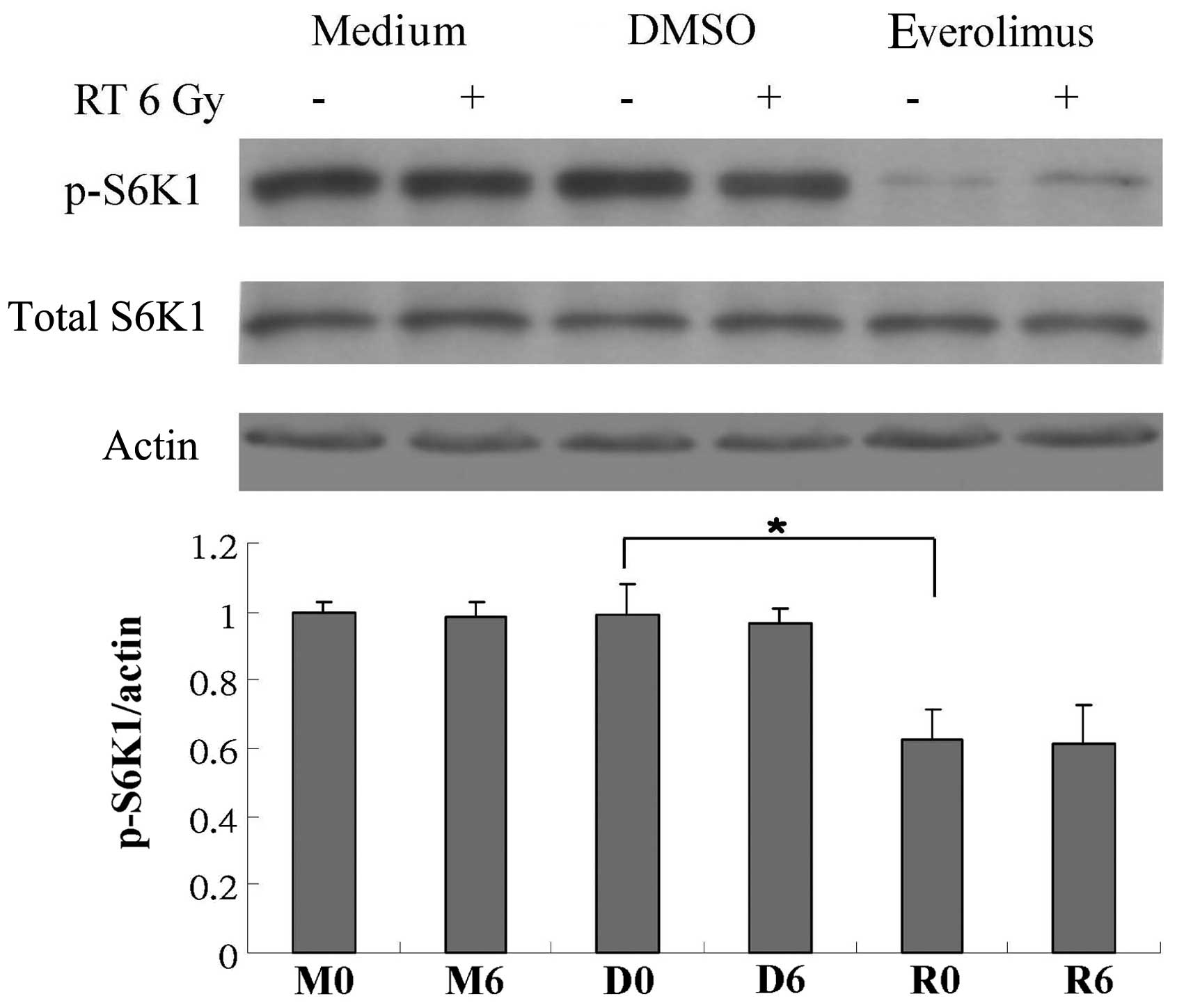
expression of p-mTOR, mTOR, p-S6K1, S6K1, p-4EBP1, 4BEP1 and eIF4E.
There was no significant change observed in eIF4E expression (data
not shown). However, treatment with everolimus alone attenuated the
phosphorylation of mTOR and S6K1 (Figs. 6 and 7), and everolimus in combination with
radiation markedly suppressed the phosphorylation of 4EBP1
(Fig. 8).
Discussion
Activating Ras mutations are frequently observed in
a number of human cancers and are strongly correlated with
radiation resistance (3,4,11).
Colorectal cancer is one of the most frequently diagnosed cancers
with aberrant Ras activation and a major cause of cancer-related
mortality in Taiwan (2,3,12).
In the present study, we demonstrated the radiosensitizing activity
of everolimus in Ras-transformed cells, suggesting a potential new
application of this compound in cancer therapy.
Different types of cell death with possible
associations to everolimus-mediated radiosensitization were
examined. Our results revealed that radiation-induced apoptotic
changes (sub-G1 cell accumulation, membrane inversion
and DNA fragmentation) were not enhanced by treatment with
everolimus (Table I; Figs. 2 and 3). Notably, the percentage of apoptotic
cells decreased following treatment with everolimus combined with
radiation relative to the percentage following radiation alone,
suggesting that other non-apoptotic pathways may be involved in the
everolimus-mediated radiosensitization. Indeed, increasing evidence
indicates that non-apoptotic mechanisms, such as autophagy, are
crucial for treatment-induced tumor-cell death (13,14). Whether the cell death caused by
everolimus-mediated radiosensitization is induced by autophagy was
examined by assessing the protein expression of LC3. Western blot
analysis revealed that treatment with everolimus plus radiation
markedly increased LC3-II expression (Fig. 4). In addition, siRNA targeting
Beclin 1 was used to further determine whether the
everolimus-mediated radiosensitizing effects are dependent on
autophagy. As shown in Fig. 1,
the radiosensitizing effects of everolimus were reversed following
transfection with Beclin 1 siRNA. These data indicate that the
autophagy pathway is involved in the everolimus-mediated
radiosensitizing effects.
Multiple signaling pathways, including the
PI3K/AKT/mTOR and Raf/MEK/ERK pathways, are associated with the
pharmacological activity of everolimus (15,16). The importance of the PI3K/AKT/mTOR
pathway in tumor progression has been investigated (17). In addition, it has been shown that
mTOR inhibitors target a number of cancers and regulate abnormal
signaling (18). mTOR signaling
is mediated through two multiprotein complexes, the mTOR complex 1
(mTORC1) and 2 (mTORC2). mTORC1 can activate 4EBP1 and S6K1 and
lead to translation, increasing protein synthesis and growth
(18). Several studies have
demonstrated that mTOR inhibitors modulate autophagy. A previous
study suggested that p-4EBP1 binds to and stabilizes mTORC1 in
vitro (19). Another study
demonstrated that the effects of 4EBP1 on autophagy were
significant, as evidenced by the increased baseline levels of
autophagy, the increased rapamycin-induced autophagy and the
elimination of 4EBP1 in the presence of the autophagy inhibitor
(20). Furthermore, S6K1 is
sensitive to the modulation of autophagy and requires a much less
stringent inhibition of mTORC1 to be targeted (21). In the present study, the
combination of everolimus and radiation markedly suppressed the
phosphorylation of 4EBP1 (Fig.
8). In addition, treatment with everolimus alone attenuated the
phosphorylation of S6K1 (Fig. 7).
These findings suggest that 4EBP1 and S6K1 interact with specific
proteins involved in autophagy and may have as yet unrecognized
actions that affect mTOR signaling and/or autophagy.
In conclusion, the present study demonstrates that
everolimus may be used as a novel radiosensitizer in
Ras-transformed cells, and that the mechanisms responsible for the
everolimus-mediated radiosensitization are partially dependent on
the autophagy pathway.
Acknowledgements
The present study was supported by the Buddhist
Dalin Tzu Chi General Hospital [DTCRD102 (2)-I-15].
References
|
1
|
Kolch W: Ras/Raf signalling and emerging
pharmacotherapeutic targets. Expert Opin Pharmacother. 3:709–718.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Bos JL, Fearon ER, Hamilton SR, et al:
Prevalence of ras gene mutations in human colorectal cancers.
Nature. 327:293–297. 1987. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kiaris H and Spandidos D: Mutations of ras
genes in human tumors (Review). Int J Oncol. 7:413–421.
1995.PubMed/NCBI
|
|
4
|
Bernhard EJ, McKenna WG, Hamilton AD, et
al: Inhibiting Ras prenylation increases the radiosensitivity of
human tumor cell lines with activating mutations of ras
oncogenes. Cancer Res. 58:1754–1761. 1998.PubMed/NCBI
|
|
5
|
Russell JS, Lang FF, Huet T, et al:
Radiosensitization of human tumor cell lines induced by the
adenovirus-mediated expression of an anti-Ras single-chain antibody
fragment. Cancer Res. 59:5239–5244. 1999.PubMed/NCBI
|
|
6
|
Gupta AK, Bakanauskas VJ, Cerniglia GJ, et
al: The Ras radiation resistance pathway. Cancer Res. 61:4278–4282.
2001.PubMed/NCBI
|
|
7
|
Budde K, Lehner F, Sommerer C, et al:
Conversion from cyclosporine to everolimus at 4.5 months
posttransplant: 3-year results from the randomized ZEUS study. Am J
Transplant. 12:1528–1540. 2012.PubMed/NCBI
|
|
8
|
Yunokawa M, Koizumi F, Kitamura Y, et al:
Efficacy of everolimus, a novel mTOR inhibitor, against basal-like
triple-negative breast cancer cells. Cancer Sci. 103:1665–1671.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Wolin EM: PI3K/Akt/mTOR pathway inhibitors
in the therapy of pancreatic neuroendocrine tumors. Cancer Lett.
335:1–8. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fu SL, Huang YJ, Liang FP, et al:
Malignant transformation of an epithelial cell by v-Src via
tv-a-mediated retroviral infection: a new cell model for
studying carcinogenesis. Biochem Biophys Res Commun. 338:830–838.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Samid D, Miller AC, Rimoldi D, Gafner J
and Clark EP: Increased radiation resistance in transformed and
nontransformed cells with elevated ras proto-oncogene expression.
Radiat Res. 126:244–250. 1991. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Chen LT and Whang-Peng J: Current status
of clinical studies for colorectal cancer in Taiwan. Clin
Colorectal Cancer. 4:196–203. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Ito H, Daido S, Kanzawa T, Kondo S and
Kondo Y: Radiation-induced autophagy is associated with LC3 and its
inhibition sensitizes malignant glioma cells. Int J Oncol.
26:1401–1410. 2005.PubMed/NCBI
|
|
14
|
Okada H and Mak TW: Pathways of apoptotic
and non-apoptotic death in tumour cells. Nat Rev Cancer. 4:592–603.
2004. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Cavazzoni A, Bonelli MA, Fumarola C, et
al: Overcoming acquired resistance to letrozole by targeting the
PI3K/AKT/mTOR pathway in breast cancer cell clones. Cancer Lett.
323:77–87. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Endo M, Yamamoto H, Setsu N, et al:
Prognostic significance of AKT/mTOR and MAPK pathways and antitumor
effect of mTOR inhibitor in NF1-related and sporadic malignant
peripheral nerve sheath tumors. Clin Cancer Res. 19:450–461. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Majumder PK, Febbo PG, Bikoff R, et al:
mTOR inhibition reverses Akt-dependent prostate intraepithelial
neoplasia through regulation of apoptotic and HIF-1-dependent
pathways. Nat Med. 10:594–601. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Strimpakos AS, Karapanagiotou EM, Saif MW
and Syrigos KN: The role of mTOR in the management of solid tumors:
an overview. Cancer Treat Rev. 35:148–159. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang L, Rhodes CJ and Lawrence JC Jr:
Activation of mammalian target of rapamycin (mTOR) by insulin is
associated with stimulation of 4EBP1 binding to dimeric mTOR
complex 1. J Biol Chem. 281:24293–24303. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Balakumaran BS, Porrello A, Hsu DS, et al:
MYC activity mitigates response to rapamycin in prostate cancer
through eukaryotic initiation factor 4E-binding protein 1-mediated
inhibition of autophagy. Cancer Res. 69:7803–7810. 2009. View Article : Google Scholar
|
|
21
|
Nyfeler B, Bergman P, Triantafellow E, et
al: Relieving autophagy and 4EBP1 from rapamycin resistance. Mol
Cell Biol. 31:2867–2876. 2011. View Article : Google Scholar : PubMed/NCBI
|