Introduction
Osteoblasts and osteoclasts are functional cells,
which are responsible for bone formation and bone resorption,
respectively (1). These two types
of cells are major regulators of bone metabolism. Bone remodeling
is essential for maintaining the structure and strength of skeletal
tissue and is caused by the coupling processes of osteoclastic bone
resorption and osteoblastic bone formation. It is generally
recognized that numerous humoral factors, including prostaglandins
(PGs) and cytokines play important roles in the process of bone
remodeling (2). Osteoblasts
possess the receptors of a number of bone resorptive factors, such
as parathyroid hormone, which promotes the formation of functional
osteoclasts through the upregulation of receptor activator of
nuclear factor-κB (RANK) ligand (RANKL) (1,2).
Therefore, osteoblasts are considered to play pivotal roles in the
regulation of not only bone formation, but also bone resorption.
Osteoprotegerin (OPG) is a glycoprotein, belonging to the tumor
necrosis factor receptor superfamily (3). OPG, which is produced and secreted
from osteoblasts, binds to RANKL as a decoy receptor and inhibits
the binding of RANKL to RANK, an essential step of
osteoclastogenesis (development of osteoclast-precursor cells into
mature osteoclasts) (3,4). Thus, it is currently recognized that
the RANK/RANKL/OPG axis is an important regulatory system for
functional osteoclast formation (5).
It has been firmly established that PGs act as
autocrine and paracrine regulators for osteoblasts and play
essential roles in the regulation of bone metabolism (6,7).
Among these, prostaglandin E2 (PGE2) is a
potent stimulator of bone resorption as it enhances osteoclast
formation (8). As regards the
intracellular signaling of PGE2 in osteoblasts, we have
previously demonstrated that PGE2 stimulates
interleukin-6 (IL-6) synthesis through Ca2+ mobilization
and cAMP production in osteoblast-like MC3T3-E1 cells (9). In addition, we have previously
demonstrated that PGE2 stimulates the induction of heat
shock protein 27 (HSP27) through Ca2+ mobilization and
protein kinase C (PKC)-dependent activation of both p44/p42
mitogen-activated protein (MAP) kinase and p38 MAP kinase in
MC3T3-E1 cells (10). However,
the effects of PGE2 on the synthesis of OPG in
osteoblasts and the mechanisms involved have not yet been
elucidated.
Resveratrol, which is a natural polyphenol
abundantly found in red grapes and berries, is recognized as
possessing antioxidant properties that exert various beneficial
effects on human health (11). It
has been reported that resveratrol increases the life span of lower
organisms by activating sirtuin 1 (SIRT1), a nicotinamide adenine
dinucleotide (NAD+)-dependent class III deacetylase
(12). The various beneficial
effects of resveratrol are considered to be mediated through the
activation of SIRT1 in mammalians as well (13). As regards bone cells, we have
recently reported that resveratrol suppresses the synthesis of
vascular endothelial growth factor (VEGF) stimulated by bone
morphogenetic protein-4 (BMP-4) in a SIRT1 activation-dependent
manner in osteoblast-like MC3T3-E1 cells (14). However, the mechanisms underlying
the effects of resveratrol on bone metabolism remain to be
elucidated.
In the present study, we investigated the mechanisms
of PGE2-induced OPG synthesis and the effects of
resveratrol on OPG synthesis by PGE2 in osteoblast-like
MC3T3-E1 cells. We demonstrate that resveratrol suppresses the
PGE2-stimulated OPG synthesis by inhibiting p44/p42 MAP
kinase, p38 MAP kinase and stress-activated protein
kinase/c-Jun N-terminal kinase (SAPK/JNK) in osteoblasts and
that the inhibitory effects are independent of the activation of
SIRT1.
Materials and methods
Materials
Resveratrol, SRT1720, PD98059, SB203580 and SP600125
were obtained from Calbiochem-Novabiochem Co. (La Jolla, CA, USA).
PGE2 was obtained from Sigma Chemical Co. (St. Louis,
MO, USA). A mouse OPG enzyme-linked immunosorbent assay (ELISA) kit
was obtained from R&D Systems, Inc. (Minneapolis, MN, USA).
Phospho-specific p44/p42 MAP kinase antibodies, p44/p42 MAP kinase
antibodies, phospho-specific p38 MAP kinase antibodies, p38 MAP
kinase antibodies, phospho-specific SAPK/JNK antibodies and
SAPK/JNK antibodies were obtained from Cell Signaling Technology
(Beverly, MA, USA). An ECL Western Blotting Detection system was
obtained from GE Healthcare (Buckinghamshire, UK). Other materials
and chemicals were obtained from commercial sources.
PGE2 was dissolved in ethanol. Resveratrol, SRT1720,
PD98059, SB203580 and SP600125 were dissolved in dimethyl
sulfoxide. The maximum concentration of ethanol or dimethyl
sulfoxide was 0.1%, which did not affect either the assay for OPG
or the detection of the protein level using western blot
analysis.
Cell culture
Cloned osteoblast-like MC3T3-E1 cells derived from
newborn mouse calvaria (15) were
generously provided by Dr M. Kumegawa (Meikai University, Sakado,
Japan) and were maintained as previously described (16). Briefly, the cells were cultured in
α-minimum essential medium (α-MEM) containing 10% fetal bovine
serum (FBS) at 37°C in a humidified atmosphere of 5%
CO2/95% air. The cells were seeded into 35-mm diameter
dishes (5×104 cells/dish) or 90-mm diameter dishes
(2×105 cells/dish) in α-MEM containing 10% FBS. After 5
days, the medium was exchanged for α-MEM containing 0.3% FBS. The
cells were used for experiments after 48 h.
Assay for OPG
The cultured cells were pre-treated with various
concentrations of resveratrol, SRT1720, PD98059, SB203580 or
SP600125 for 60 min, and then stimulated by 10 μM of
PGE2 or the vehicle [mast cell medium (MCM); pH 7.4, 150
mM NaCl, 5 mM KCl, 5.5 mM glucose, 0.8 mM MgSO4, 1 mM CaCl2, 5 mM
HEPES, 0.1% bovine serum albumin] in 1 ml of α-MEM containing 0.3%
FBS for the indicated periods of time. The conditioned medium was
collected at the end of the incubation period, and the OPG
concentration was then measured using the OPG ELISA kit according
to the manufacturer’s instructions.
Real-time RT-PCR
The cultured cells were pre-treated with 50 μM of
resveratrol or the vehicle for 60 min and were then stimulated with
10 μM of PGE2 or the vehicle in α-MEM containing 0.3%
FBS for 3 h. Total RNA was isolated and reverse transcribed into
complementary DNA (cDNA) using TRIzol reagent (Invitrogen,
Carlsbad, CA, USA) and the Omniscript Reverse Transcriptase kit
(Qiagen Inc., Valencia, CA, USA). Real-time RT-PCR was performed
using LightCycler® Capillaries and FastStart DNA Master
SYBR-Green I provided with the kit (Roche Diagnostics, Basel,
Switzerland). Sense and antisense primers for mouse OPG mRNA or
GAPDH mRNA were purchased from Takara Bio Inc. (Tokyo, Japan)
(primer set ID: MA026526). The amplified products were determined
using a melting curve analysis and agarose electrophoresis. The OPG
mRNA levels were normalized to those of GAPDH mRNA.
Western blot analysis
The cultured cells were pre-treated with various
concentrations of resveratrol or SRT1720 for 60 min and were then
stimulated with PGE2 or the vehicle in α-MEM containing
0.3% FBS for the indicated periods of time. The cells were washed
twice with phosphate-buffered saline and then lysed, homogenized
and sonicated in lysis buffer containing 62.5 mM Tris/HCl, pH 6.8,
2% sodium dodecyl sulfate (SDS), 50 mM dithiothreitol and 10%
glycerol. SDS-polyacrylamide gel electrophoresis (PAGE) was
performed according to the method of Laemmli (17) in 10% polyacrylamide gels. The
protein was fractionated and transferred onto
Immun-Blot® polyvinylidene difluoride (PVDF) membranes
(Bio-Rad Laboratories, Hercules, CA, USA). The membranes were
blocked with 5% fat-free dry milk in Tris-buffered saline with
Tween-20 (TBST; 20 mM Tris-HCl, pH 7.6, 137 mM NaCl, 0.1% Tween-20)
for 2 h prior to incubation with primary antibodies. Western blot
analysis was performed as previously described (18) using phospho-specific p44/p42 MAP
kinase antibodies, p44/p42 MAP kinase antibodies, phospho-specific
p38 MAP kinase antibodies, p38 MAP kinase antibodies,
phospho-specific SAPK/JNK antibodies or SAPK/JNK antibodies as
primary antibodies, with peroxidase-labeled antibodies raised in
goat against rabbit IgG being used as secondary antibodies. The
primary and secondary antibodies were diluted at 1:1,000 with 5%
fat-free dry milk in TBST. The peroxidase activity on the PVDF
membranes was visualized on X-ray film using the ECL Western
Blotting Detection system.
Densitometric analysis
Densitometric analysis was performed using a scanner
and image analysis software (ImageJ software version 1.47). The
phosphorylated protein levels were calculated as follows: the
background-subtracted signal intensity of each phosphorylation
signal was respectively normalized to the total protein signal and
plotted as an absolute value.
Statistical analysis
All the data were analyzed by ANOVA followed by the
Bonferroni method for multiple comparisons between pairs, and a
value of P<0.05 was considered to indicate a statistically
significant difference. All data are presented as the means ±
standard error of the mean (SEM) of triplicate determinations
obtained from 3 independent cell preparations.
Results
Effects of resveratrol on the
PGE2-stimulated release of OPG in MC3T3-E1 cells
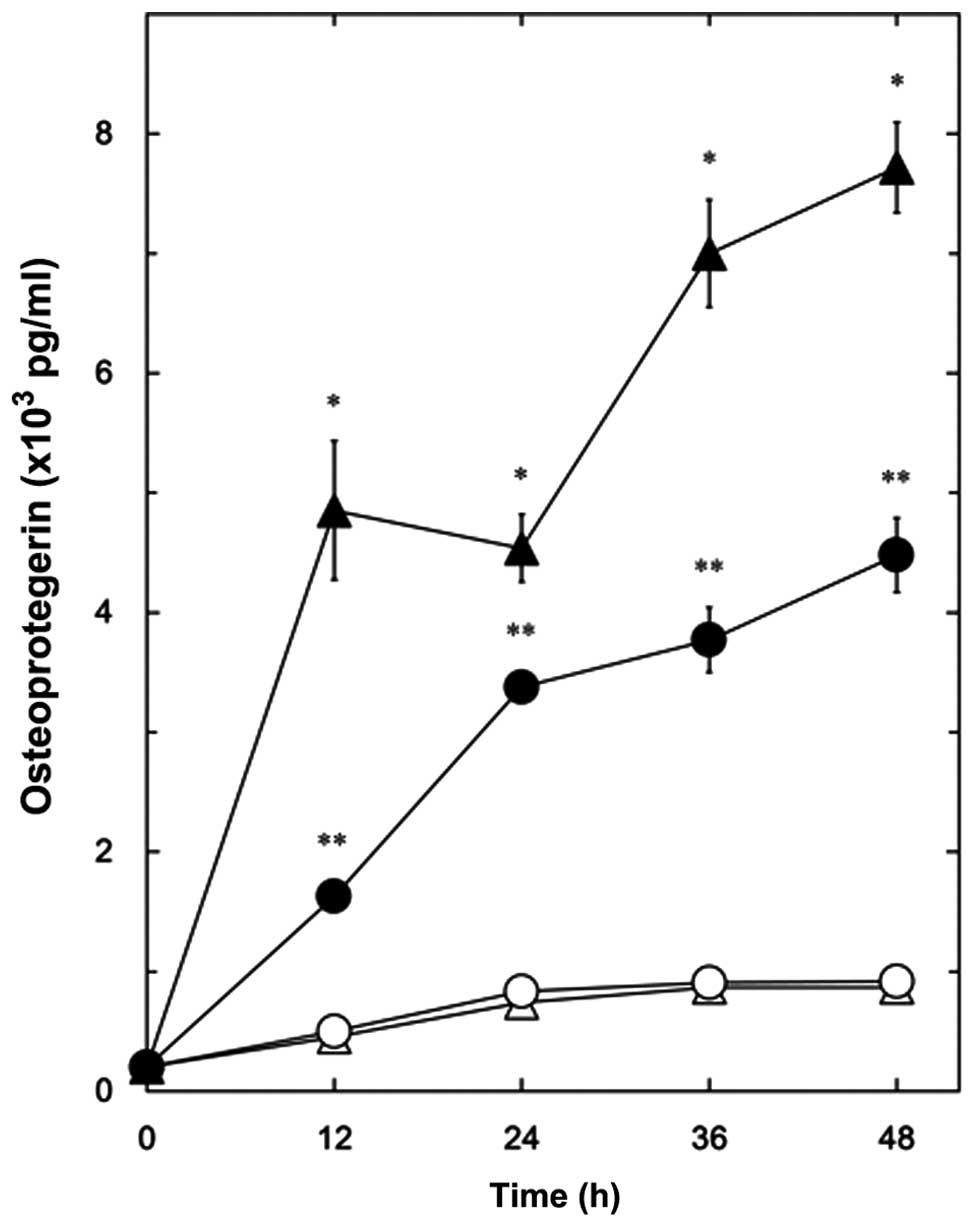
First we examined whether PGE2 stimulates
the release of OPG in osteoblast-like MC3T3-E1 cells.
PGE2 significantly promoted the release of OPG in a
time-dependent manner up to 48 h (Fig. 1). We then investigated the effects
of resveratrol on the PGE2-stimulated release of OPG in
these cells. Resveratrol, which alone did not affect the OPG levels
compared with the vehicle, significantly suppressed the
PGE2-stimulated release of OPG (Fig. 1). The inhibitory effects of
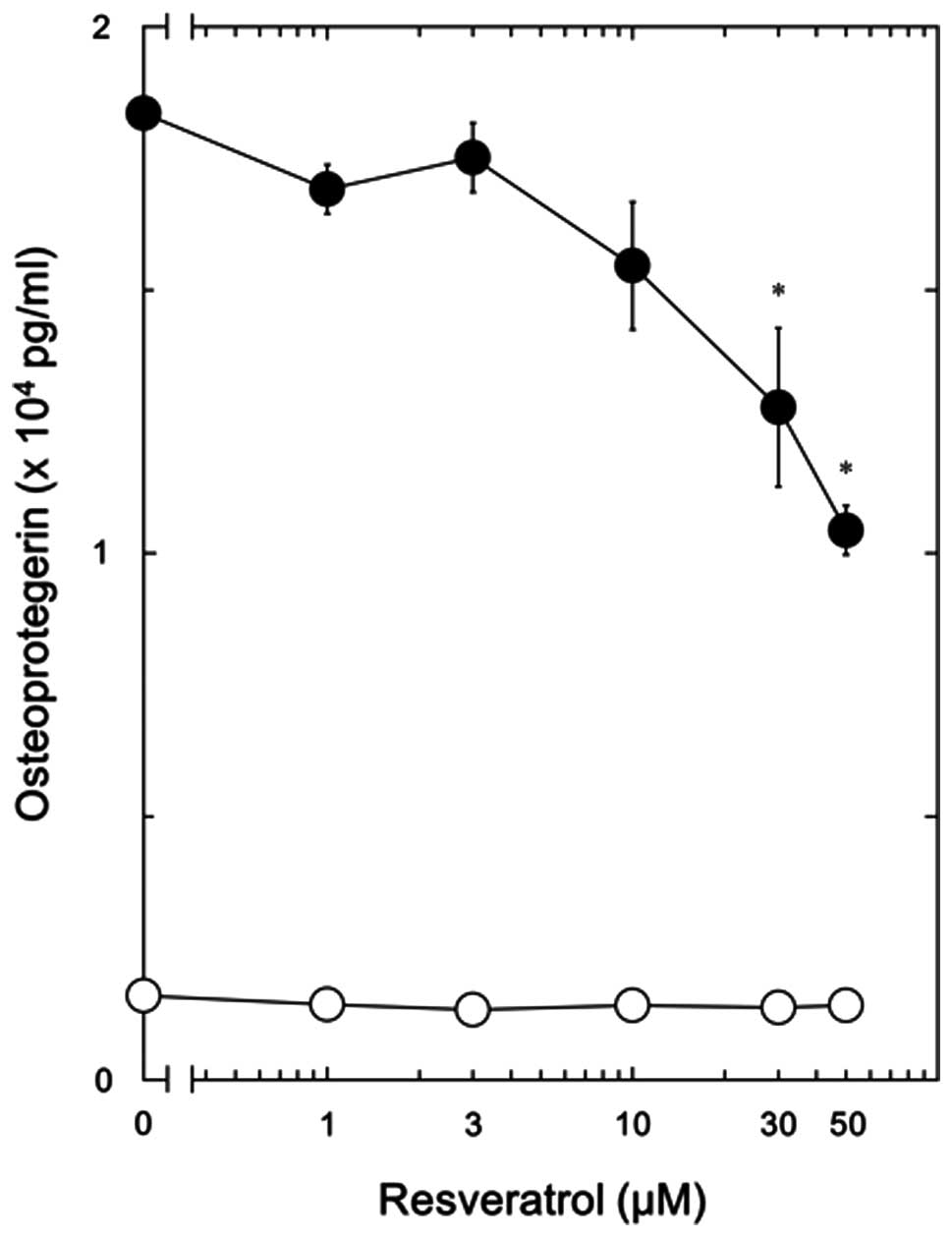
resveratrol were dose-dependent in the dose range between 1 and 50
μM (Fig. 2). The most prominent
effects of resveratrol were observed at the dose of 50 μM, which
inhibited the effects of PGE2 by approximately 50%.
Effects of SRT1720 on the
PGE2-stimulated release of OPG in MC3T3-E1 cells
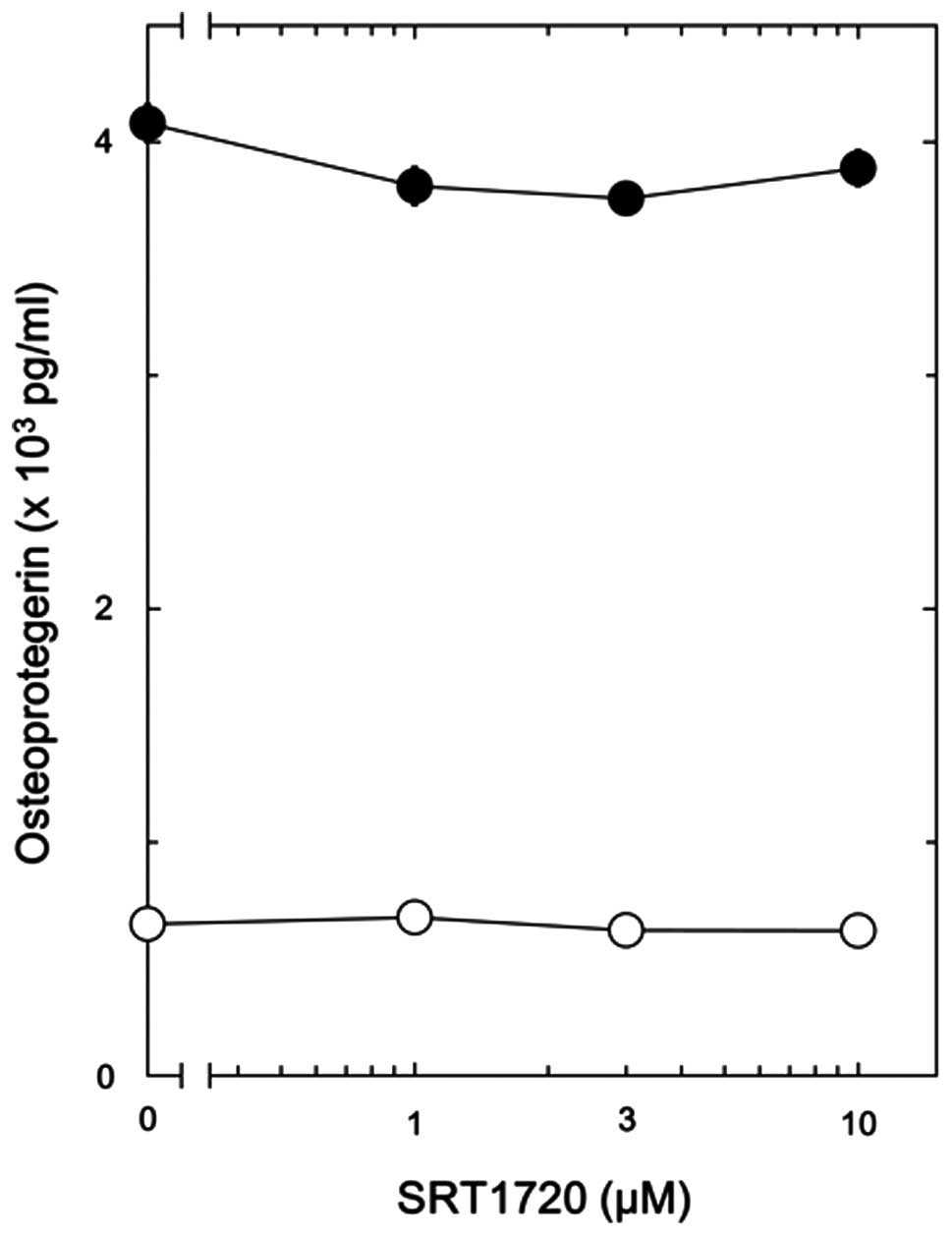
SRT1720 is known as an activator of SIRT1 with a
potency 1,000-fold greater than that of resveratrol (19). To investigate whether the effects
of resveratrol on the PGE2-stimulated release of OPG are
mediated by the activation of SIRT1, we examined the effects of
SRT1720 on the PGE2-stimulated release of OPG in the
osteoblast-like MC3T3-E1 cells. SRT1720 failed to affect the
PGE2-stimulated release of OPG from these cells
(Fig. 3).
Effects of resveratrol on the
PGE2-induced mRNA expression of OPG in MC3T3-E1
cells
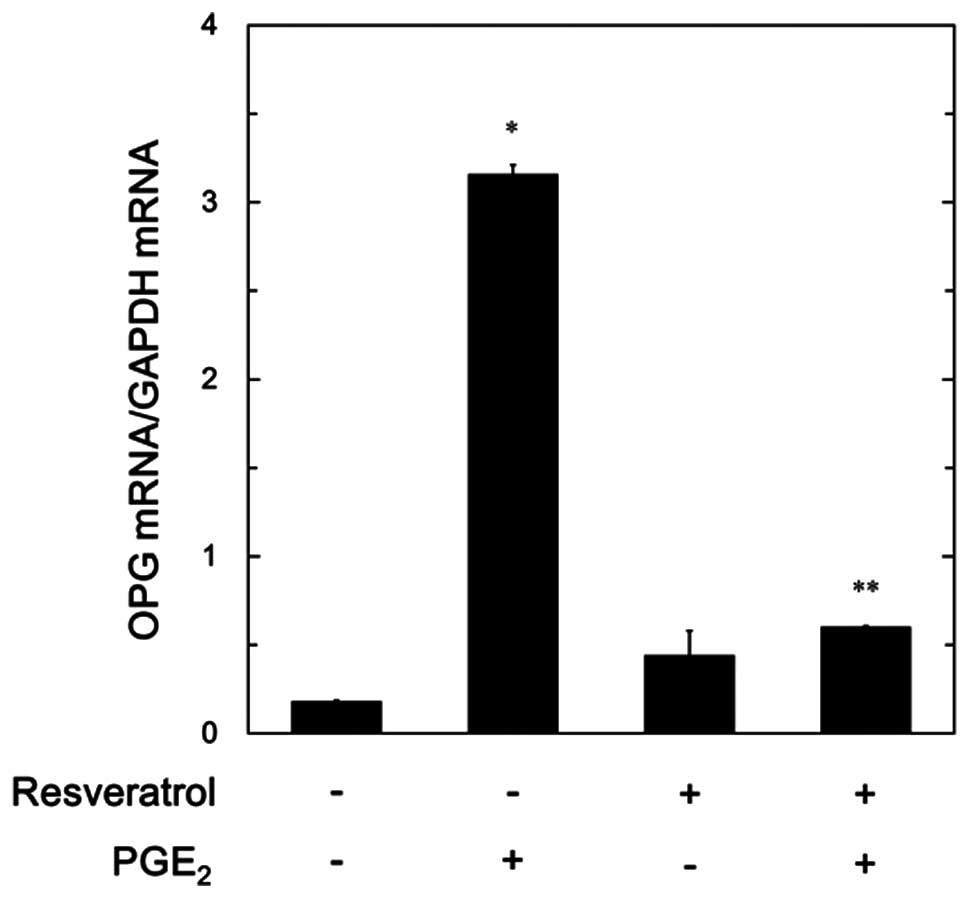
In order to elucidate whether the suppressive
effects of resveratrol on the PGE2-stimulated release of
OPG are mediated through transcriptional events, we further
examined the effects of resveratrol on the PGE2-induced
mRNA expression of OPG by real-time RT-PCR. Resveratrol, which on
its own exerted minimal effects on the mRNA levels of OPG, markedly
decreased the PGE2-induced mRNA expression levels of OPG
(Fig. 4).
Effects of PD98059, SB203580 or SP600125
on the PGE2-stimulated release of OPG in MC3T3-E1
cells
It is firmly established that the major MAP kinase
superfamily, including p44/p42 MAP kinase, p38 MAP kinase and
SAPK/JNK, is a central element used by mammalian cells to transduce
various messages of extracellular stimuli (20). As regards PGE2
intracellular signaling in osteoblasts, we have previously reported
that p44/p42 MAP kinase and p38 MAP kinase are involved in the
HSP27 induction by PGE2 in osteoblast-like MC3T3-E1
cells (10). In order to clarify
whether p44/p42 MAP kinase, p38 MAP kinase or SAPK/JNK are involved
in the PGE2-stimulated OPG synthesis in osteoblast-like
MC3T3-E1 cells, we examined the effects of PD98059, a specific
inhibitor of the upstream kinase activating p44/p42 MAP kinase
(21), SB203580, a specific
inhibitor of p38 MAP kinase (22)
or SP600125, a specific inhibitor of SAPK/JNK (23), on the release of OPG stimulated by
PGE2. PD98059, SB203580 and SP600125, which alone had
little effect on the release of OPG, markedly reduced the
PGE2-stimulated release of OPG in these cells (Table I).
 | Table IEffects of PD98059, SB203580 or
SP600125 on the PGE2-stimulated release of OPG in
MC3T3-E1 cells. |
Table I
Effects of PD98059, SB203580 or
SP600125 on the PGE2-stimulated release of OPG in
MC3T3-E1 cells.
| Inhibitors |
PGE2 | OPG (pg/ml) |
|---|
| - | − | 1,128±28 |
| - | + | 11,424±857a |
| PD98059 | − | 1,310±106 |
| PD98059 | + | 7,018±510b |
| SB203580 | − | 982±1 |
| SB203580 | + | 5,809±65b |
| SP600125 | − | 1,176±97 |
| SP600125 | + | 6,508±827b |
Effects of resveratrol or SRT1720 on the
PGE2-induced phosphorylation of p44/p42 MAP kinase, p38
MAP kinase or SAPK/JNK in MC3T3-E1 cells
To investigate whether resveratrol affects the
PGE2-stimulated OPG synthesis through the activation of
p44/p42 MAP kinase, p38 MAP kinase or SAPK/JNK in osteoblast-like
MC3T3-E1 cells, we examined the effects of resveratrol on the
PGE2-induced phosphorylation of p44/p42 MAP kinase, p38
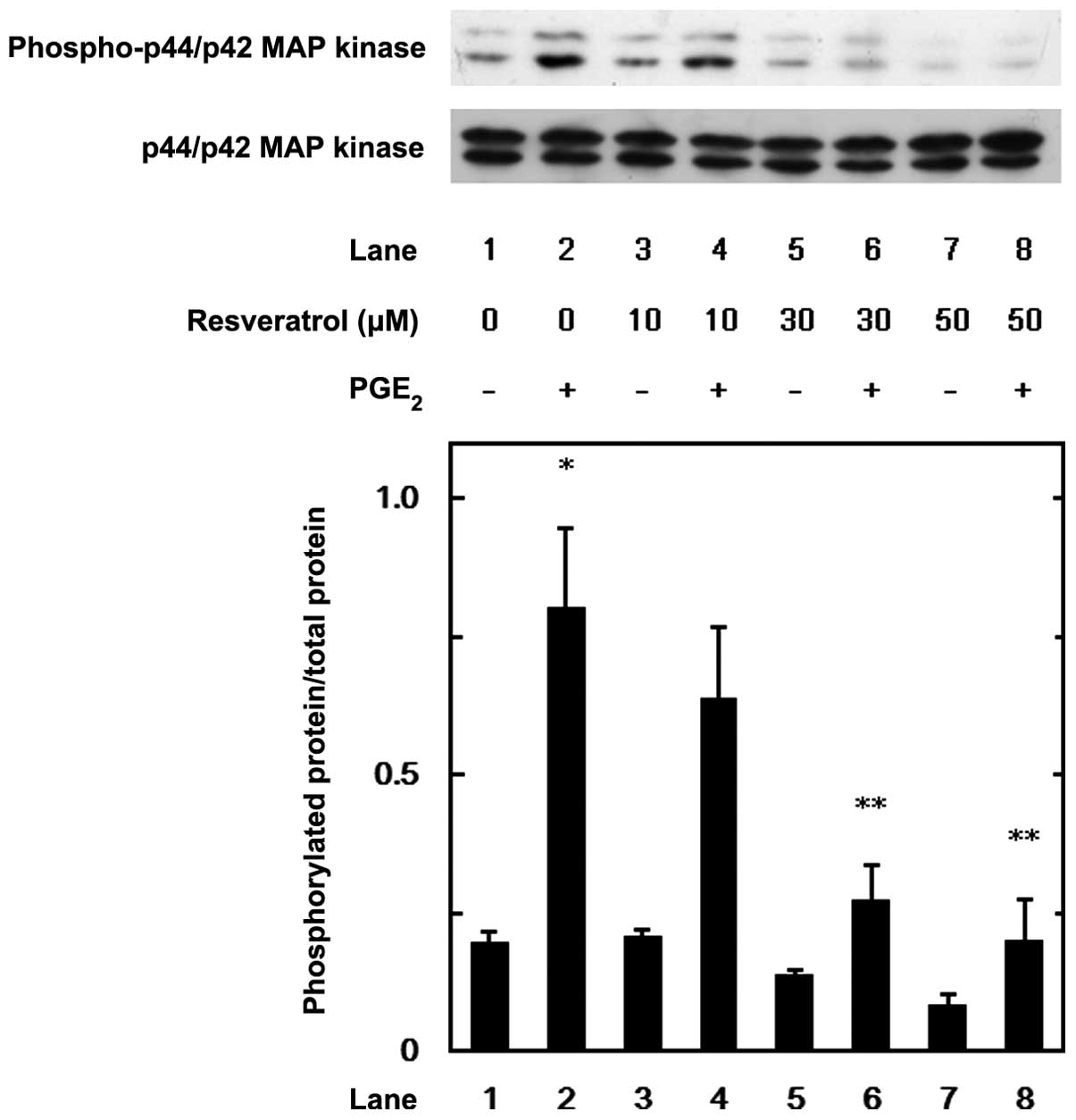
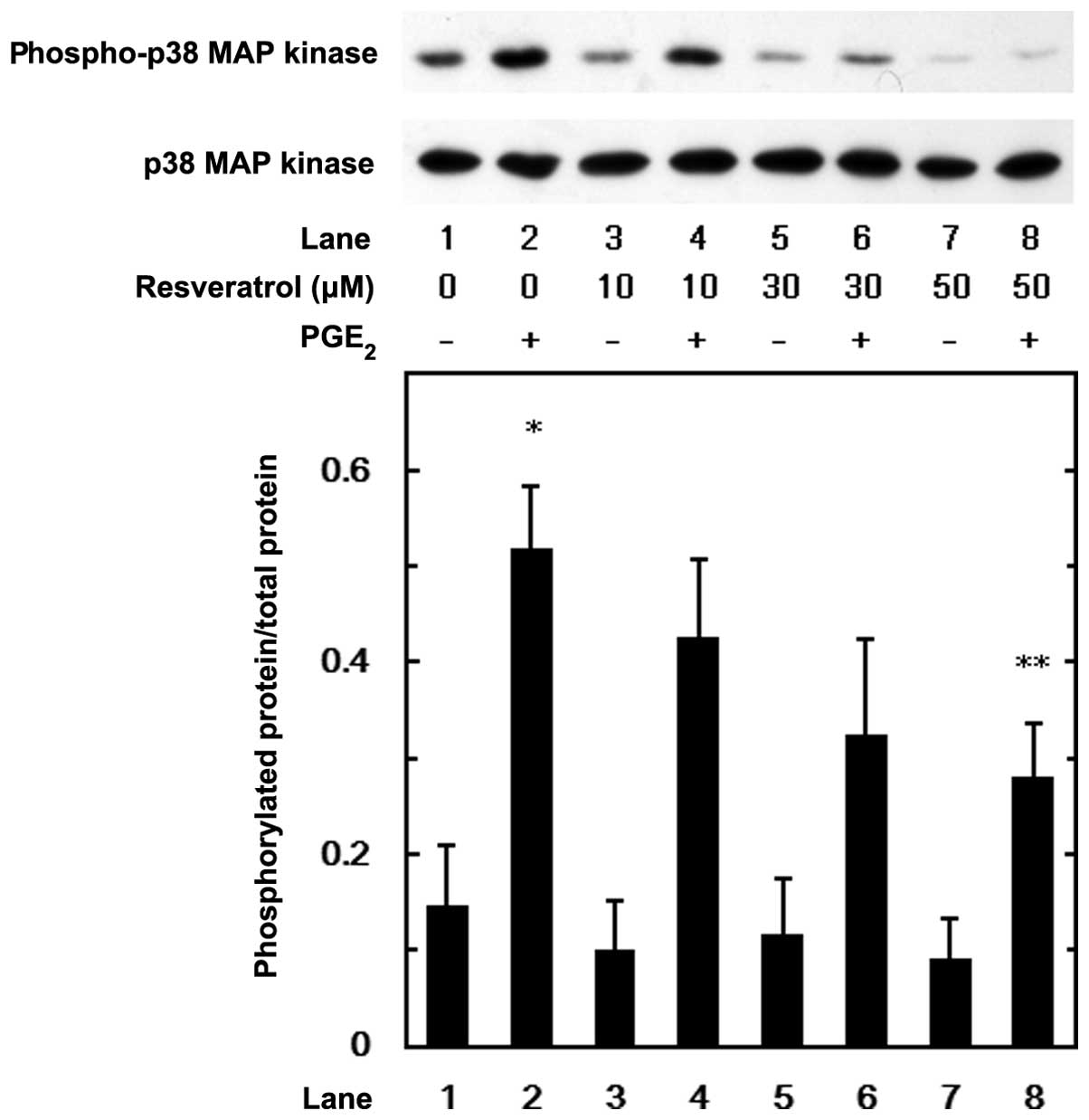
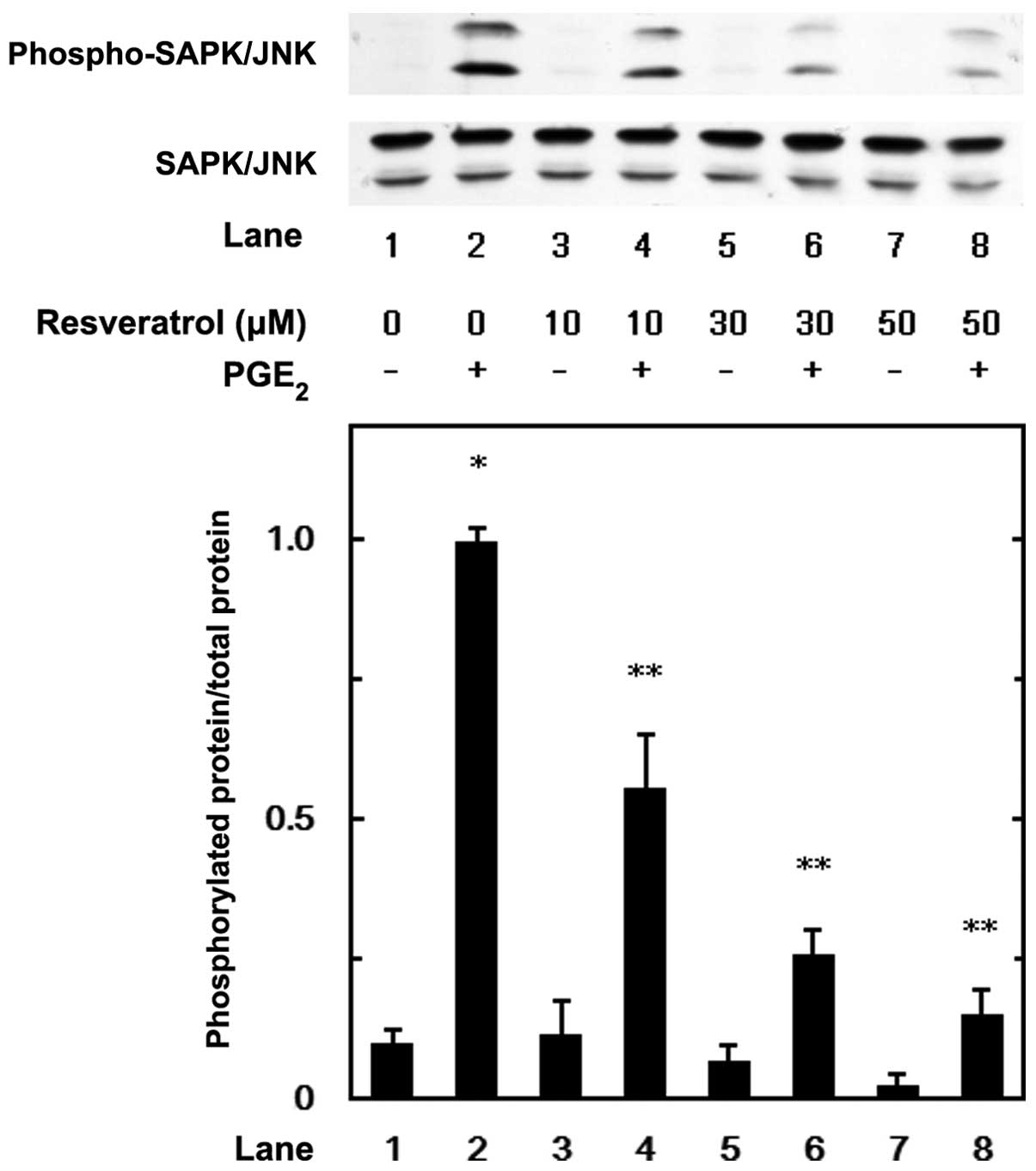
MAP kinase or SAPK/JNK. Resveratrol suppressed the
PGE2-induced phosphorylation of p44/p42 MAP kinase
(Fig. 5), p38 MAP kinase
(Fig. 6) and SAPK/JNK (Fig. 7) in a dose-dependent manner at the
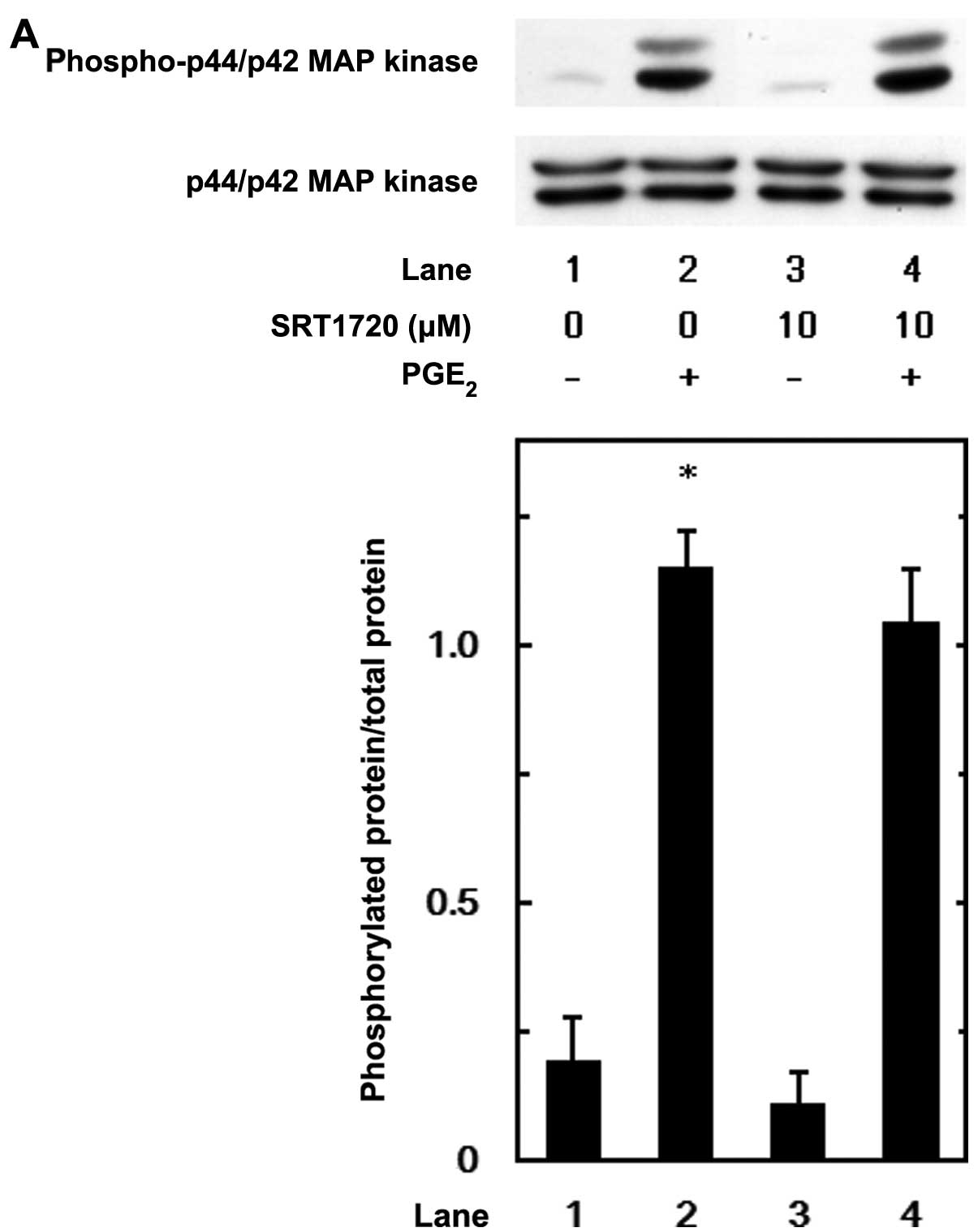
dose range between 10 and 50 μM. We further examined the effects of
SRT1720 on the PGE2-induced phosphorylation of p44/p42
MAP kinase, p38 MAP kinase or SAPK/JNK. However, SRT1720 hardly
affected the PGE2-induced phosphorylation of p44/p42 MAP
kinase, p38 MAP kinase or SAPK/JNK (Fig. 8).
Discussion
In the present study, we demonstrated that
PGE2 significantly stimulated the release of OPG in
osteoblast-like MC3T3-E1 cells and that resveratrol markedly
suppressed the PGE2-stimulated release of OPG.
Additionally, we demonstrated that PGE2 upregulated the
mRNA expression levels of OPG, and that resveratrol reduced the
mRNA expression of OPG induced by PGE2 in these cells.
Therefore, these findings suggest that the suppressive effects of
resveratrol on the PGE2-stimulated release of OPG are
mediated through transcriptional events in MC3T3-E1 cells. Thus, we
further investigated the exact mechanisms behind the inhibitory
effects of resveratrol on the PGE2-stimulated OPG
synthesis in osteoblasts.
The three major MAP kinases, p44/p42 MAP kinase, p38
MAP kinase and SAPK/JNK are recognized as central elements used by
mammalian cells to transduce diverse messages (24) and play central roles in a variety
of cellular functions, including proliferation, differentiation and
survival (20). As regards the
intracellular signaling of PGE2 in osteoblasts, we have
previously demonstrated that PGE2 induces the activation
of p44/p42 MAP kinase and p38 MAP kinase in osteoblast-like
MC3T3-E1 cells, and that PGE2 stimulates the induction
of HSP27 through the PKC-dependent activation of both p44/p42 MAP
kinase and p38 MAP kinase in MC3T3-E1 cells (10). In the present study, we found that
PGE2 stimulated the phosphorylation of SAPK/JNK in the
MC3T3-E1 cells in a time-dependent manner and the most prominent
effects of PGE2 were observed at 20 min following
stimulation (data not shown). It is generally established that MAP
kinases are activated by the phosphorylation of threonine and
tyrosine residues by dual-specificity MAP kinase kinase (24). Therefore, our findings suggest
that PGE2 stimulates the activation of SAPK/JNK in
addition to that of p44/p42 MAP kinase and p38 MAP kinase in
osteoblast-like MC3T3-E1 cells. Furthermore, we demonstrated that
PD98059, a specific inhibitor of the upstream kinase activating
p44/p42 MAP kinase (21),
SB203580, a specific inhibitor of p38 MAP kinase (22) and SP600125, a specific inhibitor
of SAPK/JNK (23) markedly
reduced the PGE2-stimulated release of OPG, suggesting
that three major MAP kinases, namely the p44/p42 MAP kinase, p38
MAP kinase and SAPK/JNK function as positive regulators in the
PGE2-stimulated OPG synthesis in these cells. In
addition, we demonstrated that resveratrol markedly suppressed the
phosphorylation of p44/p42 MAP kinase, p38 MAP kinase and SAPK/JNK
induced by PGE2 in the MC3T3-E1 cells. Taking our
findings into account, it is likely that resveratrol inhibits the
PGE2-induced OPG synthesis in osteoblast-like cells, and
that the suppressive effects of resveratrol are exerted at a point
upstream of three MAP kinases.
Evidence is accumulating that the beneficial effects
of resveratrol are mediated through SIRT1 activation (13). On the other hand, we demonstrated
that SRT1720, a potent activator of SIRT1 (19), exerted minimal effects on the
PGE2-stimulated release of OPG and on the
phosphorylation of the three MAP kinases induced by PGE2
in osteoblast-like MC3T3-E1 cells. Therefore, it seems unlikely
that the inhibitory effects of resveratrol on the
PGE2-induced events shown in our study are
SIRT1-dependent in these cells. In a previous study of ours
(14), we demonstrated that
resveratrol significantly suppressed the BMP-4-induced VEGF
synthesis, and that the effects were mediated at least in part by
the activation of SIRT1 in MC3T3-E1 cells. Based on our findings,
it is possible that the intracellular signaling mechanisms
underlying the effects of resveratrol in osteoblasts are
characterized according to each agonist or product. It has been
reported that resveratrol affects the cellular function of
adipocytes and regulates the number of fat cells in a
SIRT1-independent manner (25,26). Since both osteoblasts and
adipocytes originate from undifferentiated mesenchymal stem cells,
the SIRT-independent effects of resveratrol observed in osteoblasts
may be due to these same stem cells.
It is recognized that the upregulation of RANKL is
an essential step for the action of bone resorptive agents,
including PGE2 and promoting osteoclast formation
(8). Although OPG plays a
suppressive role in osteoclastogenesis as a decoy receptor of
RANKL, it seems that the OPG suppression of the acceleration of
bone resorption may be the initial signal to promote the bone
remodeling required for maintaining the quality of bone. The
sophisticated regulation of bone remodeling is important to
maintain the quantity and quality of bone, and proper osteoclastic
bone resorption is essential to bone turnover that involves the
removal of old fragile bone and its replacement. Therefore, the
effects of resveratrol shown in our study may provide a new aspect
of the regulatory mechanisms of bone metabolism. Further studies
are required to clarify the exact mechanisms behind the effects of
resveratrol on osteoblasts.
In conclusion, our results strongly suggest that
resveratrol reduces the PGE2-stimulated OPG synthesis
through the inhibition of p44/p42 MAP kinase, p38 MAP kinase and
SAPK/JNK in osteoblasts, and that these suppressive effects are not
mediated through the activation of SIRT1.
Acknowledgements
We are very grateful to Yumiko Kurokawa for her
skillful technical assistance. This investigation was supported in
part by Grants-in-Aid for Scientific Research (19591042) from the
Ministry of Education, Science, Sports and Culture of Japan, the
Foundation for Growth Science, the Research Funding for Longevity
Sciences (23-9, 25-4) and the National Center for Geriatrics and
Gerontology (NCGG), Japan.
References
|
1
|
Karsenty G and Wagner EF: Reaching a
genetic and molecular understanding of skeletal development. Dev
Cell. 2:389–406. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Parfitt AM: Targeted and nontargeted bone
remodeling: relationship to basic multicellular unit origination
and progression. Bone. 30:5–7. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Simonet WS, Lacey DL, Dunstan CR, Kelley
M, Chang MS, Lüthy R, Nguyen HQ, Wooden S, Bennett L, Boone T,
Shimamoto G, DeRose M, Elliott R, Colombero A, Tan HL, Trail G,
Sullivan J, Davy E, Bucay N, Renshaw-Gegg L, Hughes TM, Hill D,
Pattison W, Campbell P, Sander S, Van G, Tarpley J, Derby P, Lee R
and Boyle WJ: Osteoprotegerin: a novel secreted protein involved in
the regulation of bone density. Cell. 89:309–319. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Udagawa N, Takahashi N, Yasuda H, Mizuno
A, Itoh K, Ueno Y, Shinki T, Gillespie MT, Martin TJ, Higashio K
and Suda T: Osteoprotegerin produced by osteoblasts is an important
regulator in osteoclast development and function. Endocrinology.
141:3478–3484. 2000.PubMed/NCBI
|
|
5
|
Kwan TS, Padrines M, Théoleyre S, Heymann
D and Fortun Y: IL-6, RANKL, TNF-α/IL-1: interrelations in bone
resorption pathophysiology. Cytokine Growth Factor Rev. 15:49–60.
2004.
|
|
6
|
Nijiweide PJ, Burger EH and Feyen JH:
Cells of bone: proliferation, differentiation, and hormonal
regulation. Physiol Rev. 66:855–886. 1986.PubMed/NCBI
|
|
7
|
Smith WL: The eicosanoids and their
biochemical mechanisms of action. Biochem J. 259:315–324.
1989.PubMed/NCBI
|
|
8
|
Blackwell KA, Raisz LG and Pilbeam CC:
Prostaglandins in bone: bad cop, good cop? Trends Endocrinol Metab.
21:294–301. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Kozawa O, Suzuki A, Tokuda H, Kaida T and
Uematsu T: Interleukin-6 synthesis induced by prostaglandin
E2: cross-talk regulation by protein kinase C. Bone.
22:355–360. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Tokuda H, Kozawa O, Niwa M, Matsuno H,
Kato K and Uematsu T: Mechanism of prostaglandin
E2-stimulated heat shock protein 27 induction in
osteoblasts-like MC3T3-E1 cells. J Endocrinol. 172:271–281.
2002.
|
|
11
|
Frémont L: Biological effects of
resveratrol. Life Sci. 66:663–673. 2000.
|
|
12
|
Baur JA, Pearson KJ, Price NL, Jamieson
HA, Lerin C, Kalra A, Prabhu VV, Allard JS, Lopez-Lluch G, Lewis K,
Pistell PJ, Poosala S, Becker KG, Boss O, Gwinn D, Wang M,
Ramaswamy S, Fishbein KW, Spencer RG, Lakatta EG, Le Couteur D,
Shaw RJ, Navas P, Puigserver P, Ingram DK, de Cabo R and Sinclair
DA: Resveratrol improves health and survival of mice on a
high-calorie diet. Nature. 444:337–342. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Timmers S, Auwerx J and Schrauwen P: The
journey of resveratrol from yeast to human. Aging. 4:146–158.
2012.PubMed/NCBI
|
|
14
|
Kondo A, Otsuka T, Kuroyanagi G, Yamamoto
N, Matsushima-Nishiwaki R, Mizutani J, Kozawa O and Tokuda H:
Resveratrol inhibits BMP-4-stimulated VEGF synthesis in
osteoblasts: Suppression of S6 kinase. Int J Mol Med. 33:1013–1018.
2014.PubMed/NCBI
|
|
15
|
Sudo H, Kodama H, Amagai Y, Yamamoto S and
Kasai S: In vivo differentiation and calcification in a new clonal
osteogenic cell line derived from newborn mouse calvaria. J Cell
Biol. 96:191–198. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kozawa O, Tokuda H, Miwa M, Kotoyori J and
Oiso Y: Cross-talk regulation between cyclic AMP production and
phosphoinositide hydrolysis induced by prostaglandin E2
in osteoblast-like cells. Exp Cell Res. 198:130–134. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Laemmli UK: Cleavage of structural
proteins during the assembly of the head of bacteriophage T4.
Nature. 227:680–685. 1970. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Kato K, Ito H, Hasegawa K, Inaguma Y,
Kozawa O and Asano T: Modulation of the stress-induced synthesis of
hsp27 and αB-crystallin by cyclic AMP in C6 glioma cells. J
Neurochem. 66:946–950. 1996.
|
|
19
|
Milne JC, Lambert PD, Schenk S, Carney DP,
Smith JJ and Gagne DJ: Small molecule activators of SIRT1 as
therapeutics for the treatment of type 2 diabetes. Nature.
450:712–716. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kyriakis JM and Avruch J: Mammalian
mitogen-activated protein kinase signal transduction pathways
activated by stress and inflammation. Physiol Rev. 81:807–869.
2001.PubMed/NCBI
|
|
21
|
Alessi DR: PD98059 is a specific inhibitor
of the activation of mitogen-activated protein kinase in vitro and
in vivo. J Biol Chem. 270:27489–27494. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Cuenda A, Rouse J, Doza YN, Meier R, Cohen
P, Gallagher TF, Young PR and Lee JC: SB203580 is a specific
inhibitor of a MAP kinase homologue which is stimulated by cellular
stresses and interleukin-1. FEBS Lett. 364:229–233. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bennett BL, Sasaki DT, Murray BW, O’Leary
EC, Sakata ST, Xu W, Leisten JC, Motiwala L, Peierce S, Satoh Y,
Bhagwat SS, Manning SM and Anderson DW: SP600125, an
anthrapyrazolone inhibitor of Jun N-terminal Kinase. Proc Natl Acad
Sci USA. 98:13681–13686. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Widmann C, Gibson S, Jarpe MB and Johnson
GL: Mitogen-activated protein kinase: conservation of a
three-kinase module from yeast to human. Physiol Rev. 79:143–180.
1999.PubMed/NCBI
|
|
25
|
Mader I, Wabitsch M, Debatin KM,
Fischer-Posovsky P and Fulda S: Identification of a novel
proapoptotic function of resveratrol in fat cells:
SIRT1-independent sensitization to TRAIL-induced apoptosis. FASEB
J. 24:1997–2009. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Wang A, Liu M, Liu X, Dong LQ, Glickman
RD, Slaga TJ, Zhou Z and Liu F: Up-regulation of adiponectin by
resveratrol: the essential roles of the Akt/FOXO1 and AMP-activated
protein kinase signaling pathways and DsbA-L. J Biol Chem.
286:60–66. 2011. View Article : Google Scholar : PubMed/NCBI
|