Introduction
Atherosclerosis is a chronic disease that remains
asymptomatic for decades (1). It
is caused by the formation of multiple plaques within the arteries.
Attention has been focused on the ‘vulnerable plaque’ since the
late 1990s onwards (2).
Atherosclerosis can lead to ischemic heart disease, cerebrovascular
accidents and peripheral vascular diseases (3). Carotid intima-media thickness (cIMT)
level is known as a surrogate marker of atherosclerosis (4). A special type of carotid
atherosclerosis with CagA-positive Helicobacter pylori
(CagA+ HP) infection is common in China. An increased
serum YKL-40 level suggests plaque instability and more severe
clinical symptoms of carotid atherosclerosis with CagA+
HP infection (5). Inflammatory
cytokines induced by VEGF, such as monocyte chemoattractant protein
(MCP-1), have been previously shown to be involved in the
pathogenesis and progression of carotid atherosclerosis (6).
Metabolic syndrome may be independently associated
with the early stage but not the later and advanced stages of
carotid atherosclerosis in community residents in China (7). An in vivo 3T MRI study was
previously conducted to determine the effect of gender differences
of high-risk carotid atherosclerotic plaque with <50% stenosis
in asymptomatic patients (8).
Evaluation of carotid atherosclerosis was therefore performed from
the perspective of blood flow reflection (9). Results of a multivariate analysis
revealed plaque number by ultrasonography (P=0.023), age (P=0.001),
calcium-phosphate product (P=0.049) and serum albumin (P=0.009) as
independent risk factors (10).
Higher brachial-ankle pulse wave velocity was identified as a risk
factor for carotid atherosclerosis in patients with end-stage renal
disease (11). The relationship
between levels of circulating intercellular cell-adhesion
molecule-1 (cICAM-1) or P-selectin (cP-selectin) and the severity
of carotid atherosclerosis was also examined (12). The findings of that study showed
that cP-selectin did not increase until atherosclerosis was at an
advanced stage (12).
Inactivation of the PDZK1 gene is known to promote
the development of aortic root atherosclerosis in apolipoprotein E
(apoE) KO mice fed with a high fat/high cholesterol diet (13). Additionally, SERPINA1 was found to
be upregulated in atherosclerotic plaques (14). Angiogenesis, the process of new
capillary formation from existing blood vessels, is dysregulated in
many pathological disorders including atherosclerosis (15). In CD68+ cells from
regressing plaque of atherosclerosis, Feig et al (16) observed that genes related to cell
adhesion, such as cadherins and vinculin, were downregulated.
Transcription factors such as LDLR and TP53 are
important in the development of atherosclerosis, as identified in a
Malaysian study population (13).
It has been previously reported that miRNAs are associated with
atherosclerosis (17). However,
the association between miRNAs and atherosclerosis has not yet been
fully elucidated.
Immune-associated biological processes can affect
atherosclerosis. Growth differentiation factor-15 deficiency
inhibits the progression of atherosclerosis by controlling the
interleukin-6-dependent inflammatory response to vascular injury
(18). Vascular smooth muscle
cell (VSMC) phenotypic modulation plays a key role in
atherosclerosis. SMCs secrete cytokines and express cell adhesion
molecules such as IL-8, IL-6 and VCAM-1 (19). Prenatal arsenic exposure alters
gene expression in the adult liver to a proinflammatory state
contributing to accelerated atherosclerosis by affecting pathways
such as antigen processing and presentation (20). Vascular smooth muscle contraction
is a main effect for artery (21). Morelloflavone, a biflavonoid and
an active ingredient of the plant, has been shown to inhibit VSMC
migration through its inhibition of multiple migration-related
kinases such as focal adhesion kinase (22).
Therefore, in the present study, we initially
detected differentially expressed genes (DEGs). Subsequently, we
identified the enriched Gene Ontology (GO) terms and pathways for
the DEGs. The transcriptional and miRNA regulatory network for the
DEGs was also constructed. Cis-regulatory signals were also
investigated.
Materials and methods
Data preprocessing
The original data were downloaded from the National
Center for Biotechnology Gene Expression Omnibus (NCBI GEO)
database for atherosclerotic plaques of carotid atherosclerosis
under accession no. GSE28829 (23), including 13 chips at early stage
(EAR, pathological, intimal thickening and intimal xanthoma) and 16
chips at advanced stage (ADV, thin or thick fibrous cap atheroma).
The chip platform was GPL570, Affymetrix Human Genome U133 Plus 2.0
Array.
Background subtraction and quantile normalization
was performed using Affymetrix Power Tools (APT) (http://www.affymetrix.com/) in the Robust Multiarray
Average (RMA) algorithm (24).
Genes with a low expression were filtered, ensuring that their
plier was ≥100 in at least 2/3 of the samples (25).
Identifying differentially expressed
genes
Criteria for a differential probe included: i)
signal strength of one probe is >1.5-fold between the two
groups; ii) t-test P-value of ≤0.01; and iii) plier ≥100 in at
least 2/3 of the samples.
Functional enrichment of differential
genes
Gene Ontology (GO) and the Kyoto Encyclopedia of
Genes and Genomes (KEGG) pathway enrichment analysis was conducted
with DAVID (26) for the
differential genes between two groups. The threshold [P-value ≤0.05
and false discovery rate (FDR) ≤0.05] was calculated using Fisher’s
Exact test.
Transcriptional regulatory network
The differential genes, especially
atherosclerosis-related genes, may be regulated by different
transcriptional factors and miRNAs in different stages. We initally
extracted the sequences located at 1k bp upstream and 200 bp
downstream of 5′UTR for the transcription start site, using JASPAR
(27) (http://jaspar.genereg.net) (the score threshold was
0.95). The core transcriptional regulatory network was constructed
for the upregulated genes, with a >4-fold change in the advanced
stage and a >2-fold change in the early stage. Upregulated genes
were regulated by transcriptional factors (TFs) in the early and
advanced stages of carotid atherosclerosis. TFs act depending on
cis-regulatory motif in 5′UTR of their target genes. In
order to detect such significant enriched cis-regulatory signals,
hypergeometric distribution test was implemented (28), with a P-value of ≤0.05.
miRNA regulatory network
The 1k bp 3′UTR sequence was extracted from the
differentially expressed genes. Although the binding of miRNA and
its targets in animals are not as conservative as in plant, while
binding is relatively conservative in the seed region. RNAhybrid is
based conservation in seed region, which makes it more suitable for
animal miRNA target gene prediction. RNAhybrid (29) was subsequently used to identify
miRNA binding sites (P-value ≤0.01).
Results
Differentially expressed genes
In the present study, the early stage sample was
considered as the control group. We identified 889 differential
probes (corresponding to 707 genes), of which 413 probes (322
genes) were downregulated, and 476 probes (385 genes) were
upregulated in advanced stage samples.
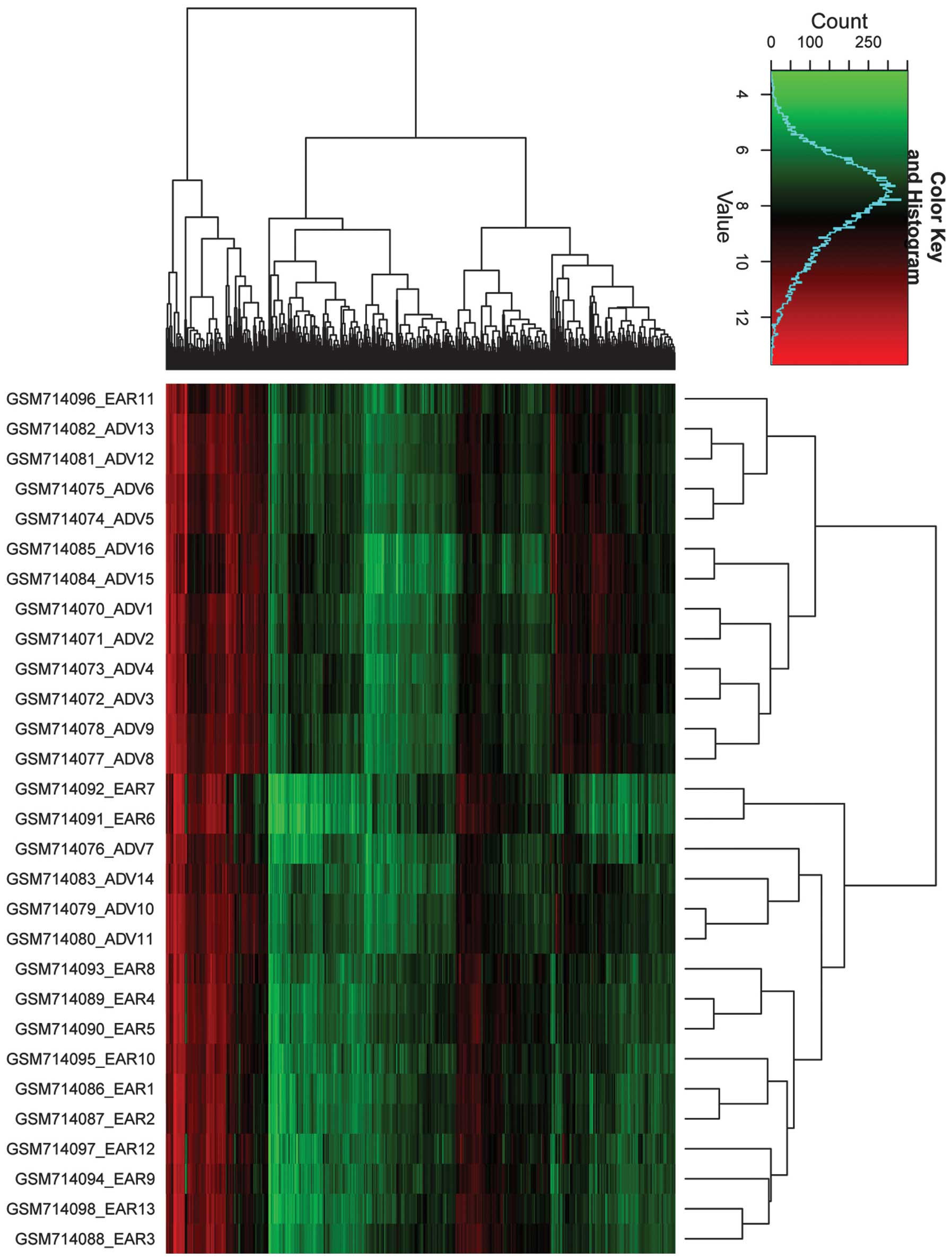
The 889 differential probes were clustered using R
hclust based on RMA log2-transformed values (Fig. 1). Thirteen samples were clustered
into one group, with only one early stage sample being
mis-clustered in this group. The standard error of the mean
distribution is shown in Fig. 2.
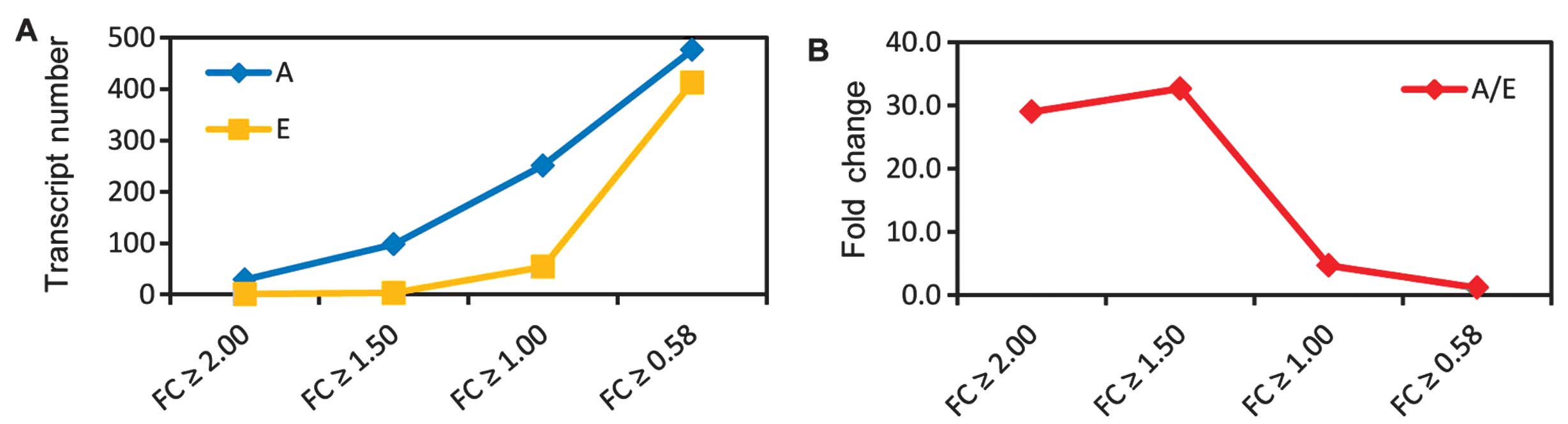
Fig. 3 shows the up- and
downregulation for the 889 differential probes (corresponding to
708 genes). As the absolute log fold change (|logFC|) becomes
larger, the number of upregulated genes were reduced (Fig. 3A). However, the fold change
increased as the |logFC| became larger (Fig. 3B). For example, when |logFC|≥1.5,
the fold change was 0.58, whereas when |logFC|≥1, the fold change
was 1. This finding suggested that a large number of genes were
activated in the advanced stage.
Enriched functional terms of differential
genes
The downregulated genes were enriched in biological
processes including cytoskeleton organization, cell adhesion,
muscle organ development, regulation of muscle contraction,
regulation of cell growth, regulation of system process, regulation
of metal ion transport, heart development, muscle contraction,
muscle cell differentiation, negative regulation of cell growth,
intracellular signaling cascade, myofibril assembly, cellular metal
ion homeostasis, regulation of calcium ion transport, negative
regulation of cell size and regulation of intracellular transport
(Table I). The findings
demonstrated that the biological processes do not correlate with
atherosclerosis.
 | Table IMajor biological processes enriched
by upregulated genes in early stage. |
Table I
Major biological processes enriched
by upregulated genes in early stage.
| Term | Count | P-value | FDR |
|---|
| GO:0007010:
Cytoskeleton organization | 25 | 2.12E-07 | 3.51E-04 |
| GO:0007155: Cell
adhesion | 29 | 1.14E-05 | 1.88E-02 |
| GO:0007517: Muscle
organ development | 14 | 4.57E-05 | 7.56E-02 |
| GO:0006937:
Regulation of muscle contraction | 7 | 1.16E-03 | 1.90E+00 |
| GO:0001558:
Regulation of cell growth | 11 | 1.38E-03 | 2.25E+00 |
| GO:0044057:
Regulation of system process | 14 | 1.80E-03 | 2.93E+00 |
| GO:0010959:
Regulation of metal ion transport | 7 | 2.13E-03 | 3.46E+00 |
| GO:0007507: Heart
development | 11 | 2.92E-03 | 4.73E+00 |
The upregulated genes enriched in biological
processes associated with atherosclerosis were: i)
immune-associated GO terms, including defense response, response to
wounding, inflammatory response, immune effector process, leukocyte
mediated immunity, antigen processing and presentation of peptide
or polysaccharide antigen via the MHC class II, adaptive immune
response, activation of immune response, lymphocyte mediated
immunity, antigen processing and presentation, behavior, B
cell-mediated immunity, antigen processing and presentation of
exogenous peptide antigen, acute inflammatory response, chemotaxis,
antigen processing and presentation of exogenous antigen; ii)
vascular-related GO terms including blood vessel development, blood
vessel morphogenesis, and angiogenesis; and iii) cell movement
homeostasis including cell adhesion, regulation of cell motion,
regulation of cell migration, regulation of locomotion, cation
homeostasis and chemical homeostasis (Table II).
| Table IIMajor biological processes enriched
by upregulated genes in advanced stage. |
Table II
Major biological processes enriched
by upregulated genes in advanced stage.
| Term | Count | P-value | FDR |
|---|
| GO:0006955: Immune
response | 86 | 4.85E-40 | 8.42E-37 |
| GO:0006952: Defense
response | 65 | 1.40E-25 | 2.43E-22 |
| GO:0009611:
Response to wounding | 57 | 1.10E-22 | 1.91E-19 |
| GO:0006954:
Inflammatory response | 43 | 2.13E-20 | 3.69E-17 |
| GO:0006935:
Chemotaxis | 27 | 2.20E-15 | 3.85E-12 |
| GO:0042330:
Taxis | 27 | 2.20E-15 | 3.85E-12 |
| GO:0002252: Immune
effector process | 21 | 2.08E-11 | 3.61E-08 |
| GO:0048584:
Positive regulation of response to stimulus | 27 | 2.37E-11 | 4.11E-08 |
| GO:0007626:
Locomotory behavior | 29 | 2.40E-11 | 4.16E-08 |
| GO:0002443:
Leukocyte mediated immunity | 17 | 7.37E-11 | 1.28E-07 |
| GO:0002504: Antigen
processing and presentation of peptide or polysaccharide antigen
via MHC class II | 12 | 8.24E-11 | 1.43E-07 |
| GO:0002250:
Adaptive immune response | 16 | 1.47E-10 | 2.55E-07 |
| GO:0002253:
Activation of immune response | 17 | 2.97E-10 | 5.16E-07 |
| GO:0002449:
Lymphocyte mediated immunity | 15 | 4.22E-10 | 7.33E-07 |
| GO:0019882: Antigen
processing and presentation | 16 | 4.49E-10 | 7.79E-07 |
| GO:0007610:
Behavior | 33 | 2.54E-08 | 4.41E-05 |
| GO:0019724: B
cell-mediated immunity | 12 | 3.99E-08 | 6.92E-05 |
| GO:0002478: Antigen
processing and presentation of exogenous peptide antigen | 7 | 5.36E-08 | 9.29E-05 |
| GO:0007155: Cell
adhesion | 40 | 1.72E-07 | 2.98E-04 |
| GO:0001568: Blood
vessel development | 22 | 1.75E-07 | 3.04E-04 |
| GO:0002526: Acute
inflammatory response | 14 | 3.02E-07 | 5.23E-04 |
| GO:0019884: Antigen
processing and presentation of exogenous antigen | 7 | 3.29E-07 | 5.70E-04 |
| GO:0001775: Cell
activation | 23 | 6.13E-07 | 1.06E-03 |
| GO:0030334:
Regulation of cell migration | 17 | 1.43E-06 | 2.48E-03 |
| GO:0048514: Blood
vessel morphogenesis | 19 | 1.45E-06 | 2.51E-03 |
| GO:0051270:
Regulation of cell motion | 18 | 1.83E-06 | 3.18E-03 |
| GO:0055066: Di-,
tri-valent inorganic cation homeostasis | 20 | 2.13E-06 | 3.69E-03 |
| GO:0045321:
Leukocyte activation | 20 | 2.56E-06 | 4.44E-03 |
| GO:0006956:
Complement activation | 9 | 3.78E-06 | 6.56E-03 |
| GO:0002541:
Activation of plasma proteins involved in acute inflammatory
response | 9 | 4.55E-06 | 7.90E-03 |
| GO:0001525:
Angiogenesis | 15 | 6.60E-06 | 1.14E-02 |
| GO:0042592:
Homeostatic process | 38 | 6.64E-06 | 1.15E-02 |
| GO:0040012:
Regulation of locomotion | 17 | 7.55E-06 | 1.31E-02 |
| GO:0009617:
Response to bacterium | 17 | 8.07E-06 | 1.40E-02 |
| GO:0055080: Cation
homeostasis | 21 | 8.17E-06 | 1.42E-02 |
| GO:0050865:
Regulation of cell activation | 16 | 1.04E-05 | 1.80E-02 |
| GO:0048878:
Chemical homeostasis | 29 | 1.48E-05 | 2.57E-02 |
| GO:0006874:
Cellular calcium ion homeostasis | 16 | 1.77E-05 | 3.07E-02 |
The enriched pathways of differential genes included
systemic lupus erythematosus, antigen processing and presentation,
complement and coagulation cascades, asthma, viral myocarditis,
lysosome, intestinal immune network for IgA production (Table III).
| Table IIIEnriched pathways of differentially
expressed transcripts. |
Table III
Enriched pathways of differentially
expressed transcripts.
| Term | Count | P-value | FDR |
|---|
| hsa05322: Systemic
lupus erythematosus | 21 | 3.65E-08 | 0.00004 |
| hsa04514: Cell
adhesion molecules (CAMs) | 23 | 2.78E-07 | 0.00033 |
| hsa04612: Antigen
processing and presentation | 18 | 3.26E-07 | 0.00039 |
| hsa04610:
Complement and coagulation cascades | 16 | 7.32E-07 | 0.00087 |
| hsa05310:
Asthma | 10 | 5.73E-06 | 0.00681 |
| hsa05416: Viral
myocarditis | 15 | 5.92E-06 | 0.00704 |
| hsa04142:
Lysosome | 19 | 1.13E-05 | 0.01347 |
| hsa04672:
Intestinal immune network for IgA production | 12 | 1.66E-05 | 0.01978 |
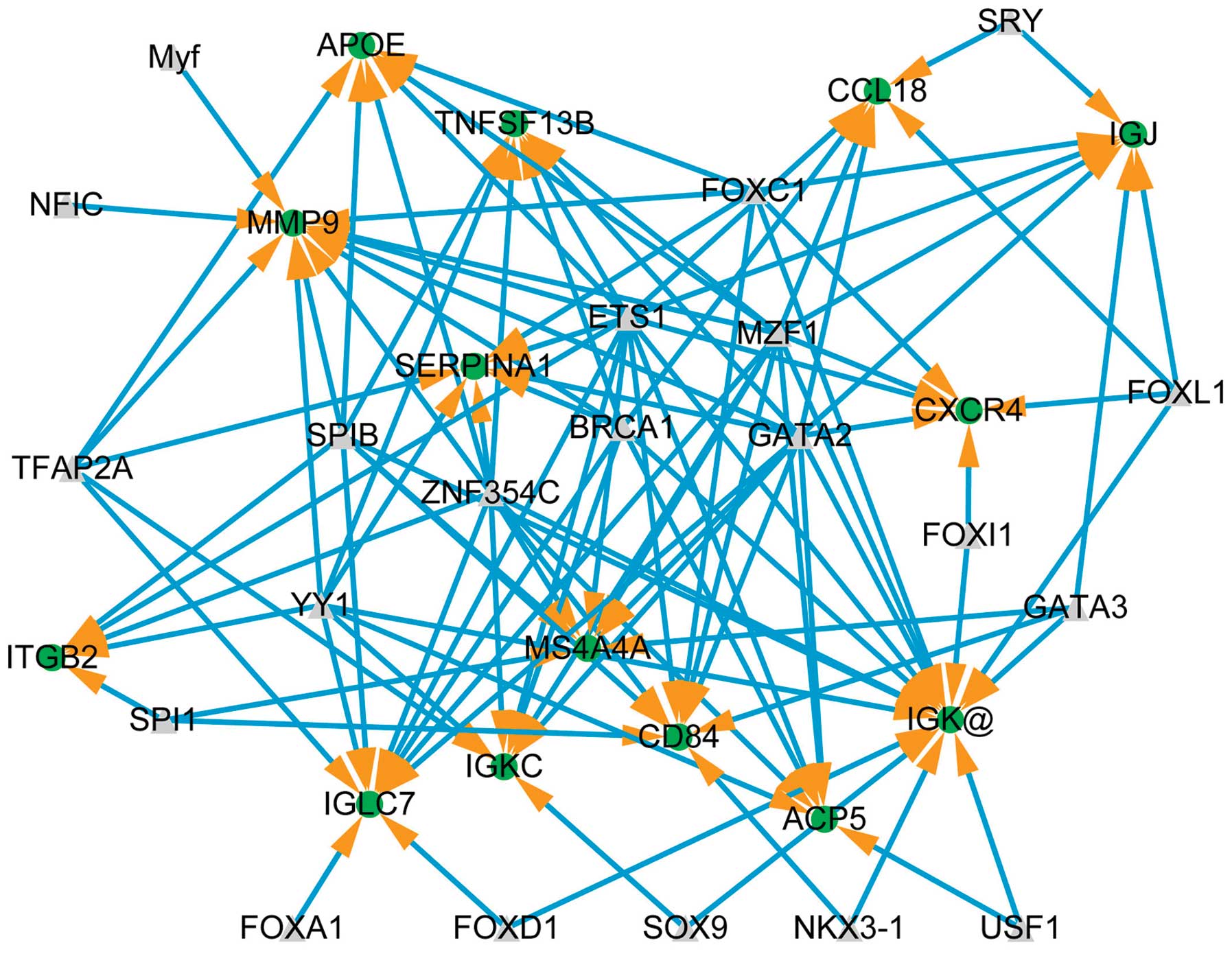
Transcriptional regulatory network
The core transcriptional regulatory network is shown
in Fig. 4 for upregulated genes
(RMA log2 transformed values >2). Fourteen upregulated genes are
shown, targeted by 21 transcription factors, constructing 111
regulatory relationships. The hub genes are IGK (15 neighbors),
MMP9 (11 neighbors) and IGLC7 (10 neighbors).
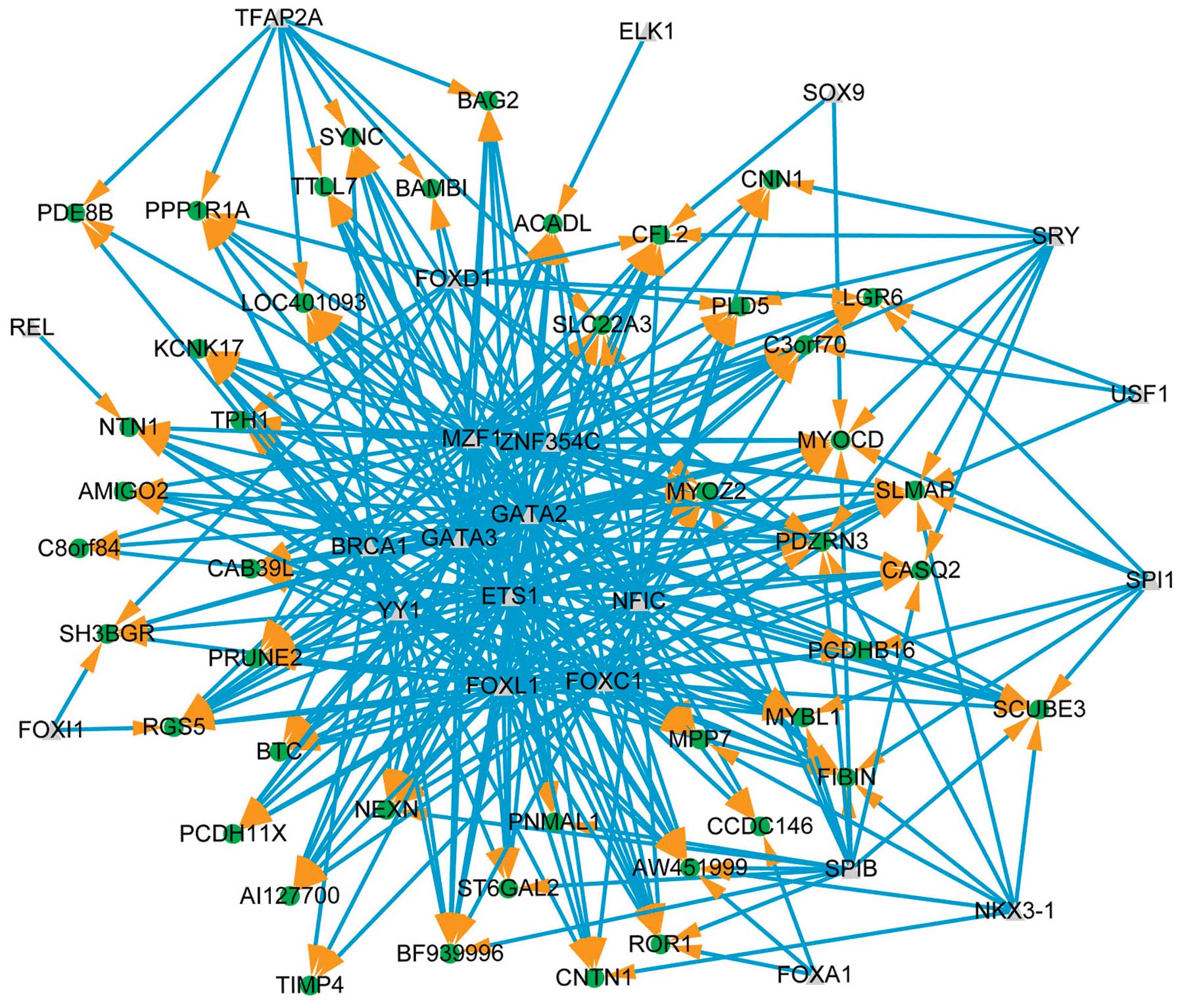
The core transcriptional regulatory network is shown
in Fig. 5 for downregulated genes
(RMA log2 transformed values <-1). Forty-five downregulated
genes, targeted by 22 transcription factors, constructing 324
regulatory relationships. The hub genes are AW451999 (10
neighbors), CFL2 (10 neighbors) and PDZRN3 (10 neighbors).
Significantly enriched cis-regulatory
signals
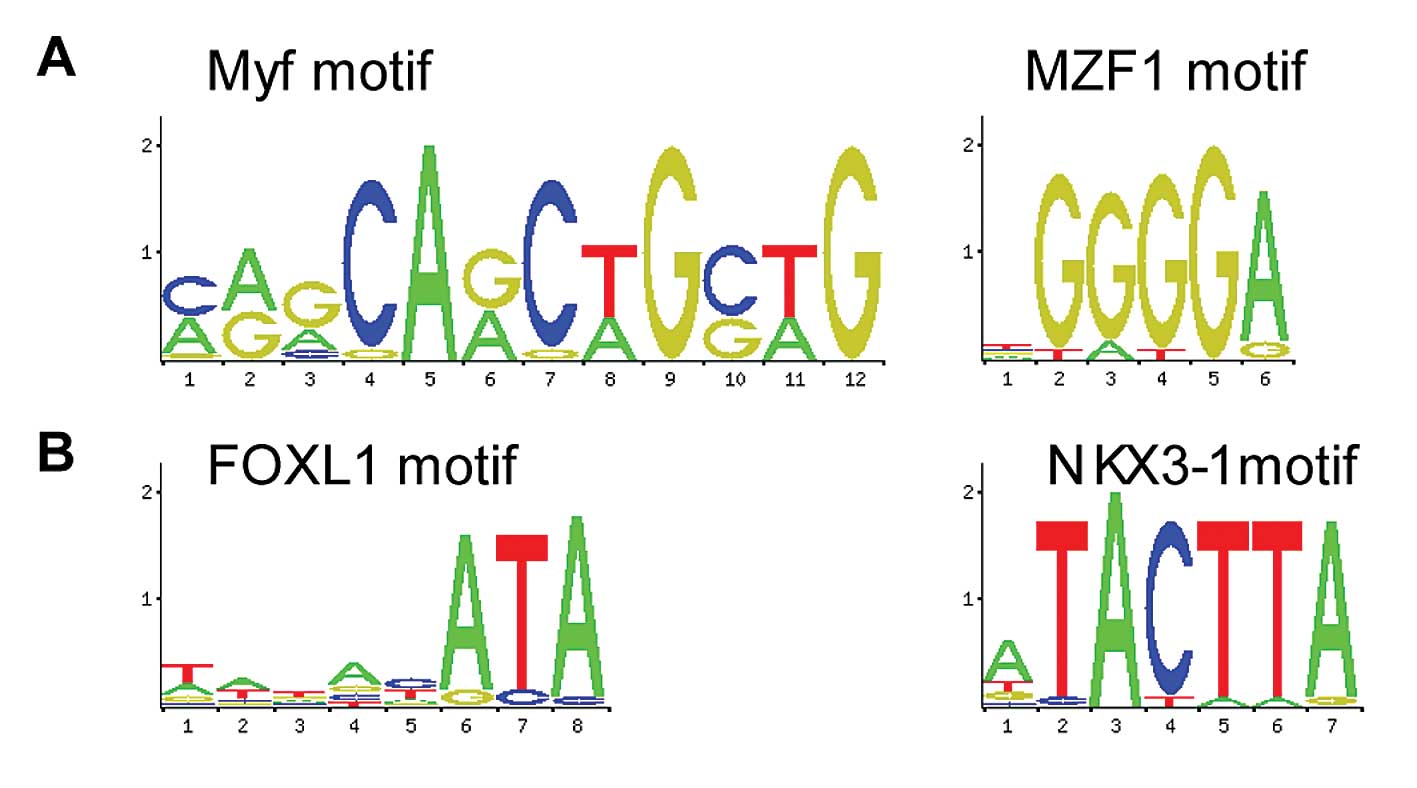
Fig. 6A shows the
5′UTR of upregulated genes enriched in TF motif Myf (P-value
1.33E-02) and MZF1 (P-value 2.47E-02). By contrast, Fig. 6B shows the 5′UTR of downregulated
genes enriched in TF motif FOXL1 (P-value, 5.95E-03) and NKX3-1
(P-value, 3.57E-02).
miRNA regulatory network
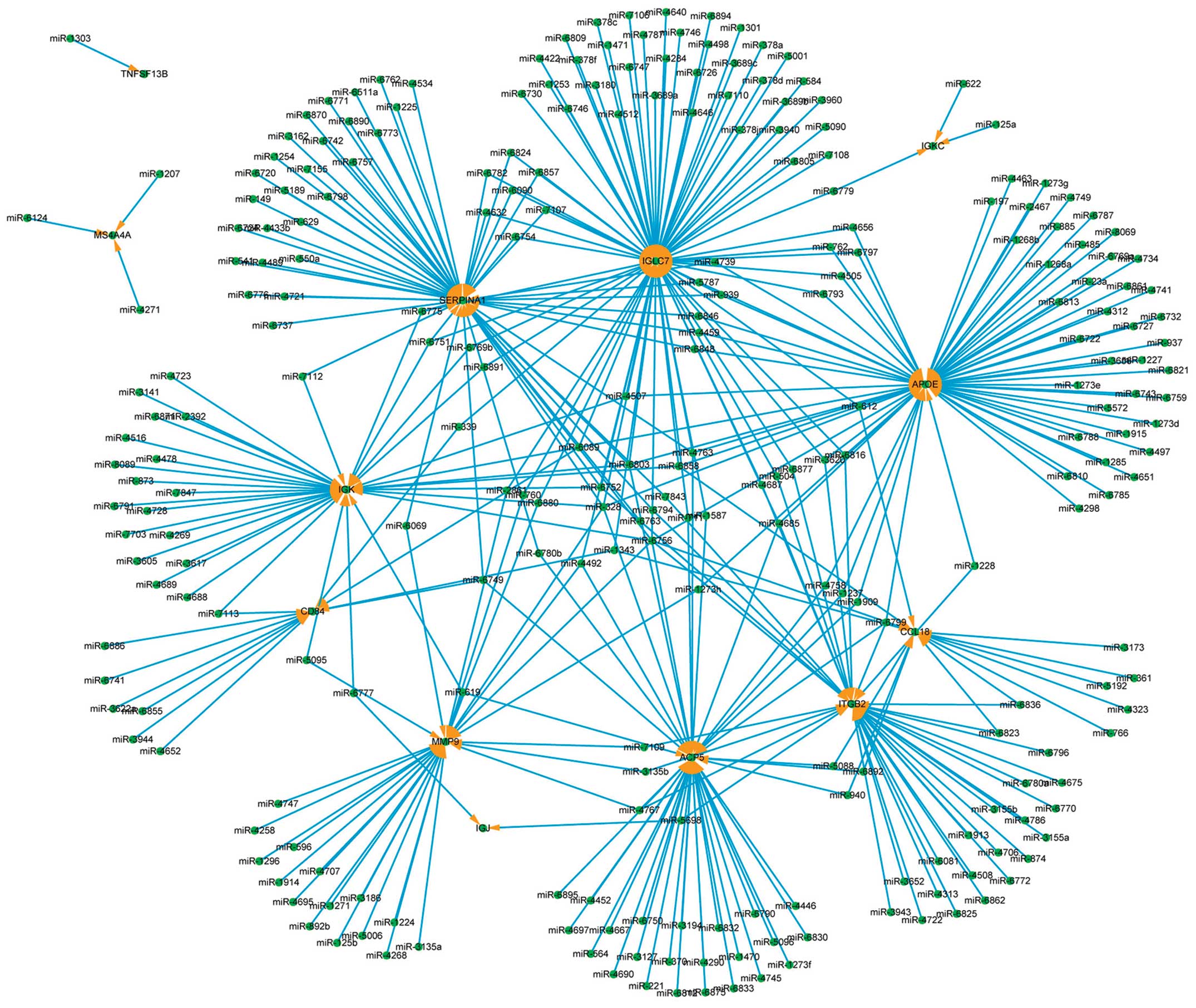
The core miRNA regulatory network is shown in
Fig. 7 for the upregulated genes
(RMA log2 transformed values >2). The genes identified by arrows
are 13 upregulated genes, targeted by 262 miRNAs, comprising 372
regulatory relationships. The hub genes are IGLC7 (78 miRNAs target
it), APOE (66 miRNAs) and SERPINA1 (53 miRNAs). The hub miRNAs are
miR-6756 (8 target genes), miR-328 (6 genes) and miR-6803 (5
genes).
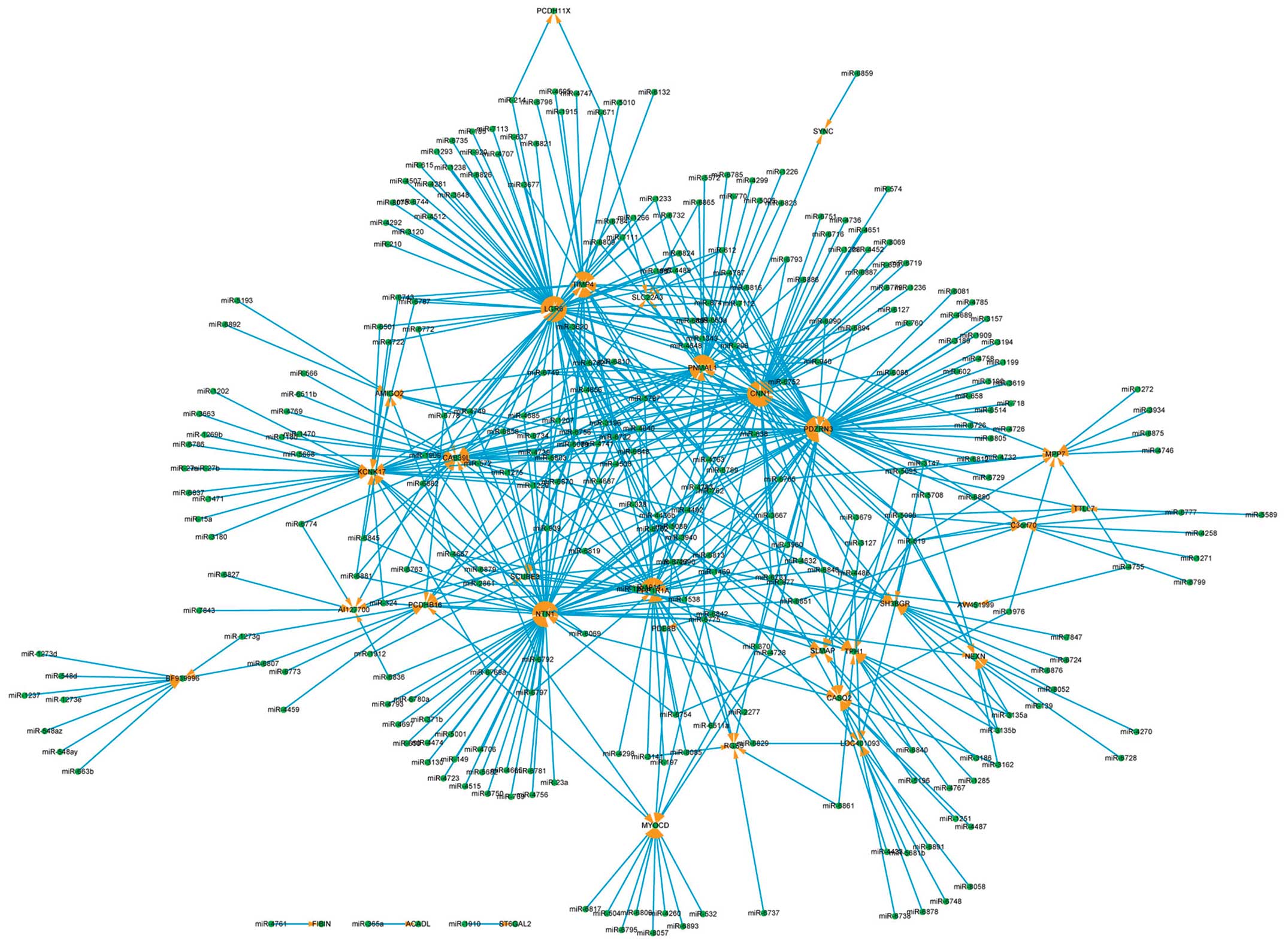
The core miRNA regulatory network is shown in
Fig. 8 for the downregulated
genes (RMA log2 transformed values >1). The genes identified by
arrows are 33 downregulated genes, targeted by 295 miRNAs,
comprising 561 regulatory relationships. The hub genes are LGR6 (71
miRNAs target it), NTN1 (61 miRNAs), CNN1 (59 miRNAs) and PDZRN3
(57 miRNAs). The hub miRNAs are miR-6756 (11 target genes), miR-619
(10 genes), miR-6089 (8 genes) and miR-6803 (8 genes).
Discussion
Carotid atherosclerosis is defined as the presence
of atherosclerotic plaques in any of the carotid vessel segments
(30). In the present study, we
aimed to identify gene expression changes and regulatory factors
for carotid atherosclerosis from an early to an advanced stage.
More genes were activated in advanced stage compared with early
stage. The upregulated genes in the advanced stage were involved in
GO terms including immune, vascular and cell movement homeostasis.
The differentially expressed genes (DEGs) were significantly
enriched in cell adhesion molecules (CAMs) and focal adhesion.
Genes such as MMP9 and CFL2 played key roles in the transcriptional
regulatory network. Moreover, miR-328 was one of the hubs in the
miRNA regulatory network.
A total of 889 transcripts were identified to be
differentially expressed from early stage plaques of carotid
atherosclerosis to advanced stage plaques. As shown in Fig. 3B, A/E increased when the fold
change threshold was elevated. The majority of the DEGs were
upregulated in the advanced stage, while they were inhibited in the
early stage.
The DEGs activated in the advanced stage may
correlate with plaques of carotid atherosclerosis and various types
of cancer. A number of immune system-related cells were detected in
human carotid atherosclerosis patients such as
monocytes/macrophages, T cells and plasmacytoid dendritic cells
(pDCs) (23). IGHG1 (Ig γ-1 chain
C region) is one gene associated with ‘innate immune response’.
IGHG1 expression has been reported to correlate with immune evasion
mechanisms, which contribute to the proliferation of human
pancreatic cancer (31).
Furthermore, inhibiting IGHG1 expression by siRNA leads to cancer
growth inhibition and apoptosis in prostate cancer (32). SPP1 (osteopontin) is involved as a
cytokine in type I immunity to elevate the product of interferon-γ
and interleukin-12. On the other hand, SPP1 reduces the expression
of interleukin-10, and IFN-γ treatment, resulting in an increase of
cytokines/cytokine receptors including CSF2, IL1R2 and SPP1
(33). In addition, smoking can
increase SPP1 expression, and subsequently induce inflammation and
emphysema (34). IGKC (Ig κ chain
C region) participates in humoral immune response, which has been
reported to play key roles in non-small cell lung cancer and breast
cancer (35–37). The genes activated in early stage
do not possess a similar function, including BTC, MYBL1 and
PLD.
Immune-associated GO terms, vascular-related GO
terms and cell movement homeostasis were identified to be
associated with atherosclerosis. The IL-6-gp130 axis is a key
regulator of inflammatory acute phase signaling in hepatocytes for
the development of atherosclerosis (38). Blood flow is crucial for blood
vessel development during embryogenesis and for the regulation of
vessel diameter in adult life. It is also a key factor in
atherosclerosis, which occurs mainly in regions of arteries that
experience disturbances in fluid flow (39). Fibrinolytic balance and the
potential contribution of PAI-1 to the regulation of cell migration
are involved in the pathogenesis of the simple atherosclerotic
lesions observed in the mouse (40).
Leukocyte transendothelial migration is one of the
earliest events of immune inflammatory responses and may contribute
to atherosclerosis (41).
Immunologic arterial injury due to allograft rejection acting in
synergy with hypercholesterolemia resulting from a dietary
supplement of cholesterol can lead to rapidly developing
atherosclerosis (42). Autoimmune
thyroid disease has a causal relationship with atherosclerosis
(even if mediated through traditional risk factors) (43). Focal adhesion plays key roles in
VSMCs. Focal adhesion pathways may be expected to facilitate the
formation of atherosclerotic plaques in ApoE-null mice (44). Systemic lupus erythematosus (SLE)
is a systemic autoimmune disease that is characterized by
autoantibody production and inflammatory disease involving multiple
organs. Premature atherosclerosis is a common complication of SLE
and results in substantial morbidity and mortality from
cardiovascular disease (CVD) (45). Diabetes and atherosclerosis are
associated with disorders of lipids and lipoproteins, notably high
apolipoprotein B (apoB) and low apolipoprotein A1 (apoA1) are well
established (46). Type I
diabetes mellitus was also enriched by differential genes in this
study. The passage of leukocytes across the endothelium and into
arterial walls is a critical step in the development of
atherosclerosis. It is consistent with our observation that DEGs
were enriched in the pathway ‘leukocyte transendothelial migration’
(47).
The expression levels of matrix metallopeptidase 9
(MMP9) were assessed. MMP9 was potentially important in the
development of atherosclerosis in a Malaysian study population
(13). Cystic fibrosis
transmembrane conductance regulator (CFTR) has a similar function
to ABCA1. Schmitz and Buechler (48) identified ABCA1 as the major
regulator of plasma high density lipoprotein (HDL) cholesterol. HDL
metabolism is crucial in the prevention of the progression of
atherosclerosis. CFL2 is an interactor of CFTR as reported by Wang
and colleagues (49). An in
vivo ApoE−/− mouse model was utilized to assess the
effects of chronic moderate exposure to arsenic on plaque formation
and composition in order to facilitate mechanistic investigations
(50). Arsenic exposure increases
oxidative stress, inflammation and atherosclerotic lesion formation
in ApoE−/− mice (51).
In human arterial tissue, SERPINA1 was upregulated (6.3-fold) in
atherosclerotic plaques (14). In
the present study, SERPINA1 was also upregulated in the advanced
stage.
In this study, we also found that miR-328 may be
crucial for atherosclerosis. miR-328 was linked to multiple
upregulated genes in advanced stage samples and miR-328 has been
found to be antiangiogenic (52).
Anti-angiogenic perfluorocarbon nanoparticles has already used for
diagnosis and treatment of atherosclerosis (53). Recently, in the ABCG2-positive
cancer cells, miR-328 has been reported to regulate the expression
of BCRP/ABCG2 (54).
Additionally, miR-328 expression in plasma was significantly
increased in atrial fibrillation (AF) patients (55).
Motifs enriched by upregulated genes in the early
and advanced stages including Myf, MZF1, FOXL1 and NKX3-1, which
were not investigated extensively were also investigated. These
transcriptional factors may play pivotal roles in carotid
atherosclerosis. In the present study, we identified gene
expression changes and regulatory factors in carotid
atherosclerosis. These results may facilitate in identifying the
mechanism involved in carotid atherosclerosis.
References
|
1
|
Ross R: The pathogenesis of
atherosclerosis: a perspective for the 1990s. Nature. 362:801–809.
1993. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Maseri A and Fuster V: Is there a
vulnerable plaque? Circulation. 107:2068–2071. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ross R: Atherosclerosis - an inflammatory
disease. N Engl J Med. 340:115–126. 1999. View Article : Google Scholar
|
|
4
|
O’Leary DH, Polak JF, Kronmal RA, Manolio
TA, Burke GL and Wolfson SK Jr: Carotid-artery intima and media
thickness as a risk factor for myocardial infarction and stroke in
older adults. Cardiovascular Health Study Collaborative Research
Group. N Engl J Med. 340:14–22. 1999.
|
|
5
|
Wu Y, Tao Z, Song C, et al: Overexpression
of YKL-40 predicts plaque instability in carotid atherosclerosis
with CagA-positive helicobacter pylori infection. PLoS One.
8:e599962013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Yamada M, Kim S, Egashira K, et al:
Molecular mechanism and role of endothelial monocyte
chemoattractant protein-1 induction by vascular endothelial growth
factor. Arterioscler Thromb Vasc Biol. 23:1996–2001. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Leng XY, Chen XY, Chook P, et al:
Association between metabolic syndrome and carotid atherosclerosis:
a community-based study in Hong Kong. Metab Syndr Relat Disord.
11:109–114. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ota H, Reeves MJ, Zhu DC, et al: Sex
differences of high-risk carotid atherosclerotic plaque with less
than 50% stenosis in asymptomatic patients: an in vivo 3T MRI
study. AJNR Am J Neuroradiol. 34:1049–1055. 2013.PubMed/NCBI
|
|
9
|
Ino-Oka E, Sekino H, Kajikawa S, Satoh T
and Inooka H: Evaluation of carotid atherosclerosis from the
perspective of blood flow reflection. Clin Exp Hypertens.
31:188–200. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Maeda S, Sawayama Y, Furusyo N, Shigematsu
M and Hayashi J: The association between fatal vascular events and
risk factors for carotid atherosclerosis in patients on maintenance
hemodialysis: plaque number of dialytic atherosclerosis study.
Atherosclerosis. 204:549–555. 2009. View Article : Google Scholar
|
|
11
|
Munakata M, Sakuraba J, Tayama J, et al:
Higher brachial-ankle pulse wave velocity is associated with more
advanced carotid atherosclerosis in end-stage renal disease.
Hypertens Res. 28:9–14. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hashimoto H, Kitagawa K, Kuwabara K, et
al: Circulating adhesion molecules are correlated with ultrasonic
assessment of carotid plaques. Clin Sci (Lond). 104:521–527. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Blin J, Ahmad Z, Rampal LR, Mohtarrudin N,
Tajudin AK and Adnan RS: Preliminary assessment of differential
expression of candidate genes associated with atherosclerosis.
Genes Genet Syst. 88:199–209. 2013.PubMed/NCBI
|
|
14
|
Inouye M, Ripatti S, Kettunen J, et al:
Novel Loci for metabolic networks and multi-tissue expression
studies reveal genes for atherosclerosis. PLoS Genet.
8:e10029072012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bateman HR, Liang Q, Fan D, Rodriguez V
and Lessner SM: Sparstolonin B inhibits pro-angiogenic functions
and blocks cell cycle progression in endothelial cells. PLoS One.
8:e705002013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Feig JE, Vengrenyuk Y, Reiser V, et al:
Regression of atherosclerosis is characterized by broad changes in
the plaque macrophage transcriptome. PLoS One. 7:e397902012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zhang E and Wu Y: Dual effects of miR-155
on macrophages at different stages of atherosclerosis: LDL is the
key? Med Hypotheses. 2014. 83:74–78. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Bonaterra GA, Zugel S, Thogersen J, et al:
Growth differentiation factor-15 deficiency inhibits
atherosclerosis progression by regulating interleukin-6-dependent
inflammatory response to vascular injury. J Am Heart Assoc.
1:e0025502012. View Article : Google Scholar
|
|
19
|
Orr AW, Hastings NE, Blackman BR and
Wamhoff BR: Complex regulation and function of the inflammatory
smooth muscle cell phenotype in atherosclerosis. J Vasc Res.
47:168–180. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
States JC, Singh AV, Knudsen TB, et al:
Prenatal arsenic exposure alters gene expression in the adult liver
to a proinflammatory state contributing to accelerated
atherosclerosis. PLoS One. 7:e387132012. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bahls M, Bidwell CA, Hu J, et al: Gene
expression differences during the heterogeneous progression of
peripheral atherosclerosis in familial hypercholesterolemic swine.
BMC Genomics. 14:4432013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Pinkaew D, Hutadilok-Towatana N, Teng BB,
Mahabusarakam W and Fujise K: Morelloflavone, a biflavonoid
inhibitor of migration-related kinases, ameliorates atherosclerosis
in mice. Am J Physiol Heart Circ Physiol. 302:H451–H458. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Doring Y, Manthey HD, Drechsler M, et al:
Auto-antigenic protein-DNA complexes stimulate plasmacytoid
dendritic cells to promote atherosclerosis. Circulation.
125:1673–1683. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Irizarry RA, Hobbs B, Collin F, et al:
Exploration, normalization, and summaries of high density
oligonucleotide array probe level data. Biostatistics. 4:249–264.
2003. View Article : Google Scholar
|
|
25
|
Philip R: Semantic similarity in a
taxonomy: an information-based measure and its application to
problems of ambiguity in natural language. J Artif Intell Res.
11:95–130. 1999.
|
|
26
|
Huang da W, Sherman BT and Lempicki RA:
Systematic and integrative analysis of large gene lists using DAVID
bioinformatics resources. Nat Protoc. 4:44–57. 2009.PubMed/NCBI
|
|
27
|
Portales-Casamar E, Thongjuea S, Kwon AT,
et al: JASPAR 2010: the greatly expanded open-access database of
transcription factor binding profiles. Nucleic Acids Res.
38:D105–D110. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wang K, Hu F, Xu K, et al: CASCADE_SCAN:
mining signal transduction network from high-throughput data based
on steepest descent method. BMC Bioinformatics. 12:1642011.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kruger J and Rehmsmeier M: RNAhybrid:
microRNA target prediction easy, fast and flexible. Nucleic Acids
Res. 34:W451–W454. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li LX, Zhao CC, Ren Y, et al: Prevalence
and clinical characteristics of carotid atherosclerosis in newly
diagnosed patients with ketosis-onset diabetes: a cross-sectional
study. Cardiovasc Diabetol. 12:182013. View Article : Google Scholar
|
|
31
|
Li X, Ni R, Chen J, et al: The presence of
IGHG1 in human pancreatic carcinomas is associated with immune
evasion mechanisms. Pancreas. 40:753–761. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Pan B, Zheng S, Liu C and Xu Y:
Suppression of IGHG1 gene expression by siRNA leads to growth
inhibition and apoptosis induction in human prostate cancer cell.
Mol Biol Rep. 40:27–33. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kitaya K, Yasuo T, Yamaguchi T, Fushiki S
and Honjo H: Genes regulated by interferon-γ in human uterine
microvascular endothelial cells. Int J Mol Med. 20:689–697.
2007.
|
|
34
|
Shan M, Yuan X, Song LZ, et al: Cigarette
smoke induction of osteopontin (SPP1) mediates TH17
inflammation in human and experimental emphysema. Sci Transl Med.
4:117ra92012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Lohr M, Edlund K, Botling J, et al: The
prognostic relevance of tumour-infiltrating plasma cells and
immunoglobulin kappa C indicates an important role of the humoral
immune response in non-small cell lung cancer. Cancer Lett.
333:222–228. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Whiteside TL and Ferrone S: IGKC and
prognosis in breast cancer - response to Schmidt. Clin Cancer Res.
19:3052013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Schmidt M, Micke P and Hengstler JG: IGKC
and prognosis in breast cancer. Clin Cancer Res. 19:3042013.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Luchtefeld M, Preuss C, Ruhle F, et al:
Gp130-dependent release of acute phase proteins is linked to the
activation of innate immune signaling pathways. PLoS One.
6:e194272011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Hahn C and Schwartz MA:
Mechanotransduction in vascular physiology and atherogenesis. Nat
Rev Mol Cell Biol. 10:53–62. 2009. View
Article : Google Scholar : PubMed/NCBI
|
|
40
|
Sjoland H, Eitzman DT, Gordon D, Westrick
R, Nabel EG and Ginsburg D: Atherosclerosis progression in LDL
receptor-deficient and apolipoprotein E-deficient mice is
independent of genetic alterations in plasminogen activator
inhibitor-1. Arterioscler Thromb Vasc Biol. 20:846–852. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Languino LR, Duperray A, Joganic KJ,
Fornaro M, Thornton GB and Altieri DC: Regulation of
leukocyte-endothelium interaction and leukocyte transendothelial
migration by intercellular adhesion molecule 1-fibrinogen
recognition. Proc Natl Acad Sci USA. 92:1505–1509. 1995. View Article : Google Scholar
|
|
42
|
Alonso DR, Starek PK and Minick CR:
Studies on the pathogenesis of atheroarteriosclerosis induced in
rabbit cardiac allografts by the synergy of graft rejection and
hypercholesterolemia. Am J Pathol. 87:415–442. 1977.PubMed/NCBI
|
|
43
|
McLeod DS: Autoimmune thyroid disease: a
novel risk factor for atherosclerosis? Endocrine. 44:8–10. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Bu DX, Rai V, Shen X, et al: Activation of
the ROCK1 branch of the transforming growth factor-β pathway
contributes to RAGE-dependent acceleration of atherosclerosis in
diabetic ApoE-null mice. Circ Res. 106:1040–1051. 2010.PubMed/NCBI
|
|
45
|
Richez C, Richards RJ, Duffau P, et al:
The effect of mycophenolate mofetil on disease development in the
gld.apoE−/− mouse model of accelerated
atherosclerosis and systemic lupus erythematosus. PLoS One.
8:e610422013.PubMed/NCBI
|
|
46
|
Hashemi M, Saadat M, Behjati M and
Kelishadi R: Comparison of serum apolipoprotein levels of diabetic
children and healthy children with or without diabetic parents.
Cholesterol. 2012:4903812012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Samson T, van Buul JD, Kroon J, et al: The
guanine-nucleotide exchange factor SGEF plays a crucial role in the
formation of atherosclerosis. PLoS One. 8:e552022013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Schmitz G and Buechler C: ABCA1:
regulation, trafficking and association with heteromeric proteins.
Ann Med. 34:334–347. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Wang X, Venable J, LaPointe P, et al:
Hsp90 cochaperone Aha1 downregulation rescues misfolding of CFTR in
cystic fibrosis. Cell. 127:803–815. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Lemaire M, Lemarie CA, Molina MF,
Schiffrin EL, Lehoux S and Mann KK: Exposure to moderate arsenic
concentrations increases atherosclerosis in ApoE−/−
mouse model. Toxicol Sci. 122:211–221. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Srivastava S, Vladykovskaya EN, Haberzettl
P, Sithu SD, D’Souza SE and States JC: Arsenic exacerbates
atherosclerotic lesion formation and inflammation in ApoE−/− mice.
Toxicol Appl Pharmacol. 241:90–100. 2009.PubMed/NCBI
|
|
52
|
Suarez Y and Sessa WC: MicroRNAs as novel
regulators of angiogenesis. Circ Res. 104:442–454. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Caruthers SD, Cyrus T, Winter PM, Wickline
SA and Lanza GM: Anti-angiogenic perfluorocarbon nanoparticles for
diagnosis and treatment of atherosclerosis. Wiley Interdiscip Rev
Nanomed Nanobiotechnol. 1:311–323. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Li X, Pan YZ, Seigel GM, Hu ZH, Huang M
and Yu AM: Breast cancer resistance protein BCRP/ABCG2 regulatory
microRNAs (hsa-miR-328, -519c and -520h) and their differential
expression in stem-like ABCG2+ cancer cells. Biochem
Pharmacol. 81:783–792. 2011. View Article : Google Scholar
|
|
55
|
Wang X, Qiu CG, Huang ZW, Han ZY, Lu WJ
and Chen XJ: The expression and clinical implication of plasma
miR-328 in patients with atrial fibrillation. Zhonghua Xin Xue Guan
Bing Za Zhi. 41:126–129. 2013.(In Chinese).
|