Introduction
Acute myeloid leukemia (AML) is defined as a clonal
disorder caused by the malignant transformation of a bone
marrow-derived, self-renewing stem cell or progenitor, which
demonstrates a decreased rate of self-destruction, as well as
aberrant differentiation (1). AML
consists of a group of malignant disorders which are characterized
by the replacement of normal bone marrow with abnormal, primitive
hematopoietic cells (2). If left
untreated, the disorder uniformly results in death, usually due to
infection or bleeding. Pediatric AML comprises up to 20% of all
childhood leukemia cases. Epigenetic disturbances have been
implicated in the development and pathogenesis of leukemia
(3). The inactivation of tumor
suppressor genes by promoter hypermethylation has been increasingly
recognized as a key event in leukemia, with the silencing of tumor
suppressor genes by aberrant DNA hypermethylation reported in
hematologic malignancies, including subsets of AML (4–6).
Previous studies have identified DNA methylation of other tumor
suppressor genes in AML, including retinoic acid receptor β
(RARβ), estrogen receptor (ER), p15 (7), E-cadherin, p16 (7), glutathione S-transferase Pi 1
(GSTP1) (8),
hypermethylated in cancer 1 (HIC1) (9) and death-associated protein kinase 1
(DAPK1) (10).
DAPK1 is the most frequently methylated gene in both
pediatric (70%) and adult AML (55%). RARβ is aberrantly
methylated in 18% of AML cases and p73 (11) is methylated in 10% of AML cases
(8). Identifying novel aberrantly
methylated genes may provide a better understanding of AML, thereby
paving the way for the development of novel tumor markers and
therapeutic targets (7,12,13).
Recently, epigenetic disturbances, such as aberrant
promoter hypermethylation and covalent histone modifications
(14), have been implicated in
the pathogenesis of leukemia (3,15).
In addition, these aberrations are responsible for enhanced
proliferation and self renewal, differentiation arrest, as well as
the impaired apoptosis of leukemic blasts (16). There are two major mechanisms of
DNA methylation-associated gene silencing. The first mechanism is
that methylation directly blocks activating transcription factors
from binding to their target sequences. A number of transcription
factors [activating protein 2 (AP-2), cAMP response element binding
protein (CREB)/activating transcription factor (ATF), c-MYC and E2F
family members] are known to bind unmethylated promoter sequences,
but fail to bind at methylated sites. The second mechanism involves
the acquisition of methyl-binding domain proteins
[methyl-CpG-binding domain protein 1 (MBD1-4), methyl CpG binding
protein 2 (Rett syndrome) (MeCP2) and Kaiso] and the recruitment of
additional factors with repressive properties, such as histone
deacetylases (17,18). The epigenetic control of gene
expression can alter gene function without modification of the DNA
coding sequence and has been suggested to play a fundamental role
in normal and tumor cells (19).
Over the past few years, the role of genetic events
in AML has been studied extensively, and a number of molecular
defects and their contribution to leukemogenesis have been
unraveled (20). However,
epigenetic alterations in AML are not yet fully understood. Zinc
finger (ZNF) proteins are among the most abundant proteins in
eukaryotic genomes. ZNF protein functions are extraordinarily
diverse and include DNA recognition, RNA packaging, transcriptional
activation, regulation of apoptosis, protein folding and assembly,
as well as lipid binding. ZNF proteins play important roles in a
variety of physiological processes. ZNF transcription factors are
involved broadly in development and tumorigenesis (21,23,29). The knockdown and overexpression of
ZNF280B protein have demonstrated that ZNF280B is involved in both
pro-growth and pro-survival functions in prostate cancer. However,
surprisingly, the pro-cancer functions of ZNF280B are mediated by
the downregulation of p53 (21).
ZNF300 expression enhances nuclear factor (NF)-κB signaling by
activating TNF receptor-associated factor 2 (TRAF2) and physically
interacting with IKKβ. Furthermore, ZNF300 overexpression increases
extracellular signal-regulated kinase (ERK)1/2 phosphorylation. It
has been demonstrated that ZNF300 overexpression stimulates
prostate cancer cell proliferation in vitro and
significantly enhances tumor development and metastasis in a mouse
xenograft model (22). The
overexpression of ZNF425 inhibits the transcriptional activities of
serum response element (SRE), AP-1 and serum response factor (SRF);
deletion analysis indicates that the C2H2 domain is the main region
responsible for the repression. The ZNF425 gene is a novel
transcriptional inhibitor that functions in the MAPK signaling
pathway (23).
ZNF382 functions as a tumor suppressor in multiple
carcinomas. It has been shown that the ectopic expression of ZNF382
in silenced tumor cells significantly inhibits their clonogenicity
and proliferation, and induces apoptosis (24). ZNF382 has been shown to be
frequently methylated in multiple primary tumors, such as
nasopharyngeal, esophageal, colon, gastric and breast cancer
(24). To the best of our
knowledge, there are no studies available to date on the expression
of ZNF382 and the methylation status of its promoter in pediatric
AML. In this study, we provide the first evidence of ZNF382
methylation in both AML cell lines and pediatric myeloid leukemia
samples. These results suggest that ZNF382 functions as a tumor
suppressor in pediatric AML.
Materials and methods
Cell lines
The leukemia cell lines, HL-60, MV4-11, U937 and
K562, were obtained from the American Type Culture Collection
(ATCC, Manassas, VA, USA). The Raji, Jurkat, 697, NB4, THP-1, Daudi
and SHI-1 cell lines were kind gifts from Professor Wang Jian-Rong
(Cyrus Tang Hematology Center of Soochow University, Suzhou,
China). All the cell lines were maintained at 37°C in RPMI-1640
medium (GibcoR; Life Technologies, Carlsbad, CA, USA) supplemented
with 10% fetal bovine serum (Invitrogen, Life Technologies,
Carlsbad, CA, USA).
Patients and samples
Bone marrow specimens were obtained at the time of
diagnosis during the routine clinical assessment of 105 pediatric
patients with AML, who presented at the Department of Hematology
and Oncology, Children’s Hospital of Soochow University between
2006 and 2011. Ethical approval was provided by the Children’s
Hospital of Soochow University Ethics Committee (nos. SUEC2006-011
and SUEC2000-021), and informed consent was obtained from the
parents or guardians. AML diagnosis was made in accordance with the
revised French-American-British (FAB) classification. ZNF382
expression and methylation were assessed in 105 Chinese pediatric
AML patients with clinical follow-up records, including patient
age, gender, FAB, cytogenetic and survival information.
Multivariate analysis of ZNF382 expression or promoter methylation
was also performed. Additionally, bone marrow samples from 12
healthy donors and 8 patients with idiopathic thrombocytopenic
purpura (ITP) were analyzed as the controls. Bone marrow
mononuclear cells (BMNCs) were isolated using Ficoll solution
within 2 h after the bone marrow samples harvested. Analysis of the
methylation status of genes in the pediatric AML samples (M1, M2,
M3, M4 and M5) and 3 NBM control samples (N1, N2 and N3) was
carried out using the NimbleGen Human DNA Methylation 385K Promoter
Plus CpG Island Array. The microarray analysis was performed by
KangChen Bio-tech, Shanghai P.R. China.
CD34+ cell purification
For the selection CD34+ cells, the
Miltenyi immunoaffinity device (VarioMACS 130-046-703) was used
according to the instructions provided by the manufacturer.
(Miltenyi Biotech, Auburn, CA, USA). Briefly, the CD34+
cells were magnetically labeled with CD34 MicroBeads. Subsequently,
the cell suspension was loaded onto a MACSR Column which was placed
in the magnetic field of a MACS Separator. The magnetically labeled
CD34+ cells were retained within the column. The
unlabeled cells ran through; the CD34+ cells were
adsorbed on the magnetic poles. Following the removal of the column
from the magnetic field, the magnetically retained CD34+
cells were eluted as the positively selected cell fraction.
Sodium bisulphite modification of genomic
DNA
High-molecular-weight genomic DNA was extracted from
the cell lines and biopsy samples by a conventional
phenol/chloroform method. The sodium bisulphite modification
procedure was carried out as previously described with slight
modifications (25). In brief,
600 ng of genomic DNA were denatured in 3 M NaOH for 15 min at
37°C, then mixed with 2 vol 2% low-melting-point agarose.
Agarose/DNA mixtures were then pipetted into chilled mineral oil to
form agarose beads. Aliquots of 200 μl of 5 M bisulphite solution
(2.5 M sodium metabisulphite and 100 mM hydroquinone, both from
Sigma-Aldrich, St. Louis, MO, USA) were added to each tube
containing a single bead. The bisulphite reaction was then carried
out by incubating the reaction mixture for 4 h at 50°C in the dark.
Treatments were terminated by equilibration against 1 ml of TE
buffer, followed by desulphonation in 500 μl of 0.2 M NaOH.
Finally, the beads were washed with 1 ml of TE buffer and used
directly for PCR.
Methylation-specific (MSP) PCR
The methylation status of the ZNF382 promoter region
was determined by MSP PCR analysis. Primers distinguishing
unmethylated (U) and methylated (M) alleles were designed with
MethPrimer (http://www.urogene.org/methprimer/) to amplify the
sequence: ZNF382 B M forward, 5′-AGGTTAGGGAGGTAATTAA GAGTTC-3′ and
reverse, 5′-GAATCACCCTCAAAATC ACG-3′; ZNF382 B U forward,
5′-GAGGTTAGGGAGGTAA TTAAGAGTTT-3′ and reverse, 5′-CAAATCACCCTCAAAA
TCACAC-3′.
Each PCR reaction contained 20 ng of sodium
bisulphite-modified DNA, 250 pmol of each primer, 250 pmol
deoxynucleoside triphosphate, 1× PCR buffer, and one unit of ExTaq
HS polymerase (Takara, Tokyo, Japan) in a final reaction volume of
20 μl. Cycling conditions were initial denaturation at 95°C for 3
min, 40 cycles of 94°C for 30 sec, 65 (M) or 63°C (U) for 30 sec
and 72°C for 30 sec. For each set of methylation-specific PCR
reactions, in vitro-methylated genomic DNA treated with
sodium bisulphite served as a positive methylation control. PCR
products were separated on 4% agarose gels, stained with ethidium
bromide and visualized under UV illumination. For cases with
borderline results, PCR analyses were repeated.
Bisulfite genomic sequencing (BGS)
BGS was performed as previously described (24). Primers were designed with
MethPrimer (http://www.urogene.org/methprimer/). The primers for
BGS analysis were from +132 to +431, including 37 CpGs: ZNF382
forward, 5′-AGGTTAGGGAGGTAATTAAGAGTT-3′ and reverse,
5′-AACACCTAAAATACAACAAATTAAAC-3′. Amplified BGS products were
TA-cloned; and 5 to 6 randomly selected colonies were sequenced.
DNA sequences were analyzed with QUMA analyzer. (http://quma.cdb.riken.jp/).
Leukemia cells treated with
5-aza-2′-deoxycytidine (5-Aza-dC)
Demethylation was induced with 5-Aza-dC (5-Aza;
Sigma-Aldrich) treatment at a concentration that induced
demethylation of the DNA without killing the cells. The culture
medium for the HL-60 and MV4-11 cells contained 5 μM 5-Aza. DNA and
RNA were extracted after 72 h of 5-Aza treatment for the following
analysis. A group treated with DMSO was used as the control
group.
Quantitative reverse-transcription PCR
(RT-qPCR) for ZNF382
RT-qPCR was performed to determine the expression
levels of the ZNF382 gene. Total RNA was reverse transcribed using
the Reverse Transcription kit, according to the manufacturer’s
instructions (Applied Biosystems Inc., Foster City, CA, USA). The
real-time PCR primers used to quantify GAPDH expression were:
forward, 5′-AGAAGGCTGGGGCTCATTTG-3′ and reverse,
5′-AGGGGCCATCCACAGTCTTC-3′; and for ZNF382 forward,
5′-TGAGGGGTCTAGAATGCCTG-3′ and reverse,
5′-TGGACACAGAGAATTTTTCCA-3′. Real-time PCR was performed using
FastStart SYBR-Green Master (cat. no. 04673484001) on a
LightCycler480 system [both from Roche Diagnostics (Schweiz) AG,
Rotkreuz, Switzerland]. The expression of ZNF382 was normalized to
endogenous GAPDH expression.
Statistical analysis
SPSS software version 11.5 (SPSS Inc., Chicago, IL,
USA) was used for statistical analysis. Data are presented as the
means ± standard deviation. A group t-test was used to compare the
expression of ZNF382 between the DMSO group and the 5-Aza group.
Statistical significance between the methylated sample data and
clinicopathological characteristics of the AML patients were
analyzed by Pearson’s Chi-square test or Fisher’s exact test.
Statistical significance of ZNF382 expression among normal bone
marrow (NBM) and pediatric AML groups was determined using one-way
ANOVA. A value of P<0.05 was considered to indicate a
statistically significant difference.
Results
ZNF382 promoter is hypermethylated in AML
cells
ZNF382 is a functional tumor suppressor frequently
methylated in multiple carcinomas, including nasopharyngeal,
esophageal, colon, gastric and breast cancer (24). However, the methylation status of
ZNF382 in the blood system is unknown, particularly in pediatric
AML. Our analyses of promoter methylation in pediatric AML
suggested that the ZNF382 promoter was hypermethylated in AML using
NimbleGen Human DNA Methylation 385K Promoter plus CpG Island
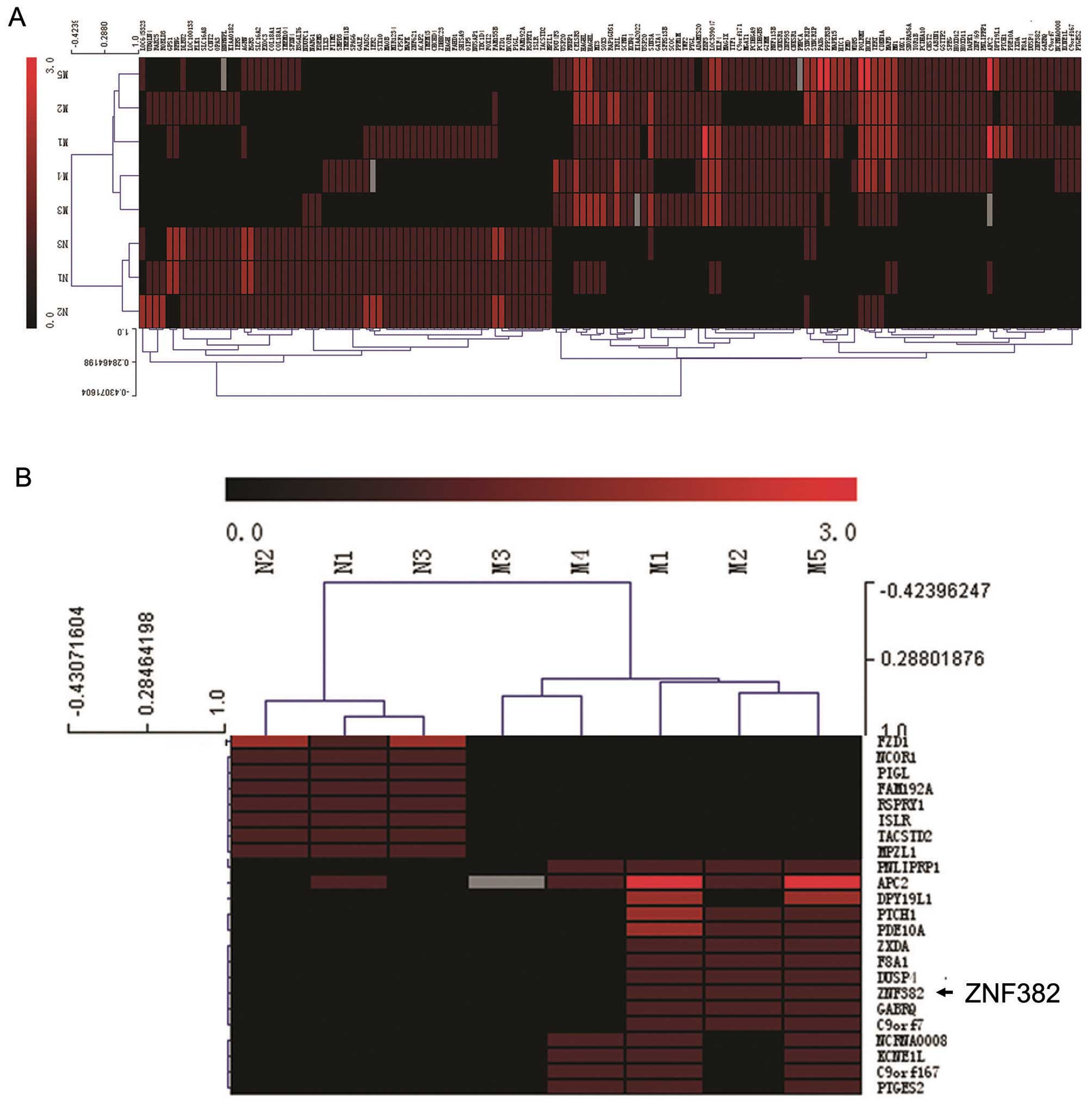
Arrays (Fig. 1A). We found that
the ZNF382 promoter was hypermethylated in 3/5 (60%) pediatric AML
samples and in 0/3 (0%) normal bone marrow samples (Fig. 1B). Subsequent analyses of the
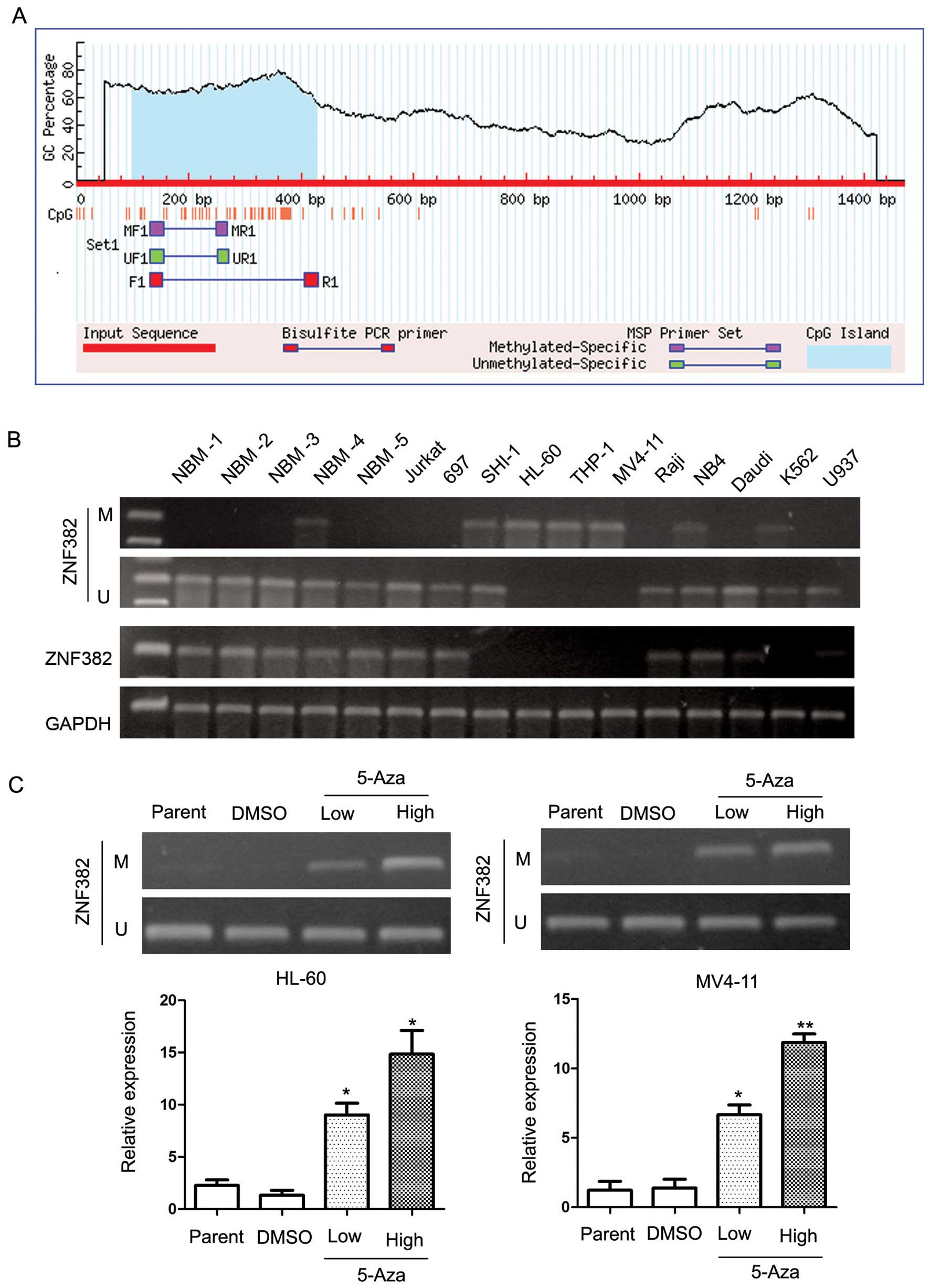
ZNF382 promoter sequence identified a large CpG island (Fig. 2A). MSP PCR assays were performed
to detect the methylation status of the ZNF382 promoter in the 11
leukemia cell lines. The MSP primer was designed using MethPrimer
(http://www.urogene.org/cgi-bin/methprimer/methprimer.cgi)
and encompassed the CpG islands of the ZNF382 promoter identified
in Fig. 2A. Our results revealed
that the ZNF382 promoter was hypermethylated in 6 leukemia cell
lines, including SHI-1, HL-60, THP-1, MV4-11, NB4 and K562 cells
(Fig. 2B). In addition, ZNF382
expression was detected in 5 cell lines (Jurkat, 697, Raji, NB4 and
Daudi) by RT-qPCR (Fig. 2B).
These results suggest that the downregulation of ZNF382 in AML
cells is a common phenomenon. To confirm the methylation of the
ZNF382 promoter, we treated the leukemia cell lines with the
demethylation reagent, 5-Aza. 5-Aza is an epigenetic modifier that
inhibits DNA methyltransferase activity through the remodeling
(opening) of chromatin, which results in DNA demethylation
(hypomethylation) and gene activation. Our results revealed that
treatment with 5-Aza significantly upregulated ZNF382 expression.
As shown in Fig. 2C, ZNF382
expression was upregulated by 11.1-fold in the HL-60 cells (5-Aza
14.83 vs. DMSO 1.33; P=0.023) and by 8.47-fold in the MV4-11 cells
(5-Aza 11.87 vs. DMSO 1.4; P<0.001). Furthermore, these results
were supported by MSP analyses, which revealed a change in the
methylation status of the ZNF382 promoter following treatment with
5-Aza. In summary, these results demonstrate that the ZNF382
promoter is consistently methylated in human myeloid leukemia cell
lines. Based on these findings, we suggest that the ZNF382 promoter
may be methylated in pediatric AML patients.
The ZNF382 promoter is methylated in
pediatric AML patients
We then examined the methylation status of the
ZNF382 promoter in pediatric AML samples and normal bone marrow
(NBM)/idiopathic thrombocytopenic purpura (ITP) control samples.
The aberrant methylation of ZNF382 was observed in 26.7% (28/105)
of the pediatric AML samples compared with 10% (2/20) of the
control samples (Fig. 3A). A
total of 3 control samples and 3 AML samples were further analyzed
by BGS (Fig. 3B). The BGS results
revealed that CpG islands in the ZNF382 promoter were methylated in
the AML samples (63.4, 70.9 and 60% in AML2#, AML4# and AML5#,
respectively). By contrast, CpG islands in the ZNF382 promoter were
unmethylated in the control samples (26.3, 29.1 and 29.1% in NBM6#,
NBM7# and NBM11#, respectively). Furthermore, these results were
supported by MSP assays.
 | Figure 3Zinc finger protein 382 (ZNF382) is
inactivated by promoter hypermethylation in pediatric acute myeloid
leukemia (AML). (A) Methylation-specific PCR (MSP) analysis of the
methylation status of ZNF382 shows aberrant methylation in
pediatric AML samples compared with normal bone marrow/idiopathic
thrombocytopenic purpura (NBM/ITP) control samples. M and U
represent MSP results using primer sets for methylated and
unmethylated ZNF382 genes, respectively. (B) A total of 3 NBM
samples and 3 AML samples were analyzed by bisulfite genomic
sequencing (BSG), ●, methylated cytosines; ○, unmethylated
cytosines. (C) The results revealed that the CpG islands in the
ZNF382 promoter were methylated in the AML samples (63.4, 70.9 and
60.0% in AML2#, AML4# and AML5#, respectively). By contrast, the
CpG islands of ZNF382 promoter in the NBM samples were unmethylated
(26.3, 29.1 and 29.1% in NBM6#, NBM7# and NBM11#,
respectively). |
ZNF382 expression is downregulated by
promoter methylation in pediatric AML patients
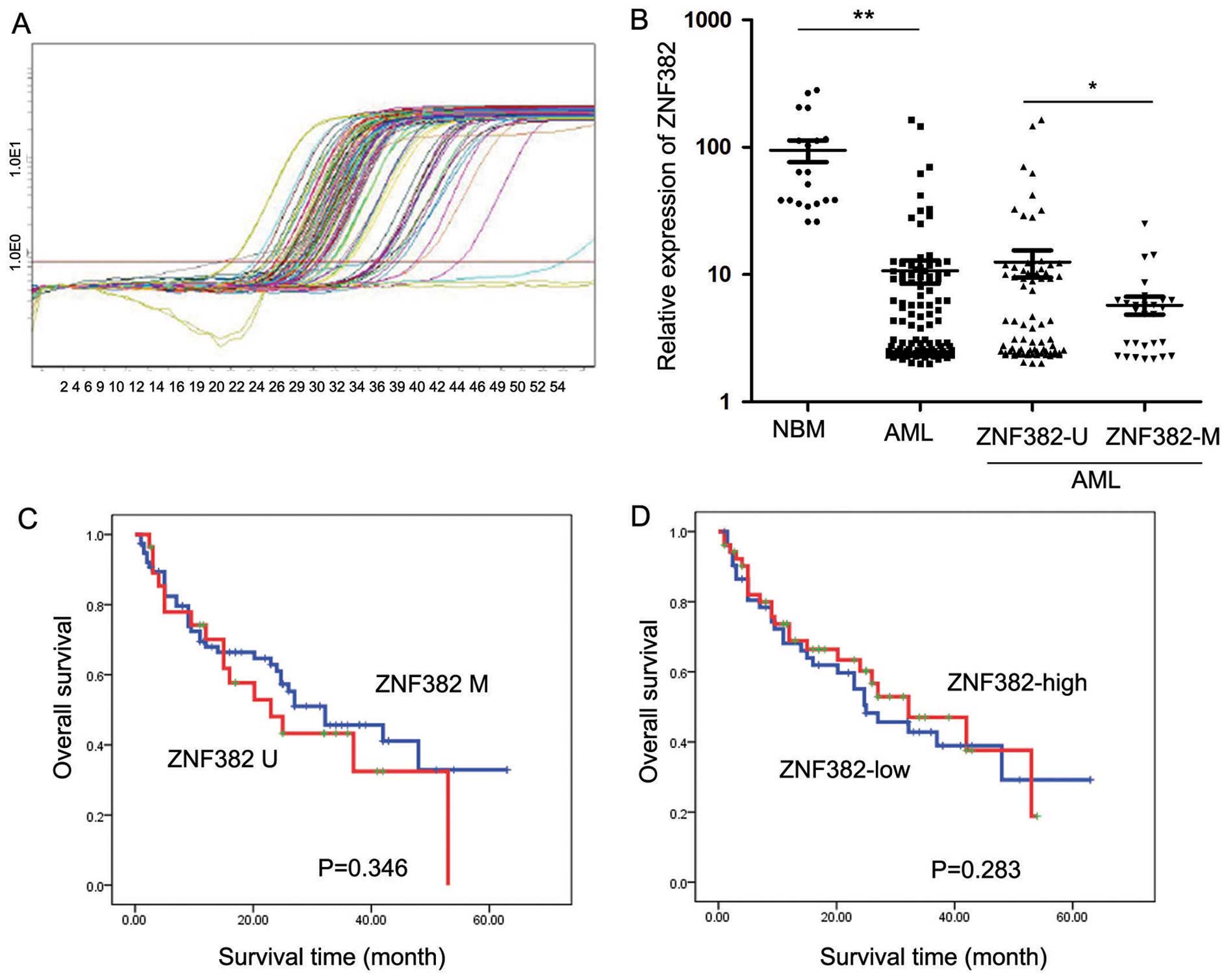
The transcript levels of ZNF382 were examined in 105
pediatric AML patients by RT-qPCR (Fig. 4C). As shown in Fig. 4D, ZNF382 expression was
significantly decreased in the 105 AML patients (10.73±23.01;
P<0.001) compared with the 20 control samples (94.74±81.62).
Patients exhibiting ZNF382 methylation (5.77±4.90, n=28) showed
lower ZNF382 transcript levels compared to the patients without
ZNF382 methylation (12.53±26.53, P=0.043; n=77). Furthermore, AML
patients with and without ZNF382 methylation presented with
significantly lower ZNF382 transcript levels compared with the
controls (Fig. 4D). The
prognostic significance of ZNF382 expression was assessed in 105
pediatric AML patients with clinical follow-up records. We found no
significant association between ZNF382 expression and patient age,
gender, FAB scores or cytogenetics (Table I). Kaplan-Meier survival analysis
of the 105 pediatric AML patients revealed similar survival times
in tumors with high or low ZNF382 expression (P=0.283) (Table III and Fig. 4B). Furthermore, multivariate
analysis revealed that ZNF382 promoter methylation failed to be an
independent prognostic factor in pediatric AML (P=0.832; Table IV).
 | Table IAssociation of ZNF382 expression with
clinicopathological characteristics in the 105 pediatric AML
samples. |
Table I
Association of ZNF382 expression with
clinicopathological characteristics in the 105 pediatric AML
samples.
| Clinical
variables | No. of
patients | ZNF382 expression
(n) | P-value |
|---|
|
|---|
| Low | High |
|---|
| Gender |
| Male | 42 | 20 | 22 | 0.633 |
| Female | 63 | 33 | 30 | |
| Age (years) |
| <6 | 60 | 34 | 26 | 0.143 |
| ≥6 | 45 | 19 | 26 | |
| Leukocytes
(/μl) |
| >10,000 | 61 | 30 | 31 | 0.755 |
| ≤10,000 | 44 | 23 | 21 | |
| FAB score |
| M1-M6 | 93 | 45 | 48 | 0.233 |
| M7 | 12 | 8 | 4 | |
| Cytogenetics |
| Favorable | 50 | 25 | 25 | 0.639 |
| Intermediate | 27 | 12 | 15 | |
| Unfavorable | 28 | 16 | 12 | |
| MRD |
| <0.25% | 49 | 24 | 25 | 0.774 |
| ≥0.25% | 56 | 29 | 27 | |
| Table IIIAssociation of ZNF382 expression and
promoter methylation with Kaplan-Meier survival in the 105
pediatric AML samples. |
Table III
Association of ZNF382 expression and
promoter methylation with Kaplan-Meier survival in the 105
pediatric AML samples.
| Variable | No. of
patients | Over survival
Median ± SE | P-value |
|---|
| Cytogenetics |
| Favorable | 50 | 46.664±3.717 | <0.001 |
| Intermediate | 27 | 29.220±3.188 | |
| Unfavorable | 28 | 11.161±1.827 | |
| FAB score |
| M1-M6 | 93 | 36.113±2.885 | <0.001 |
| M7 | 12 | 8.542±1.820 | |
| Leukocytes
(/μl) |
| >10,000 | 61 | 30.220±2.974 | 0.803 |
| ≤10,000 | 44 | 33.631±4.063 | |
| MRD |
| <0.25% | 49 | 53.627±3.151 | <0.001 |
| ≥0.25% | 56 | 18.893±2.425 | |
| ZNF382
expression |
| Low <4.03 | 52 | 32.020±3.661 | 0.283 |
| High ≥4.03 | 53 | 31.571±3.306 | |
| ZNF382
methylation |
| Negative | 77 | 34.215±3.236 | 0.346 |
| Positive | 28 | 28.051±4.275 | |
| Table IVCox multivariate analysis of ZNF382
expression, promoter methylation and clinicopathological
characteristics in pediatric AML. |
Table IV
Cox multivariate analysis of ZNF382
expression, promoter methylation and clinicopathological
characteristics in pediatric AML.
| Variable | Odds ratio | EXP (B) 95% CI | P-value |
|---|
| Cytogenetics |
| Fav. vs. inter and
unfav. | 4.569 | 2.216
(1.068–4.595) | 0.033 |
| MRD |
| <0.25 vs.
≥0.25% | 16.575 | 6.255
(2.588–15.119) | 0.000 |
| Leukocytes
(/μl) |
| >10,000 vs.
≤10,000 | 0.083 | 1.088
(0.615–1.925) | 0.773 |
| FAB score |
| M7 vs. M1-M6 | 9.265 | 3.113
(1.498–6.469) | 0.002 |
| ZNF382
expression |
| Low vs. high | 0.045 | 0.939
(0.527–1.675) | 0.832 |
| ZNF382
methylation |
| Negative vs.
positive | 1.326 | 0.690
(0.367–1.298) | 0.249 |
Promoter methylation of ZNF382 does not
correlate with survival in pediatric AML
The prognostic significance of ZNF382 promoter
methylation was assessed in 105 cases of pediatric AML patients
with clinical follow-up records. Table II shows that ZNF382 promoter
methylation correlated with minimal residual disease (MRD;
P=0.025). We found no significant differences in clinical
characteristics, such as gender, age, FAB scores or cytogenetics
between the patients with or without ZNF382 methylation (Table II). Kaplan-Meier survival
analysis revealed similar survival times in the samples with ZNF382
promoter methylation (P=0.346) (Table III and Fig. 4A). Furthermore, multivariate
analysis revealed that ZNF382 promoter methylation failed to be an
independent prognostic factor in pediatric AML (P=0.249) (Table IV).
| Table IIAssociation of ZNF382 promoter
methylation with clinicopathological characteristics in the 105
pediatric AML samples. |
Table II
Association of ZNF382 promoter
methylation with clinicopathological characteristics in the 105
pediatric AML samples.
| Clinical
variables | No. of
patients | ZNF382 methylation
(n) | P-value |
|---|
|
|---|
| Negative | Positive |
|---|
| Gender |
| Male | 42 | 30 | 12 | 0.719 |
| Female | 63 | 47 | 16 | |
| Age (years) |
| <6 | 60 | 42 | 18 | 0.372 |
| ≥6 | 45 | 35 | 10 | |
| Leukocytes
(/μl) |
| >10,000 | 61 | 43 | 18 | 0.438 |
| ≤10,000 | 44 | 34 | 10 | |
| FAB score |
| M1-M6 | 93 | 71 | 22 | 0.052 |
| M7 | 12 | 6 | 6 | |
| Cytogenetics |
| Favorable | 50 | 36 | 14 | 0.955 |
| Intermediate | 27 | 20 | 7 | |
| Unfavorable | 28 | 21 | 7 | |
| MRD |
| <0.25% | 49 | 41 | 8 | 0.025 |
| ≥0.25% | 56 | 36 | 20 | |
In summary, our results demonstrated that the ZNF382
promoter was significantly methylated in leukemia cell lines. The
aberrant methylation of ZNF382 was observed in 26.7% (28/105) of
the pediatric AML samples compared with 10% (2/20) of the control
samples. The expression of ZNF382 was significantly lower in the
pediatric AML samples compared with the control samples, and
patients with ZNF382 methylation had lower ZNF382 transcript levels
compared to the patients with no ZNF382 methylation. Furthermore,
ZNF382 promoter methylation correlated with MRD, and Kaplan-Meier
survival analysis revealed similar survival times in samples
harboring ZNF382 promoter methylation. Moreover, multivariate
analysis revealed that ZNF382 promoter methylation failed to be an
independent prognostic factor in pediatric AML.
Discussion
Aberrant DNA methylation patterns are a
characteristic feature of cancer, including myeloid malignancies,
such as AML. In AML, the presence of common methylation patterns in
a few genes, such as p15 and E-cadherin has been
independently described by several groups across larger patient
cohorts (26,27). The progression from
myelodysplastic syndrome to AML has been associated with increased
aberrant DNA methylation. Genome-wide methylation patterns in AML
have, however, thus far only been addressed in a few studies
(4,28). In particular, to the best of our
knowledge, whole-genome bisulfite sequencing analyses in AML are
limited. DNA methylation coupled with the H3K9me3-mediated gene
silencing of ZNF genes is widespread, occurring at individual ZNF
genes on multiple chromosomes and across ZNF gene family clusters
(29). In the present study,
promoter methylation in pediatric AML was analyzed using NimbleGen
Human DNA Methylation 385K Promoter plus CpG Island Arrays. This
approach revealed significant differences in the methylation status
of genes between pediatric AML and normal bone marrow samples. Our
results suggested that the ZNF382 promoter was hypermethylated in
AML. The ZNF382 promoter was hypermethylated in 3/5 (60%) of the
pediatric AML samples and in 0/3 (0%) of the normal bone marrow
samples.
ZNF382 is a functional tumor suppressor that is
frequently methylated in multiple carcinomas, including
nasopharyngeal, esophageal, colon, gastric and breast cancer
(24). A genome-wide analysis of
promoter associated CpG island methylation in acute lymphoblastic
leukemia (ALL) has revealed DNA methylation of ZNF382 in primary
ALL samples (30). To the best of
our knowledge, this is the first study to determine the expression
of ZNF382 and promoter methylation status in pediatric AML.
The molecular function of ZNF382 has been
investigated in multiple carcinomas. As previously demonstrated,
the ectopic expression of ZNF382 in silenced tumor cells
significantly inhibits clonogenicity, proliferation and induces
apoptosis. ZNF382 inhibits NF-κB and AP-1 signaling and
downregulates the expression of multiple oncogenes, including
MYC, microphthalmia-associated transcription factor
(MITF), high mobility group AT-hook 2 (HMGA2)
and cyclin-dependent kinase 6 (CDK6), as well as the NF-κB
upstream factors, including signal transducer and activator of
transcription (STAT)3, STAT5B, inhibitor of DNA binding 1, dominant
negative helix-loop-helix protein (ID1) and inhibitor of kappa
light polypeptide gene enhancer in B-cells, kinase epsilon (IKBKE),
most likely through heterochromatin silencing (24). ZNF family proteins play important
roles in recognition for methylated CpG. Two recent structures of
C2H2 ZNF proteins in complex with methylated DNA reveal a common
recognition mode for 5-methylcytosine (5 mC) that involves a 5
mC-Arg-G triad. In the two ZNF proteins, an arginine that precedes
the first Zn-binding histidine (RH motif) can interact with a 5
mCpG or TpG dinucleotide. RH-ZNF motifs provide specificity for 5
mCpG, whereas the neighboring ZNFs recognize the surrounding DNA
sequence context (31). The ZNF
protein, Kaiso, recognizes methylated and specific DNA sequences.
Kaiso is a bifunctional Cys(2)His(2) ZNF protein implicated in
tumor cell proliferation. Kaiso binds to both methylated CpG (mCpG)
sites and specific unmethylated DNA motifs (TCCTGCNA) and represses
transcription by recruiting chromatin remodeling corepression
machinery to target genes (32).
The ZNF proteins, zinc finger and BTB domain containing 4 (ZBTB4)
and zinc finger and BTB domain containing 38 (ZBTB38), can also
recognize methylated DNA. Kaiso, ZBTB4 and ZBTB38 can bind
methylated DNA in a sequence-specific manner (33), suggesting that many other ZNF
proteins, including ZNF382, may also have a similar function.
Currently, the molecular function of ZNF382 in pediatric AML
remains unknown and further investigations are required to
elucidate the role of ZNF382 in pediatric leukemia. The epigenetic
inactivation of ZNF382 by promoter hypermethylation can be observed
in both AML cell lines and pediatric AML samples. Therefore, our
study suggests that ZNF382 may be considered a putative tumor
suppressor gene in pediatric AML. In addition, our findings
indicate that ZNF382 promoter methylation correlates with MRD.
Furthermore, Kaplan-Meier survival analysis revealed similar
survival times in samples with ZNF382 promoter methylation.
However, further studies focusing on the mechanisms responsible for
ZNF382 methylation in pediatric leukemia are required.
Acknowledgements
This study was supported by grants from the National
Key Basic Research Program no. 2010CB933902, grants from key
medical subjects of Jiangsu province (XK201120), Innovative team of
Jiangsu province (LJ201114, LJ201126), Special Clinical Medical
Science and Technology of Jiangsu province (BL2012050, BL2013014),
Key Laboratory of Suzhou (SZS201108, SZS201307), National Natural
Science Foundation (81100371, 81370627, 81300423, 81272143).
Natural Science Foundation of Jiangsu province no. BK2011308,
Universities Natural Science Foundation of Jiangsu Province no.
11KJB320014 and Talent’s subsidy project in Science and Education
of Department of Public Health of Suzhou City no. SWKQ1020, and
Major scientific and technological special project for ‘significant
new drugs creation’ no. 2012ZX09103301-040.
References
|
1
|
Estey E and Dohner H: Acute myeloid
leukaemia. Lancet. 368:1894–1907. 2006. View Article : Google Scholar
|
|
2
|
Wlodarska I, Mecucci C, Baens M, Marynen P
and van den Berghe H: ETV6 gene rearrangements in hematopoietic
malignant disorders. Leuk Lymphoma. 23:287–295. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Plass C, Oakes C, Blum W and Marcucci G:
Epigenetics in acute myeloid leukemia. Semin Oncol. 35:378–387.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Figueroa ME, Lugthart S, Li Y, et al: DNA
methylation signatures identify biologically distinct subtypes in
acute myeloid leukemia. Cancer Cell. 17:13–27. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Wang N, Pan J, Cao L, et al: The
expression and clinical significance of miR-203 in pediatric acute
leukemia. Zhonghua Xue Ye Xue Za Zhi. 34:777–781. 2013.(In
Chinese).
|
|
6
|
Jian P, Yan WS, Chao SL, et al: Promoter
of TFPI-2 is hypermethylated in Chinese pediatric acute myeloid
leukemia. J Pediatr Hematol Oncol. 34:43–46. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Rodrigues EF, Santos-Reboucas CB,
Goncalves Pimentel MM, et al: Epigenetic alterations of p15(INK4B)
and p16(INK4A) genes in pediatric primary myelodysplastic syndrome.
Leuk Lymphoma. 51:1887–1894. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Ekmekci CG, Gutierrez MI, Siraj AK, Ozbek
U and Bhatia K: Aberrant methylation of multiple tumor suppressor
genes in acute myeloid leukemia. Am J Hematol. 77:233–240. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Britschgi C, Jenal M, Rizzi M, et al: HIC1
tumour suppressor gene is suppressed in acute myeloid leukaemia and
induced during granulocytic differentiation. Br J Haematol.
141:179–187. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Voso MT, Scardocci A, Guidi F, et al:
Aberrant methylation of DAP-kinase in therapy-related acute myeloid
leukemia and myelodysplastic syndromes. Blood. 103:698–700. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sahu GR, Mishra R, Nagpal JK and Das BR:
Alteration of p73 in acute myelogenous leukemia. Am J Hematol.
79:1–7. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Tao YF, Jian N, Jun L, et al: The promoter
of miR-663 is hypermethylated in Chinese pediatric acute myeloid
leukemia (AML). BMC Med Genet. 14:742013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Paz-Priel I and Friedman A: C/EBPα
dysregulation in AML and ALL. Crit Rev Oncog. 16:93–102. 2011.
|
|
14
|
Tao YF, Pang L, Du XJ, et al: Differential
mRNA expression levels of human histone-modifying enzymes in normal
karyotype B cell pediatric acute lymphoblastic leukemia. Int J Mol
Sci. 14:3376–3394. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Issa JP: CpG island methylator phenotype
in cancer. Nat Rev Cancer. 4:988–993. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Boultwood J and Wainscoat JS: Gene
silencing by DNA methylation in haematological malignancies. Br J
Haematol. 138:3–11. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gibbons RJ, McDowell TL, Raman S, et al:
Mutations in ATRX, encoding a SWI/SNF-like protein, cause diverse
changes in the pattern of DNA methylation. Nat Genet. 24:368–371.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Klose RJ and Bird AP: Genomic DNA
methylation: the mark and its mediators. Trends Biochem Sci.
31:89–97. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Baylin SB and Ohm JE: Epigenetic gene
silencing in cancer - a mechanism for early oncogenic pathway
addiction? Nat Rev Cancer. 6:107–116. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Flach J, Dicker F, Schnittger S, et al: An
accumulation of cytogenetic and molecular genetic events
characterizes the progression from MDS to secondary AML: an
analysis of 38 paired samples analyzed by cytogenetics, molecular
mutation analysis and SNP microarray profiling. Leukemia.
25:713–718. 2011. View Article : Google Scholar
|
|
21
|
Gao S, Hsieh CL, Zhou J and Shemshedini L:
Zinc Finger 280B regulates sGCα1 and p53 in prostate cancer cells.
PLoS One. 8:e787662013.PubMed/NCBI
|
|
22
|
Wang T, Wang XG, Xu JH, et al:
Overexpression of the human ZNF300 gene enhances growth and
metastasis of cancer cells through activating NF-κB pathway. J Cell
Mol Med. 16:1134–1145. 2012.PubMed/NCBI
|
|
23
|
Wang Y, Ye X, Zhou J, et al: A novel human
KRAB-related zinc finger gene ZNF425 inhibits mitogen-activated
protein kinase signaling pathway. BMB Rep. 44:58–63. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cheng Y, Geng H, Cheng SH, et al: KRAB
zinc finger protein ZNF382 is a proapoptotic tumor suppressor that
represses multiple oncogenes and is commonly silenced in multiple
carcinomas. Cancer Res. 70:6516–6526. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Olek A, Oswald J and Walter J: A modified
and improved method for bisulphite based cytosine methylation
analysis. Nucleic Acids Res. 24:5064–5066. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bullinger L, Ehrich M, Dohner K, et al:
Quantitative DNA methylation predicts survival in adult acute
myeloid leukemia. Blood. 115:636–642. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Deneberg S, Grovdal M, Karimi M, et al:
Gene-specific and global methylation patterns predict outcome in
patients with acute myeloid leukemia. Leukemia. 24:932–941. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Cancer Genome Atlas Research Network.
Genomic and epigenomic landscapes of adult de novo acute myeloid
leukemia. N Engl J Med. 368:2059–2074. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Severson PL, Tokar EJ, Vrba L, Waalkes MP
and Futscher BW: Coordinate H3K9 and DNA methylation silencing of
ZNFs in toxicant-induced malignant transformation. Epigenetics.
8:1080–1088. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Kuang SQ, Tong WG, Yang H, et al:
Genome-wide identification of aberrantly methylated promoter
associated CpG islands in acute lymphocytic leukemia. Leukemia.
22:1529–1538. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Liu Y, Zhang X, Blumenthal RM and Cheng X:
A common mode of recognition for methylated CpG. Trends Biochem
Sci. 38:177–183. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Buck-Koehntop BA, Stanfield RL, Ekiert DC,
et al: Molecular basis for recognition of methylated and specific
DNA sequences by the zinc finger protein Kaiso. Proc Natl Acad Sci
USA. 109:15229–15234. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sasai N, Nakao M and Defossez PA:
Sequence-specific recognition of methylated DNA by human
zinc-finger proteins. Nucleic Acids Res. 38:5015–5022. 2010.
View Article : Google Scholar : PubMed/NCBI
|