1. Introduction
Cancer, particularly malignant tumors, is the
leading cause of mortality in developed countries and the second
leading cause of mortality in developing countries (1). For decades, advancements have been
made in traditional cancer treatment regimens, chiefly surgery,
chemotherapy and radiation. Although these treatment regimens have
had certain success, they are not entirely adequate or optimal for
tumors that have migrated to areas that are inaccessible by surgery
or chemotherapy/radiation is not permissible. Therefore,
immunotherapy, which may produce fewer side-effects and prevent
metastasis in comparison to traditional treatments, has become of
increasing interest in the area of cancer treatment.
In the field of immunotherapy, increasing attention
has been focused on the use of cancer vaccines that activate T
cells to treat growing tumors (2). The development of peptide-based
vaccines has taken >20 years. A vaccine specific for tumor
antigens may have wide application and utility in the prevention of
the recurrence in numerous different malignancies. In cancer
patients, the body masks tumor antigens by the addition of
carbohydrate moieties to avoid an uncontrolled autoimmune response;
however, in the process, the elimination of threatening tumor cells
is also impeded (3). Therefore,
the development of peptide-based vaccines that directly stimulate
the immune system would be highly significant.
Peptides, which are composed of several amino acids
and are absent of oncogenic potential, are antigenic epitopes
derived from tumor-associated antigens (TAA) or tumor-specific
antigens (TSA) (3,4). Peptide-based vaccines are designed
to elicit specific T cells against antigens selectively expressed
by tumor cells (5). In comparison
to traditional treatment, peptide-based vaccines significantly
prolonged the overall survival rate and spare normal tissue due to
its low toxic effect (6–8). Evidently, there is a large number of
cancer patients requiring the development of novel approaches for
immunotherapy.
2. Mechanism of antitumor immunity by
peptide-based vaccines
The application of peptide-based vaccines is based
on three distinct steps to create a specific antitumor immune
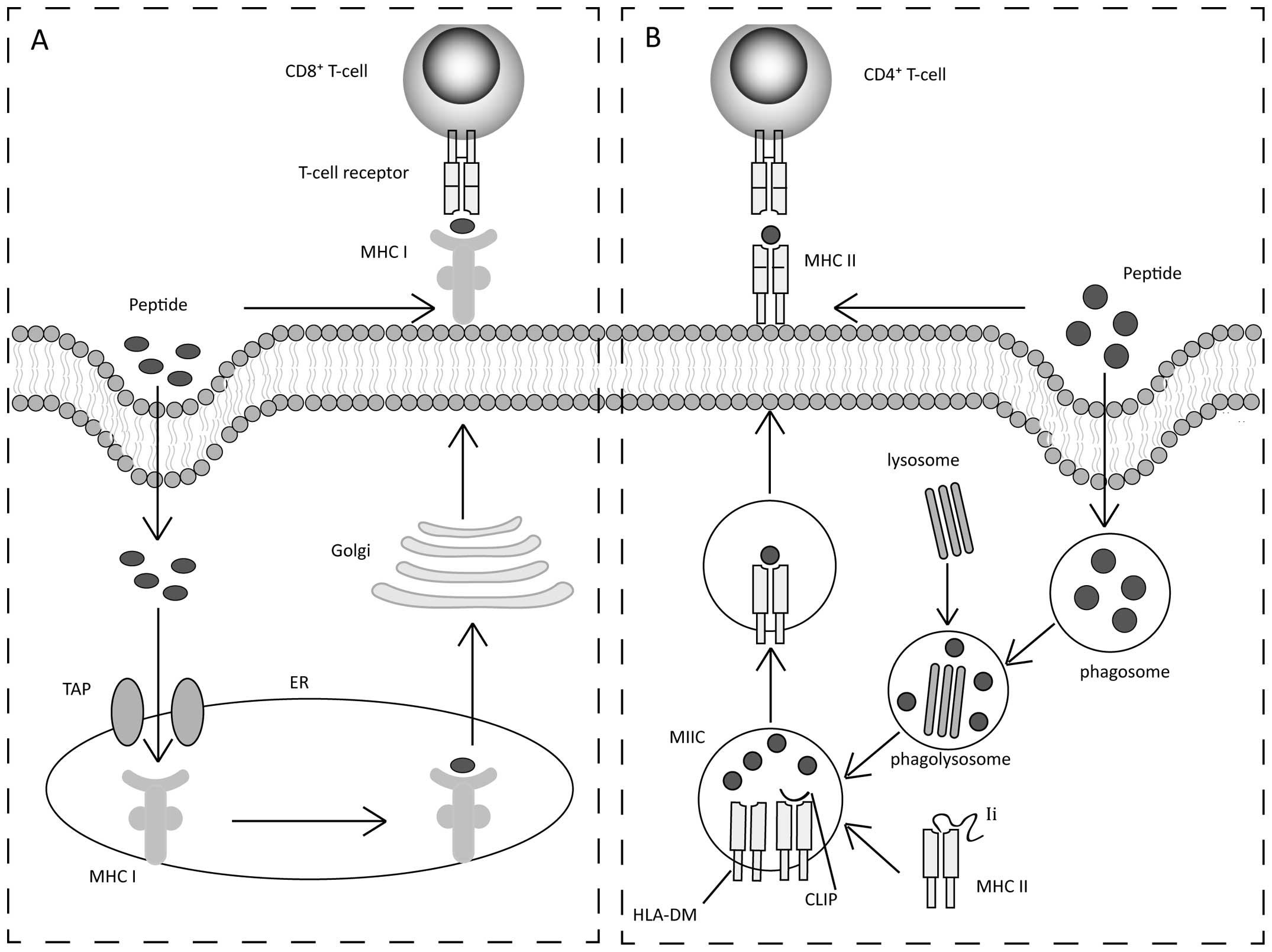
response. To initiate immunity, dentritic cells (DCs), which are
taken up exogenously as part of a therapeutic vaccine (9), differentiate into immunogenic mature
DCs (10). DCs enable the
presentation of peptides on major histocompatibility complex (MHC)
class I and II molecules (Fig.
1). Previously, the majority of peptide-based vaccines target
MHC class I peptides to stimulate cytotoxic T lymphocyte (CTL)
responses. MHC class I binds with peptides that are ~8–12 amino
acids in length (Fig. 1A)
(11). Peptides are transported
into the endoplasmic reticulum (ER) by a transporter associated
with antigen presentation (12).
Subsequently, peptide-MHC class I complexes go through the Golgi
and are delivered to the cell surface for recognition by cluster of
differentiation 8+ (CD8+) CTLs (13). Concurrently, a small proportion of
MHC class II-restricted peptides stimulate CD4+ T helper
cells. MHC class II-restricted peptides are generally 12–20 amino
acids in length (Fig. 1B)
(3). Extracellular peptides are
taken up by antigen-presenting cells and placed into phagosomes,
which fuse with lysosomes to form phagolysosomes (14). MHC class II assembled in the ER
associates with the invariant chain (Ii), and the Ii-MHC class II
complex is transported to phagolysosome and is known as the MHC
class II compartment (MIIC). In the MIIC, Ii is degraded, leaving a
residual class II-associated Ii peptide in the peptide-binding
groove (12). MHC class II
requires human leukocyte antigen (HLA)-DM (H2-DM in mice) to
completely expose the peptide-binding groove, binding with a
specific peptide (15).
Peptide-MHC class II complexes are delivered to the cell surface
for recognition by CD4+ T helper cells.
Subsequently, in lymphoid organs the peptide-loaded
DCs trigger specific T-cell responses (10). These T cells, such as
CD4+ T helper cells and CD8+ T cells, show
potent cytotoxic effects by two signals (16). One signal is from the T cell
receptor (TCR) interacting with peptide-MHC complexes on the DCs
(17). The other signal is from
the interaction of DC surface receptor CD80/CD86 with T-cell
co-stimulatory molecule, CD28 (17,18).
Finally, specific T cells must migrate to the tumor
microenvironment to cause the cytotoxic response. Considerable
knowledge has been obtained on CD8+ CTLs that have been
identified as the most powerful effector cells (19). CD4+ T helper 1 cells
(Th1) secrete several cytokines, such as IFN-γ, TNF-α and
interleukin-2 (IL-2) (20,21).
These cytokines exert direct antitumor immunity and antiangiogenic
effects (22). Notably, specific
CD4+ T helper cells have been found to enhance
CD8+ CTL recruitment and proliferation (21).
3. Peptide selection
During the past two decades, a majority of antigenic
peptides have been discovered. In principle, there are two types of
tumor antigens. One type is from TSAs, which are expressed
exclusively by tumors. TSAs generally arise from viral infections
or genetic mutations (23). The
second more common type is derived from TAAs. TAAs are found on
malignant and normal cells, but in a significantly higher number in
the former.
From TSAs
Peptide-based vaccines targeting viral oncogene
products are ideal candidates to elicit strong immune responses
without generating autoimmunity. As these viral oncoproteins are
not expressed in normal cells and their expression is required to
maintain the malignant phenotype, the viral protein is considered
as a potential target for cancer immunotherapy. Recently, a variety
of viral oncoproteins in virus-associated cancers have been used as
vaccines to induce T-cell responses. For instance, injection of
human papillomavirus type 16 E5 peptide + CpG resulted in strong
T-cell immunity and inhibited tumor growth, whilst prolonging the
survival time in animals with cervical cancer (24). Similarly, two recombinant
Epstein-Barr virus antigenic peptides, EBNA1 fused with LMP2,
boosted T-cell immunity and was proven to be safe and had low
immunogenicity in the phase I clinical trial for nasopharyngeal
carcinoma (7).
The antigens occurring in a number of different
proteins expressed by tumor cells are the result of genetic
mutations. A controversy exists over the idea that a single human
tumor, as in a mouse system, can express multiple mutated antigens
and generate new ones during progression, thereby making their
characterization even more complex. However, in the last few years,
the situation has slowly changed. Several studies (69–79) have described such antigens as
peptide epitopes recognized by T cells in combination with MHC
class I and II in human tumors, such as melanoma, non-small cell
lung cancer, renal cancer and head/neck cancer (Table I). The presumed advantages of
mutated antigens are based on the potential to be recognized as
non-self by the immune system and their potential resistance to
negative selection if the mutated protein participates in cell
survival (25).
 | Table IAntigens from genetic mutation
recognized by HLA class I and class II restricted T cells. |
Table I
Antigens from genetic mutation
recognized by HLA class I and class II restricted T cells.
| Antigen | Tumor | Refs. |
|---|
| HLA class
I-restricted |
| β-catenin | Melanoma | (69) |
| CDK-4 | Melanoma | (70) |
| MART-2 | Melanoma | (71) |
| MUM-1/2 | Melanoma | (72) |
| MUM-3 | Melanoma | (73) |
| HSP70-2 | Renal cancer | (74) |
| Caspase-8 | Head/neck
cancer | (75) |
| p21/ras | Pancreatic,
colorectal, lung cancer | (76) |
| HLA class
II-restricted |
| p53 | Head/neck
cancer | (77) |
| TPI | Melanoma | (78) |
| CDC27 | Melanoma | (79) |
From TAAs
TAAs can be divided into four major categories: i)
Differentiation antigens; ii) cancer/testis antigens shared by germ
and tumor cells; iii) overexpressed antigens, such as normal
proteins whose expression is upregulated in tumor cells; and iv)
universal tumor antigens.
Differentiation-antigens
These antigens are expressed by the normal tissue
and tumor originating from these tissues. The majority of these
antigens have been applied to treat melanoma, such as melanoma Ag
recognized by T cells (MART-1)/Melan A, gp100, tyrosinase,
tyrosinase-related protein (TRP-1) and TRP-2. Gp100, which were
initially identified, were reported as non-mutated differentiation
antigens expressed by a melanocytic lineage, including normal
melanocytes, pigmented retinal cells and melanomas, but not in
other normal tissues or non-melanoma tumors. In a current clinical
trial with tumor-free lymph nodes of stage I to III melanoma
patients, immunization with modified gp100209–2M peptide
without co-administration of CD4+ cell-restricted
antigens induced the effective expansion of tumor-reactive memory
CD8+ T cells with high proliferation potential (26). In another clinical study, the
response rate was higher and the progression-free survival rate was
longer with gp100:209–217 (210 M) peptide vaccine plus IL-2
compared to IL-2 alone in patients with metastatic melanoma
(27). These studies demonstrate
that differentiation proteins may be suitable targets for
immunotherapy.
Cancer/testis (CT) antigens
Expression of these antigens is restricted to human
germ cells within the testis and trophoblasts, and is also
expressed on a variety of types of human cancers. The antigens in
testis do not induce an immune response, as testis cells do not
express MHC class I. Since CT antigens are not expressed in normal
tissue, these antigens may be potentially useful targets for
tumor-specific immunotherapy. More than 40 antigens have been
identified as CT antigens, including melanoma antigen (MAGE), B
melanoma antigen, New York oesophageal squamous cell carcinoma 1
(NY-ESO-1) and G antigen 1. The first CT antigen was discovered
from a patient with melanoma who was identified as having cytotoxic
T cells that recognized autologous tumor cells (28). Through DNA-cloning methodology,
the gene encoding the tumor antigen MZ2-E was cloned and was termed
MAGE1. Fujie et al (29)
proposed that through using the MAGE-1-encoded peptide it was
possible to immunize an increased number of patients by means of
such peptide-based immunotherapeutic approaches to MAGE-1-positive
malignant tumors. Thus far, NY-ESO-1 is the most studied due to its
strong capacity to induce a tumor-specific immune response
(30). Previously, a completed
clinical study using co-administration of CpG 7909 and Montanide
ISA-51 with peptide NY-ESO-1 p157–165 showed the vaccine to be
capable of inducing CTLs, resulting in an extended survival time in
the majority of vaccinated patients (31). Currently, a number of clinical
trials treating various cancers are being performed using antigenic
peptide NY-ESO-1 combined with differing adjuvants.
Overexpressed-antigens
In healthy tissues, these antigens are expressed at
low levels on the surface of normal cells. Conversely, in the
majority of cancers these antigens are overexpressed but with no
preferential expression on certain tumor types, involving human
epidermal growth factor receptor-2 (HER-2), human mucin 1 (MUC1)
and cyclin B1. In humans, the HER-2 protein is expressed during
fetal development but is weakly detectable in the epithelial cells
of a number of normal tissues in adults. Overexpression of the
HER-2 protein has been identified in numerous types of human
cancers, such as breast, ovarian and non-small-cell lung cancer.
Immunizing patients with peptides derived from HER-2/neu protein
admixed with granulocyte-macrophage colony-stimulating factor
(GM-CSF) have been indicated to result in the generation of T-cell
immunity specific for the HER-2/neu (32). In addition, MUC1 has been studied
as a target for immunotherapy following a long developmental phase.
Transmembrane glycoprotein MUC1 is expressed on the apical surface
of polarized epithelial cells. However, in the majority of
epithelial malignancies, MUC1 is overexpressed and loses its
polarity of expression (33). In
pre-clinical studies using primates, MUC1 tandem repeat peptide
administered with LeIF elicited T helper cells and CTL responses
(34). This study showed the
peptide-based vaccine to be safe and to possibly be a vaccine that
induces MUC1-specific immune responses in patients with cancer.
Universal tumor antigens
Over the past decade, numerous TAAs have been
reported. However, for any particular TAA, expression is restricted
to several tumor types. To circumvent this, a new category of TAAs,
known as ‘universal tumor antigens,’ has been described. Such
universal tumor antigens are highly expressed in tumor cells of
different tissue origins with minimal expression in normal
counterparts. Survivin and telomerase have been reported to be
suitable as target universal tumor antigens for active immunization
of cancer patients. In a previous study, telomerase was expressed
in 85–90% of cancer patients and was an attractive universal tumor
antigen (35). Telomerase helps
to mediate functional telomeres, maintaining at the end of
chromosomes, and prevent cells from going into senescence,
particularly in cancer cells. The majority of human cells do not
express telomerase activity, but the majority of human tumors
exhibit strong activity (36). In
2006, Brunsvig et al (37)
conducted a phase I/II clinical study in patients with non-small
cell lung cancer (NSCLC), and the results demonstrated that
intradermal injections of GV1001 (hTERT: 611–626) was immunogenic,
safe and induced strong specific immune responses. Based on these
initial encouraging results, the study reported further clinical
studies of GV1001 in NSCLC patients. Vaccination with GV1001 was
indicated to exhibit low toxicity, induced considerable immune
response rate and established durable T-cell memory (38).
4. Strategies to induce and maintain T
cells
Peptide-based vaccines present certain objective
limitations. Free peptides have poor immunogenicity or no tertiary
structure, and thus are rapidly degraded by tissue and serum
peptidases prior to being loaded onto DCs. Recent studies indicate
that optimal strategies to induce and maintain T cells include
adjuvants, cytokines, HLA class II-restricted helper epitopes,
immune-modulating antibodies and low-affinity peptides combined
with high-affinity peptides, which are described in the
following.
Adjuvants
Peptide-based vaccines require additional adjuvant
to elicit efficient immunological response. Conjugates of peptides
with heat shock proteins (HSPs) (39,40) or ligands of toll-like receptors
(TLRs) (41–43) have been applied to a broad range
of vaccines as adjuvants to enhance the immunogenicity of peptides.
In 2000, the study by Cho et al (44) reported that HSP65 fusion proteins
stimulated DCs to increase expression of MHC (class I and II) and
co-stimulatory (B7.2) molecules. This study suggested a mechanism
in which the HSP fusion proteins induced CTLs to peptides without
requiring exogenous adjuvants or the participation of
CD4+ T cells (44).
However, TLRs may improve vaccination efficacy through activating
DC maturation, thereby upregulating the expression of MHC molecules
and enhancing antigen uptake. Khan et al (45) reported that TLR ligand-peptide
conjugates improved intracellular trafficking and processing
pathways, triggering optimal antigen presentation and T cells
priming.
Cytokines
Cytokines, such as GM-CSF and IL-12, are used as an
adjuvant in vaccines. GM-CSF increases the number of immature DCs
and migration, and it induces MHC class II expression and
activation by macrophages. In melanoma patients, subcutaneous
injection with GM-CSF modestly increased the immune response
against peptide vaccines (46).
The cytokine IL-12 augments antitumor immunity through promoting
Th1 cell differentiation and stimulating the production of IFN-γ
from CD4+ Th1 and CD8+ T cells (47). In preclinical and clinical
studies, a significant proportion of patients with resected
melanomas experienced an improved performance of the peptide
vaccines when it was combined with properly dosed IL-12 therapy
(48).
HLA class II-restricted helper
epitopes
HLA class II-restricted helper epitopes enhance
specific CTLs and generate T-cell memory (49–51). CD4+ T helper cells can
activate DCs to enhance antigen presentation, resulting in the
secretion of IL-2 and other cytokines from DCs that may help to
direct the immune response. Furthermore, cytokines secreted by Th1,
such as IL-2 and IFN-γ, are required for the generation of CTLs, as
well as in promoting CD8+ memory T-cell development.
IL-2 induces CTL activation and proliferation (52). Simultaneously, IFN-γ controls the
migration of CTLs (53). IL-2 is
essential for programming the ability of CD8+ memory T
cells to re-expand upon secondary infection in vivo
(52,54). Knutson et al (55) evaluated whether active
immunization with HER-2/neu helper peptides generated
CD4+ and CD8+ T-cell responses in patients.
Following vaccination, HER-2/neu-specific CD8+ T-cells
increased in the majority of patients. Additionally, the specific T
cells were able to lyse tumors and the immunity was long-lived.
Subsequently, Gritzapis et al (56) indicated that in comparison to
Her-2 (435–443) CTL peptide alone, mice vaccinated with Her-2
(435–443) plus Her-2 (776–790) exhibited longer lasting antitumor
responses and induced memory immunity. Thus, there is a rationale
for induction of CD4+ T cells with peptide-based
vaccines, either in combination with stimulation of CD8+
T cells or on their own.
Immune-modulating antibodies
Combining peptide-based vaccines with
immune-modulating antibodies may be a novel strategy to overcome
immune suppression (17,57,58). Several antibody therapies that are
either agonistic or inhibit receptors have shown benefits in cancer
treatment (59–61). Specific recognition by T cells is
a two-step process. In addition to the interaction of TCRs with
MHC-peptide complexes as the first signal for T cell recognition, a
second signal co-stimulates the receptors that determine whether
the T cell will become activated or anergic. These surface
co-stimulatory receptors transmit agonistic or inhibitory signals
through engagement of specific ligands. The co-stimulatory activity
of these receptors can be mimicked by antibodies modulating T-cell
proliferation, cytokine secretion and cytolysis (62). There are a number of known
agonistic receptors, including 4–1BB (CD137), OX40, CD27, GITR and
CD28 (62–64). The immune response against a
peptide vaccine combined with the systemic delivery of anti-4–1BB
antibodies resulted in considerably improved antitumor therapeutic
activity through CTLs, NK cells, neutrophils and IFN-γ (65). Receptors that serve as targets for
inhibitory antibodies include CTLA-4, PD-1 and BTLA. In one study,
a combination of peptide-based vaccines with blocking co-inhibitory
receptor signaling, resulted in antibody blockage of the
co-inhibitory receptors, CTLA-4 and PD-1, decreased T cell anergy
and allowed specific T cells to carry out their effector function
(66).
Low-affinity peptides combined with
high-affinity peptides
Peptides that bind to MHC molecules with high
affinity usually induce high-avidity T cells. Administration of
high-affinity peptides of exogenous antigens (such as viruses) is
necessary for their elimination. However, if the antigen is
autologous, high-affinity peptides possibly lead to tolerance.
Thus, for a peptide-based vaccine the most appropriate peptides may
be low-to-medium-affinity peptides (67). However, low-affinity peptides are
difficult to identify. In order to overcome this difficulty,
attempts have been made to raise the affinity of peptides for the
MHC through binding with high-affinity peptides. For example,
synthetic peptides, such as TERT578–592 combined with
two peptides derived from TERT, as high-affinity forms strongly
stimulated antitumor immune responses (22). In a parallel study, Disis et
al (68) synthesized four
peptides from HER-2/neu protein and two were shown to be avid
binders to HLA-A2.1, whereas the other two may be shown to elicit
peptide-specific CTL in vivo.
5. Conclusions
The significant advantage of peptide-based vaccines
is that they are easily synthesized, chemically stable entities and
notably, are absent of oncogenic potential. Antigenic peptides
offer a simple and flexible way to deal with the complexity of
tumor antigens through bypassing the requirements for antigen
processing.
As discussed previously, several promising
preclinical and clinical studies for peptide-based vaccines are
currently being carried out. Thus far however, there is no
peptide-based vaccine currently available on the market. There are
drawbacks that may hinder the peptide vaccine therapy. First, tumor
cells can downregulate MHC molecules or disable other components of
the antigen processing machinery. Second, peptides are HLA class I
and II restricted and, consequently, restrict the treatment of
patients in the clinical trials. The majority of previous cancer
vaccines target MHC class I-restricted peptides to stimulate CTL
responses. However, the clinical effect of CTL peptide-based
vaccines remains modest. Third, tumor cells may upregulate surface
ligands (such as PD-L1), which engage inhibitory receptors on the
surfaces of activated T cells (PD-1), to mediate T-cell anergy.
These drawbacks emphasize the requirement for significant changes
in the applications of peptide-based vaccines. Various
combinational approaches have been carried out to raise the
efficacy of peptide-based therapies.
In conclusion, there is evidence that peptide-based
vaccines have increased responses and prolonged survival rates in
patients with cancer. Firstly, designing a peptide-based vaccine
for cancer immunotherapy is challenging, involving the selection of
appropriate antigenic peptides. Strategies to increase the effects
to generate antitumor CD4+ cells that recognize MHC
class II-restricted peptides may have impact due to the importance
of CD4+ cells in enhancing activation and survival of
CD8+ effector cells, as well as generating
CD8+ memory T cells. Synthetic peptides that have
antigenic low-affinity combined with high-affinity peptides raise
the affinity of the peptides for the MHC and may significantly
enhance antitumor response. Furthermore, increasing numbers of
peptide-based vaccines with the co-administration of adjuvants,
cytokines or immunomodulatory antibodies have been shown to induce
and maintain immune responses. Finally, further studies are
required to focus on the synergy of peptide vaccination with
chemotherapy, involving larger studies providing evidence to
evaluate the curative effects ex vivo and in
vivo.
Acknowledgements
The present study was supported by grants from the
Natural Science Foundation of China (no. 81301946, 81301947 and
81202015), and Nature Science Foundation of Jiangsu Province (no.
BK2012146).
References
|
1
|
Yong X, Xiao YF, Luo G, et al: Strategies
for enhancing vaccine-induced CTl antitumor immune responses. J
Biomed Biotechnol. 2012:6050452012. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Rosenberg SA, Yang JC and Restifo NP:
Cancer immunotherapy: moving beyond current vaccines. Nat Med.
10:909–915. 2004. View
Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lazoura E and Apostolopoulos V: Rational
peptide-based vaccine design for cancer immunotherapeutic
applications. Curr Med Chem. 12:629–639. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Buhrman JD, Jordan KR, Munson DJ, Moore
BL, Kappler JW and Slansky JE: Improving antigenic peptide vaccines
for cancer immunotherapy using a dominant tumor-specific T cell
receptor. J Biol Chem. 288:33213–33225. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Milani A, Sangiolo D, Montemurro F,
Aglietta M and Valabrega G: Active immunotherapy in HER2
overexpressing breast cancer: current status and future
perspectives. Ann Oncol. 24:1740–1748. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tanaka T, Kitamura H, Inoue R, et al:
Potential survival benefit of anti-apoptosis protein:
survivin-derived peptide vaccine with and without interferon alpha
therapy for patients with advanced or recurrent urothelial cancer -
results from phase I clinical trials. Clin Dev Immunol.
2013:262967l2013. View Article : Google Scholar
|
|
7
|
Hui EP, Taylor GS, Jia H, et al: Phase I
trial of recombinant modified vaccinia ankara encoding epstein-BARR
viral tumor antigens in nasopharyngeal carcinoma patients. Cancer
Res. 73:1676–1688. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Asahara S, Takeda K, Yamao K, Maguchi H
and Yamaue H: Phase I/II clinical trial using HLA-A24-restricted
peptide vaccine derived from KIF20A for patients with advanced
pancreatic cancer. J Transl Med. 11:2912013. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Basha G, Lizée G, Reinicke AT, et al: MHC
class I endosomal and lysosomal trafficking coincides with
exogenous antigen loading in dendritic cells. PLoS One.
3:e32472008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mellman I, Coukos G and Dranoff G: Cancer
immunotherapy comes of age. Nature. 480:480–489. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Flutter B and Gao B: MHC class I antigen
presentation-recently trimmed and well presented. Cell Mol Immunol.
1:22–30. 2004.
|
|
12
|
Neefjes J, Jongsma MLM, Paul P and Bakke
O: Towards a systems understanding of MHC class I and MHC class II
antigen presentation. Nat Rev Immunol. 11:823–836. 2011.PubMed/NCBI
|
|
13
|
Van Kaer L: Major histocompatibility
complex class I-restricted antigen processing and presentation.
Tissue Antigens. 60:1–9. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Vyas JM, Van der Veen AG and Ploegh HL:
The known unknowns of antigen processing and presentation. Nat Rev
Immunol. 8:607–618. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Pieters J: MHC class II-restricted antigen
processing and presentation. Adv Immunol. 75:159–208. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Callan MF, Fazou C, Yang H, et al: CD8(+)
T-cell selection, function, and death in the primary immune
response in vivo. J Clin Invest. 106:1251–1261. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Vanneman M and Dranoff G: Combining
immunotherapy and targeted therapies in cancer treatment. Nat Rev
Cancer. 12:237–251. 2012. View
Article : Google Scholar : PubMed/NCBI
|
|
18
|
Shashidharamurthy R, Bozeman EN, Patel J,
Kaur R, Meganathan J and Selvaraj P: Immunotherapeutic strategies
for cancer treatment: a novel protein transfer approach for cancer
vaccine development. Med Res Rev. 32:1197–1219. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Vesely MD, Kershaw MH, Schreiber RD and
Smyth MJ: Natural innate and adaptive immunity to cancer. Annu Rev
Immunol. 29:235–271. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kennedy R and Celis E: Multiple roles for
CD4+ T cells in anti-tumor immune responses. Immunol
Rev. 222:129–144. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bos R and Sherman LA: CD4+
T-cell help in the tumor milieu is required for recruitment and
cytolytic function of CD8+ T lymphocytes. Cancer Res.
70:8368–8377. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Dosset M, Godet Y, Vauchy C, et al:
Universal cancer peptide-based therapeutic vaccine breaks tolerance
against telomerase and eradicates established tumor. Clin Cancer
Res. 18:6284–6295. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wei HJ, Wu AT, Hsu CH, et al: The
development of a novel cancer immunotherapeutic platform using
tumor-targeting mesenchymal stem cells and a protein vaccine. Mol
Ther. 19:2249–2257. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liao SJ, Deng DR, Zeng D, et al: HPV16 E5
peptide vaccine in treatment of cervical cancer in vitro and in
vivo. J Huazhong Univ Sci Technolog Med Sci. 33:735–742. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Palucka K, Banchereau J and Mellman I:
Designing vaccines based on biology of human dendritic cell
subsets. Immunity. 33:464–478. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Walker EB, Miller W, Haley D, Floyd K,
Curti B and Urba WJ: Characterization of the class I-restricted
gp100 melanoma peptide-stimulated primary immune response in
tumor-free vaccine-draining lymph nodes and peripheral blood. Clin
Cancer Res. 15:2541–2551. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Schwartzentruber DJ, Lawson DH, Richards
JM, et al: gp100 peptide vaccine and interleukin-2 in patients with
advanced melanoma. N Engl J Med. 364:2119–2127. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Knuth A, Wölfel T, Klehmann E, Boon T and
Meyer zum Buschenfelde KH: Cytolytic T-cell clones against an
autologous human melanoma: specificity study and definition of
three antigens by immunoselection. Proc Natl Acad Sci USA.
86:2804–2808. 1989. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Fujie T, Tahara K, Tanaka F, Mori M,
Takesako K and Akiyoshi T: A MAGE-1-encoded HLA-A24-binding
synthetic peptide induces specific anti-tumor cytotoxic T
lymphocytes. Int J Cancer. 80:169–172. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Simpson AJ, Caballero OL, Jungbluth A,
Chen YT and Old LJ: Cancer/testis antigens, gametogenesis and
cancer. Nat Rev Cancer. 5:615–625. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Karbach J, Gnjatic S, Bender A, et al:
Tumor-reactive CD8+ T-cell responses after vaccination
with NY-ESO-1 peptide, CpG 7909 and montanide ISA-51: association
with survival. Int J Cancer. 126:909–918. 2010.
|
|
32
|
Disis ML, Gooley TA, Rinn K, et al:
Generation of T-cell immunity to the HER-2/neu protein after active
immunization with HER-2/neu peptide-based vaccines. J Clin Oncol.
20:2624–2632. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Sangha R and Butts C: L-BLP25: a peptide
vaccine strategy in non small cell lung cancer. Clin Cancer Res.
13:s4652–s4654. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Barratt-Boyes SM, Vlad A and Finn OJ:
Immunization of chimpanzees with tumor antigen MUC1 mucin tandem
repeat peptide elicits both helper and cytotoxic T-cell responses.
Clin Cancer Res. 5:1918–1924. 1999.PubMed/NCBI
|
|
35
|
Bernhardt SL, Gjertsen MK, Trachsel S, et
al: Telomerase peptide vaccination of patients with non-resectable
pancreatic cancer: a dose escalating phase I/II study. Br J Cancer.
95:1474–1482. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kim NW, Piatyszek MA, Prowse KR, et al:
Specific association of human telomerase activity with immortal
cells and cancer. Science. 266:2011–2015. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Brunsvig PF, Aamdal S, Gjertsen MK, et al:
Telomerase peptide vaccination: a phase I/II study in patients with
non-small cell lung cancer. Cancer Immunol Immunother.
55:1553–1564. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Brunsvig PF, Kyte JA, Kersten C, et al:
Telomerase peptide vaccination in NSCLC: a phase II trial in stage
III patients vaccinated after chemoradiotherapy and an 8-year
update on a phase I/II trial. Clin Cancer Res. 17:6847–6857. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Ding Z, Ou R, Ni B, Tang J and Xu Y:
Cytolytic activity of the human papillomavirus type 16 E711-20
epitope-specific cytotoxic t lymphocyte is enhanced by heat shock
protein 110 in HLA-A*0201 transgenic mice. Clin Vaccine
Immunol. 20:1027–1033. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yang J, Zhang Y, Wang H, et al:
Vaccination with the repeat β-hCG C-terminal peptide carried by
heat shock protein-65 (HSP65) for inducing antitumor effects. Tumor
Biol. 33:1777–1784. 2012. View Article : Google Scholar
|
|
41
|
Koido S, Homma S, Okamoto M, et al:
Combined TLR2/4-activated dendritic/tumor cell fusions induce
augmented cytotoxic T lymphocytes. PLoS One. 8:e592802013.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Muraoka D, Kato T, Wang L, et al: Peptide
vaccine induces enhanced tumor growth associated with apoptosis
induction in CD8+ T cells. J Immunol. 185:3768–3776.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Speiser DE, Liénard D, Rufer N, et al:
Rapid and strong human CD8+ T cell responses to
vaccination with peptide, IFA, and CpG oligodeoxynucleotide 7909. J
Clin Invest. 115:739–746. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Cho BK, Palliser D, Guillen E, et al: A
proposed mechanism for the induction of cytotoxic T lymphocyte
production by heat shock fusion proteins. Immunity. 12:263–272.
2000. View Article : Google Scholar
|
|
45
|
Khan S, Bijker MS, Weterings JJ, et al:
Distinct uptake mechanisms but similar intracellular processing of
two different toll-like receptor ligand-peptide conjugates in
dendritic cells. J Biol Chem. 282:21145–21159. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Weber J, Sondak VK, Scotland R, et al:
Granulocyte- macrophage-colony-stimulating factor added to a
multipeptide vaccine for resected stage II melanoma. Cancer.
97:186–200. 2003. View Article : Google Scholar
|
|
47
|
Hamid O, Solomon JC, Scotland R, et al:
Alum with interleukin-12 augments immunity to a melanoma peptide
vaccine: correlation with time to relapse in patients with resected
high-risk disease. Clin Cancer Res. 13:215–222. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Lee P, Wang F, Kuniyoshi J, et al: Effects
of interleukin-12 on the immune response to a multipeptide vaccine
for resected metastatic melanoma. J Clin Oncol. 19:3836–3847.
2001.PubMed/NCBI
|
|
49
|
Izumoto S: Peptide vaccine. Adv Exp Med
Biol. 746:166–177. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
May RJ, Dao T, Pinilla-Ibarz J, et al:
Peptide epitopes from the wilms’ tumor 1 oncoprotein stimulate
CD4+ and CD8+ T cells that recognize and kill
human malignant mesothelioma tumor cells. Clin Cancer Res.
13:4547–4555. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Fujiki F, Oka Y, Tsuboi A, et al:
Identification and characterization of a WT1 (Wilms Tumor Gene)
protein-derived HLA-DRB1*0405-restricted 16-mer helper
peptide that promotes the induction and activation of WT1-specific
cytotoxic T lymphocytes. J Immunother. 30:282–293. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Pipkin ME, Sacks JA, Cruz-Guilloty F,
Lichtenheld MG, Bevan MJ and Rao A: Interleukin-2 and inflammation
induce distinct transcriptional programs that promote the
differentiation of effector cytolytic T cells. Immunity. 32:79–90.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Nakanishi Y, Lu B, Gerard C and Iwasaki A:
CD8+ T lymphocyte mobilization to virus-infected tissue
requires CD4+ T-cell help. Nature. 462:510–513. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Williams MA, Tyznik AJ and Bevan MJ:
Interleukin-2 signals during priming are required for secondary
expansion of CD8+ memory T cells. Nature. 441:890–893.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Knutson KL, Schiffman K and Disis ML:
Immunization with a HER-2/neu helper peptide vaccine generates
HER-2/neu CD8 T-cell immunity in cancer patients. J Clin Invest.
107:477–484. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Gritzapis AD, Voutsas IF, Lekka E,
Papamichail M and Baxevanis CN: Peptide vaccination breaks
tolerance to HER-2/neu by generating vaccine-specific FasL(+)
CD4(+) T cells: first evidence for intratumor apoptotic regulatory
T cells. Cancer Res. 70:2686–2696. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Wang Y, Wang XY, Subjeck JR, Shrikant PA
and Kim HL: Temsirolimus, an mTOR inhibitor, enhances anti-tumour
effects of heat shock protein cancer vaccines. Br J Cancer.
104:643–652. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Arens R, van Hall T, van der Burg SH,
Ossendorp F and Melief CJM: Prospects of combinatorial synthetic
peptide vaccine-based immunotherapy against cancer. Semin Immunol.
25:182–190. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Gray JC, French RR, James S, Al-Shamkhani
A, Johnson PW and Glennie MJ: Optimising anti-tumour CD8 T-cell
responses using combinations of immunomodulatory antibodies. Eur J
Immunol. 38:2499–2511. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Fransen MF, Sluijter M, Morreau H, Arens R
and Melief CJ: Local activation of CD8 T cells and systemic tumor
eradication without toxicity via slow release and local delivery of
agonistic CD40 antibody. Clin Cancer Res. 17:2270–2280. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Ascierto PA, Simeone E, Sznol M, Fu YX and
Melero I: Clinical experiences with anti-CD137 and anti-PD1
therapeutic antibodies. Semin Oncol. 37:508–516. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Croft M: The role of TNF superfamily
members in T-cell function and diseases. Nat Rev Immunol.
9:271–285. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Croft M: Co-stimulatory members of the
TNFR family: keys to effective T-cell immunity? Nat Rev Immunol.
3:609–620. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Topalian SL, Weiner GJ and Pardoll DM:
Cancer immunotherapy comes of age. J Clin Oncol. 29:4828–4836.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Sin JI, Kim H, Ahn E, et al: Combined
stimulation of TLR9 and 4.1BB augments Trp2 peptide
vaccine-mediated melanoma rejection by increasing Ag-specific CTl
activity and infiltration into tumor sites. Cancer Lett.
330:190–199. 2013. View Article : Google Scholar
|
|
66
|
Curran MA, Montalvo W, Yagita H and
Allison JP: PD-1 and CTLA-4 combination blockade expands
infiltrating T cells and reduces regulatory T and myeloid cells
within B16 melanoma tumors. Proc Natl Acad Sci USA. 107:4275–4280.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
67
|
Apostolopoulos V: Peptide-based vaccines
for cancer: are we choosing the right peptides? Expert Rev
Vaccines. 8:259–260. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Disis ML, Smith JW, Murphy AE, Chen W and
Cheever MA: In vitro generation of human cytolytic T-cells specific
for peptides derived from the HER-2/neu protooncogene protein.
Cancer Res. 54:1071–1076. 1994.PubMed/NCBI
|
|
69
|
Robbins PF, El-Gamil M, Li YF, et al: A
mutated beta-catenin gene encodes a melanoma-specific antigen
recognized by tumor infiltrating lymphocytes. J Exp Med.
183:1185–1192. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Wölfel T, Hauer M, Schneider J, et al: A
p16INK4a-insensitive CDK4 mutant targeted by cytolytic T
lymphocytes in a human melanoma. Science. 269:1281–1284. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Kawakami Y, Wang X, Shofuda T, et al:
Isolation of a new melanoma antigen, MART-2, containing a mutated
epitope recognized by autologous tumor-infiltrating T lymphocytes.
J Immunol. 166:2871–2877. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Coulie PG, Lehmann F, Lethé B, et al: A
mutated intron sequence codes for an antigenic peptide recognized
by cytolytic T lymphocytes on a human melanoma. Proc Natl Acad Sci
USA. 92:7976–7980. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Baurain JF, Colau D, van Baren N, et al:
High frequency of autologous anti-melanoma CTL directed against an
antigen generated by a point mutation in a new helicase gene. J
Immunol. 164:6057–6066. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Gaudin C, Kremer F, Angevin E, Scott V and
Triebel F: A hsp70-2 mutation recognized by CTl on a human renal
cell carcinoma. J Immunol. 162:1730–1738. 1999.PubMed/NCBI
|
|
75
|
Mandruzzato S, Brasseur F, Andry G, Boon T
and van der Bruggen P: A CASP-8 mutation recognized by cytolytic T
lymphocytes on a human head and neck carcinoma. J Exp Med.
186:785–793. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
76
|
Bristol JA, Schlom J and Abrams SI:
Development of a murine mutant ras CD8+ CTL peptide
epitope variant that possesses enhanced MHC class I binding and
immunogenic properties. J Immunol. 160:2433–2441. 1998.PubMed/NCBI
|
|
77
|
Couch ME, Ferris RL, Brennan JA, et al:
Alteration of cellular and humoral immunity by mutant p53 protein
and processed mutant peptide in head and neck cancer. Clin Cancer
Res. 13:7199–7206. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Pieper R, Christian RE, Gonzales MI, et
al: Biochemical identification of a mutated human melanoma antigen
recognized by CD4(+) T cells. J Exp Med. 189:757–766. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Wang RF, Wang X, Atwood AC, Topalian SL
and Rosenberg SA: Cloning genes encoding MHC class II-restricted
antigens: mutated CDC27 as a tumor antigen. Science. 284:1351–1354.
1999. View Article : Google Scholar : PubMed/NCBI
|