Introduction
Bone marrow (BM)-derived endothelial progenitor
cells (EPCs) play a crucial role in neovascularazation. In the past
decades, the regenerative potential of EPCs in the injured
endothelia, including myocardial ischemia (1), carotid artery injury (2) and limb ischemia (3), has been extensively investigated.
However, it is now known that the hostile wound environment in
settings, such as chronic wound healing and ischemic myocardial
infarction characterized by hypoxia, and increased inflammation and
increased free radicals, has an adverse effect on the survival and
function of transplanted EPCs, thereby compromising their full
therapeutic benefit (4). In
addition, there is a study that heart failure patients with high
circulating levels of tumor necrosis factor-α (TNF-α), a potent
pro-inflammatory cytokine, are associated with significantly lower
EPC counts as compared to patients treated with a TNF-α inhibitor
(5). Therefore, modulation of the
local tissue microenvironment by anti-inflammatory factors can
confer an improved stem cell survival and function, and improved
clinical efficacy.
Interleukin-10 (IL-10), a potent anti-inflammatory
cytokine, attenuates inflammatory response and suppresses various
pro-inflammatory mediators (6–8).
IL-10 plays a role not only in immunoregulation and inhibition of
pro-inflammatory cytokine synthesis, but also in directly
regulating the growth and survival of noninflammatory cells.
Several studies have demonstrated that BM-mononuclear cells (MNCs),
as well as mesenchymal stem cells (MSCs), have the ability to
immunoregulate and improve tissue repair through IL-10 secretion
(9–11). A further study by Krishnamurthy
et al (12) demonstrated
that IL-10 has a role on EPC mobilization following myocardial
injury. Therefore, we hypothesized that IL-10 modulates EPC biology
and enhances its function via activation of the signal transducer
and activator of transcription 3 (STAT3) signaling pathway.
Materials and methods
Major reagents
Lentivirus vector-IL-10-green fluorescent protein
(LV-IL-10-GFP) and LV-noncontain-GFP (LV-NC-GFP) were obtained from
Xi’An Kewei Biological Technology Company (Xi’an, China).
Horseradish peroxidase affinipure goat anti-rabbit immunoglobulin G
(E030120-02) and goat anti-mouse (E030110-02) were purchased from
EarthOx, LLC (San Francisco, CA, USA). The anti-matrix
metallopeptidase-9 (MMP-9) antibody (ab58803), anti-vascular
endothelial growth factor (VEGF) antibody (ab46154), anti-STAT3
antibody (ab11935) and anti-fluorescein isothiocyanate
(FITC)-conjugated von Willebrand factor (vWF) antibody (ab8822)
were purchased from Abcam (Cambridge, UK). Anti-phycoerythrin
(PE)-conjugated VEGF receptor (VEGFR2) antibody (YM-0565R) was
purchased from Shanghai Yanmeng Biological Technology Co.
(Shanghai, China). Phosphorylated-STAT3 (p-STAT3; #9145s) was
purchased from Cell Signaling Technology (Danvers, MA, USA).
Electrochemiluminescence (ECL) was obtained from Millipore
Corporation (Billerica, MA, USA).
Animals
Postnatal 7–9-day-old male Wistar rats were
purchased from the Academy of Military Medical Sciences (Beijing,
China). All the animals in the study were cared, used and treated
in strict accordance to the ARVO statement for the use of animals
in ophthalmic and vision research.
Isolation and culture of EPCs
BM-derived EPCs were cultured according to
established methods (13,14). In brief, postnatal 7–9-day-old
rats were sacrificed by cervical dislocation and the tibias were
extracted. The BM was flushed out of the tibias with sterile
phosphate-buffered saline (PBS) using a syringe, and MNCs were
isolated by Ficoll gradient centrifugation. Following washing twice
with PBS, MNCs were seeded on a human fibronectin (FN;
Sigma-Aldrich, St. Louis, MO, USA) coated 6-well plate at a density
of 5×106 cell/well and cultured in endothelial cell
growth medium-2 (EGM-2; Lonza, Walkersville, MD, USA) at 37°C, 5%
CO2. EGM-2 was changed every two days and non-adherent
cells were removed. After 7 days in culture, cells were identified
by fluorescent-activated cell sorting (FACS; Calibur flow
cytometer, Becton Dickinson, CA, USA) and immunofluorescence.
Characterization of BM-derived EPCs
Identification of EPCs was first by fluorescent
labeling, direct binding of FITC-conjugated Ulex europaeus
agglutinin (UEA-1; Sigma-Aldrich) and uptake of
1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine percholate
(DiI)-labeled acetylated low-density lipoprotein (acLDL; Biomedical
Technologies, Stoughton, MA, USA) were performed. For these assays,
the cells were first incubated with DiI-ac-LDL (10 μg/ml in
EBM-2 medium) at 37°C for 4 h and were subsequently fixed with 4%
paraformaldehyde (PFA) and counterstained with UEA-1 (10
μg/ml in 0.9% saline). Subsequently, images were captured
under an inverted fluorescence microscope (Olympus, Tokyo,
Japan).
For extracellular labeling of vWF and VEGFR2, cells
were fixed with 4% PFA for 30 mins and blocked in 5% bovine serum
albumin and labeled sequentially with anti-vWF antibody conjugated
to FITC and anti-VEGFR2 antibody conjugated to PE. The cells were
subsequently washed three times in PBS, counterstained with
diamidino-phenyl-indole for 5 mins. Following washing, images were
captured by fluorescence microscopy.
Lentiviral transduction
Following identification, EPCs were transfected with
LV-IL-10-GFP (EPC-LV-IL-10-GFP) and LV-NC-GFP (EPC-LV-NC-GFP) as
the control group, whereas the LV expressing enhanced GFP was used
to measure transduction efficiency. To achieve optimal gene
transfer, polybrene (1:1000; Biomedical Technologies) was used, and
the medium was changed 4 h later. Two days later, transfection
efficiency was observed under an inverted fluorescence microscope
and the transfected cells were used for all the experiments. Before
being used, a section of the cells were starved for 12 h and were
subsequently stimulated with recombinant rat TNF-α (PeproTech,
Rocky Hill, NJ, USA) at 10 ng/ml, and the medium was changed 12 h
after stimulation.
Assessment of TNF-α, IL-10, IL-8 and VEGF
by ELISA
The concentrations of TNF-α, IL-10, IL-8 and VEGF in
EGM-2 medium were measured by rat enzyme-linked immunosorbent assay
(ELISA) kits (R&D Systems, Inc., Minneapolis, MN, USA)
according to the manufacturer’s instructions. Subsequently, all the
groups of supernatants were collected and the assays for the levels
of TNF-α, IL-10, IL-8 and VEGF were performed.
Migration assay of IL-10-modified
EPCs
Cell migration was tested in Transwell 12-well
plates (Corning, New York, NY, USA). Cells were digested and
re-suspended in serum-free EGM-2 medium, and subsequently,
2×104 cells were loaded into the upper chambers. The
lower chamber was filled with EGM-2 medium. After 6 h of incubation
at 37°C, cells were stained with crystal violet and counted in 3
random fields (magnification, ×200) in each well.
Adhesion assay of IL-10-modified
EPCs
Cell adhesion was examined in 96-well plates coated
overnight (4°C) with FN. Cells were digested, re-suspended and
plated at a concentration of 5×104 cells/well in 100 ml
of serum-free EGM-2 medium. Adhesion was carried out for 4 h at
37°C. Removal of non-adherent cells was achieved by two washing
steps with serum-free EGM-2 medium. Adhesion was quantified by
counting adherent cells in 3 random fields (magnification, ×200) in
each well.
Tube formation assay of IL-10-modified
EPCs
Serum-starved cells (1×105) were seeded
onto a matrigel (BD Bioscience, San Jose, CA, USA)-coated plate in
EGM-2 medium and incubated at 37°C for 10 h. To quantify the length
of newly-formed tubes, photomicrographs in 3 random fields
(magnification, ×100) per well were captured, and the total tubule
lengths were measured using WimTube Image software (Wimasis GmbH,
Munich, Germany).
Cell cycle analysis of IL-10-modified
EPCs by flow cytometry
Cells (1×106) were digested, re-suspended
and added to ice-cold methanol for fixation and stored at 4°C.
Samples were warmed to room temperature and rehydrated by rinsing
twice in PBS. Samples were centrifuged (250 × g, 10 min at 4°C) and
incubated in 500 μl PI/RNase staining buffer (BD
Biosciences) for 15 min. Then the cells were analyzed on a flow
cytometry. The data were analyzed by FlowJo 7.6 software (Tree
Star, Inc., Ashland, OR, USA).
Western blot analysis
Proteins from cells were extracted with lysis buffer
(Beyotime, Beijing, China) with a 1% volume of phosphatase
inhibitor (Roche Diagnostics, Basel, Switzerland) mixture. Total
protein was measured using a standard bicinchoninic acid assay
(Pierce Biotechnology, Inc., Rockford, IL, USA). Protein samples
were fractioned in a 10% SDS-polyacrylamide gel and transferred to
a polyvinylidene fluoride membrane. After blocking by 5% skimmed
milk for 1 h, western blot analysis was performed with each of the
indicated primary antibodies at 4°C overnight and secondary
antibodies for 1 h. Immunodetection was performed using enhanced
electrochemiluminescence.
Statistical analyses
All the data are presented as the means ± standard
deviation. Difference among groups was compared using one-way
analysis of variance followed by the least significant difference
test. Values of P<0.05 were considered to indicate a
statistically significant difference. Experiments of all the groups
were performed in triplicate.
Results
Characterization of BM-derived EPCs and
detection of transfected EPCs
The isolated EPCs exhibited a spindle appearance
with distinct colony formations by 72 h (Fig. 1A). The 7-day cultured EPCs were
identified by Dil-ac-LDL and FITC-lectin double staining (Fig. 1B). To further confirm the
endothelial profile of EPCs, the expression of endothelial markers
was examined by immunofluorescent staining. The results showed that
the majority of cultured EPCs expressed endothelial markers,
including vWF (Fig. 1C) and
VEGFR-2 (Fig. 1D), indicating
their potential in differentiating into endothelial lineage. In
order to enhance the therapeutic potential of EPCs, LV-IL-10-GFP
and LV-NC-GFP were transfected into EPCs. GFP-labeled EPCs were
detected by a fluorescence microscope, indicating high transfection
efficiency (Fig. 1E and F). All
the above results confirmed the successful isolation and
transfection of BM-derived EPCs.
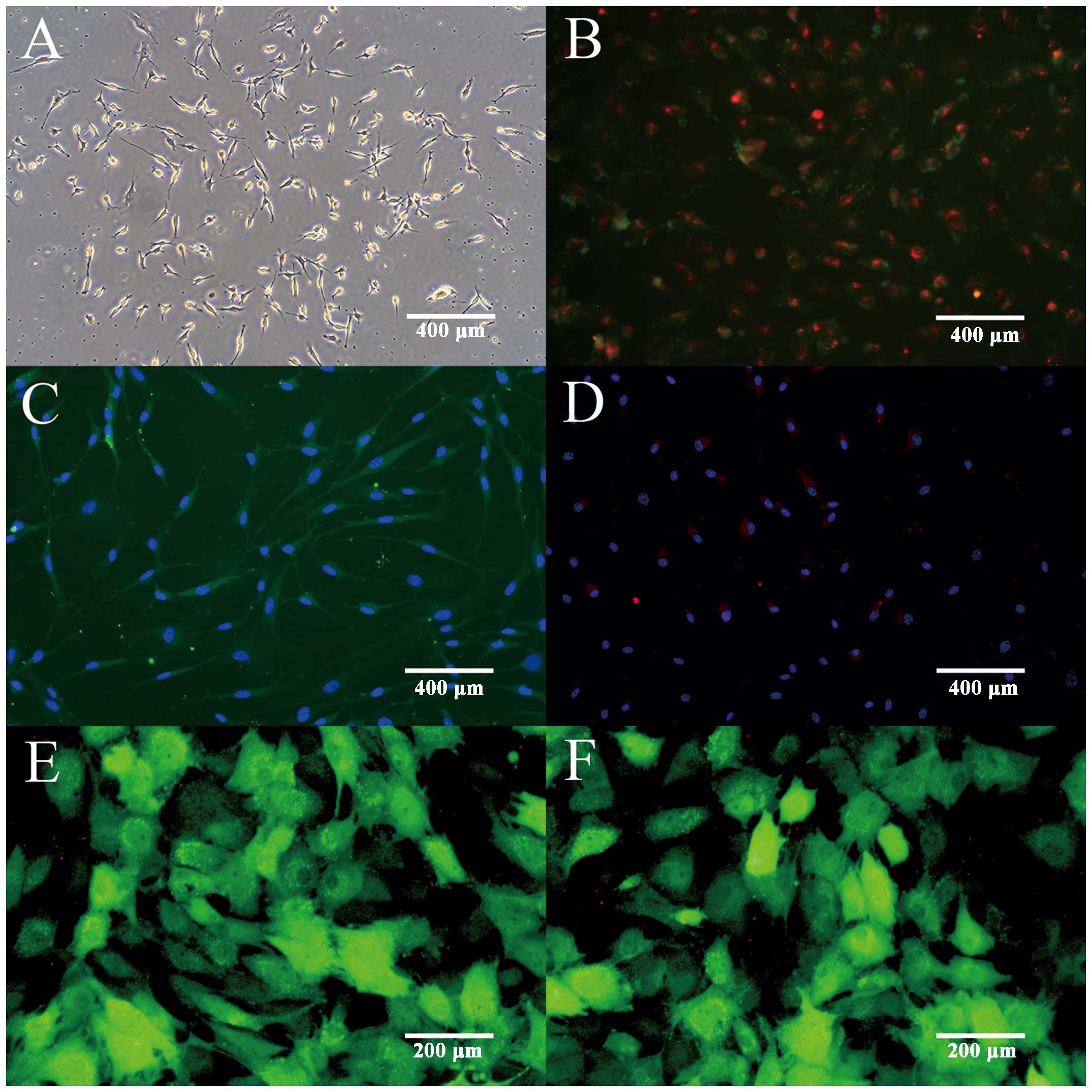 | Figure 1Characterization of EPCs and
transfected EPCs. (A) Isolation of EPCs exhibited a spindle
appearance after culturing for 72 h (magnification, ×100).
Fluorescence microscopy illustrated that adherent cells were
positive for uptake of Dil-acLDL and FITC-UEA-1. (B) The
double-positive cells were recognized as EPCs (magnification,
×100). (C) Immunoflurescent staining demonstrated that adherent
cells had positive expression of endothelial marker FITC-vWF
(magnification, ×100) and (D) PE-VEGFR2 (magnification, ×100).
Fluorescence microscopy showed EPCs expression of GFP following
transfection with (E) LV-IL-10-GFP (magnification, ×200) and (F)
LV-NC-GFP (magnification, ×200). EPC, endothelial progenitor cells;
Dil-acLDL, 1,1′-dioctadecyl-3,3,3′,3′-tetramethylindocarbocyanine
percholate-labeled acetylated low-density lipoprotein; FITC-UEA-1,
fluorescein isothiocyanate-Ulex europaeus agglutinin; vWF, von
Willebrand factor; PE-VEGFR2, phycoerythrin-vascular endothelial
growth factor receptor 2; GFP, green fluorescent protein; LV,
lentivirus vector; IL-10, interleukin-10; NC, noncontain. |
Concentrations of cytokines in EGM-2
The levels of TNF-α and IL-8 were significantly
decreased in the EPC-LV-IL-10-GFP group compared to the EPC and
EPC-LV-NC-GFP groups (P<0.05 for all), and there was no
statistically significant difference between the EPC and
EPC-LV-NC-GFP group (P>0.05). By contrast, the levels of IL-10
and VEGF were contrasting in association to these. The levels
significantly increased in the EPC-LV-IL-10-GFP group compared to
the EPC and EPC-LV-NC-GFP groups (P<0.05 for all), and there was
no statistically significant difference between the EPC and
EPC-LV-NC-GFP groups (P>0.05) ELISA analysis of EGM-2 with TNF-α
were consistent with EGM-2 without TNF-α. The bar graphs of the
ELISA analysis are shown in Fig.
2. The results showed that IL-10-transfected EPCs were more
resistant to an inflammatory stimulus, such as TNF-α, leading to
the decline of pro-inflammatory factors. Therefore, the function of
IL-10-modified EPCs may be improved in the inflammatory
environment.
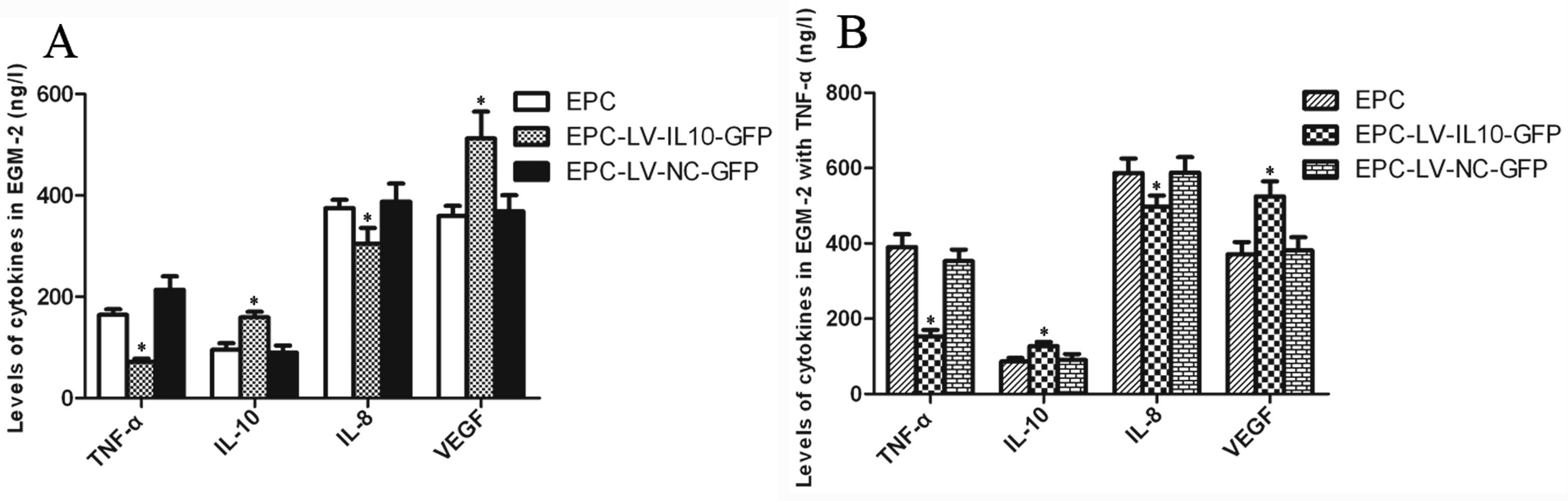 | Figure 2Bar graphs of the ELISA analysis of
EPCs. (A) The levels of TNF-α and IL-8 significantly decreased in
the EPC-LV-IL-10-GFP group compared to the EPC and EPC-LV-NC-GFP
groups, and there was no significant difference between the EPC and
EPC-LV-NC-GFP group. By contrast, the levels of IL-10 and VEGF were
significantly increased in the EPC-LV-IL-10-GFP group compared to
the EPC and EPC-LV-NC-GFP groups, and there was no statistically
significant difference between the EPC and EPC-LV-NC-GFP group. (B)
ELISA analysis of EGM-2 with TNF-α was consistent with EGM-2
without TNF-α (*P<0.05). EPCs, endothelial progenitor
cells; TNF-α, tumor necrosis factor-α; IL-8, interleukin-8; LV,
lentivirus vector; GFP, green fluorescent protein; NC, noncontain;
VEGF, vascular endothelial growth factor; EGM-2, endothelial cell
growth medium-2. |
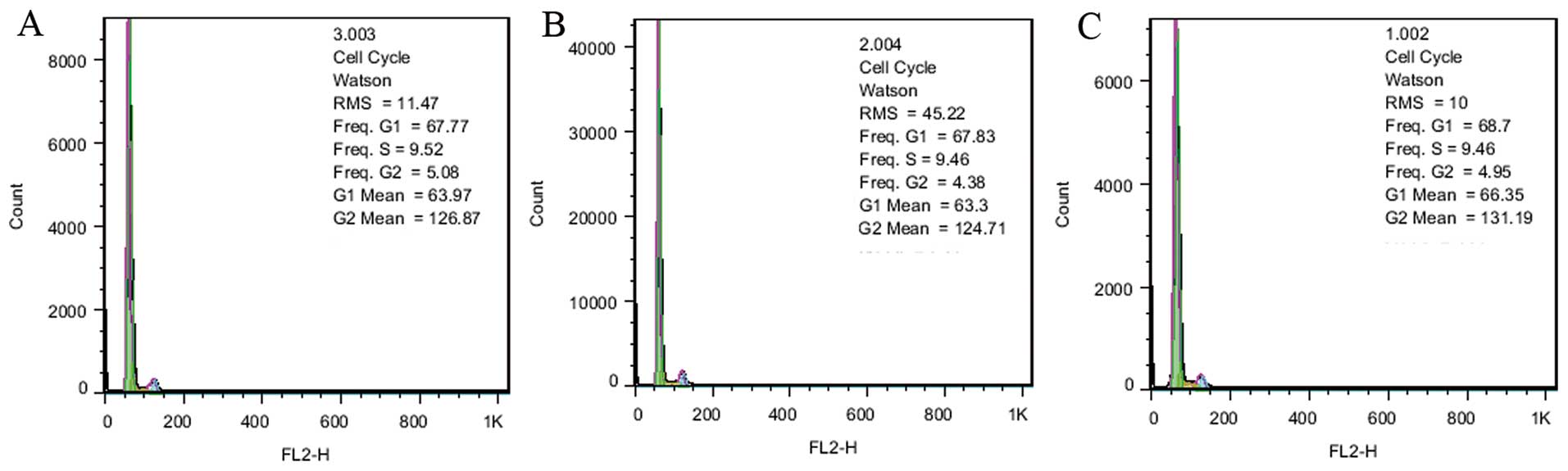
Overexpression of IL-10 modulates EPC
function
To explore the role of IL-10 in EPC function, EPCs
were transfected with LV-IL-10-GFP and LV-NC-GFP was the control
group. Analysis of the cell cycle by flow cytometry with PI/RNase
staining indicated that cells in the S phase were 9.52, 9.46 and
9.46% in the EPC, EPC-LV-IL-10-GFP and EPC-LV-NC-GFP groups,
respectively. The result revealed that there was no significant
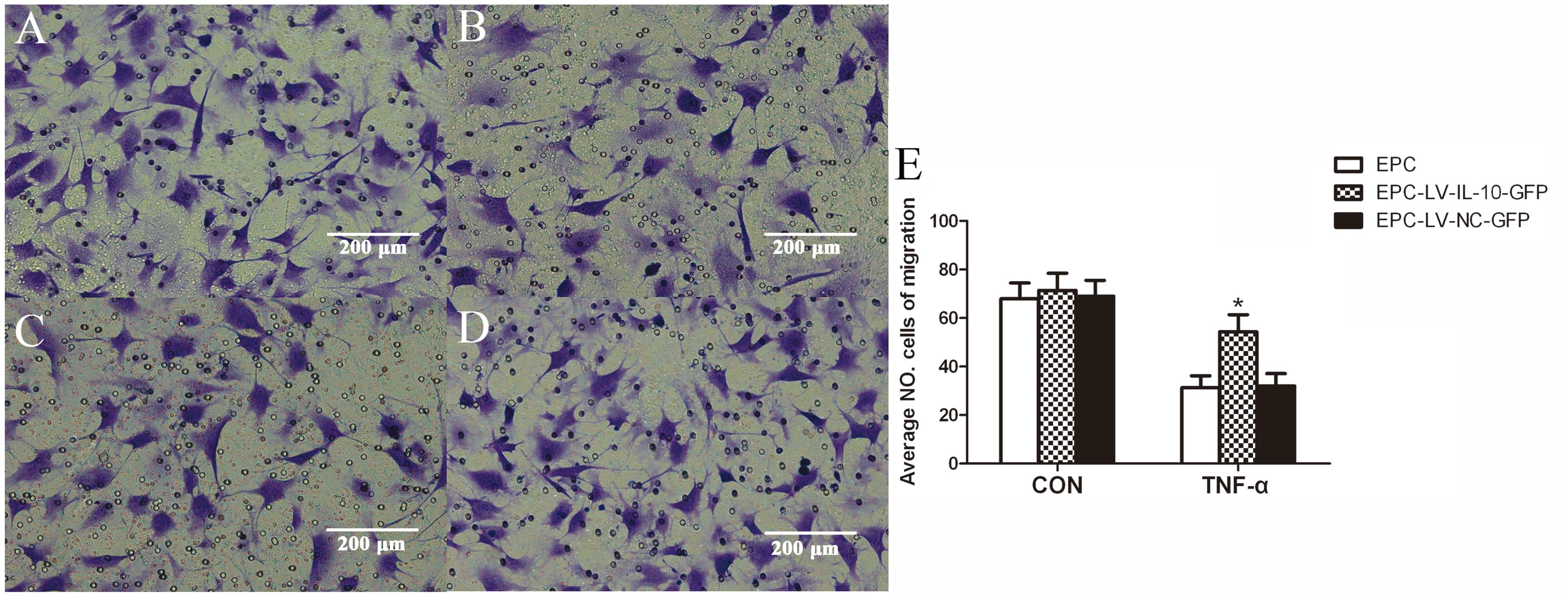
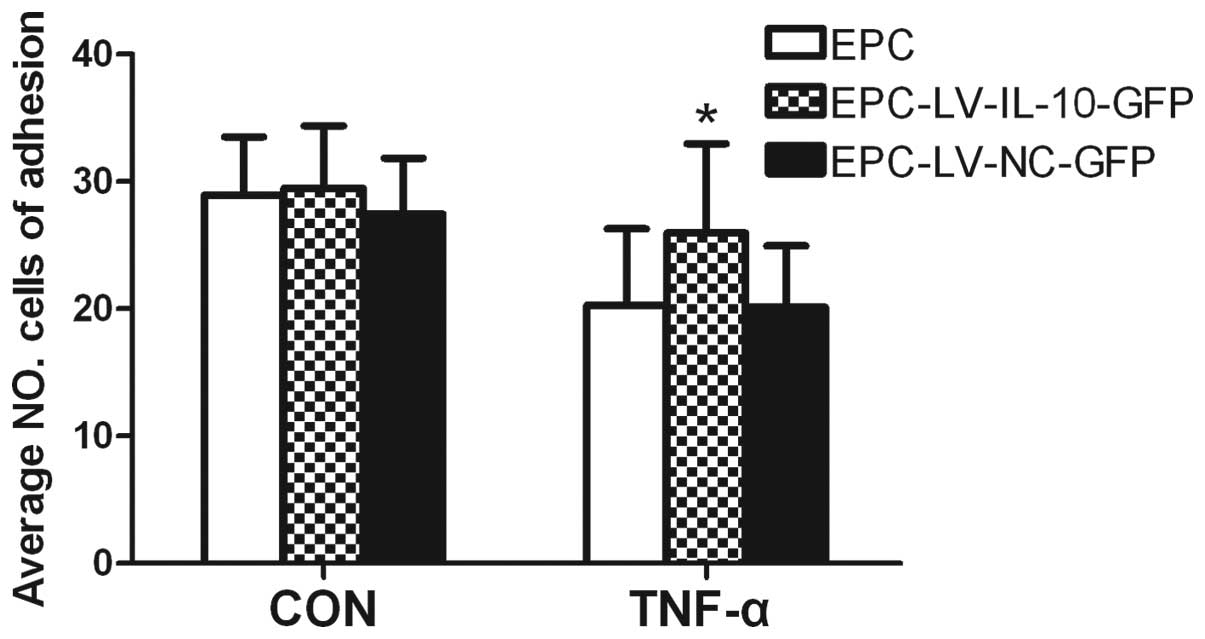
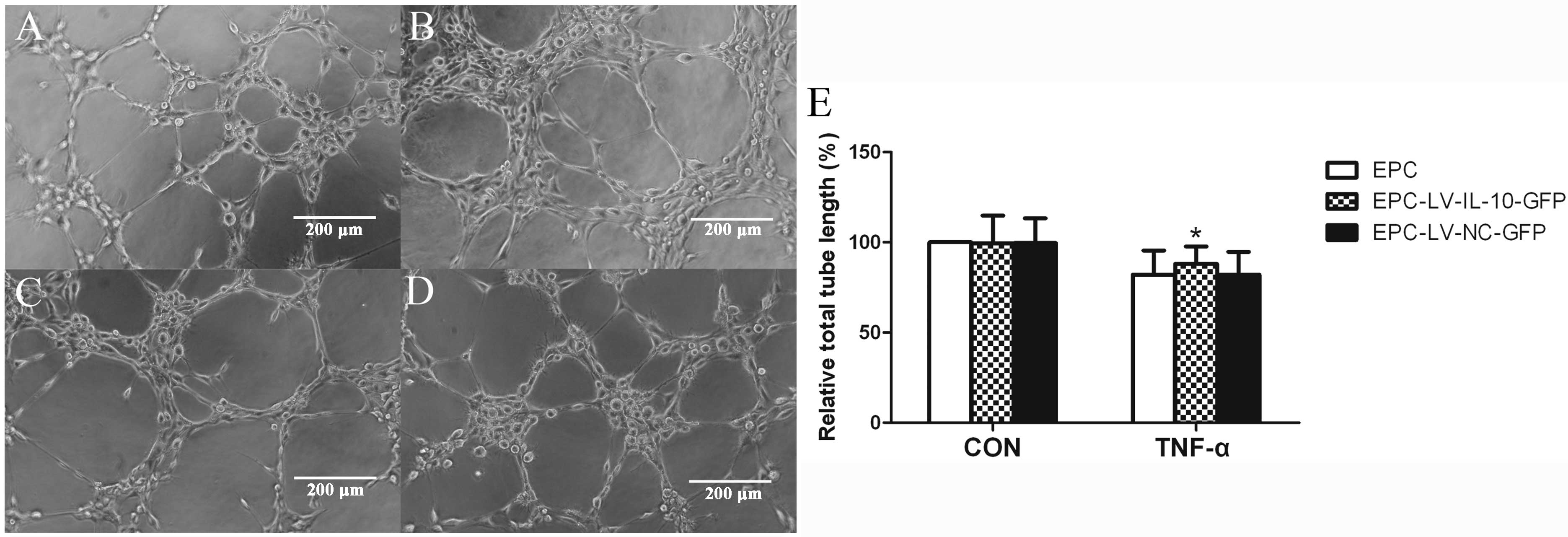
difference in cell growth between the three groups (Fig. 3). The data showed that the average
number of EPCs undergoing migration, adhesion and total lengths of
EPCs were similar among the EPC, EPC-LV-IL-10-GFP and EPC-LV-NC-GFP
groups without TNF-α (P>0.05 for all) (Figs. 4E, 5 and 6E). Further study showed that
EPC-LV-IL-10-GFP with TNF-α significantly enhanced EPC migration
(Fig. 4), adhesion (Fig. 5) and promoted EPC tube formation
(Fig. 6) compared to
EPC-LV-NC-GFP with TNF-α and EPC with TNF-α (P<0.05 for all).
The bar graph regarding the groups with or without TNF-α are shown
in Figs. 4E, 5E and 6E. In summary, these data confirmed that
the overexpression of IL-10 had no effect on migration, adhesion,
tubule formation and cell growth of EPCs without TNF-α.
Furthermore, the overexpression of IL-10 improved the function,
including the migration, adhesion and tubule formation of EPCs
stimulated with TNF-α.
Expression of proteins in EPCs
To study the mechanism of IL-10 in EPCs, the
expression of MMP-9, VEGF, STAT-3 and p-STAT3 was examined. Western
blot analysis revealed that the expression of VEGF, MMP-9 and
p-STAT3 significantly increased in the EPC-LV-IL-10-GFP group
compared to the other groups. By contrast, STAT-3 expression
decreased in the EPC-LV-IL-10-GFP group compared to the other
groups (Fig. 7). The results
suggested that improvement in the function of IL-10-modified EPCs
was possibly through the activation of the STAT3 signaling
pathway.
 | Figure 7Western blot analysis for the EPCs.
The results revealed that the expression of VEGF, MMP-9 and p-STAT3
significantly increased in the EPC-LV-IL-10-GFP group compared to
the EPC-LV-NC-GFP and EPC groups. By contrast, STAT-3 expression
decreased in the EPC-LV-IL-10-GFP group compared to the
EPC-LV-NC-GFP and EPC groups. EPCs, endothelial progenitor cells;
VEGF, vascular endothelial growth factor; MMP-9, matrix
metalloproteinase 9; p-STAT3, phosphorylated-signal transducer and
activator of transcription 3; LV, lentivirus vector; IL-10,
interleukin-10; GFP, green fluorescent protein; NC, noncontain. |
Discussion
A progenitor cell is a biological cell that, similar
to stem cells (SCs), has a tendency to differentiate into a
specific type of cell. However, it is already more specific than an
SC and is pushed to differentiate into its ‘target’ cell. EPCs are
believed to be derived from the BM and are able to differentiate
into mature endothelial cells (ECs). EPCs play a crucial role in
the neovascularization of ischemic tissue and in the maintenance of
endothelial cell integrity. Since Asahara et al (15) first identified circulating EPCs in
1997, increasing evidence suggests that BM-derived EPCs
functionally contribute to neovascularization in several models of
tissue injury and remodeling, including wound healing, myocardial
ischemia, retinopathy, stroke and peripheral vascular disease. Thus
far, techniques aimed at enhancing ex vivo expansion and the
therapeutic potential of EPCs, such as epigenetic and genetic
modifications of EPCs, are also being extensively studied (16). However, the effect and mechanism
remain unclear.
In the past decades, the potential pathogenesis of
inflammation in cardiovascular disease (17), lung cancer (18), pulmonary hypertension (19), vascular injury (20) and diabetes (21) has been extensively investigated.
Thus, anti-inflammation mediators play a critical role in treatment
for diseases. IL-10 is a 35-kDa homodimeric cytokine that is
produced by a variety of cell types. IL-10, a potent
anti-inflammatory cytokine, attenuates inflammatory response and
suppresses various pro-inflammatory mediators (6). Thus, the aim of the present study
was to observe the influence of IL-10 overexpression on EPCs and to
clarify the possible mechanism. IL-10-modified EPCs may be a
potentially therapeutic method for numerous vascular diseases.
In the present study, the data indicate that IL-10
does not influence migration, adhesion, tubule formation and cell
growth of EPCs. TNF-α is one of the critical pro-inflammatory
cytokines. Peplow (22) reported
that TNF-α has an unfavorable influence on cell migration and
apoptosis. In order to further study the influence of IL-10 on
EPCs, EPC functions were tested under the inflammatory
microenvironment induced by TNF-α. The results showed that IL-10
enhanced the function of EPCs stimulated with TNF-α, including
migration, adhesion and tubule formation in vitro. In
addition, a previous study demonstrated that EPC was able to carry
and express hNIS in glioma following intravenous administration
(23). Based on this, it is
possible to use EPCs as carrier delivery vehicles or therapeutic
genes, which can be administered either systemically or locally. In
addition, IL-10-modified EPCs possibly have a clear positive
clinical effect, due to the improvement of the local inflammatory
microenvironment and promotion of EPC function.
The IL-10 signaling pathway has been primarily
elucidated in monocytes. However, the mechanism of EPCs affected by
IL-10 remains unclear. There are numerous studies that exhibit the
association between IL-10 and STAT3 (24,25). STAT3 is known to be involved in
the development and progression of a number of different types of
tumor. The STAT signaling pathway is activated by a diverse array
of cytokines and growth factors, and has been indicated in a
variety of cellular functions, including inflammatory processes
(26). IL-10 dimerizes to bind to
a tetramer receptor complex that comprises two molecules of IL-10
receptor 1 (R1) and two molecules of IL-10R2, which permits
phosphorylation and dimerization of STAT3. Phosphorylated-STAT3
translocates to the nucleus to activate the downstream target
genes. Therefore, we hypothesize that IL-10 promotes the function
of EPCs through activating the STAT3 signaling pathway.
Western blot analysis suggests that the expression
of VEGF, MMP-9 and p-STAT3 significantly increased in the
EPC-LV-IL-10-GFP group compared to the EPC and EPC-LV-NC-GFP
groups, whereas the expression of STAT3 is contrasting in
association to these. VEGF is the noteworthy candidate for
stimulating new vessel formation and vascular hyperpermeability and
appears to have important roles for mobilization of EPCs. VEGF
stimulates proliferation, migration and survival of endothelial
cells, which in turn facilitated the endothelialization and
recovery of endothelial function of EPCs (27).
MMPs are well known as important modulators of
innate inflammation and are associated with angiogenesis and
vascular remodeling. Active MMPs degrade vascular basement membrane
and other extracellular matrix (ECM) proteins leading to detaching
of ECs, which is necessary for proliferation (28), migration (29) and even apoptosis of ECs (30) and is an initial step of
angiogenesis or vascular formation. In addition, MMPs can modify
non-ECM molecules, such as VEGFR, to affect EC vitality and
behavior (31). In addition,
deficiency of MMP-9 attenuates ischemia-induced neovascularization
through an impairment of bone marrow-derived EPC adhesion,
migration and proangiogenic functions (32). Therefore, the present result
suggests that IL-10 promotes the EPC migration, adhesion and tube
formation by increasing MMP-9 and VEGF expression, and activating
the STAT3 signaling pathway.
In conclusion, the present study demonstrated that
the overexpression of IL-10 had no effect on migration, adhesion,
tubule formation and cell growth of EPCs without stimulation with
TNF-α. Furthermore, in the EPCs stimulated with TNF-α, the
overexpression of IL-10 improved EPC function, including migration,
adhesion and tubule formation through activating the STAT3
signaling pathway. The delivery of IL-10-modified EPCs to the sites
of the different lesions may be a novel therapeutic target.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81371038 and
91442124).
References
|
1
|
Huang H, Huang F and Huang JP:
Transplantation of bone marrow-derived endothelial progenitor cells
overexpressing Delta-like-4 enhances functional neovascularization
in ischemic myocardium. Mol Med Rep. 8:1556–1562. 2013.PubMed/NCBI
|
|
2
|
Barsotti MC, Santoni T, Picoi ME, et al:
Endothelial progenitor cell homing in human myocardium in patients
with coronary artery disease. Int J Cardiol. 15:516–517. 2014.
View Article : Google Scholar
|
|
3
|
Long J and Wang S, Zhang Y, Liu X, Zhang H
and Wang S: The therapeutic effect of vascular endothelial growth
factor gene - or heme oxygenase-1 gene-modified
endothelialprogenitor cells on neovascularization of rat hindlimb
ischemia model. J Vasc Surg. 58:756–765. 2013. View Article : Google Scholar
|
|
4
|
Roncalli JG, Tongers J, Renault MA and
Losordo DW: Endothelial progenitor cells in regenerative medicine
and cancer: a decade of research. Trends Biotechnol. 26:276–283.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Grisar J, Aletaha D, Steiner CW, et al:
Depletion of endothelial progenitor cells in the peripheral blood
of patients with rheumatoid arthritis. Circulation. 111:204–211.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Krishnamurthy P, Lambers E, Verma S,
Thorne T, Qin G, Losordo DW and Kishore R: Myocardial knockdown of
mRNA-stabilizing protein HuR attenuates post-mi inflammatory
response and left ventricular dysfunction in IL-10-null mice. FASEB
J. 24:2484–2494. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Krishnamurthy P, Rajasingh J, Lambers E,
Qin G, Losordo DW and Kishore R: Il-10 inhibits inflammation and
attenuates left ventricular remodeling after myocardial infarction
via activation of STAT3 and suppression of HuR. Circ Res.
104:e9–e18. 2009. View Article : Google Scholar :
|
|
8
|
Yao L, Huang K, Huang D, Wang J, Guo H and
Liao Y: Acute myocardial infarction induced increases in plasma
tumor necrosis factor-alpha and interleukin-10 are associated with
the activation of poly (ADP-ribose) polymerase of circulating
mono-nuclear cell. Int J Cardiol. 123:366–368. 2008. View Article : Google Scholar
|
|
9
|
Jui HY, Lin CH, Hsu WT, et al: Autologous
mesenchymal stem cells prevent transplant arteriosclerosis by
enhancing local expression of interleukin-10, interferon-gamma, and
indoleamine 2,3-dioxygenase. Cell Transplant. 21:971–984. 2012.
View Article : Google Scholar
|
|
10
|
Kawamoto A, Iwasaki H, Kusano K, et al:
CD34-positive cells exhibit increased potency and safety for
therapeutic neovascularization after myocardial infarction compared
with total mononuclear cells. Circulation. 114:2163–2169. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhao W, Li JJ, Cao DY, et al: Intravenous
injection of mesenchymal stem cells is effective in treating liver
fibrosis. World J Gastroenterol. 18:1048–1058. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Krishnamurthy P, Thal M, Verma S, et al:
Interleukin-10 deficiency impairs bone marrow-derived endothelial
progenitor cell survival and function in ischemic myocardium. Circ
Res. 109:1280–1289. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Chen DD, Dong YG, Yuan H and Chen AF:
Endothelin 1 activation of endothelin A receptor/NADPH oxidase
pathway and diminished antioxidants critically contribute to
endothelial progenitor cell reduction and dysfunction in
salt-sensitive hypertension. Hypertension. 59:1037–1043. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Marrotte EJ, Chen DD, Hakim JS and Chen
AF: Manganese superoxide dismutase expression in endothelial
progenitor cells accelerates wound healing in diabetic mice. J Clin
Invest. 120:4207–4219. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
15
|
Asahara T, Murohara T, Sullivan A, et al:
Isolation of putative progenitor endothelial cells for
angiogenesis. Science. 275:964–967. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Peng LH, Tsang SY, Tabata Y and Gao JQ:
Genetically-manipulated adult stem cells as therapeutic agents and
gene delivery vehicle for wound repair and regeneration. J Control
Release. 157:321–330. 2012. View Article : Google Scholar
|
|
17
|
Golia E, Limongelli G, Natale F, et al:
Inflammation and cardiovascular disease: from pathogenesis to
therapeutic target. Curr Atheroscler Rep. 16:4352014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pu X, Wang L, Chang JY, et al:
Inflammation-related genetic variants predict toxicity following
definitive radiotherapy for lung cancer. Clin Pharmacol Ther. Jul
23–2014.(Epub ahead of print). View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Guignabert C and Dorfmuller P: Pathology
and pathobiology of pulmonary hypertension. Semin Respir Crit Care
Med. 34:551–559. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Inoue T, Croce K, Morooka T, Sakuma M,
Node K and Simon DI: Vascular inflammation and repair: implications
for re-endothelialization, restenosis, and stent thrombosis. JACC
Cardiovasc Interv. 4:1057–1066. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Tang J and Kern TS: Inflammation in
diabetic retinopathy. Prog Retin Eye Res. 30:343–358. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Peplow PV: Influence of growth factors and
cytokines on angiogenic function of endothelial progenitor cells: a
review of in vitro human studies. Growth Factors. 32:83–116. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Palenski TL, Sorenson CM and Sheibani N:
Inflammatory cytokine-specific alterations in retinal endothelial
cell function. Microvasc Res. 89:57–69. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Lafarge S, Hamzeh-Cognasse H, Chavarin P,
Genin C, Garraud O and Cognasse F: A flow cytometry technique to
study intracellular signals NF-kappaB and STAT3 in peripheral blood
mononuclear cells. BMC Mol Biol. 8:642007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Maher K, Završnik J, Jerič-Kokelj B,
Vasiljeva O, Turk B and Kopitar-Jerala N: Decreased IL-10
expression in stefin B-deficient macrophages is regulated by the
MAP kinase and STAT-3 signaling pathways. FEBS Lett. 588:720–726.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Leonard WJ and O’Shea JJ: Jaks and STATS:
biological implications. Annu Rev Immunol. 16:293–322. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Jin K, Kashiwagi K, Iizuka Y, Tanaka Y,
Imai M and Tsukahara S: Matrix metalloproteinases in human diabetic
and nondiabetic vitreous. Retina. 21:28–33. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Mazor R, Alsaigh T, Shaked H, et al:
Matrix metalloproteinase-1-mediated up-regulation of vascular
endothelial growth factor-2 in endothelial cells. J Biol Chem.
288:598–607. 2013. View Article : Google Scholar :
|
|
29
|
Genís L, Gonzalo P, Tutor AS, et al:
Functional interplay between endothelial nitric oxide synthase and
membrane type 1-matrix metalloproteinase in migrating endothelial
cells. Blood. 110:2916–2923. 2007. View Article : Google Scholar
|
|
30
|
Shapiro S, Khodalev O, Bitterman H,
Auslender R and Lahat N: Different activation forms of MMP-2
oppositely affect the fate of endothelial cells. Am J Physiol Cell
Physiol. 298:C942–C951. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Ito TK, Ishii G, Saito S, Yano K, Hoshino
A, Suzuki T and Ochiai A: Degradation of soluble vegf receptor-1 by
MMP-7 allows VEGF access to endothelial cells. Blood.
113:2363–2369. 2009. View Article : Google Scholar
|
|
32
|
Huang PH, Chen YH, Wang CH, et al: Matrix
metalloproteinase-9 is essential for ischemia-induced
neovascularization by modulating bone marrow-derived endothelial
progenitor cells. Arterioscler Thromb Vasc Biol. 29:1179–1184.
2009. View Article : Google Scholar : PubMed/NCBI
|