Introduction
Natural killer (NK) cells are large granular
lymphocytes that function as effectors of innate immune activity
and modulators of adaptive immune responses (1,2).
NK cells kill stressed and abnormal cells, such as virus-infected
cells and tumor cells, and can also produce cytokines. NK
cell-mediated cytotoxicity and cytokine production are regulated by
NK cell surface receptors, such as activating, inhibitory,
adhesion, cytokine and chemotactic receptors, and the relative
balance of these receptor-mediated signals fine-tunes the various
biological activities of NK cells. NK cells circulate in the
peripheral blood NK (termed pNK cells) (3). NK cells are also present in the
uterus (4). In the human
placenta, there are two distinct fetal maternal interfaces, the
first is the decidua and the second is the intervillous space where
maternal blood bathes the floating chorionic villous tree. A
specific population of NK cells accumulates within the decidua.
These NK cells, referred to as decidual NK (dNK) cells, are
considered to play a pivotal role in placentation, including
trophoblast invasion and uterine spiral artery remodeling (5,6).
As pNK cells can come into direct contact with chorionic villi at
the intervillous space, mechanisms of resistance to pNK
cytotoxicity may be involved at the second interface. This second
interface constitutes a privileged site where the secretion of
placental microvesicles, including exosomes, into the maternal
peripheral blood occurs. Hedlund et al reported that the
human placenta secretes KLRK1 ligands via exosomes that induce the
downregulation of the KLRK1 receptor on pNK cells, leading to a
reduction in their cytotoxicity in vitro (7). The syncytiotrophoblast covering
chorionic villi may evade NK cytotoxicity from these cells.
MicroRNAs (miRNAs or miRs) are small non-coding RNAs
that play a pivotal role in post-transcriptional gene regulation by
targeting the 3′-untranslated region (3′-UTR) of specific target
mRNAs for endonucleolytic cleavage or translational repression
(8). With regard to human NK cell
miRNAs, genome-wide comparisons have been made for human
lymphocytes subsets, including NK cells (9,10).
Two studies have also reported the miRNA profiles of resting and
cytokine-activated pNK cells using next-generation sequencing
(11,12). Despite such progress, knowledge of
the NK cell miRNA profiles and their physiological roles remain
incomplete. Moreover, little is known about the miRNA-gene
regulatory relationships that may be relevant for the functions of
maternal NK cells during pregnancy.
In the present study, to determine the roles of
miRNAs within gene regulatory networks of maternal pNK cells during
pregnancy, we performed comprehensive miRNA and gene expression
profiling of NK cells isolated from the peripheral blood of healthy
pregnant females and analyzed these differential expression levels
between first- and third-trimester pNK cells. We explored NK cell
function-associated genes that were negatively correlated with
miRNA expression levels and computationally predicted to be miRNA
targets. Finally, we constructed a regulatory network for
miRNA-mediated gene expression in pNK cells during pregnancy using
miRNA and gene expression profiles.
Materials and methods
pNK cell isolation from pregnant
females
Samples of peripheral blood were obtained from
pregnant females after obtaining informed consent. For the
comprehensive analysis of mRNA and gene expression profiles in pNK
cells, samples were obtained from the same healthy pregnant females
during the first (gestational age, 7–11 weeks), second (19–23
weeks) and third (36–38 weeks) trimesters of gestation (n=5 each),
and from other females who had a normal pregnancy 4 days following
delivery (n=5). For the validation of miRNA expression levels by
reverse transcription quantitative PCR (RT-qPCR, real-time PCR) in
pNK cells, a different set of experiments with other healthy
pregnant females was performed; samples were obtained from the same
females in the first, second and third trimesters of gestation (n=5
each), and from other females who had a normal pregnancy 4 days
following delivery (n=5). The study protocols were approved by the
Ethics Committees of Jichi Medical University (Tochigi, Japan) and
Nippon Medical School (Tokyo, Japan). Peripheral blood mononuclear
cells were isolated from heparinized venous blood using Lymphoprep
(Axis-Shield PoC AS, Oslo, Norway) as previously described
(13). NK cells were isolated
from the peripheral blood mononuclear cells using the Dynabeads
Untouched NK Cells kit (Invitrogen, Carlsbad, CA, USA) according to
the manufacturer’s instructions.
Total RNA within the cells was extracted using
RNAiso reagent (Takara Bio, Inc., Shiga, Japan) according to the
manufacturer’s instructions. The integrity of the RNA was
determined using an Agilent 2100 Bioanalyzer (Agilent Technologies,
Santa Clara, CA, USA); samples with an RNA integrity number >7
were used.
Quantitative PCR-based array analysis of
miRNAs
We performed real-time PCR-based array analysis to
quantitatively and comprehensively examine the expression levels of
756 miRNAs in the pNK cells obtained from pregnant females. Total
RNA from each specimen (each 30 ng) was reverse transcribed using
Megaplex RT Primers (Applied Biosystems, Foster City, CA, USA). The
cDNA was then pre-amplified using Megaplex PreAmp Primers (Applied
Biosystems). The pre-amplified products were subjected to real-time
PCR using TaqMan Array Human MicroRNA Cards (A and B, version 2.0)
on a 7900HT Fast Real-Time PCR System (Applied Biosystems)
according to the manufacturer’s instructions. The miRNA sequences
were annotated using the Sanger database (miRBase), release 14.
Data obtained with this assay were analyzed using RQ Manager 1.2
(Applied Biosystems). The relative Ct method (ΔΔCt method) was
applied for the quantification of each miRNA expression level, as
previously described (14). The
data were normalized relative to the expression of
RNU6-2.
Real-time PCR analysis for the validation
of miRNA expression levels
Real-time PCR for miRNA expression was carried out
using TaqMan MicroRNA Assays on a 7300 Real-Time PCR System (both
from Applied Biosystems). Briefly, total RNA (each 10 ng) was
reverse transcribed with a High Capacity cDNA Reverse Transcription
kit (Applied Biosystems). The RT products were subsequently
subjected to real-time PCR using TaqMan 2X Universal PCR Master Mix
(Applied Biosystems). To normalize the expression levels of the
miRNAs, SNORD44 was used as an endogenous internal control.
Primers for miR-517a-3p (assay ID 002402) and
miR-518b (assay ID 001156) were from Applied Biosystems.
DNA microarray analysis
We performed microarray analysis of the gene
expression levels in the first- and third-trimester pNK cells. Of
the total RNA obtained, 20 ng was used in a labeling reaction with
a Low-Input Quick Amp Labeling kit, one-color (Agilent
Technologies), and the quality and yield of the labeled cRNA were
evaluated on an Agilent 2100 Bioanalyzer. Gene expression profiling
was conducted using an Agilent Microarray (Human GE 4×44K, version
2). The resulting signals were normalized relative to the 75th
percentile signal intensity and spots with low reproducibility were
omitted (coefficient of variation <50%). The processed data were
filtered for P<0.05 (paired t-test) using the GeneSpring GX
software (version 11.5; Agilent Technologies). The mRNA array data
are publicly available (Gene Expression Omnibus, accession no.
GSE64884; http://www.ncbi.nlm.nih.gov/geo/).
Pathway and network analysis
Molecular networks of the selected molecules and
specific pathways were analyzed by the Core Analysis of fold change
for the third- vs. first-trimester pNK cell group in Ingenuity
Pathway Analysis (IPA; www.ingenuity.com). A total of 715 NK cell
function-related genes have been previously reported (1,15).
Of these 715 NK cell function-related genes, 69 were included in
the 1,173 differentially expressed genes (P<0.05) in the
above-mentioned microarray data and were used for IPA. In addition
to IPA, the miRNA.org database (August 2010 release; http://www.microrna.org/microrna/getMirnaForm.do)
was also used to predict the targets of miRNAs.
Statistical analysis
We conducted all analyses using the SPSS statistical
software package (Windows version 20; IBM-SPSS, Inc., Chicago, IL,
USA). The significance of between-group differences was assessed
using ANOVA followed by Tukey’s test, or the Kruskal-Wallis test,
and a value of P<0.05 was considered to indicate a statistically
significant difference.
Results
Differential expression of miRNAs in pNK
cells during and after pregnancy
We performed real-time PCR-based array analysis of
the expression levels of 756 miRNAs in pNK cells at different time
points of pregnancy (first, second and third trimester of
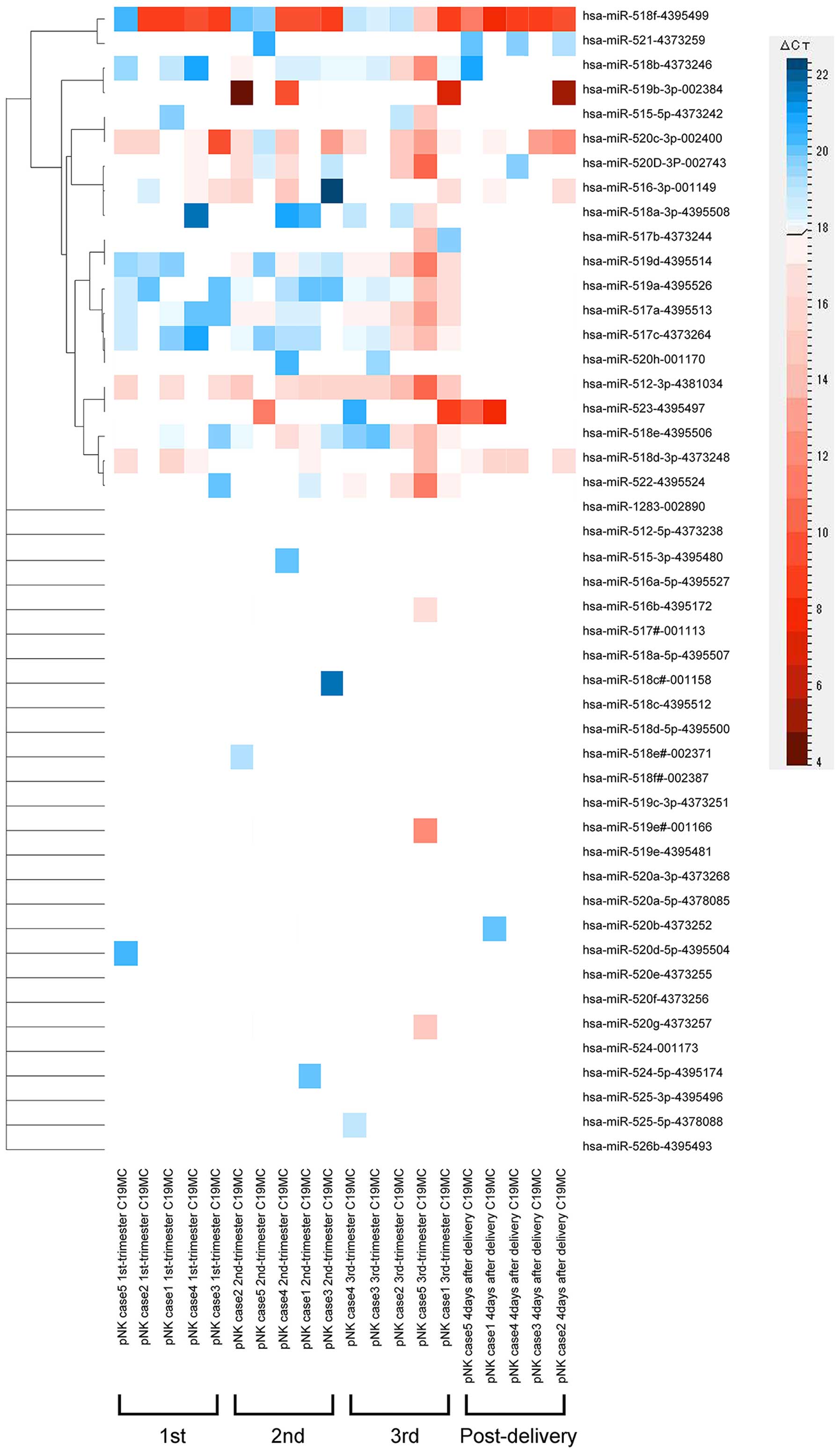
gestation, and post-delivery). Among the 756 miRNAs examined, 293,
293, 294 and 325 miRNAs were detected in the pNK cells obtained
from females in the first, second and third trimester of gestation,
and post-delivery, respectively. Of the 47 mature chromosome 19
miRNA cluster (C19MC) miRNAs, which are expressed exclusively in
the placenta (16–18), probed in the array cards, 12, 15,
and 18 placental C19MC miRNAs (26, 32 and 38% of the C19MC miRNAs,
respectively) were detected in the pNK cells obtained from the
females in the first, second and third trimester of gestation,
respectively (Fig. 1). By
contrast, the number of detectable C19MC miRNAs decreased following
delivery; only six C19MC miRNAs (13%) were detected in the pNK
cells obtained from females following delivery (Fig. 1).
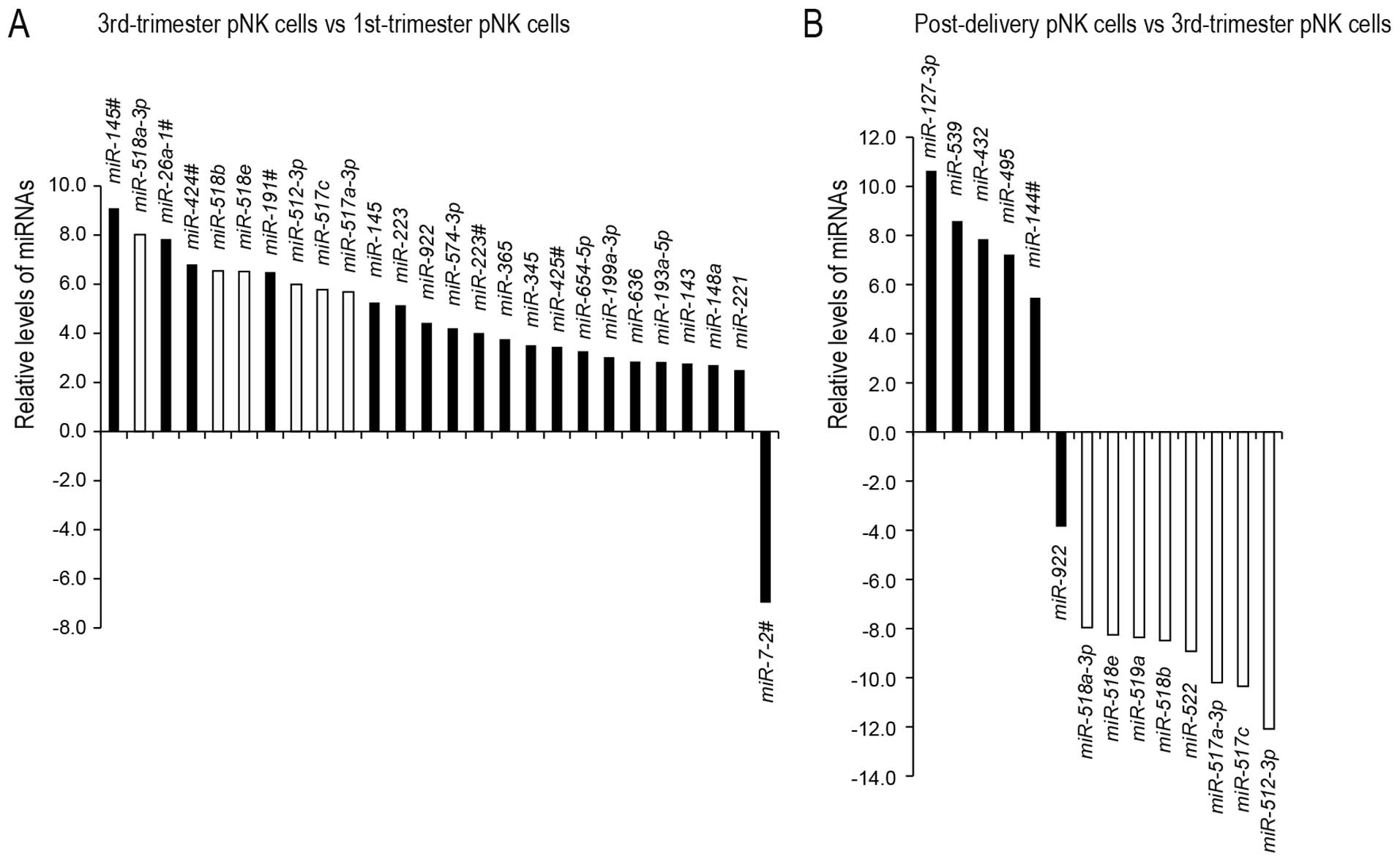
A comparison of the miRNA expression profiles
revealed that 25 miRNAs were significantly upregulated in the
third-compared to the first-trimester pNK cells (P<0.05;
Fig. 2A). The upregulated miRNAs
included six C19MC miRNAs (24% of the upregulated miRNAs). Only one
miRNA, miR-7-2# [currently defined as miR-7-2-3p in
miRBase release 21 (http://www.mirbase.org/)], was significantly
downregulated in the third-trimester pNK cells compared to those in
the first trimester. Subsequently, a comparison of the miRNA
expression profiles between the third-trimester and post-delivery
pNK cells identified nine miRNAs, including eight C19MC miRNAs (89%
of the downregulated miRNAs), which were significantly
downregulated in the post-delivery pNK cells compared to the
third-trimester pNK cells (Fig.
2B). Five miRNAs were significantly upregulated in the
post-delivery pNK cells compared to the third-trimester pNK
cells.
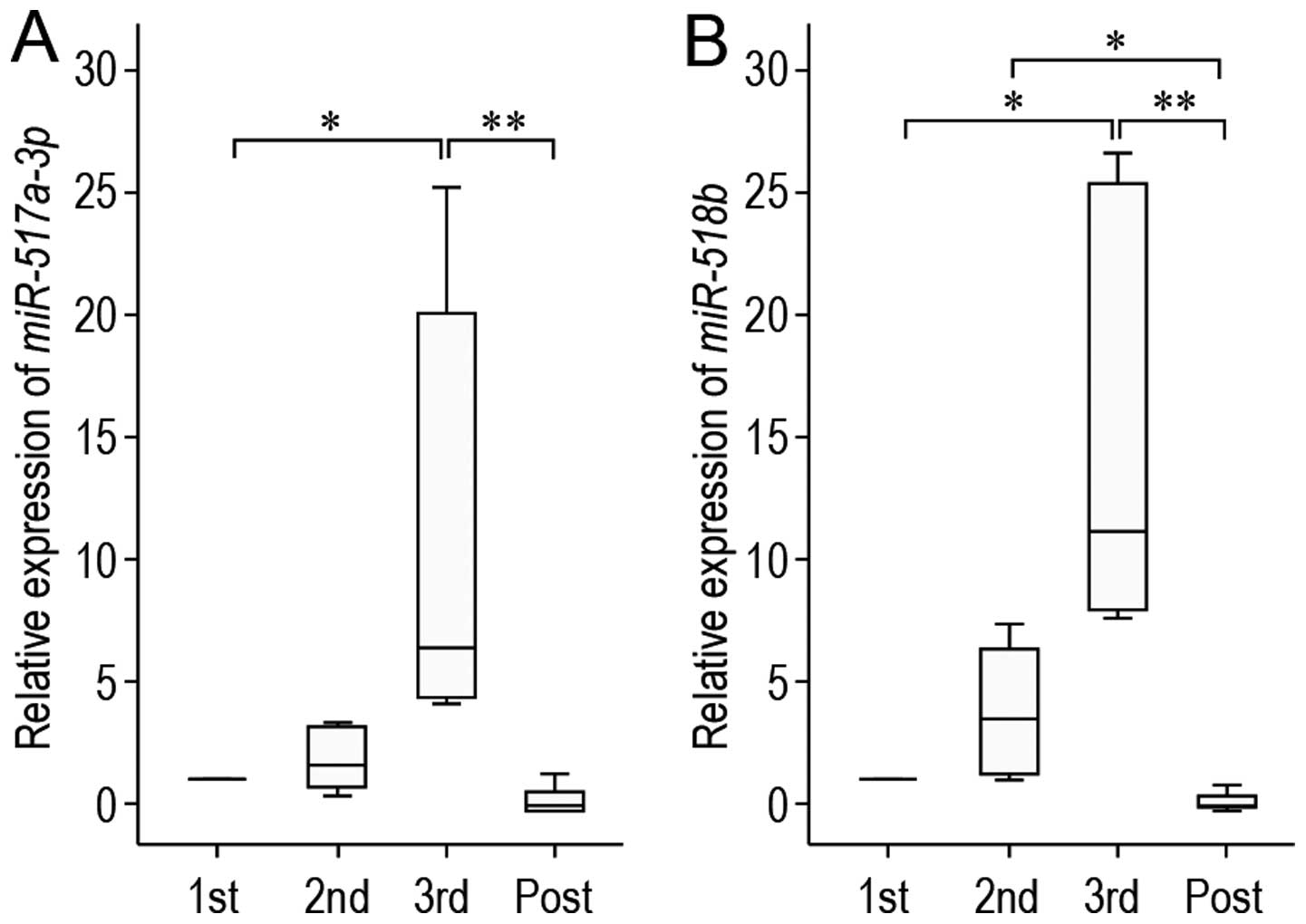
To confirm the differential C19MC miRNA expression
identified by real-time PCR-based array assay, we investigated the
expression of miR-517a-3p and miR-518b in the pNK
cells at different time points of pregnancy by RT-qPCR (Fig. 3) as these miRNAs were included in
the 25 upregulated miRNAs of the third-trimester pNK cells. A
different set of experiments with other pregnant females was
performed. In the pNK cells obtained during pregnancy,
miR-517a-3p and miR-518b were detected; these C19MC
miRNAs were significantly upregulated in a gestational
stage-dependent manner (Fig. 3).
On the other hand, the levels of miR-517a-3p and
miR-518b were significantly decreased at 4 days following
delivery (Fig. 3). These results
suggest that the transfer of placental C19MC miRNAs into maternal
pNK cells occurs during pregnancy.
Comparison of gene expression between
first- and third-trimester pNK cells
Using DNA microarray analysis, we assessed
differentially expressed genes in the pNK cells during pregnancy to
identify genes whose expression levels negatively correlated with
those of the miRNAs. At the transcriptional level, the
third-trimester pNK cells differed markedly from the first-trimes
ter pNK cells. DNA microarray analysis revealed the differential
expression of 1,173 genes among the 27,958 transcripts examined in
this study (P<0.05) (a data set of the differential expression
of 1,173 genes is available upon request). Among the 1,173 genes,
264 genes were significantly downregulated in the third- compared
to the first-trimester cells. On the other hand, the expression of
909 genes was found to be significantly increased in the third-
compared to the first-trimester cells.
Furthermore, we focused on the 715 genes which have
been previously reported to be related to NK cell functions (i.e.,
cytolytic pathway, cytokines/chemokines and their receptors,
secretory signature, cell surface/adhesion molecules, cell cycle
and proliferation and signal transduction) (1,15).
Among these 715 genes, 69 NK cell function-related genes were
differentially expressed between the first- and third-trimester pNK
cells (P<0.05); these included genes encoding NK cell surface
and adhesion molecules, as well as cell cycle-related genes
(Table I). Thirteen genes were
significantly downregulated in the third- compared to the
first-trimester cells. Of note, as regards cell surface/adhesion
molecules, killer-cell immunoglobulin-like receptor (KIR) genes
(e.g., KIR2DL4) showed significantly a decreased expression
in the third- compared to the first-trimester cells. By contrast,
the expression levels of leukocyte Ig-like receptor (LILR) genes
(e.g., LILRB2) in the third-trimester cells were
significantly higher than those in the first-trimester cells. With
regard to cell cycle and proliferation, the third-trimester pNK
cells showed a high-level expression of a group of genes involved
in regulating cell cycle transitions (e.g., CDK2 and
CCNA1) (Table I).
 | Table INK cell function-related genes that
were differentially expressed between the first- and
third-trimester pNK cells in DNA microarray analysis. |
Table I
NK cell function-related genes that
were differentially expressed between the first- and
third-trimester pNK cells in DNA microarray analysis.
| NK cell
function | Gene symbol | Log fold change
(3rd vs. 1st) | P-value | miRNAa |
|---|
| Cytolytic
pathway | CTSZ | 1.44 | 0.0025 | |
| Cytolytic
pathway |
TNFSF13B | 1.29 | 0.0009 | |
| Cytolytic
pathway | TNFAIP2 | 3.26 | 0.0009 | |
| Cytokines,
chemokines and their receptors | CXCR7
(CMKOR1) | −1.06 | 0.0009 | |
| Cytokines,
chemokines and their receptors | IL13RA1 | 3.22 | 0.0053 | |
| Cytokines,
chemokines and their receptors | IRF2BP2 | 0.76 | 0.0130 | |
| Cytokines,
chemokines and their receptors | IL17RA | 1.01 | 0.0067 | |
| Secretory
signature, cell cycle, proliferation | CDKN3 | 2.74 | 0.0009 | |
| Secretory
signature | HM13 | 0.83 | 0.0013 | |
| Secretory
signature | DEGS1 | 1.07 | 0.0012 | |
| Secretory
signature | GLRX | 0.72 | 0.0017 | |
| Cell surface and
adhesion molecules | ITM2C | 2.57 | 0.0009 | |
| Cell surface and
adhesion molecules | CD59 | 2.18 | 0.0009 | |
| Cell surface and
adhesion molecules | CD244 (2B4A,
2B4) | −0.70 | 0.0018 | miR-143 |
| Cell surface and
adhesion molecules | CD24 | 4.59 | 0.0010 | |
| Cell surface and
adhesion molecules | CD82 | 3.22 | 0.0009 | |
| Cell surface and
adhesion molecules | CD63 | 1.02 | 0.0016 | |
| Cell surface and
adhesion molecules | CD302 | 3.57 | 0.0009 | |
| Cell surface and
adhesion molecules | CD200R1 | 1.81 | 0.0082 | |
| Cell surface and
adhesion molecules |
KIR2DL5A | −0.86 | 0.0000 | miR-148a,
miR-221, miR-654-5p |
| Cell surface and
adhesion molecules | KIR3DL1 | −0.84 | 0.0009 | miR-221,
miR-654-5p |
| Cell surface and
adhesion molecules | KIR2DS4 | −0.80 | 0.0003 | miR-512-3p,
miR-654-5p |
| Cell surface and
adhesion molecules | KLRK1 (NKG2D,
CD314) | −0.83 | 0.0051 | miR-143,
miR-365, miR-636, miR-654-5p |
| Cell surface and
adhesion molecules | KIR2DL4 | −0.93 | 0.0009 |
miR-654-5p |
| Cell surface and
adhesion molecules | LILRA2 | 3.51 | 0.0009 | |
| Cell surface and
adhesion molecules | LILRA4 | 2.40 | 0.0050 | |
| Cell surface and
adhesion molecules | LILRA5 | 2.65 | 0.0009 | |
| Cell surface and
adhesion molecules | LILRA6 | 3.58 | 0.0014 | |
| Cell surface and
adhesion molecules | LILRB2 | 4.16 | 0.0009 | |
| Cell surface and
adhesion molecules | LILRB3 | 2.58 | 0.0015 | |
| Cell surface and
adhesion molecules | CEACAM1 (BGP,
CD66A) | 3.87 | 0.0015 | |
| Cell surface and
adhesion molecules | CEACAM6
(CD66C) | 4.73 | 0.0009 | |
| Cell cycle,
proliferation | TFDP1 | 1.19 | 0.0009 | |
| Cell cycle,
proliferation | MCM6 | 1.12 | 0.0012 | |
| Cell cycle,
proliferation | E2F1 | 1.78 | 0.0009 | |
| Cell cycle,
proliferation | GTSE1 | 2.31 | 0.0010 | |
| Cell cycle,
proliferation | CKS2 | 1.04 | 0.0009 | |
| Cell cycle,
proliferation | MKI67 | 3.45 | 0.0011 | |
| Cell cycle,
proliferation | PCNA | 1.13 | 0.0016 | |
| Cell cycle,
proliferation | CHEK1 | 1.86 | 0.0009 | |
| Cell cycle,
proliferation | CKS1B | 1.11 | 0.0008 | |
| Cell cycle,
proliferation | CDK2 | 0.73 | 0.0095 | |
| Cell cycle,
proliferation | KNTC1 | 0.96 | 0.0009 | |
| Cell cycle,
proliferation | MCM2 | 1.41 | 0.0009 | |
| Cell cycle,
proliferation | CCNB1 | 1.60 | 0.0009 | |
| Cell cycle,
proliferation | CCNE2 | 2.15 | 0.0009 | |
| Cell cycle,
proliferation | E2F2 | 3.35 | 0.0009 | |
| Cell cycle,
proliferation | CCNB2 | 3.61 | 0.0009 | |
| Cell cycle,
proliferation | KPNA2 | 0.65 | 0.0009 | |
| Cell cycle,
proliferation | RBBP8 | 1.91 | 0.0009 | |
| Cell cycle,
proliferation | CCNA2 | 2.83 | 0.0009 | |
| Cell cycle,
proliferation | GADD45A | 1.99 | 0.0011 | |
| Cell cycle,
quiescence | FBXO9 | 1.28 | 0.0011 | |
| Cell cycle,
quiescence | DHRS7 | −0.25 | 0.0044 | |
| JAK/STAT
pathway | JAK2 | 0.61 | 0.0009 | |
| JAK/STAT
pathway | SLA2 | −0.99 | 0.0022 | |
| JAK/STAT
pathway, | IL1R1 | 3.99 | 0.0024 | |
| NF-κB pathway
regulation | | | | |
| JAK/STAT
pathway | IL6R | 3.43 | 0.0017 | |
| TGF-β pathway | RUNX2 | 1.08 | 0.0052 | |
| TGF-β pathway | BMP2 | 2.49 | 0.0019 | |
| TGF-β pathway | BMPR1A | −1.22 | 0.0068 | miR-143,
miR-654-5p, miR-922 |
| P25K pathway | AKT2 | −0.55 | 0.0082 | miR-143,
miR-148a, miR-193a-5p, miR-221, miR-345, miR-365, miR-574-3p,
miR-654-5p, miR-922 |
| P43K pathway | FOXM1 | 2.83 | 0.0009 | |
| P50K pathway | PAK1 | 1.95 | 0.0009 | |
| NF-κB pathway
regulation | IRAK3 | 3.93 | 0.0010 | |
| NF-κB pathway
regulation | TLR2 | 2.41 | 0.0024 | |
| NF-κB target
genes | DUSP2 | −1.05 | 0.0058 | |
| NF-κB target
genes | LITAF | −0.54 | 0.0049 | miR-636,
miR-654-5p |
| NF-κB target
genes | MARCKS | 4.25 | 0.0009 | |
Bioinformatics analysis of functional
pathways and network of the differentially expressed genes
We then used the IPA tool to search for molecular
pathways associated with pNK cells during pregnancy. A data set of
69 NK cell function-associated genes mentioned above was included
in the IPA Core Analysis that allowed us to interpret the data set
in the context of biological processes, pathways and gene networks.
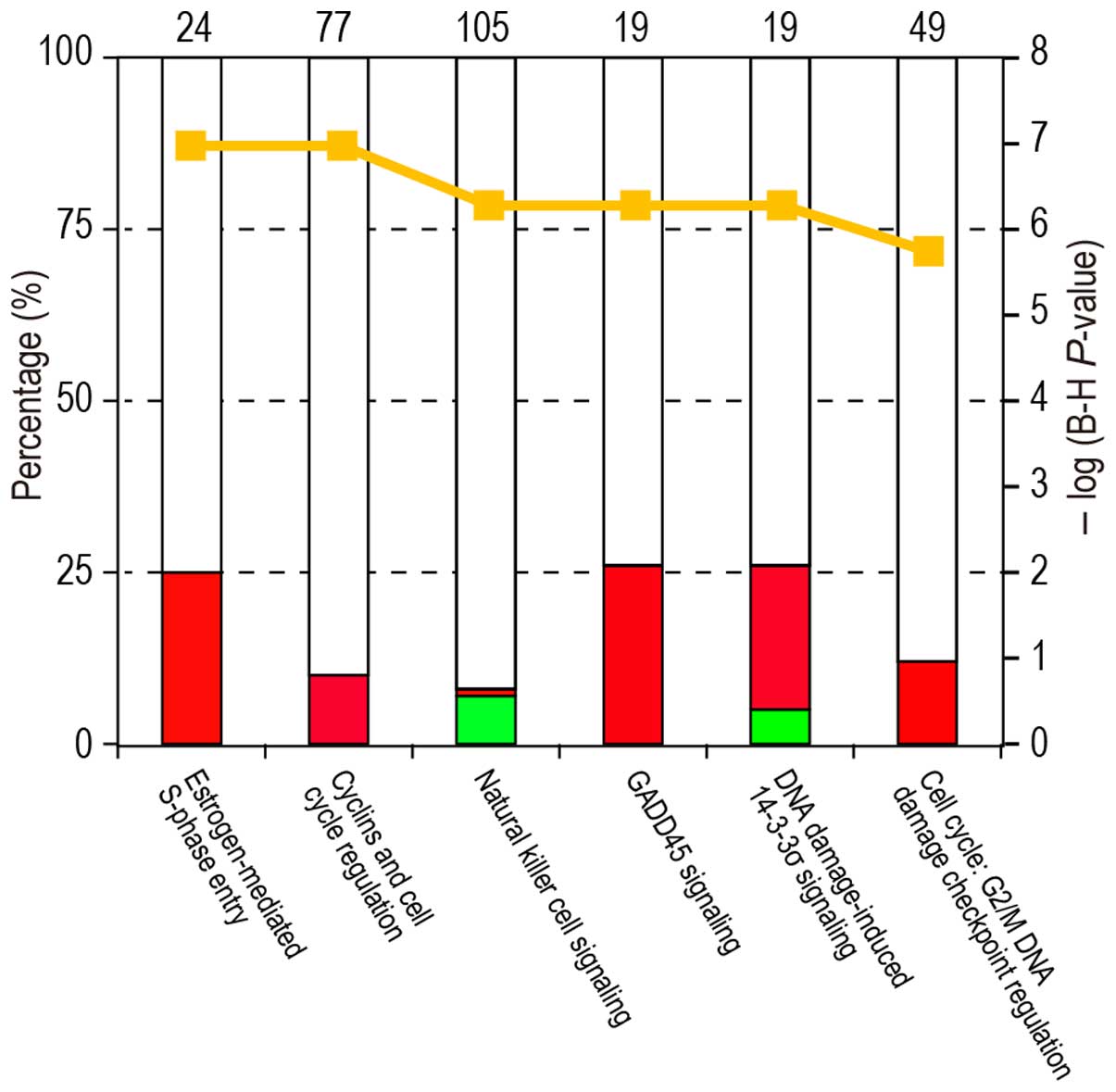
The top canonical pathways and gene networks that were most
significantly affected in the pNK cells during pregnancy are shown
in Fig. 4. High-ranking canonical
pathways included the NK cell signaling pathway
(P=5.13×10−7), as well as cell cycle regulation pathways
(e.g., estrogen-mediated S-phase entry pathway,
P=1.02×10−7). The NK cell signaling pathway contained
downregulated KIR genes (i.e., KIR2DL4, KIR2DL5A,
KIR2DS4 and KIR3DL1).
We found that 136 gene-related functions with a
minimum of ten genes per category in the IPA defined Diseases and
Bio Functions category (an IPA data set of the 136 functions is
available upon request). Moreover, we filtered the 136 functions by
NK cell context with a minimum of ten genes per category. The
filtering found 16 functions for NK cells; 54 genes were selected
among the 69 NK cell function-associated genes (Table II). Among the 16 functions, the
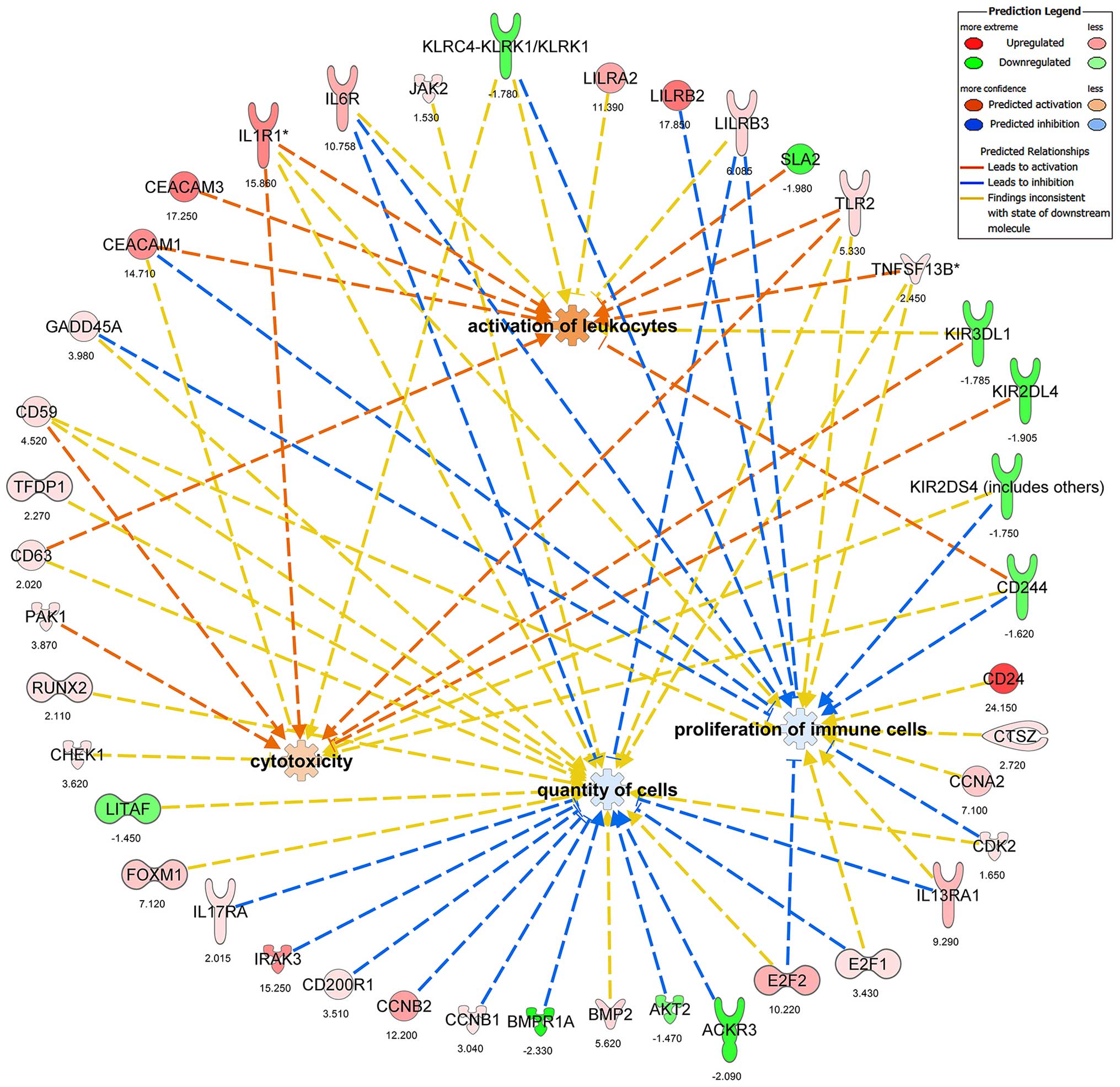
top scoring functions were the proliferation of immune cells (rank
8 relative to the 136 functions with P=5.96×10−10), the
quantity of cells (rank 11, P=1.02×10−9), the activation
of leukocytes (rank 15, P=3.68×10−9) and cytotoxicity
(rank 19, P=6.25×l0−9). Downstream Effects Analysis (as
defined in IPA) for visualization of the biological trends in this
study revealed that the proliferation of immune cells and quantity
of cells were predicted to decrease in the third- compared to the
first-trimester pNK cells (Fig.
5). By contrast, the activation of leukocytes and cytotoxicity
were predicted to increase in the third- compared to the
first-trimester pNK cells (Fig.
5). The combined network demonstrated that many genes in the
data set were upregulated (e.g., 1L1R1 and CEACAM1)
and that KIR genes were downregulated (Fig. 5).
| Table IIGene-related functions with a minimum
of ten genes per category in the IPA defined Diseases and Bio
Functions category as revealed by IPA Core Analysis. |
Table II
Gene-related functions with a minimum
of ten genes per category in the IPA defined Diseases and Bio
Functions category as revealed by IPA Core Analysis.
| Category | Function | Disease or function
annotation | P-value | P-value ranking of
136 gene-related functions | P-value ranking
filtered by NK cell context | Genea |
|---|
| Cellular
development, cellular growth and proliferation, hematological
system development and function | Proliferation | Proliferation of
immune cells |
5.96×l0−10 | 8 | 1 | CCNA2, CD24,
CD244, CD59, CDK2, CEACAM1, CTSZ, E2F1, E2F2, GADD45A,
IL13RA1,IL1R1, IL6R, JAK2, KIR2DS4 (includes
others), KLRC4-KLRK1/KLRK1, LILRB2, LILRB3, TLR2,
TNFSF13B |
| Tissue
morphology | Quantity | Quantity of
cells |
1.02×l0−9 | 11 | 2 | ACKR3, AKT2,
BMP2, BMPR1A, CCNA2, CCNB1, CCNB2, CD200R1, CD244, CD59, CD63,
CDK2, E2F1, E2F2, FOXM1, GADD45A, IL13RA1, IL17RA, IL1R1, IL6R,
IRAK3, JAK2, LILRB3, LITAF, RBBP8, RUNX2, TFDP1, TLR2,
TNFSF13B |
| Cell-to-cell
signaling and interaction, hematological system development and
function, immune cell trafficking, inflammatory response | Activation | Activation of
leukocytes |
3.68×l0−9 | 15 | 3 | CD244, CD59,
CD63, CEACAM1, CEACAM3, CEACAM6, GADD45A, IL1R1,
IL6R, JAK2, KIR3DL1, KLRC4-KLRK1/KLRK1, LILRA2,
LILRB2, LILRB3, SLA2, TLR2, TNFSF13B |
| Cell death and
survival | Cytotoxicity | Cytotoxicity |
6.25×l0−9 | 19 | 4 | CD244, CD59,
CEACAM1, CHEK1, IL1R1, KIR2DL4,KIR2DS4 (includes
others),KIR3DL1 KLRC4-KLRK1/KLRK1,PAK1,
TLR2 |
| Cell death and
survival, cellular compromise | Toxicity | Toxicity of
cells |
4.13×10−8 | 28 | 5 | CD244, CD59,
CEACAM1, IL1R1, KIR2DL4, KIR2DS4 (includes others),
KIR3DL1, KLRC4-KLRK1/KLRK1, PAK1,
TLR2 |
| Cell-to-cell
signaling and interaction, hematological system development and
function, immune cell trafficking, inflammatory response | Activation | Activation of
mononuclear leukocytes |
4.47×l0−8 | 29 | 6 | CD244, CD59,
CEACAM1, CEACAM6, GADD45A, IL6R, KIR3DL1,
KLRC4-KLRK1/KLRK1, LILRA2, LILRB2, L1LRB3, SLA2, TLR2,
TNFSF13B |
| Cell-to-cell
signaling and interaction, hematological system development and
function, immune cell trafficking, inflammatory response | Activation | Activation of
lymphocytes |
1.82×l0−7 | 38 | 7 | CD244, CD59,
CEACAM1, CEACAM6, GADD45A, IL6R, KIR3DL1,
KLRC4-KLRK1/KLRK1, LILRB2, LILRB3, SLA2, TLR2,
TNFSF13B |
| Cell-to-cell
signaling and interaction, hematological system development and
function, immune cell trafficking, inflammatory response | Activation | Activation of T
lymphocytes |
5.90×10−7 | 44 | 8 | CD244, CD59,
CEACAM1, CEACAM6, GADD45A, IL6R, KIR3DL1,
KLRC4-KLRK1IKLRK1, LILRB2, SLA2, TLR2 |
| Cellular function
and maintenance | Homeostasis | Lymphocyte
homeostasis |
1.34×10−6 | 53 | 9 | AKT2, CCNB2,
CD244, CDK2, CHEK1, E2F1, E2F2, IL17RA, IL1R1, LILRB3, RUNX2, TLR2,
TNFSF13B |
| Cell-mediated
immune response, cellular function and maintenance, hematological
system development and function | Homeostasis | T cell
homeostasis |
4.69×10−6 | 63 | 10 | AKT2, CCNB2,
CD244, CDK2, CHEK1, E2F1, E2F2, IL17RA, IL1R1, RUNX2, TLR2,
TNFSF13B |
| Hematological
system development and function, tissue morphology | Quantity | Quantity of
lymphocytes |
6.38×10−6 | 67 | 11 | ACKR3, CCNB2,
CD244, E2F1, E2F2, GADD45A, IL13RA1, IL17RA, IL1R1, JAK2, LILRB3,
RUNX2, TLR2, TNFSF13B |
| Immunological
disease | Systemic autoimmune
syndrome | Systemic autoimmune
syndrome |
1.20×10−5 | 77 | 12 | CCNB1, CD24,
CD244, CDK2, CEACAM1, CEACAM8, DUSP2, E2F1, IL17RA,
IL1R1, IL6R, IRAK3, JAK2, LILRB3, TLR2, TNFSF13B |
| Connective tissue
disorders, inflammatory disease, skeletal and muscular
disorders | Rheumatic
disease | Rheumatic
disease |
1.59×10−5 | 80 | 13 | BMP2, CCNB1,
CD200R1, CD24, CD244, CD59, CDK2, CEACAM1, CEACAM8,
DUSP2, IL17RA, IL1R1, IL6R, JAK2, KIR3DL1, TLR2,
TNFSF13B |
| Cell-to-cell
signaling and interaction | Communication | Communication of
cells |
1.11×10−4 | 108 | 14 | AKT2, BMP2,
CD244, CD59, CEACAM1, CEACAM6, IL13RA1, IL1R1, JAK2,
KIR2DL4, KLRC4-KLRK1/KLRK1, LILRA2, LILRB2, LILRB3,
LITAF, PAR1, SLA2, TLR2, TNFSF13B |
| Connective tissue
disorders, inflammatory disease, skeletal and muscular
disorders | Arthritis | Arthritis |
3.42×10−4 | 118 | 15 | BMP2, CCNB1,
CD200R1, CD24, CD244, CD59, DUSP2, IL17RA, IL1R1, IL6R, JAK2,
KIR3DL1, TLR2, TNFSF13B |
| Cancer | Breast or
colorectal cancer | Breast or
colorectal cancer |
2.44×10−3 | 136 | 16 | ACKR3,
AKT2, BMP2, BMPR1A,
CCNA2, CCNB1, CCNB2,
CD24, CD244, CDK2,
CDKN3, CEACAM1, CEACAM6,
CEACAM8, CHEK1, CTSZ,
DHRS7, DUSP2, FOXM1,
GLRX, HM13, IL17RA,
IL6R, IRF2BP2, JAK2,
KIR2DL4, KIR3DL1, KNTC1,
KPNA2, LILRA6, LILRB3,
MCM6, MKI67, PAK1,
PCNA, RBBP8, RUNX2,
TFDP1, TLR2, TNFAIP2 |
In silico analysis of miRNA-mRNA
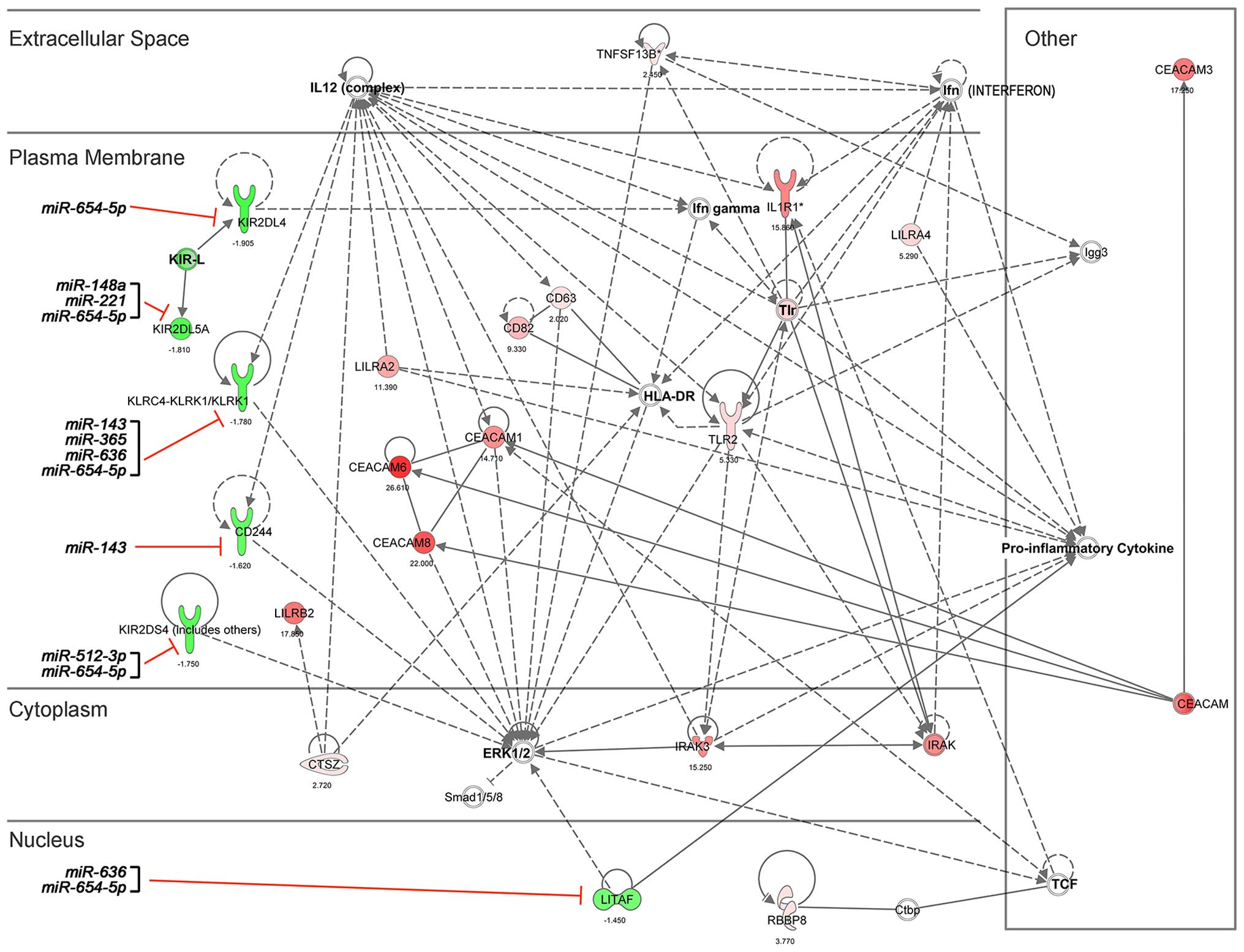
interactions in pNK cells during pregnancy
As upregulated miRNAs promoted mRNA degradation or
repression in pNK cells during pregnancy, a subset of downregulated
genes would include miRNA target candidates. To identify these
targets as well as effector miRNAs, we used IPA and the
microRNA.org database (http://www.microrna.org/microrna/getMirnaForm.do)
to search for over-represented miRNA seed sequences. We identified
27 seeds originating from the 12 miRNAs that were significantly
upregulated in the third- compared to the first-trimester pNK cells
(Table I). At least one miRNA
target site was identified in 69% (9 of 13) of the downregulated NK
cell function-related genes, with an average of three miRNA target
sites per gene. Note that the target site of one upregulated C19MC
miRNA, miR-512-3p, was present in the 3′-UTR of the
upregulated KIR2DS4 gene. The mRNA-miRNA network that was
filtered by NK cell context with a minimum of 10 genes per category
is depicted in Fig. 6. One miRNA,
miR-7-2#, which was significantly downregulated in the
third- compared to the first-trimester cells did not have the
target sites of upregulated NK cell function-associated genes
during pregnancy.
Discussion
The miRNAs within the human imprinted C19MC, a
primate-specific miRNA cluster encompassing 46 miRNA genes in the
human genome, are expressed exclusively in the placenta; i.e.,
placenta-associated miRNAs (16–18). Placenta-associated miRNAs are
expressed in human villous trophoblasts and are secreted into the
maternal circulation via exosomes (19). During pregnancy, exosomes are
involved in cell-to-cell communication between the placenta and
peripheral blood immune cells. Taylor et al reported that
placenta-derived exosomes are present in the maternal circulation
and suppress T cells with exosome-associated proteins (e.g., Fas
ligand) (20). Although it is
likely that exosomal C19MC miRNAs contribute to placental-maternal
communication, few studies have examined whether exosomal
placenta-associated miRNAs modulate the expression of their targets
in recipient maternal immune cells. Recently, we demonstrated the
exosome-mediated transfer of a placenta-associated miRNA,
miR-517a-3p, into maternal pNK cells with the subsequent
modulation of a target, PRKG1 (13). We hypothesized that
placenta-associated miRNAs, as well as proteins (i.e., KLRK1
ligands) may be transferred via exosomes from placental
trophoblasts into maternal NK cells and modulate gene expression,
which possibly contributes to the maintenance of pregnancy.
In the present study, we performed comprehensive
miRNA and gene expression profiling of maternal pNK cells using a
combination of real-time PCR-based array and DNA micro-array
analyses. In particular, we focused on placenta-associated C19MC
miRNAs in pNK cells and found that C19MC miRNAs were incorporated
into maternal pNK cells during pregnancy (Figs. 1Figure 23). Moreover, the levels of incorporated
C19MC miRNAs were increased in the pNK cells in a gestational
stage-dependent manner and decreased rapidly following delivery
(Figs. 1Figure 23). These results are consistent with
those of our previous study, showing that the levels of
incorporated miR-517a-3p and miR-518b differed
significantly before and after delivery (13). These results, together with data
from our previous study, provide evidence that the transfer of
placental C19MC miRNAs into maternal pNK cells occurs during
pregnancy.
With respect to the transcriptional level, we
observed differential gene expression between the first- and
third-trimester pNK cells on DNA microarray analysis. The
differentially expressed genes included many NK cell
function-associated genes (Table
I). IPA Core Analysis revealed that many cell cycle regulation
pathways were included in high-ranking canonical pathways with the
upregulation of cell cycle- and proliferation-related genes
(Fig. 4). For example, the
estrogen-mediated S-phase entry pathway was significantly altered
as the highest-ranking canonical pathway. Estrogens induce the
proliferation of estrogen receptor-positive cells by stimulating
G1/S transition and the subsequent progression of the cell cycle
(21). NK cells express estrogen
receptors (22). The number of
uterine NK cells increase by the elevation of plasma estrogen
levels during pregnancy (23). In
the present study, cell cycle regulation pathways showed a trend
toward cell cycle progression in pNK cells during pregnancy. These
pathways may be of interest for further study.
On the other hand, the interpretation of other
functions in pNK cells affected by pregnancy may be more complex
than that of the cell cycle. The network generated by IPA
Downstream Effects Analysis revaled complex prediction of the
effects of gene expression changes in the third- compared to the
first-trimester pNK cells (Fig.
5). For example, the downregulation of KIR2DL4 and
KIR3DL1 indicates a prediction of leading to the activation
of NK cell cytotoxicity. Yusa et al examined the inhibitory
functions of the KIR2DL4 cytoplasmic domain using NK-like cell
lines that had a chimeric receptor of the KIR2DL4 cytoplasmic
domain fused to the extracellular and transmembrane domains of this
chimeric receptor KIR3DL1 and revealed that both receptors
exhibited similar capacity to inhibit cytotoxicity (24). Both KIR2DL4 and KIR3DL1 molecules
can decrease the cytotoxicity of NK cells. In the network, the
upregulation of CEACAM1, a member of the carcinoembryonic
antigen gene family, indicates a prediction of leading to the
activation of leukocytes. Lu et al investigated Ceacam1 as a
regulator of graft-vs.-host disease and graft-vs.-tumor after
allogeneic bone marrow transplantation in mouse models, and
demonstrated that Ceacam1 regulated T-cell activation and the
number of donor T cells in lymphoid organs and graft-vs.-host
disease target tissues (25). In
the network, the upregulation of LILRB2 indicates a
prediction of leading to the inhibition of immune cell
proliferation. Human LILRB2 inhibits NK T-cell activation by
directly binding to CD1D and preventing antigen loading, thus
leading to reduction of T-cell proliferation (26–28). In the network, another upregulated
LILR gene, LILRB3, indicates a prediction of leading to the
inhibition of cell quantity. Older Lilrb3−/− mice
have been shown to have an increased number of peritoneal B1 B
lymphocytes (29), indicating
that the upregulation of Lilrb3 decreased the number of B1 B cells.
Taken together, it is inferred from IPA Downstream Effects Analysis
that gene expression changes in pNK cells may contribute to the
increase in the cytotoxicity of third-compared to first-trimester
pNK cells. In addition to NK cell cytotoxicity, pNK cells may also
influence adaptive immune responses by acting on dendritic cells
and T/B cells, and NK cells may be involved in controlling the
proliferation of these immune cells. However, it should be noted
that the changes in expression of genes indicated with yellow lines
in the network shown in Fig. 5
were inconsistent with the predicted expression changes of these
genes with respect to NK functions generated in this study.
NK cells consist of two distinct subsets based on
the levels of expression of the low-affinity Fc γ receptor IIIA
(CD16) and neural cell adhesion molecule (CD56) receptors.
CD56dim CD16pos NK cells predominantly
mediate cytotoxic effector functions (i.e., natural cytotoxic
activity and antibody-dependent cellular cytotoxicity), while
CD56bright CD16neg NK cells primarily produce
cytokines. The pNK cells are mainly CD56dim
CD16pos (30,31). The majority of pNK cells appear to
produce low levels of cytokines, but show a high-level expression
of KIRs. Thus, pNK cells are potent cytotoxic effector cells. In
this study, an activating KIR gene (KIR2DS4) and activating
receptor NKG2-D type II integral membrane protein gene
(KLRK1) showed a significantly decreased expression during
pregnancy, and inhibitory KIR genes (KIR2DL4, KIR2DL5A, and
KIR3DL1) simultaneously showed a decreased expression
(Table I). By contrast, the
expression levels of both an activating LILR gene (LILRA2)
and inhibitory LILR genes (LILRB2 and LILRB3) in the
third-trimester pNK cells were significantly higher than those in
the first-trimester cells (Table
I). These differentially expressed genes of immune receptors
did not represent a universal shift toward the same direction of
functional response of pNK cells to cytotoxicity. However, previous
studies have suggested that NK cells can adapt their responsiveness
to a changing environment in vivo (32,33). It is possible that pNK cells
adjust to different gestational conditions during pregnancy.
In conclusion, the present study provides details of
the miRNA and gene expression profiles in pNK cells during
pregnancy. PCR-based array analysis indicated that 25 miRNAs,
including six placenta-derived miRNAs (C19MC miRNAs), were
significantly upregulated in the third- compared to the
first-trimester pNK cells. pNK cells incorporated C19MC miRNAs, and
these interactions were possibly mediated via exosomes. The rapid
clearance of C19MC miRNAs also occurred in the NK cells following
delivery. DNA microarray analysis revealed that 69 NK cell
function-related genes were differentially expressed between the
first- and third-trimester pNK cells. Pathway and network analysis
suggested that these gene expression changes of pNK cells
contribute to the increase in the cytotoxicity, as well as the cell
cycle progression of third-trimester pNK cells. Thirteen of the 69
NK cell function-related genes were significantly downregulated
between the first- and third-trimester pNK cells. Nine of the 13
downregulated NK function-associated genes were in silico
target candidates of 12 upregulated miRNAs, including C19MC miRNA
miR-512-3p. Further in vitro experiments are required
to validate the NK cell function-related genes as real targets of
miRNAs identified in this study. Although recent studies have
revealed that miRNAs regulate fundamental NK cell processes,
including cytotoxicity, cytokine production and proliferation
(34,35), the functions of placental C19MC
miRNAs in maternal plasma also remain to be elucidated. Such
studies are currently in progress in our laboratory. Further
characterization of the subsets of maternal NK cells and the
discovery of target genes of C19MC miRNAs may provide further
insight into the understanding of maternal NK cell functions (i.e.,
the establishment and maintenance of pregnancy, and parturition
outbreak mechanism) during pregnancy.
Acknowledgments
The authors are indebted to Mr. Takuji Kosuge and
Ms. Kazuko Tanaka (Nippon Medical School) for their technical
assistance. This study was supported in part by Grants-in-Aids for
Scientific Research (no. 23390386 to S. Saito; no. 24390383 to T.
Takizawa) from the Ministry of Education, Culture, Sports, Science
and Technology (MEXT)/Japan Society for the Promotion of Science
(Japan), by Grant-in-Aid [no. H26-itaku(nan)-ippan-082 to S. Saito]
from the Ministry of Health, Labour and Welfare (Japan), and by a
grant from the MEXT-Supported Program for the Strategic Research
Foundation at Private Universities (2013–2017) to T. Takizawa.
References
|
1
|
Vivier E, Tomasello E, Baratin M, Walzer T
and Ugolini S: Functions of natural killer cells. Nat Immunol.
9:503–510. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
2
|
Vivier E, Raulet DH, Moretta A, Caligiuri
MA, Zitvogel L, Lanier LL, Yokoyama WM and Ugolini S: Innate or
adaptive immunity? The example of natural killer cells. Science.
331:44–49. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Carrega P and Ferlazzo G: Natural killer
cell distribution and trafficking in human tissues. Front Immunol.
3:3472012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kalkunte S, Chichester CO, Gotsch F,
Sentman CL, Romero R and Sharma S: Evolution of non-cytotoxic
uterine natural killer cells. Am J Reprod Immunol. 59:425–432.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Santoni A, Carlino C, Stabile H and
Gismondi A: Mechanisms underlying recruitment and accumulation of
decidual NK cells in uterus during pregnancy. Am J Reprod Immunol.
59:417–424. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Moffett A and Colucci F: Uterine NK cells:
Active regulators at the maternal-fetal interface. J Clin Invest.
124:1872–1879. 2014. View
Article : Google Scholar : PubMed/NCBI
|
|
7
|
Hedlund M, Stenqvist AC, Nagaeva O,
Kjellberg L, Wulff M, Baranov V and Mincheva-Nilsson L: Human
placenta expresses and secretes NKG2D ligands via exosomes that
down-modulate the cognate receptor expression: Evidence for
immunosuppressive function. J Immunol. 183:340–351. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bartel DP: MicroRNAs: Target recognition
and regulatory functions. Cell. 136:215–233. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Rossi RL, Rossetti G, Wenandy L, Curti S,
Ripamonti A, Bonnal RJ, Birolo RS, Moro M, Crosti MC, Gruarin P, et
al: Distinct microRNA signatures in human lymphocyte subsets and
enforcement of the naive state in CD4+ T cells by the
microRNA miR-125b. Nat Immunol. 12:796–803. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Allantaz F, Cheng DT, Bergauer T,
Ravindran P, Rossier MF, Ebeling M, Badi L, Reis B, Bitter H,
DAsaro M, et al: Expression profiling of human immune cell subsets
identifies miRNA-mRNA regulatory relationships correlated with cell
type specific expression. PLoS One. 7:e299792012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wang P, Gu Y, Zhang Q, Han Y, Hou J, Lin
L, Wu C, Bao Y, Su X, Jiang M, et al: Identification of resting and
type I IFN-activated human NK cell miRNomes reveals microRNA-378
and microRNA-30e as negative regulators of NK cell cytotoxicity. J
Immunol. 189:211–221. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Liu X, Wang Y, Sun Q, Yan J, Huang J, Zhu
S and Yu J: Identification of microRNA transcriptome involved in
human natural killer cell activation. Immunol Lett. 143:208–217.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kambe S, Yoshitake H, Yuge K, Ishida Y,
Ali MM, Takizawa T, Kuwata T, Ohkuchi A, Matsubara S, Suzuki M, et
al: Human exosomal placenta-associated miR-517a-3p modulates the
expression of PRKG1 mRNA in Jurkat cells. Biol Reprod. 91:1292014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
15
|
Dybkaer K, Iqbal J, Zhou G, Geng H, Xiao
L, Schmitz A, d’Amore F and Chan WC: Genome wide transcriptional
analysis of resting and IL2 activated human natural killer cells:
Gene expression signatures indicative of novel molecular signaling
pathways. BMC Genomics. 8:2302007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Morales-Prieto DM, Ospina-Prieto S,
Schmidt A, Chaiwangyen W and Markert UR: Elsevier Trophoblast
Research Award Lecture: Origin, evolution and future of placenta
miRNAs. Placenta. 35(Suppl): pp. S39–S45. 2014, View Article : Google Scholar
|
|
17
|
Bentwich I, Avniel A, Karov Y, Aharonov R,
Gilad S, Barad O, Barzilai A, Einat P, Einav U, Meiri E, et al:
Identification of hundreds of conserved and nonconserved human
microRNAs. Nat Genet. 37:766–770. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Noguer-Dance M, Abu-Amero S, Al-Khtib M,
Lefèvre A, Coullin P, Moore GE and Cavaillé J: The primate-specific
microRNA gene cluster (C19MC) is imprinted in the placenta. Hum Mol
Genet. 19:3566–3582. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Luo SS, Ishibashi O, Ishikawa G, Ishikawa
T, Katayama A, Mishima T, Takizawa T, Shigihara T, Goto T, Izumi A,
et al: Human villous trophoblasts express and secrete
placenta-specific microRNAs into maternal circulation via exosomes.
Biol Reprod. 81:717–729. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Taylor DD, Akyol S and Gercel-Taylor C:
Pregnancy-associated exosomes and their modulation of T cell
signaling. J Immunol. 176:1534–1542. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Doisneau-Sixou SF, Sergio CM, Carroll JS,
Hui R, Musgrove EA and Sutherland RL: Estrogen and antiestrogen
regulation of cell cycle progression in breast cancer cells. Endocr
Relat Cancer. 10:179–186. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Curran EM, Berghaus LJ, Vernetti NJ,
Saporita AJ, Lubahn DB and Estes DM: Natural killer cells express
estrogen receptor-alpha and estrogen receptor-beta and can respond
to estrogen via a non-estrogen receptor-alpha-mediated pathway.
Cell Immunol. 214:12–20. 2001. View Article : Google Scholar
|
|
23
|
van den Heuvel MJ, Xie X, Tayade C,
Peralta C, Fang Y, Leonard S, Paffaro VA Jr, Sheikhi AK, Murrant C
and Croy BA: A review of trafficking and activation of uterine
natural killer cells. Am J Reprod Immunol. 54:322–331. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yusa S, Catina TL and Campbell KS: SHP-1-
and phosphotyrosine-independent inhibitory signaling by a killer
cell Ig-like receptor cytoplasmic domain in human NK cells. J
Immunol. 168:5047–5057. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Lu SX, Kappel LW, Charbonneau-Allard AM,
Atallah R, Holland AM, Turbide C, Hubbard VM, Rotolo JA, Smith M,
Suh D, et al: Ceacam1 separates graft-versus-host-disease from
graft-versus-tumor activity after experimental allogeneic bone
marrow transplantation. PLoS One. 6:e216112011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Van Kaer L: Regulation of immune responses
by CD1d-restricted natural killer T cells. Immunol Res. 30:139–153.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ueda N, Kuki H, Kamimura D, Sawa S, Seino
K, Tashiro T, Fushuku K, Taniguchi M, Hirano T and Murakami M:
CD1d-restricted NKT cell activation enhanced homeostatic
proliferation of CD8+ T cells in a manner dependent on
IL-4. Int Immunol. 18:1397–1404. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li D, Wang L, Yu L, Freundt EC, Jin B,
Screaton GR and Xu XN: Ig-like transcript 4 inhibits lipid antigen
presentation through direct CD1d interaction. J Immunol.
182:1033–1040. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Ujike A, Takeda K, Nakamura A, Ebihara S,
Akiyama K and Takai T: Impaired dendritic cell maturation and
increased T(H)2 responses in PIR-B(−/−) mice. Nat Immunol.
3:542–548. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Cooper MA, Fehniger TA and Caligiuri MA:
The biology of human natural killer-cell subsets. Trends Immunol.
22:633–640. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Jabrane-Ferrat N and Siewiera J: The up
side of decidual natural killer cells: New developments in
immunology of pregnancy. Immunology. 141:490–497. 2014. View Article : Google Scholar :
|
|
32
|
Joncker NT, Shifrin N, Delebecque F and
Raulet DH: Mature natural killer cells reset their responsiveness
when exposed to an altered MHC environment. J Exp Med.
207:2065–2072. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Elliott JM, Wahle JA and Yokoyama WM: MHC
class I-deficient natural killer cells acquire a licensed phenotype
after transfer into an MHC class I-sufficient environment. J Exp
Med. 207:2073–2079. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Beaulieu AM, Bezman NA, Lee JE, Matloubian
M, Sun JC and Lanier LL: MicroRNA function in NK-cell biology.
Immunol Rev. 253:40–52. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Leong JW, Sullivan RP and Fehniger TA:
microRNA management of NK developmental and functional programs.
Eur J Immunol. 44:2862–2868. 2014. View Article : Google Scholar : PubMed/NCBI
|