Introduction
Prostate cancer is a pathological change, associated
with urinary dysfunction, in the prostate, which is one of the
three most life threatening diseases in males worldwide (1). Although it has been observed that
the incidence rate of prostate cancer is decreasing, its incidence
remains the highest in the world, particularly in the United States
(2). The mortality rate of
prostate cancer is lower than the sum of the mortality rates of
lung and bronchus cancer (3). A
number of males with prostate cancer never present with symptoms,
therefore, early detection and prevention is challenging (4).
Conventional treatments for prostate cancer include
surgery, chemotherapy, radiation therapy, radiofrequency ablation,
high-intensity focused ultrasound, cryosurgery and hormonal therapy
(5–7). The outcome of these therapeutic
approaches is beneficial for benign prostate tumor, however, the
survival rate of patients who present with cancer metastasis is low
(8). It has been reported that
the median overall survival rate of patients treated by
chemotherapy with enzalutamide is 18.4 months (9). Therefore, a novel therapeutic
strategy for prostate cancer is required. Gene therapy is one of
the most promising therapeutic approaches, as it delivers
therapeutic DNA into damaged cells, leading to fundamental healing
in the patients (10). One of the
most important issues is the identification of a specific gene
responsible for the prostate cancer and universally impacting on
different cell types in prostate cancer.
Garcia-Gonzalo et al reported that tectonic-1
(TCTN1) transports MKS1 and other MKS proteins to the transition
zone, between the basal body and ciliary axoneme (12). MKS1, as a basal body protein, has
a potential role in regulating Wnt signaling (11,12). The ciliary phenotype directly
represents the de-regulation of Wnt signaling in vitro and
in vivo (13,14). Wnt signaling regulates cell
proliferation, contributing to high proliferation rates in the
mutant kidney (15,16). Disturbance of Wnt signaling have
been demonstrated to cause varies diseases, including breast and
prostate cancer (17,18).
It has also been reported that MKS1 acts upstream of
Patched, and the loss of MKS1, which leads to Sonic hedgehog (Shh)
signaling causes a reduction in high-level Shh signaling (19). Shh is the most extensively
investigated ligand of the hedgehog signaling pathway among the
mammalian signaling pathway families (20). It is critical in the
differentiation and development of organs (21). MKS1 mutation in Hedgehog signaling
leads to hypoplasia, in various types of tumor (22), and inhibition of the Shh signaling
pathway has been identified as a possible treatment strategy for
gastric cancer (23).
Therefore, changes in the gene expression levels of
TCTN1 causes a chain reaction. Initially, it directly affects the
expression level of MKS1, the further actions of which affect Wnt
and Shh signaling. Variation in these two signaling pathways can
lead to overproliferation of cells and the progression of
cancer.
Despite substantial investigations in the gene
therapy field (24–27), the mechanism of action of a
specific gene target in prostate cancer remains unclear. Although
the effect of the TCTN1 gene on prostate cancer has been revealed
(28,29), a direct link between the TCTN1
gene and its effects on the viability of prostate cancer cells
remains to be elucidated. To investigate the role of the TCTN1 gene
in prostate cancer, the expression of TCTN1 gene was knocked down
using RNA interference lentivirus system in four prostate cancer
cell lines, PC-3, DU145, LNCaP and 22RV1. Biological function was
further evaluated by analyzing the effects of TCTN1 on cell growth,
cell cycle progression and cell migration. We aimed to reveal its
contribution to the progression of prostate cancer
Materials and methods
Cell culture
DU145, PC-3, LNCaP, 22RVI and 293T cells were
obtained from the Cell Bank of Chinese Academy of Sciences
(Shanghai, China). The DU145 cells were cultured in Ham's F-12
supplemented with 10% fetal bovine serum (FBS) and 1% NEAA. The
PC-3 cells were cultured in Ham's F-12 supplemented with 10% FBS.
LNCaP and 22RVI cells were cultured in RPMI-1640 supplemented with
10% FBS. The 293T cells were cultured in Dulbecco's modified
Eagle's medium (DMEM) supplemented with 10% FBS. All cell lines
were cultured at 37°C in humidified air with 5% CO2.
Lentivirus vector design and
production
The two lentivirus vectors were designed to knock
down the TCTN1 gene (NM_001082537.2) and to avoid the non-specific
knockdown effect, respectively. The short hairpin RNA (shRNA;
Shanghai Hollybio, Shanghai, China), designed to silence TCTN1 had
the following sequence:
5′-GCTCAGATGCATCAGTTCCTTCTCGAGAAGGAACTGATGCATCTGAGCTTTTTT-3′. The
control shRNA had the following sequence:
5′-GCGGAGGGTTTGAAAGAATATCTCGAG ATATCTTTCAAACCCTCCGCTTTTTT-3′. The
stem-loop-stem oligos (shRNAs) were synthesized, annealed and
ligated into a NheI/PacI-linearized pFH-L vector
(Shanghai Hollybio) containing the green fluorescent protein (GFP)
gene as a reporter. Following DNA sequencing confirmation, using
Lipofectamine® 2000 (Invitrogen Life Technologies,
Carlsbad, CA, USA), the successfully constructed vectors,
containing the pVSVG-I and pCMVΔR8.92 plasmids (Shanghai Hollybio),
were transfected into the 80% confluent 293T cells for 48 h at
37°C, which were harvested 72 h following transfection and then
purified by ultracentrifugation.
Lentivirus transduction and gene
knockdown
The PC-3 and DU145 cells (5×104
cells/well) were seeded into 6-well plates and transduced with
either the TCTN1 shRNA lentivirus (Lv-shTCTN1) or control shRNA
lentivirus (Lv-shCon) at a multiplicity of infection of 40,
respectively. Fluorescence microscopy was then used to observe the
tranduction efficiency 96 h post-infection.
Quantitative polymerase chain reaction
(qPCR)
qPCR was performed to determine the gene expression
levels of TCTN1 in the PC-3, DU145, LNCaP and 22RV1 cell lines on a
Bio-Rad Connet Real-Time PCR platform (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). Total RNA was extracted using
TRIzol® reagent (Invitrogen Life Technologies) and
reverse-transcribed into cDNA using Moloney Murine Leukemia Virus
Reverse Transcriptase (Promega Corp., Madison, WI, USA). The qPCR
reaction system consisted of 2X SYBR Premix Ex Taq (10 µl),
forward and reverse primers (2.5 µM; 0.8 µl), cDNA (5
µl) and ddH2O (4.2 µl). The β-actin gene
was used as an internal control. For β-actin, the forward primer
sequences were as follows: Forward 5′-GTGGACATCCGCAAAGAC-3′ and
reverse 5′-AAAGGGTGTAACGCAACTA-3′. For TCTN1, the primer sequences
were as follows: Forward 5′-CCTTTGCGTGAATGTTGTTC-3′ and reverse
5′-AGAGGGACTGGCTGGGTATT-3′. The qPCR cycle was performed as
follows: Initial denaturation at 95°C for 1 min, denaturation at
95°C for 5 sec and annealing extension at 60°C for 20 sec. A total
of 40 cycles were performed. The cycle threshold (Ct) value,
normalized with that of β-actin was used to determine the rela-tive
expression of TCTN1, using the 2−ΔΔCt formula (30).
Western blotting
The lentivirus-transduced cells (10,000 cells/well)
were lysed in 2X SDS sample buffer, containing 100 mM Tris-HCl (pH
6.8), 10 mM EDTA, 4% SDS and 10% glycine. The proteins were
separated by 10% sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE). In each lane of the gels, 30 µg
protein was added and electrophoresis was performed under 50 V for
3 h. Subsequently a polyvinylidene fluoride (PVDF; Millipore,
Bedford, MA, USA) transmembrane procedure was performed under 300
mA for 1.5 h. The membrane was then incubated with indicated
primary antibodies [rabbit anti-TCTN1 (1:1,000; Cat. no.
SAB3500518; Sigma-Aldrich, St. Louis, MO, USA) and rabbit
anti-GAPDH (1:60,000; Cat. no. 10494-1-AP; Proteintech Group, Inc.,
Chicago, IL, USA)] at 4°C overnight, followed by incubation with
horseradish peroxidase-conjugated secondary antibodies (1:5,000;
Cat. no. sc-2054; Santa Cruz Biotechnology, Inc., Santa Cruz, CA,
USA) at room temperature for 2 h. Horseradish peroxidase
glyceraldehyde 3-phosphate dehydrogenase protein was used as a
loading control.
MTT assay
Following lentivirus transduction, the DU145 (2,500
cells/well) and PC-3 (2,000 cells/well) cells were seeded into
96-well plates, respectively. The numbers of cells were measured at
time‑points indicated in the figures. MTT solution (5 mg/ml; 20
µl) was added into each well. The MTT solution was aspirated
off following incubation for 4 h at 37°C. Subsequently, 100
µl acidic isopropanol, containing 10% SDS, 5% isopropanol
and 0.01 mol/L HCl, was added. The absorbance of each plate was
measured at 595 nm using spectrophotometer (Epoch; BioTek,
Winooski, VT, USA).
Colony formation assay
Following lentivirus tranduction, the PC-3 (300
cells/well) and DU145 (500 cells/well) cells were seeded into
6-well plates and maintained at 37°C for 8 days, respectively. The
culture media were replaced every 2–3 days. When the colonies were
formed, the plate was washed and fixed with paraformaldehyde,
stained with crystal purple for 20 min and washed three times using
ddH2O, sequentially. These cells were then photographed
using a digital camera (D7000; Nikon Corp., Tokyo, Japan). The
number of colonies containing >50 cells/colony was then
counted.
Flow cytometric analysi
Following lentivirus transduction, the PC-3 and
DU145 cells were seeded into 6-cm dishes at a density of
1×105 and 2×105 cells/dish, respectively. The
cells were harvested following trypsinization, washed with
phosphate‑buffered saline (PBS) and fixed in 80% ethanol at −20°C
for 24 h. The cells were then collected and centrifuged at 214.2 x
g for 10 min at 4°C, resuspended in the staining solution
containing 100 µg/ml RNase A and 50 µg/ml propidium
iodide in PBS, and incubated for 1 h at 37°C. The stained cells
were subjected to flow cytometric analysis using a FACSCalibur II
sorter and CellQuest FACS system (BD Biosciences, San Jose, CA,
USA).
Transwell migration assay
The cell migration was determined by the number of
cells that migrated through an 8-µm pore Transwell
polycarbonate membrane (Corning, Inc., Union City, CA, USA),
separating the upper and lower chamber. An equal number of cells
(5×104 PC-3 cells or 1×105 DU145 cells) in
the three groups were seeded into the upper chamber with 200
µl of serum-free medium. Subsequently, 500 µl medium,
containing 10% FBS for the PC-3 cells or 20% FBS for the DU145
cells was added to the lower chamber. The fully prepared Transwell
migration system was then placed in an incubator for 24 h at 37°C
in 5% CO2. Finally, the cells on the surface of the
basement membrane were removed, and the migrated cells were stained
using crystal violet (0.05%) and counted under a microscope, in
which five randomly-selected fields were observed for each
sample.
Statistical analysis
Statistical analysis was performed using Prism 5 for
Windows software (GraphPad Software, San Diego, CA, USA) Data are
presented as the mean ± standard deviation from at least three
independent experiments, performed in triplicate. Statistical
significance was determined using Student's t-test. P<0.05 was
considered to indicate a statistically significant difference.
Results
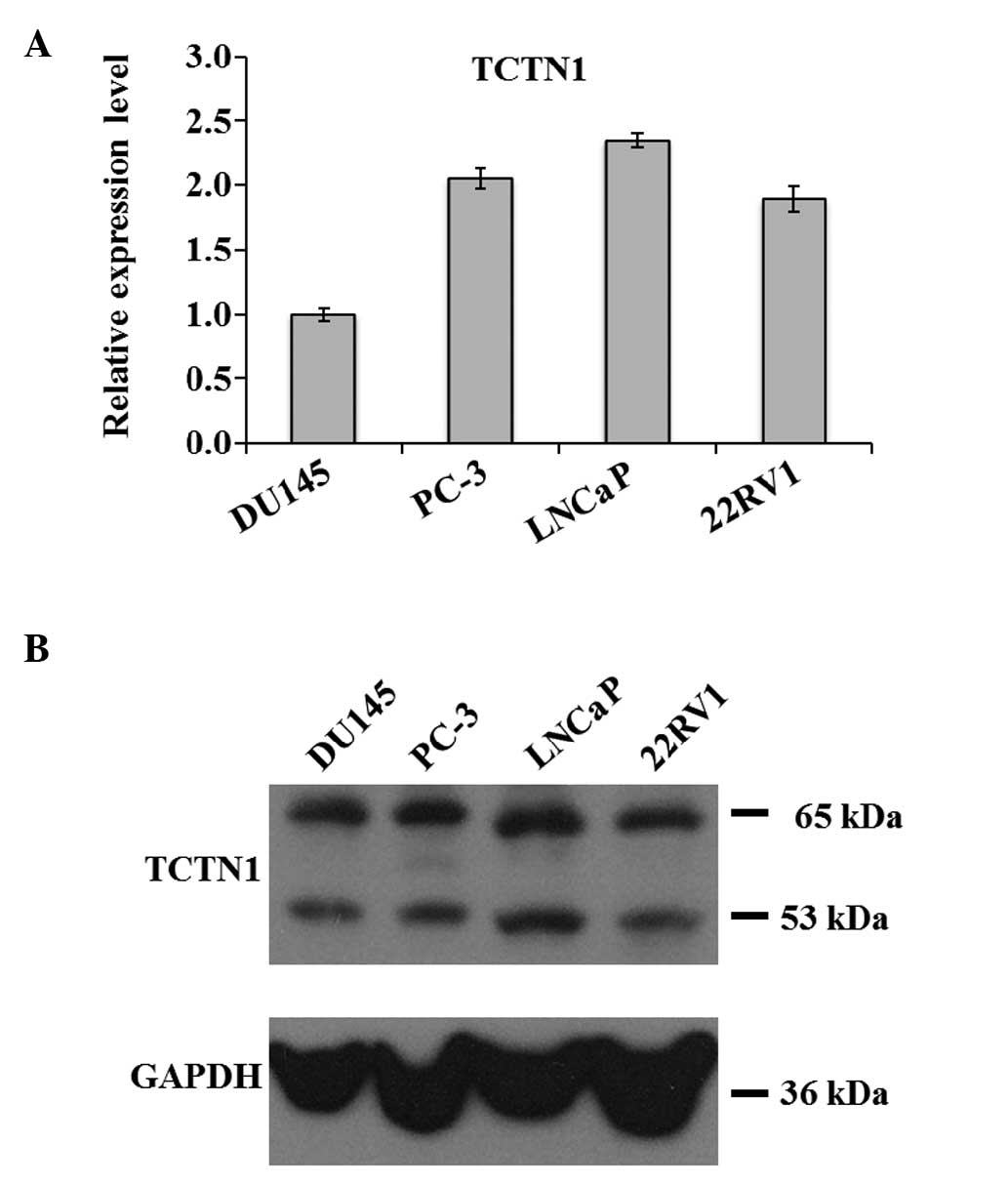
Expression of TCTN1 is upregulated in
prostate cancer cells
The DU145, PC-3, LNCaP and 22RV1 human prostate
cancer cell lines were cultured to measure the expression levels of
TCTN1 using qPCR and western blot analysis. TCTN1 mRNA was detected
in all the prostate cancer cell lines (Fig. 1A). The protein expression of TCTN1
was also confirmed in all the cell lines (Fig. 1B). These results indicated that
TCTN1 was involved in prostate cancer. As the expression of TCTN1
was consistently high in these four cell lines, it may serve as a
specific biomarker for the early detection of prostate cancer.
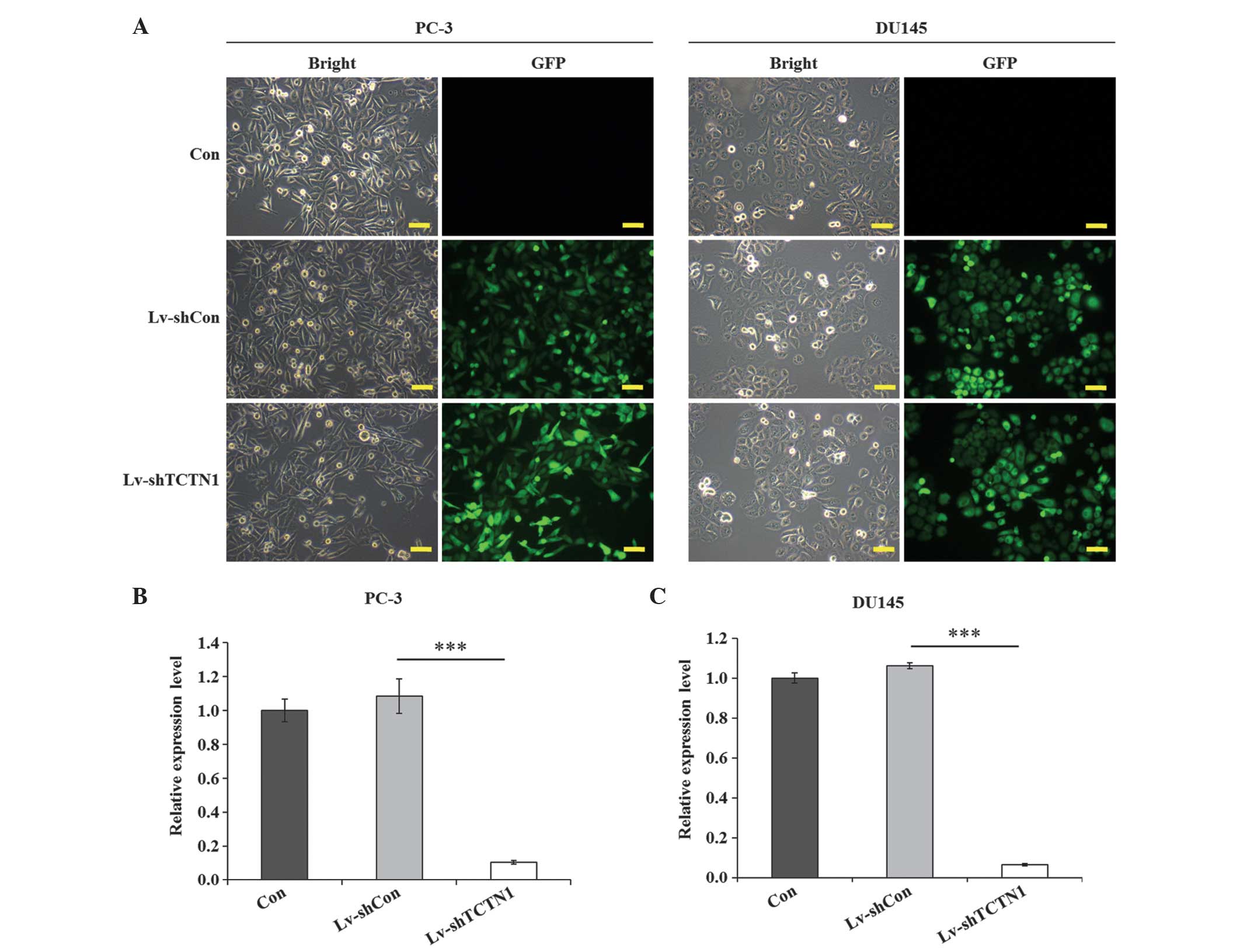
Expression of TCTN1 is successfully
downregulated by lentivirus-mediated RNAi in prostate cancer
cells
A specific Lv-shTCTN1 system was designed to silence
the TCTN1 gene, and a control shRNA (Lv-shCon) was also constructed
to eliminate the non-specific gene-silencing effect of the
lentivirius alone. The lentiviral transduction efficiency was
recorded using a fluorescence microscope (BX50; Olympus, Tokyo,
Japan), in which >80% of the cells expressed fluorescence in the
lentivirus transfection groups in the PC-3 and DU145 cell lines,
whereas the non-infected control groups exhibited no fluorescence
(Fig. 2A). This suggested the
efficiency of the lentiviral transduction was stable and
substantial. qPCR was then performed to evaluate the knockdown
efficiency of TCTN1 following lentivirus tranduction. The
expression levels of TCTN1 in the control (Con) and Lv-shCon groups
were similar, however, compared with the Lv-shCon group, the
expression of TCTN1 was reduced by 90.3% in the PC-3 cells
(P<0.01; Fig. 2B) and 94.0% in
the DU145 cells (P<0.001; Fig.
2C). This indicated that the expression of TCTN1 was
specifically knocked down by the lentivirus system in the two cell
lines. Therefore, Lv-shTCTN1 was a stable lentivirus system which
efficiently downregulated the expression of TCTN1 in the prostate
cancer cells.
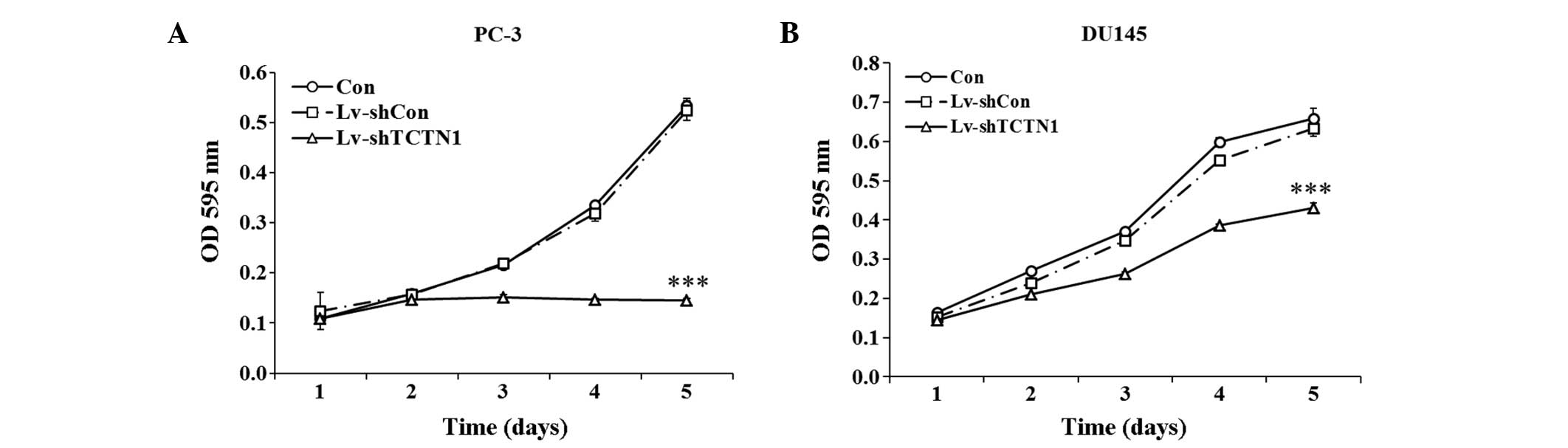
Knockdown of the TCTN1 gene inhibits the
viability and proliferation of prostate cancer cells
In the PC-3 cells, the growth curves of the Con and
Lv-shCon groups were overlapping, with almost no difference
observed between them (Fig. 3A).
By contrast, the growth curve of the Lv-shTCTN1 group was markedly
lower, compared with that of the Lv-shCon group from day 2. On day
5, the optical density (OD) value at 595 nm of the PC-3 cells in
the Lv-shTCTN1 group remained at 0.1454±0.0042, which was markedly
lower than that in the Con group (0.5334±0.0148) and Lv-shCon group
(0.5242±0.0199). However, cell proliferation increased
exponentially on day 5 in the Con and Lv-shCon groups. Similar
growth curve trends were observed in the DU145 cells, as shown in
Fig. 3B. The number of viable
cells in the Lv-shTCTN1 group was markedly lower than that in
either the Con group or Lv-shCon group. On day 5, the OD value at
595 nm of the DU145 cells in the Lv-shTCTN1 group was
0.4312±0.0115, which was lower than that in the Con group
(0.6594±0.0257) and Lv-shCon group (0.6334±0.0211).
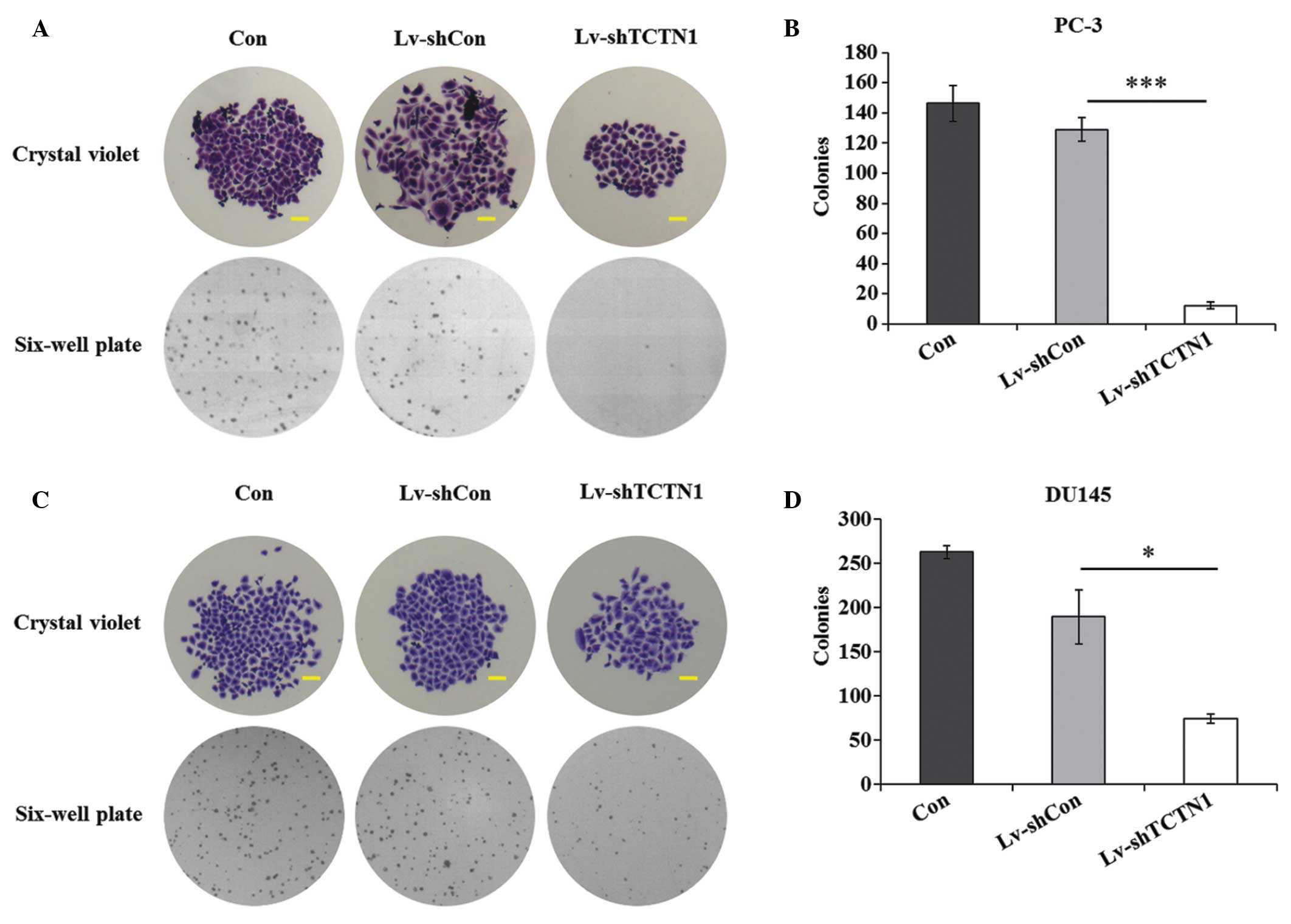
The present study also performed a colony formation
assay to determine the effect of TCTN1 on long-term cell
proliferation in the PC-3 and DU145 cell lines. Compared with the
Con and Lv-shCon groups, the size of a single colony in the
Lv-shTCTN1 group was substantially smaller in the images captured
of the crystal violet staining and bright field (Fig. 4A and C). In addition, there were
fewer colonies in the Lv-shTCTN1 group, compared with the Con and
Lv-shCon groups. Compared with the Lv-shCon group, the number of
colonies in the Lv-shTCTN1 group were reduced by 90.7% (Fig. 4B). Similar results were observed
in the DU145 cells (Fig. 4D).
Taken together, it is reasonable to conclude that
lentivirus-mediated TCTN1 silencing had a suppressive effect on
cell viability and proliferation of the prostate cancer cells.
Therefore it was suggested that TCTN1 may act as a potential
therapeutic target in prostate cancer.
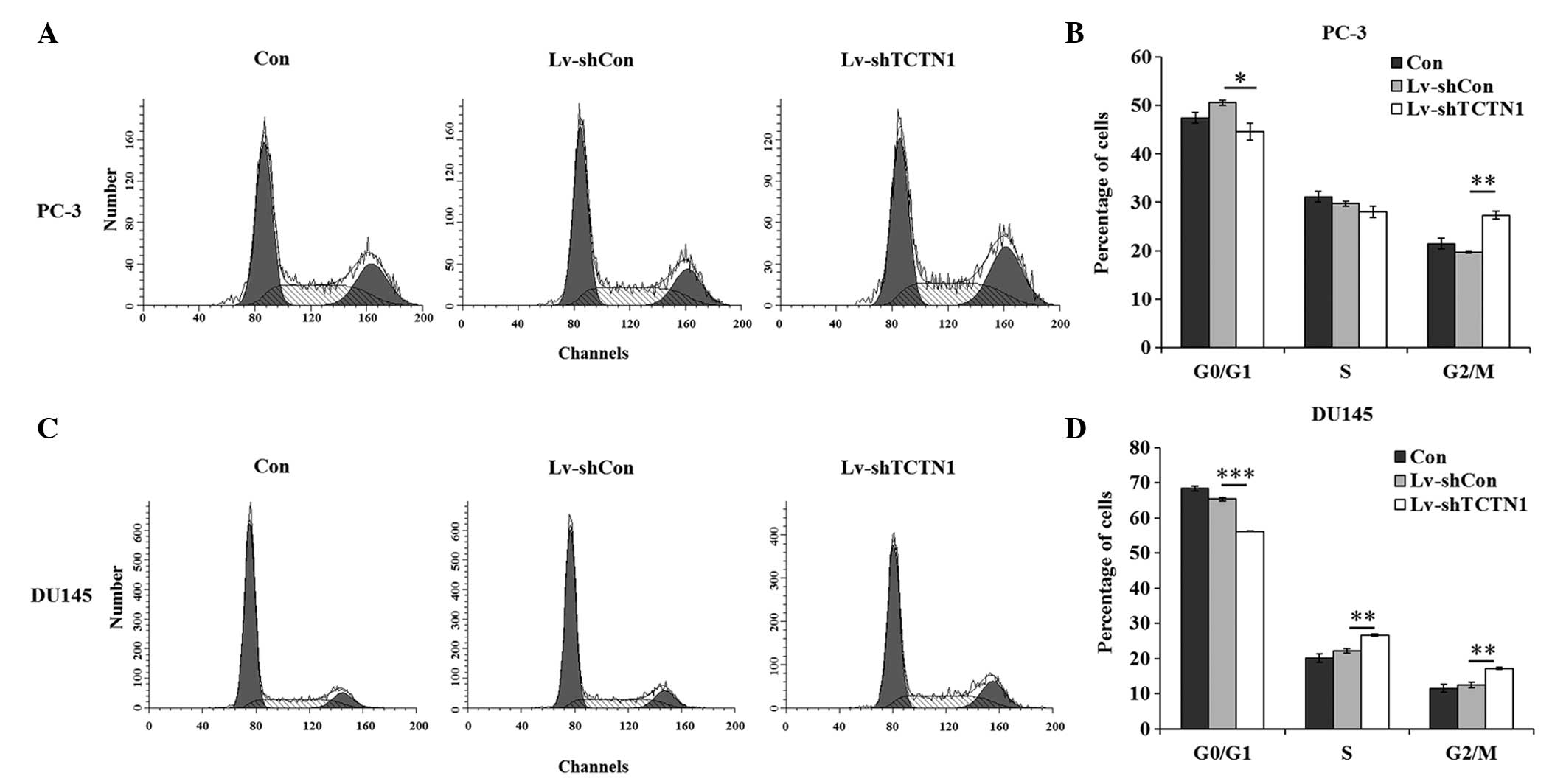
Knockdown of the TCTN1 gene inhibits the
cell cycle progression of prostate cancer cells
Flow cytometry, in conjunction with modeling
algorithms, distinguished cells in the different stages of the cell
cycle. TCTN1 silencing had a marked effect on cell cycle
progression (Fig. 5A and C). In
the PC-3 cells, the percentage of cells in the G0/G1 phase
decreased from 50.60±0.52% in the Lv-shCon group to 44.68±1.77% in
the Lv-shTCTN1 group. The percentage of cells in the G2/M phase
increased from 19.69±0.17% in the Lv-shCon group to 27.34±0.77% in
the Lv-shTCTN1 group. However, no significant difference was
observed between the Con and Lv-shCon groups (Fig. 5B). In the DU145 cells, the number
of cells in the G2/M phase in the Lv-shTCTN1 group was also lower,
compared with those in the Con and Lv-shCon groups. The percentages
of cells in the S phase and G2/M phase were markedly elevated in
the DU145 cells following TCTN1 silencing (Fig. 5D). There was a marked change in
the number of cells in the S phase, which may have been due to the
specific cell type. In conclusion, silencing of the TCTN1 gene
arrested cells at the G2/M phase, which impaired cell
proliferation.
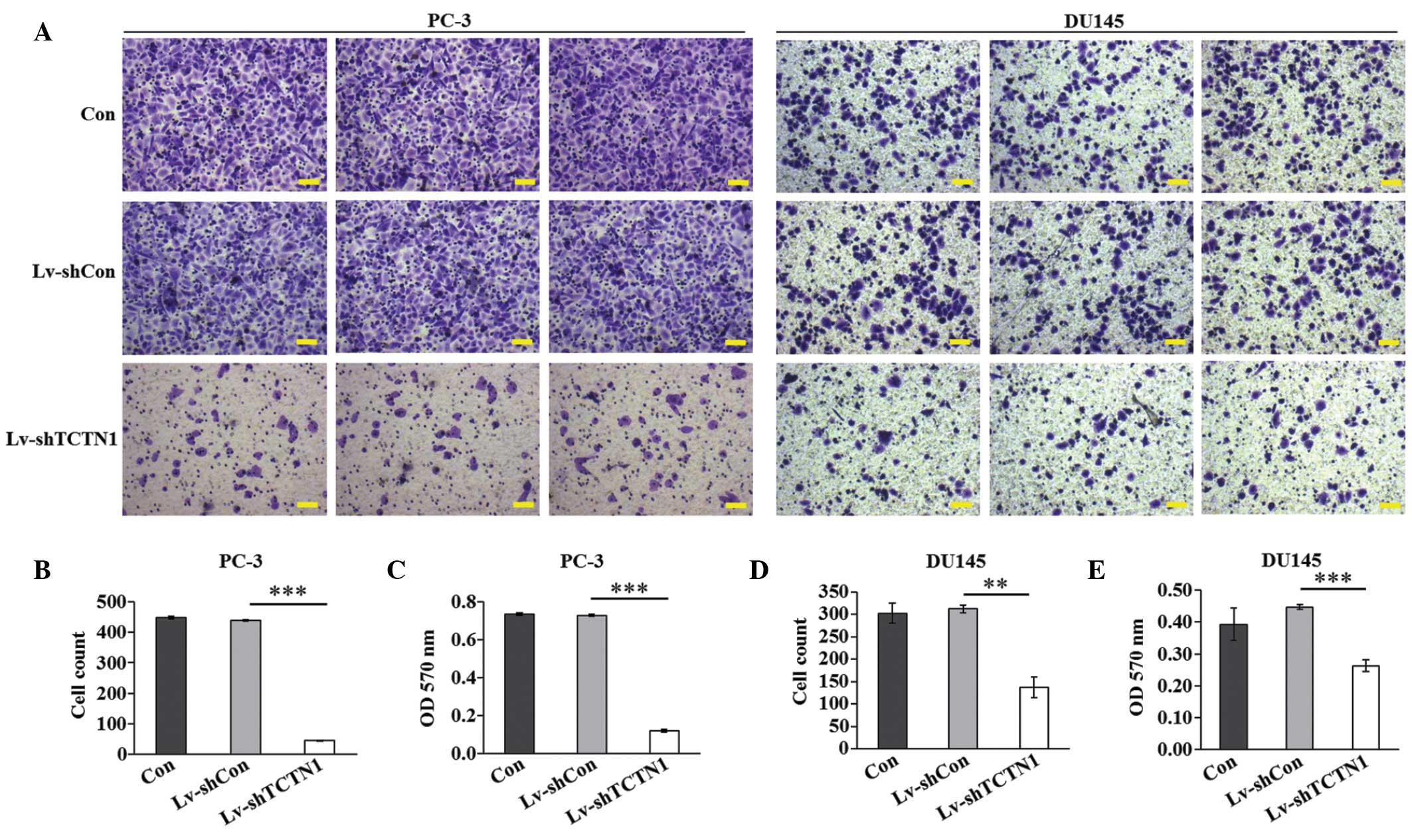
Knockdown of the TCTN1 gene inhibits the
migration of prostate cancer cells
Cell migration is a critical step during cancer
progression. The present study subsequently aimed to determine the
effect of TCTN1 knockdown in regulating prostate cancer cell
migration using a Transwell assay (Fig. 6A). In the PC-3 cells, fewer cells
in the Lv-shTCTN1 group (44.7±1.6) migrated to the lower surface of
the membrane, compared with the cells in the Con group (447.9±4.3)
or Lv-shCon group (440.1±2.3; Fig.
6B). In addition, the crystal violet staining intensity was
significantly lower in the Lv‑shTCTN1 group than in the Con and
Lv-shCon groups (Fig. 6C).
Similar results were observed in the DU145 cells (Fig. 6D and E), with knock-down of the
TCTN1 gene also disrupting the migration of the DU145 cells. These
results suggested that TCTN1 may be key in prostate cancer
metastasis.
Discussion
Prostate cancer has the highest incidence rate among
all types of cancers in males, and the mortality rate of prostate
cancer remains high, as some prostate cancer cells migrate into
lymph glands and through the lymphatic and circulatory systems
(31,32). Traditional treatment approaches
for prostate cancer are unable to cure it, with recurrence often
occurring shortly following treatment (33,34). It is well accepted that cancer
formation is a mutli-step process, involving continuous gene
mutation (35,36). Gene therapy is designed to deliver
an effective gene into a target site to regulate the expression of
a specific gene (37). Thus, it
is important to identify the gene responsible for the viability and
migration of prostate cancer cells to suppress cell growth in
situ.
It has been reported that the TCTN1 gene regulates
the expression of MKS1, which is associated with the expression of
the Wnt signaling and Shh signaling pathways. These two signaling
pathways share are involved in cell ove-proliferation, which is one
of six hallmarks of cancer cells (38). These findings prompted the present
study to investigate the TCTN1 gene as a target site in prostate
cancer therapy.
In the present study, the association between TCTN1
and characteristics of prostate cancer cells were initally
examined. The expression of TCTN1 was detected in four prostate
cancer cell lines, PC-3, DU145, LNCaP and 22RV1, which indicated
that high expression levels of TCTN1 may be associated with
prostate cancer. Subsequently, lentivirus-based shRNA expression
systems were introduced to specifically knock down the expression
of TCTN1 in the PC-3 and DU145 cells. In the absence of TCTN1, the
viability and colony formation ability were impaired in the PC-3
and DU145 cell lines, which suggested that TCTN1 may be essential
for the growth of prostate cancer cells in vitro. To
determine the cause of this cell growth suppression, flow
cytometery was performed to examine cell cycle progression. The
results indicated that the number of cells in the G2/M phase were
markedly increased in the Lv-shTCTN1 group, compared with the Con
and Lv-shCon groups. It has been reported that TCTN1 is a complex
that is localized at the transition zone of primary cilia (12), and that cilia are necessary for
tissue development and homeostasis, which emerge during interphase
prior to mitosis (39). Thus, the
knockdown of TCTN1 gene may arrest cells at the M phase, which is
consistent with the results of the present study. The arrest of the
cell cycle was essential in reducing the rate of cell growth. TCTN1
regulates the primary cilia, which are critical in modeling
cytoskeletal changes that impinge on cell migration (40,41). TCTN1 silencing also markedly
inhibited the ability of cells to migrate in prostate cancer.
In conclusion, the present study demonstrated that
TCTN1 was associated with the growth and migration of prostate
cancer cells in vitro. Silencing of the TCTN1 gene markedly
inhibited cell viability, proliferation and migration. Therefore,
TCTN1 may offer potential as biomarker for the treatment of
prostate cancer.
Acknowledgments
This study was supported by grants from the National
Natural Science Foundation of China (grant. no. 81170637), the
Shanghai Committee of Science and Technology General Program for
Medicine (grant. no. 11JC1402302), the Key Project of Science and
Innovation Foundation of Shanghai Ministry of Education (grant. no.
14zz084), and the Military Fund for Health Care (grant. no.
13BJZ29).
References
|
1
|
Calle EE, Rodriguez C, Walker-Thurmond K
and Thun MJ: Overweight, obesity, and mortality from cancer in a
prospectively studied cohort of US adults. New Engl J Med.
348:1625–1638. 2003. View Article : Google Scholar
|
|
2
|
Siegel R, Naishadham D and Jemal A: Cancer
Statistics, 2012. Ca Cancer J Clin. 62:10–29. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Parkin DM, Bray F, Ferlay J and Pisani P:
Global cancer statistics, 2002. CΑ Cancer J Clin. 55:74–108. 2005.
View Article : Google Scholar
|
|
4
|
Smolensky M: Book review: Light pollution
as a new risk factor for human breast and prostate cancers
(Springer pp168, 2013). Chronobiol Int. 30:1203–1204. 2013.
View Article : Google Scholar
|
|
5
|
Peyromaure M, Valéri A, Rebillard X, et
al: Characteristics of prostate cancer in men less than
50-year-old. Prog Urol. 19:803–809. 2009.In French. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Crouzet S, Rouviere O, Martin X and Gelet
A: High-intensity focused ultrasound as focal therapy of prostate
cancer. Curr Opin Urol. 24:225–230. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zhao X and Chua KJ: Regulating the
cryo-freezing region of biological tissue with a controlled thermal
device. Med Eng Phys. 36:325–334. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Baade PD, Youlden DR and Krnjacki LJ:
International epidemiology of prostate cancer: geographical
distribution and secular trends. Mol Nutr Food Res. 53:171–184.
2009. View Article : Google Scholar
|
|
9
|
Scher HI, Fizazi K, Saad F, et al:
Increased survival with enzalutamide in prostate cancer after
chemotherapy. New Engl J Med. 367:1187–1197. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Fitzgerald KA, Evans JC, McCarthy J, et
al: The role of transcription factors in prostate cancer and
potential for future RNA interference therapy. Expert Opin Ther
Targets. 18:633–649. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Wheway G, Abdelhamed Z, Natarajan S,
Toomes C, Inglehearn C and Johnson CA: Aberrant Wnt signalling and
cellular over-proliferation in a novel mouse model of Meckel-Gruber
syndrome. Dev Biol. 377:55–66. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Garcia-Gonzalo FR, Corbit KC,
Sirerol-Piquer M, et al: A transition zone complex regulates
mammalian ciliogenesis and ciliary membrane composition. Nat Genet.
43:776–784. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
13
|
Corbit KC, Shyer AE, Dowdle WE, Gaulden J,
Singla V, Chen MH, Chuang PT and Reiter JF: Kif3a constrains
beta-catenin-dependent Wnt signalling through dual ciliary and
non-ciliary mechanisms. Nat Cell Biol. 10:70–76. 2008. View Article : Google Scholar
|
|
14
|
Gerdes JM, Liu Y, Zaghloul NA, et al:
Disruption of the basal body comprises proteasomal function and
perturbs intracellular Wnt response. Nat Genet. 39:1350–1360. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Bohnenpoll T, Trowe MO, Wojahn I, Taketo
MM, Petry M and Kispert A: Canonical Wnt signaling regulates the
proliferative expansion and differentiation of fibrocytes in the
murine inner ear. Dev Biol. 391:54–65. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Liu YZ, Wu K, Huang J, et al: The
PTEN/PI3K/Akt and Wnt/β-catenin signaling pathways are involved in
the inhibitory effect of resveratrol on human colon cancer cell
proliferation. Int J Oncol. 45:104–112. 2014.PubMed/NCBI
|
|
17
|
Logan CY and Nusse R: The Wnt signaling
pathway in development and disease. Annu Rev. Cell Dev Biol.
20:781–810. 2004. View Article : Google Scholar
|
|
18
|
Komiya Y and Habas R: Wnt signal
transduction pathways. Organogenesis. 4:68–75. 2008. View Article : Google Scholar
|
|
19
|
Vierkotten J, Dildrop R, Peters T, Wang B
and Ruether U: Ftm is a novel basal body protein of cilia involved
in Shh signalling. Development. 134:2569–2577. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Ye W, Shimamura K, Rubenstein JL, Hynes MA
and Rosenthal A: FGF and Shh signals control dopaminergic and
serotonergic cell fate in the anterior neural plate. Cell.
93:755–766. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Dahmane N and Ruiz i Altaba A: Sonic
hedgehog regulates the growth and patterning of the cerebellum.
Development. 126:3089–3100. 1999.PubMed/NCBI
|
|
22
|
Hinterseher U, Wunderlich A, Roth S, et
al: Expression of hedgehog signalling pathway in anaplastic thyroid
cancer. Endocrine. 45:439–447. 2014. View Article : Google Scholar
|
|
23
|
Wan J, Zhou J, Zhao H, et al: Sonic
hedgehog pathway contributes to gastric cancer cell growth and
proliferation. Biores Open Access. 3:53–59. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Nishikawa R, Goto Y, Kojima S, et al:
Tumor-suppressive microRNA-29s inhibit cancer cell migration and
invasion via targeting LAMC1 in prostate cancer. Int J Oncol.
45:401–410. 2014.PubMed/NCBI
|
|
25
|
Tetsu O and McCormick F: Beta-catenin
regulates expression of cyclin D1 in colon carcinoma cells. Nature.
398:422–426. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Scholzen T and Gerdes J: The Ki-67
protein: From the known and the unknown. J Cell Physiol.
182:311–322. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ambrosini G, Adida C and Altieri DC: A
novel anti-apoptosis gene, survivin, expressed in cancer and
lymphoma. Nat Med. 3:917–921. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Shaw G, Price AM, Ktori E, Bisson I,
Purkis PE, McFaul S, Oliver RT and Prowse DM: Hedgehog signalling
in androgen independent prostate cancer. Eur Urol. 54:1333–1343.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Jamaluddin M, Liang Z, L J, Yao Q and Chen
C: Tectonic1, a new hedgehog signaling molecule, upregulates
endothelial nitric oxide synthase in human endothelial cells. J
Surg Res. 179(331)2013. View Article : Google Scholar
|
|
30
|
Gu J, Li K, Li M, Wu X, Zhang L, Ding Q,
Wu W, Yang J, Mu J, Wen H, et al: A role for p21-activated kinase7
in the development of gastric cancer. FEBS J. 280:46–55. 2013.
View Article : Google Scholar
|
|
31
|
Revelo MP, Cookson MS, Chang SS, Shook MF,
Smith JA Jr and Shappell SB: Incidence and location of prostate and
urothelial carcinoma in prostates from cystoprostatectomies:
implications for possible apical sparing surgery. J Urol. 171(2 Pt
1): 646–651. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Revelo MP, Cookson MS, Chang SS, Shook MF,
Smith JA Jr and Shappell SB: Incidence and location of prostate and
urothelial carcinoma in prostates from cystoprostatectomies:
Implications for possible apical sparing surgery. J Urol. 179(Suppl
5): S27–S32. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Bolla M, Gonzalez D, Warde P, et al:
Improved survival in patients with locally advanced prostate cancer
treated with radiotherapy and goserelin. N Engl J Med. 337:295–300.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
D'Amico AV, Whittington R, Malkowicz SB,
et al: Biochemical outcome after radical prostatectomy, external
beam radiation therapy, or interstitial radiation therapy for
clinically localized prostate cancer. JAMA. 280:969–974. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hollstein M, Sidransky D, Vogelstein B and
Harris CC: p53 mutations in human cancers. Science. 253:49–53.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Vogelstein B, Fearon ER, Hamilton SR, et
al: Genetic alterations during colorectal-tumor development. N Engl
J Med. 319:525–532. 1988. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Gleich LL: Gene therapy for head and neck
cancer. Laryngoscope. 110:708–726. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Hanahan D and Weinberg RA: Hallmarks of
cancer: the next generation. Cell. 144:646–674. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Parker JD, Hilton LK, Diener DR, et al:
Centrioles are freed from cilia by severing prior to mitosis.
Cytoskeleton (Hoboken). 67:425–430. 2010. View Article : Google Scholar
|
|
40
|
Christensen ST, Pedersen SF, Satir P,
Veland IR and Schneider L: Chapter 10 The primary cilium
coordinates signaling pathways in cell cycle control and migration
during development and tissue repair. Current Topics in
Developmental Biology. Bradley KY: 85. Academic Press; pp. 261–301.
2008, View Article : Google Scholar
|
|
41
|
Schneider L, Cammer M, Lehman J, et al:
Directional cell migration and chemotaxis in wound healing response
to PDGF‑AA are coordinated by the primary cilium in fibroblasts.
Cell Physiol Biochem. 25:279–292. 2010. View Article : Google Scholar :
|