Introduction
The local metabolism and synthesis of estrogens,
resulting from the interactions of various enzymes, is considered
to play a crucial role in the pathogenesis and development of
hormone-dependent carcinomas, but also in various organs and
systems, including female reproductive organs, bone, liver, central
nervous system and vascular system (1–3).
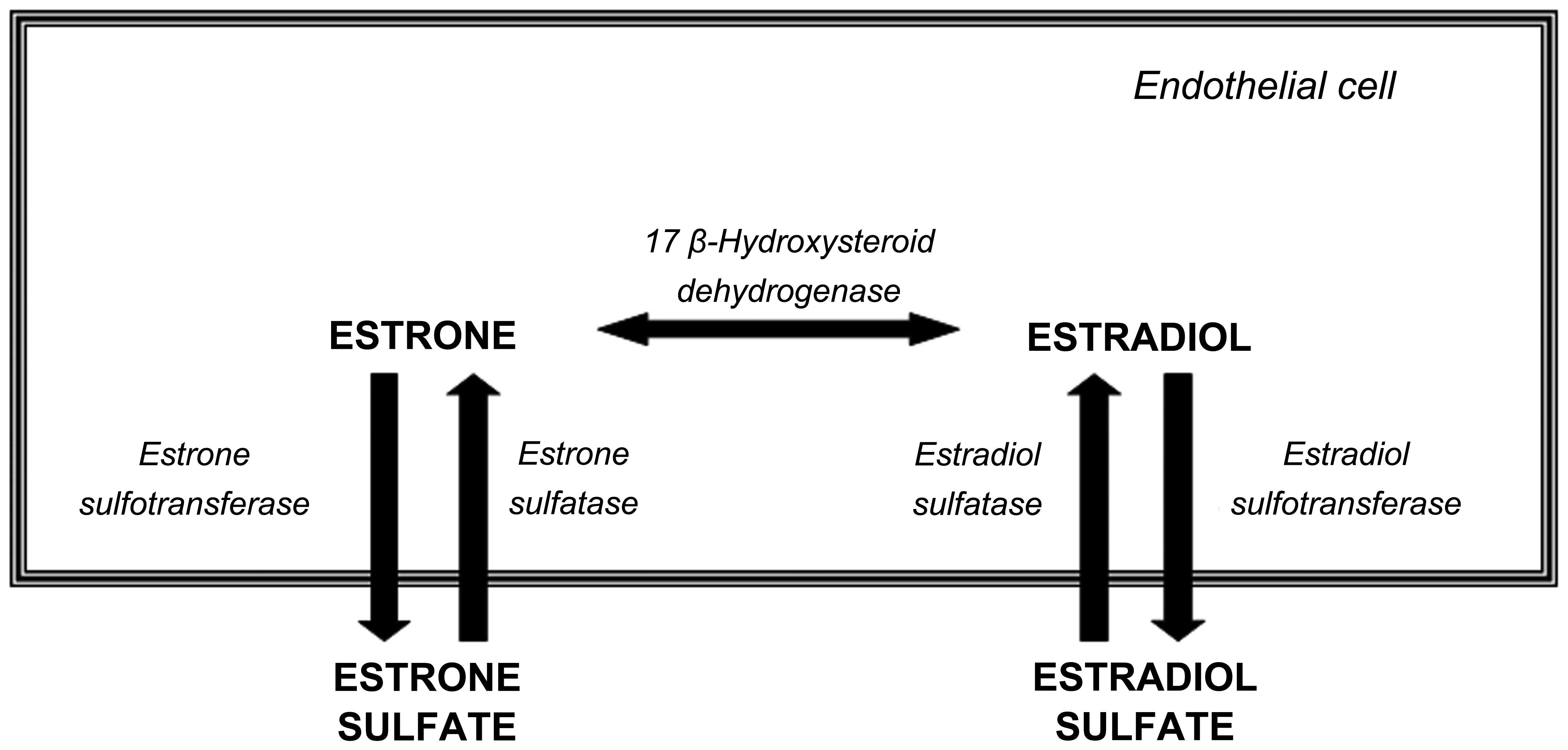
The major circulating form of plasma estrogen is estrone sulfate, a
biologically inactive form of estrogen, which acts as an estrogen
reserve, which can be formed from sulfated estrogens by steroid
sulfatase (STS), which is present in both normal and tumor tissues
(Fig. 1). The plasma
concentrations of estrogen sulfates are 50- to 15-fold higher than
those of unconjugated estrogens, such as estrone, estradiol and
estriol, and their half-life in plasma (10–12 h) is considerably
longer than that of estrone and estradiol (20–30 min) (4,5).
Estrogen sulfates are therefore thought to act as a reservoir for
the formation of active estrogens via the action of STS (6), which converts inactive estrogen
sulfates to estrone and 17β-estradiol. Estrone is then reduced to
the biologically active estrogen, 17β-estradiol, by
17β'-hydroxysteroid dehydrogenase type 1 (17β-HSD1) (7). STS is ubiquitously expressed in
mammalian tissues and target organs, including the liver,
endometrium, ovaries, bone, brain, prostate, white blood cells and
adipocytes, but it is particularly prevalent in the placenta and in
breast carcinomas (6,7). The potency of estrogenic hormones is
generally reduced during the metabolic process to inactive products
in target tissues, and this contributes to the modulation of the
overall biological actions of estrogens. Estrogen sulfotransferase
(EST) sulfonates estrogens to form biologically inactive estrogen
sulfates (8) and it is
distributed in a wide range of normal human tissues, such as the
kidneys, liver, gastrointestinal tract, vascular tissue and breasts
(Fig. 1). In breast carcinoma,
EST activity and expression tend to be decreased (9). Therefore, STS and EST are considered
to be involved in the regulation of in situ estrogen levels
in various human tissues and hormone-dependent tumors (6,10).
Melatonin, the main secretory product of the pineal
gland, is well known to reduce the growth and development of
estrogen-responsive breast cancers (11–14). Melatonin exerts its oncostatic
effects in hormone-dependent breast cancer by interfering at
different levels of the estrogen signaling pathways (15,16). One of the mechanisms through which
this occurs involves the regulation by melatonin of the expression
and activity of several enzymes (aromatase, STS, 17β-HSD1 and EST)
that are responsible for the local synthesis of estrogens; thus,
melatonin functions as a selective estrogen enzyme modulator
(17,18). Melatonin exerts its regulatory
actions on estrogen-producing enzymes in tumor cells, surrounding
fibroblasts and endothelial cells. In endothelial cells, melatonin
regulates the aromatase pathway, which transforms androgens into
estrogens (19–21). Melatonin reduces the activity and
expression of aromatase mainly by inducing a significant
downregulation in aromatase expression, specifically driven by
promoter I.7, the major promoter directing aromatase expression in
endothelial cells (19). In
addition, melatonin has been shown to exert anti-angiogenic effects
by reducing endothelial cell proliferation, invasion, migration and
tube formation, through a downregulatory effect on vascular
endothelial growth factor (VEGF) in endothelial cells (22,23). These results piqued our interest
into the possible effects of melatonin on other enzymes involved in
the local biotransformation of steroids in endothelial cells. Thus,
our objective in the present study, was to analyze the activity and
expression of EST and STS in human umbilical vein endothelial cells
(HUVECs) and to elucidate the possible modulatory effects and
mechanisms of melatonin.
Materials and methods
Cells and culture conditions
The HUVECs were purchased from the American Tissue
Culture Collection (ATCC; Rockville, MD, USA). They were maintained
as monolayer cultures in 75 cm2 plastic culture flasks
in Vascular Cell Basal Medium (VCBM) supplemented with Endothelial
Cell Growth kit-BBE (both from ATCC) which consisted of 2% fetal
bovine serum (FBS), 0.2% bovine brain extract, 5 ng/ml recombinant
human epidermal growth factor (rhEGF), 10 mM L-glutamine, 0.75 U/ml
heparin sulfate, 1 µg/ml hydrocortisone hemisuccinate, 50
µg/ml ascorbic acid, penicillin (20 U/ml) and streptomycin
(20 µg/ml) (Sigma-Aldrich, Madrid, Spain) at 37°C in a humid
atmosphere containing 5% CO2. To avoid genetic mutation
and low viability, HUVECs at no more than 6 passages were used in
the following experiments.
Measurement of STS activity
STS activity in the HUVECs was assayed by the
formation of estrone from a labeled substrate
([6,7-3H(N)]-estrone sulfate ammonium salt) (24). The HUVECs were seeded onto 75
cm2 plastic culture flasks in VCBM supplemented with 2%
FBS. When a homogenous monolayer of pre-confluent HUVECs was
established, the medium was aspirated and the cultured cells were
harvested and seeded onto 6-well plates (3×105
cells/well) in VCBM supplemented with 2% FBS. One day later, the
medium was replaced with fresh medium (1 ml/plate) containing 20 nM
[6,7-3H(N)]-estrone sulfate ammonium salt (NEN Life
Science Products, Boston, MA, USA) (57.3 Ci/mM) in the presence of
either 1 mM, 1 µM or 1 nM melatonin, or its diluent
(control; ethanol at a final concentration <0.001%). Following
20 h of incubation, the culture dishes were placed on ice for 15
min to condense any water vapour and 0.5 ml of the medium was
transferred to tubes containing 2.5 ml of toluene, vortexed for 45
sec and centrifuged at 1,000 × g for 10 min at 4°C. The resulting
organic phase was added to vials with scintillation cocktail
(National Diagnostics, Atlanta, GA, USA) and counted using a beta
counting system (Beckman LS60001C Scintillation Counter; Beckman
Instruments, Atlanta, GA, USA). The amount of radioactivity
measured in the [3H]-toluene was corrected by
subtracting the blank values from each sample, obtained by
incubating dishes containing medium but no cells with the tritiated
estrone. As previously described (18), the values were also corrected by
taking into account the fractional retention of tritium in medium
throughout the procedure of incubation and processing, utilizing
parallel dishes containing medium plus known amounts of
[3H] estrone (NEN Life Science Products). The fractional
retention of tritium in medium throughout the incubation and
processing of samples was always >92%.
Measurement of EST activity
EST activity in the HUVECs was evaluated by the
formation of E1-S from
[2,4,6,7-3H(N)]-estrone, as previously described
(25). The HUVECs were seeded
onto 75 cm2 plastic culture flasks in VCBM supplemented
with 2% FBS. When a homogenous monolayer of pre-confluent HUVECs
was reached, the medium was aspirated and the cultured cells were
harvested and seeded onto 6-well plates (3×105
cells/well) in VCBM supplemented with 2% FBS. One day later, the
medium was replaced with fresh medium (1 ml/plate) supplemented
with 2% dicyclohexylcarbodiimide (DCC), containing 2 µM
estrone including 10 nM [2,4,6,7-3H(N)]-estrone (NEN
Life Science Products) (100 Ci/mM), 7 mM MgCl2, 50 mM
Tris-HCl pH 7.4, phosphoadenosine phosphosulfate (PAPS) at a final
concentration of 20 µM, in the presence of either melatonin
(1 mM, 1 µM or 1 nM), or its diluent (control; ethanol at a
final concentration <0.001%). Following 48 h of incubation, the
medium was transferred to tubes containing a cold mix of 4 ml of
chloroform plus 0.375 ml of 0.25 M Tris-HCl pH 8.7, vortexed for 45
sec and centrifuged at 600 × g for 5 min at 4°C. The resulting
aqueous phase was added to vials with scintillation cocktail and
counted using a beta counting system. Values were corrected for
blanks and tritium recovery (>87%) was as described above.
Measurement of mRNA expression of STS and
EST
The mRNA expression of different enzymes was carried
out by reverse transcription-quantitative polymerase chain reaction
(RT-qPCR) of the HUVECs, after incubation of cells with either
melatonin or the vehicle for 4 h, as described in a previous study
of ours (39). Total cellular RNA
was purified using the NucleoSpin RNA II kit (Macherey-Nagel GmbH
& Co., Düren, Germany) following the manufacturer's
instructions. The integrity of the RNA was assessed by
electrophoresis in ethidium bromide-stained 1.2%
agarose-Tris-borate EDTA gels. The absorbance ratio A260/A280 nm
was >1.8. For cDNA synthesis, 1 µg of total RNA was
denaturated at 65°C for 10 min and reverse transcribed for 50 min
at 45°C using the cDNA synthesis kit (Bioline, London, UK) in a
final volume of 20 µl in the presence of 500 ng of
oligo(dT)12–18 primer. qPCR (MX3000; Stratagene, La Jolla, CA, USA)
was performed using Brilliant SYBR-Green PCR Master Mix
(Stratagene) following the manufacturer's instructions. The pairs
of human oligonucle-otides (Sigma Genosys Ltd., Cambridge, UK) used
as primers are indicated in Table
I. S14 mRNA expression was used as a control. PCRs were
performed for 40 cycles for quantitative analysis using the
annealing temperature indicated in Table I for 60 sec, the extension being
carried out at 72°C for 45 sec and the denaturation at 95°C for 50
sec. Each product was electrophoresed on ethidium bromide-stained
2% agarose-Tris-borate gels to corroborate that the product
amplified corresponded to the adequate length. Finally, the mRNA
expression levels of target genes were normalized to S14 mRNA
expression. Each reaction was run in triplicate. Melting curve
analysis was performed using dissociation curves to verify that
only a single product with no primer-dimers was amplified.
 | Table IPrimers used for the amplification of
mRNA transcripts of STS, EST and S14 (control). |
Table I
Primers used for the amplification of
mRNA transcripts of STS, EST and S14 (control).
| mRNA | Sequence | bp | [ ] nM | T |
|---|
| STS | F:
5′-TCCGTTCCTGCTTGTCTTGTC-3′ | 197 | 200 | 57°C |
| R:
5′-CCTGGTCCGATGTGAAGTAGATG-3′ | | 200 | 57°C |
| EST | F:
5′-AGACTCATTTGCCACCTGAACTTC-3 | 133 | 200 | 59°C |
| R:
5′-GGATGACCAGCCACCATTAGAAAG-3′ | | 100 | 59°C |
| S14 | F:
5′-TCACCGCCCTACACATCAAAC-3′ | 159 | 100 | |
| R:
5′-TCCTGCGAGTGCTGTCAGAG-3′ | | 100 | |
Statistical analysis
The data are expressed as the means ± standard
errors of the mean (SEM) of 6 independent experiments. Statistical
differences between groups were analyzed by one way analysis of
variance (ANOVA), followed by the Student-Newman-Keuls test.
Results were considered statistically significant difference at a
value of p<0.05.
Results
Effects of melatonin on STS activity and
expression
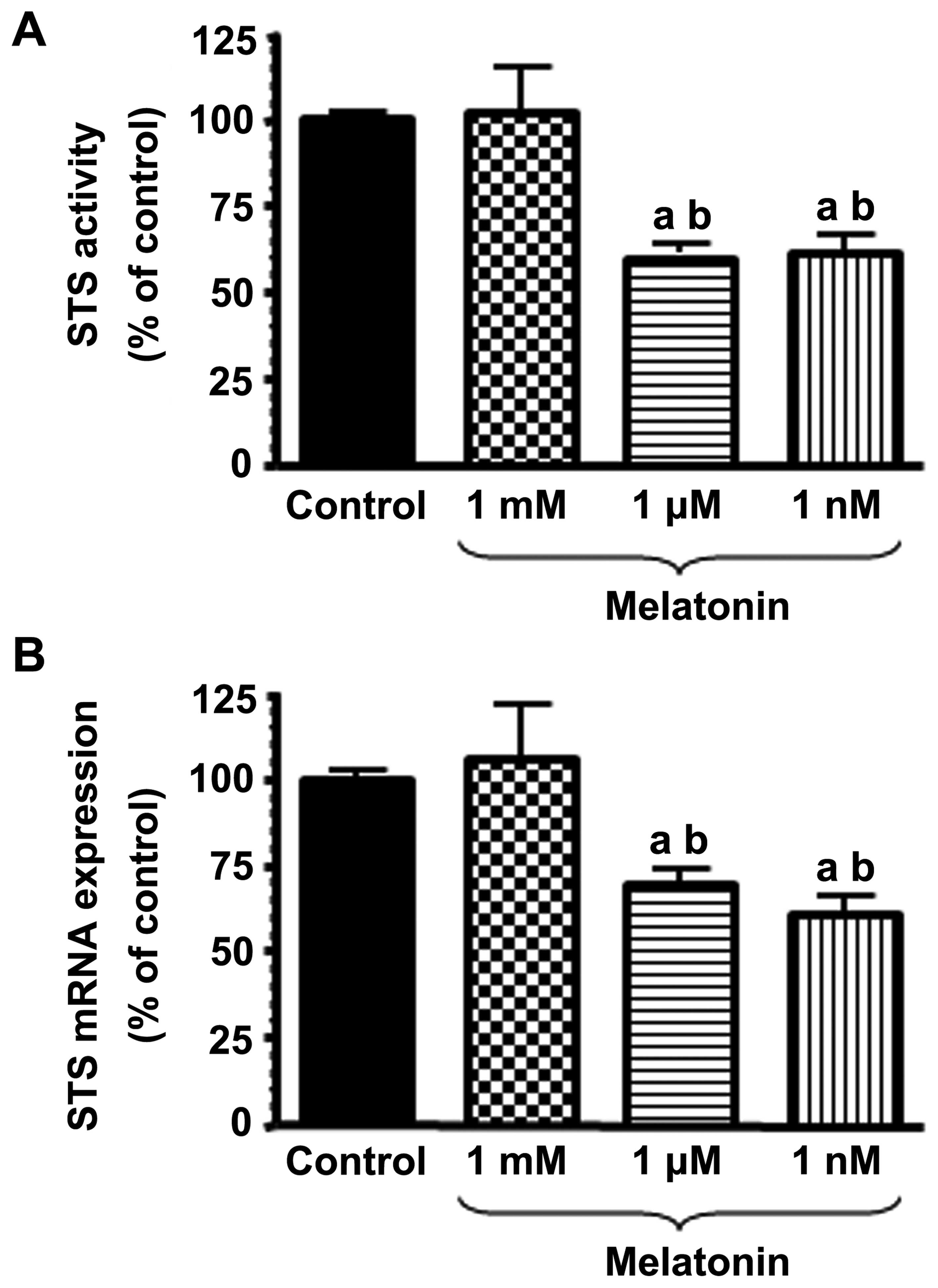
We first examined the effects of various
concentrations of melatonin on the activity and mRNA expression of
STS. Physiological concentrations of melatonin (1 nM) induced a
significant (p<0.001) inhibition of STS activity in the HUVECs
(Fig. 2A). The concentrations of
melatonin in the micromolar range (1 µM) were also effective
at reducing (p<0.001) the conversion of estrone sulphate to
estrone.
With the aim of determining whether the inhibitory
effects of melatonin on STS activity are due to the downregulation
of STS expression, we incubated the HUVECs with either melatonin (1
mM, 1 µM or 1 nM) or the vehicle [the diluent (control),
ethanol at a final concentration <0.001%] for 4 h, and total RNA
was isolated to perform RT-qPCR with specific primers for human
STS. As a control, the same samples were subjected to RT-qPCR with
specific primers for the housekeeping gene, S14, a ribosomal
protein component of the 40S subunit. Treatment of the HUVECs with
physiological concentrations of melatonin (1 nM) led to a
significant and potent inhibition (40%) of STS mRNA expression
compared to the controls (Fig.
2B). The supraphysiological concentrations of melatonin (1
µM) were also effective at reducing (p<0.001) STS mRNA
expression.
Effects of melatonin on EST activity and
expression
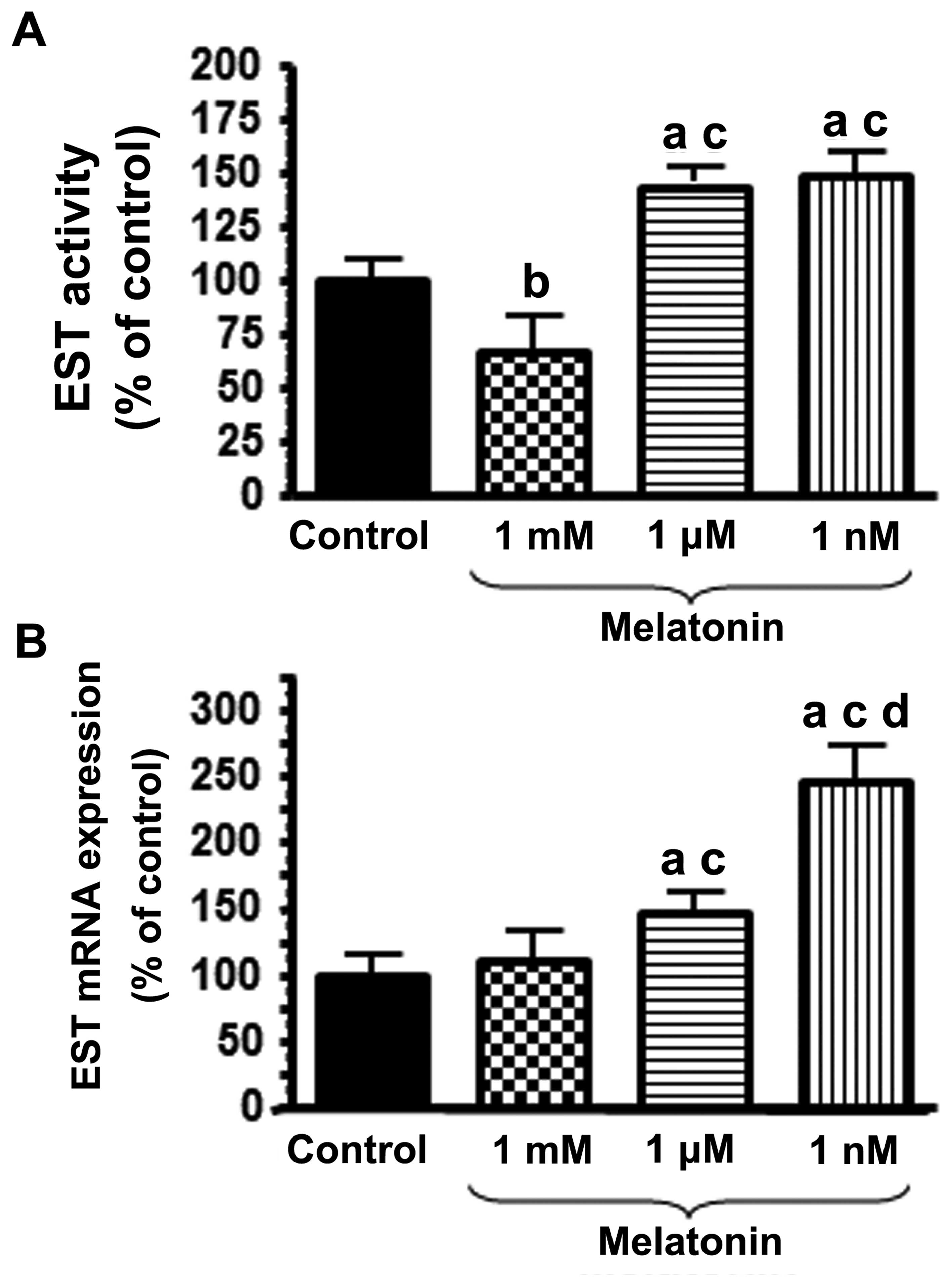
In a second set of experiments, the effects of
melatonin on the activity and mRNA expression of EST were examined.
The activity of EST was significantly stimulated by treatment of
the HUVECs with physiological concentrations (1 nM) of melatonin
(Fig. 3A). When the HUVECs were
incubated for 4 h with 1 nM melatonin, the mRNA expression of EST
was significantly increased (p<0.001) (Fig. 3B). Higher concentrations of
melatonin (1 µM) also induced an increase in the mRNA
expression of EST, although to a lesser extent than that triggered
by treatment with 1 nM melatonin (Fig. 3B).
Discussion
Estrogens are potent steroid hormones with
pleiotropic effects potentially causing diverse modulatory effects
on vascular functions and the endothelial dysfunction that is
associated with an increased risk of cancer and cardiovascular
disease (26–28). Estrogens synthesized in
peritumoral endothelial cells also play a role in the growth and
progression of estrogen-dependent tumors, such as breast tumors
(4). Estrogens are secreted
primarily by the ovaries, the placenta and the testes, and they are
also produced by peripheral steroidogenic conversion. Two major
pathways are implicated in providing peripheral sources of estrogen
in human peripheral tissues. One is the aromatase pathway which
transforms androgens into estrogens (29,30), and the other is the sulfatase
pathway which converts estrogen sulfates into estrogens by the
enzyme, STS (5). During the past
decades, research has been directed mainly towards the development
of aromatase inhibitors (31),
and as a result, potent, well-tolerated and highly selective
aromatase inhibitors are now available for use in clinical practice
(32). However, there is a
growing awareness of the role that STS may play in regulating the
formation of estrogenic steroids since the estrone sulfatase
pathway is the major route of estrogen formation in breast tumors
(6,33). Estrone sulfate is quantitatively
the most important precursor of estradiol and STS activity is
40–500-fold greater than aromatase activity in breast cancer
tissues. Whereas STS decreases estrogen sulfate levels and
increases active estrogen levels, EST sulfonates estrogens to
biologically inactive estrogen sulfates and increases the level of
estrogen sulfates (4,9,34).
Melatonin is well known for its oncostatic
properties (11,12,14,16). This indoleamine exerts its
oncostatic effects on estrogen-dependent breast tumors mainly
through two mechanisms: interfering with the estrogen signaling
pathways at the estrogen receptor level (16,35,36) and regulating the activity and
expression of the enzymes involved in local estrogen biosynthesis
in tumor cells and peritumoral fibroblasts (16,20,21,37,38).
To date, it has not been clearly determined whether
STS and EST activity and expression are present in HUVECs. The
present study firstly demonstrated that STS and EST are expressed
in HUVECs, and are able to modulate the concentration of estrogen
sulfates, and therefore, estrogen synthesis in endothelial cells.
Melatonin, at physiological (1 nM) concentrations, induced a
significant inhibition of STS activity in the HUVECs and this
inhibitory effect was due to the downregulation of STS expression.
To the best of our knowledge, this is the first time that the
effects of melatonin on STS activity and expression in endothelial
cells have been established. In relation to the modulatory effects
of melatonin on STS, it is known that melatonin inhibits STS
activity and expression in vitro in human breast cancer
cells (18) and glioma cells
(39). This modulatory effect of
melatonin on STS has also been described in vivo, in rats
bearing 7,12-dimethylbenz[a]anthracene (DMBA)-induced mammary
tumors (40). The inhibition of
STS activity and expression in breast cancer with high doses of
synthetic antiestrogens, such as tamoxifen, has been also described
(41).
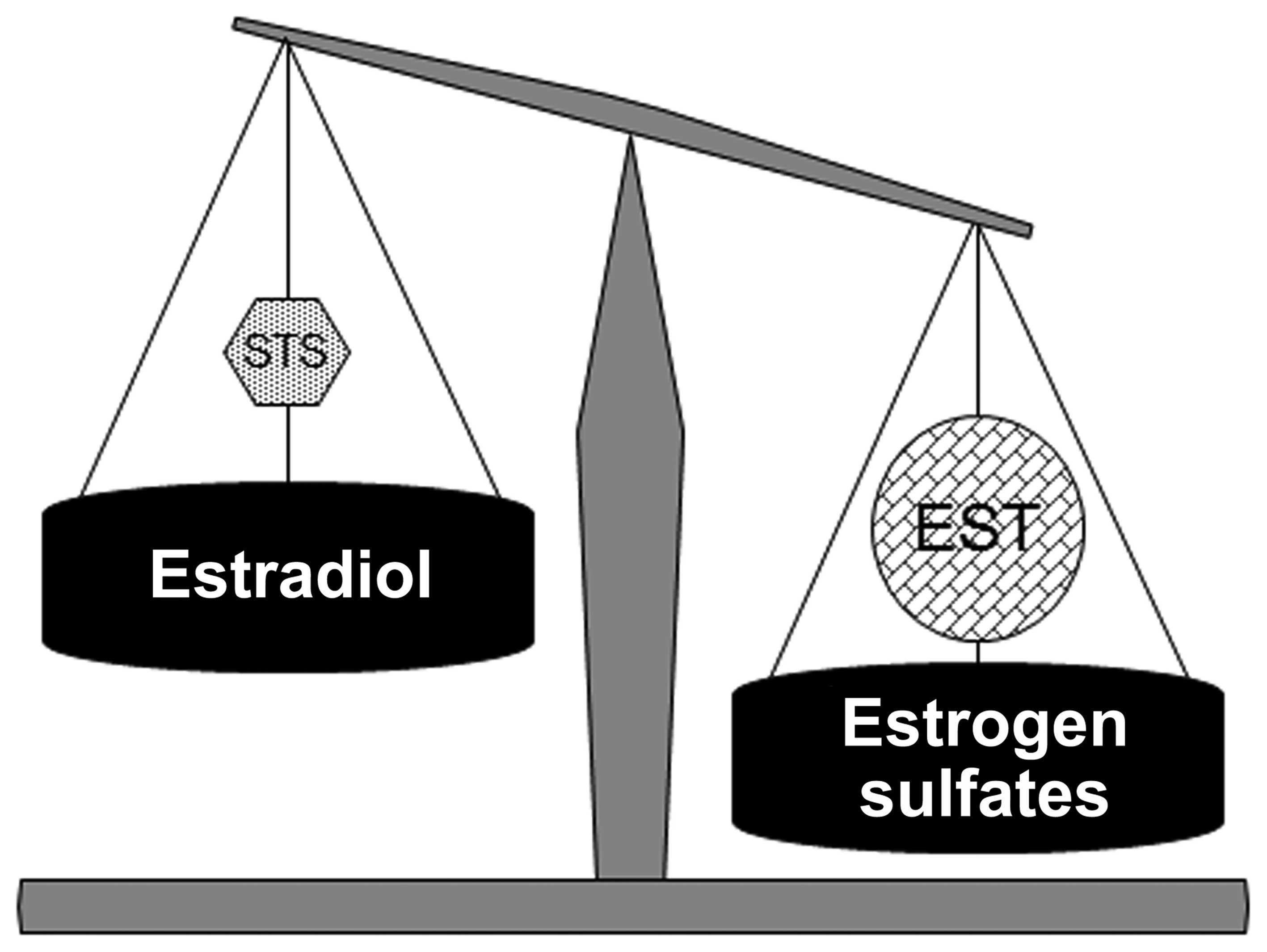
Estrogen sulfate concentrations are dependent on its
inactivation by STS and its production by EST (balance of STS/EST).
Recently, it has been described that EST is highly expressed in
human vascular endothelial and smooth muscle cells and the
modulation of its activity and expression regulates the levels of
active estrogens in these cells, thereby affecting the physiology
and pathophysiology of vascular walls (42,43). In this study, physiological
concentrations of melatonin (1 nM) significantly increased EST
activity and expression in HUVECs. This upregulation of EST induced
by melatonin has also been described in human breast cancer cells
(9,18,20). This indolamine, at physiological
concentrations in human breast cancer cells, increased EST activity
by almost 2-fold and EST expression up to 3-fold (9,18,20). These results concur with the
findings of other previous on estrogen-responsive human breast
cancer cells. Melatonin was shown to exert a direct
concentration-dependent anti-proliferative effect and only
melatonin concentrations closer to 1 nM (concentrations similar to
those found in the serum of most mammals during the nocturnal
period) were effective at decreasing cell proliferation, whereas
supra- or subphysiological concentrations lacked these
antiproliferative effects (11,12,14).
STS and EST are involved in the regulation of in
situ estrogen sulfate levels in HUVECs. The balance between
these two enzymes determines the concentration of estrogen sulfates
and consequently, the concentration of active estrogens in
endothelial cells (4,6). In normal breast tissue, STS activity
and expression are very weak whereas, the activity and expression
of EST are very high. The equilibrium between the two enzymes is
displaced towards the accumulation of estrogen sulfates, the major
precursor of local estrogen production. However, in breast
carcinoma tissue the formation of the active 17β-estradiol
predominates, as STS tends to be overexpressed, whereas the
expression of EST is frequently decreased (4,9).
In HUVECs, since melatonin decreases the activity and expression of
STS, which transforms the estrone sulfates to estrogens, and
increases the activity and expression of EST, which inactivates
estrogens, it results in the modification of the dynamic
steady-state equilibrium of estrogen sulfates by increasing
estrogen sulfate levels and decreasing active estrogen levels
(Fig. 4). Melatonin regulates the
activity and expression of the enzymes involved in the local
synthesis of estrogens in endothelial cells in a similar manner to
the activity and expression of enzymes in normal mammary normal
tissue.
In conclusion, the findings presented herein point
to a role for melatonin in mediating the synthesis and
transformation of biologically active estrogens in HUVECs by
inhibiting the mRNA expression and activity of the enzyme involved
in estradiol formation (STS) and by stimulating the mRNA expression
and activity of the enzyme responsible for the inactivation of
estradiol and formation of the biologically inactive estrogen
sulfates (EST). Melatonin decreased STS and increased EST levels,
and as a result, modified the dynamic steady-state equilibrium of
estrogen sulfates through increased estrogen sulfate levels and
decreased active estrogen levels. Thus, melatonin may modulate the
physiology and pathophysiology of vascular tissue.
Acknowledgments
This study was supported by grants from the Spanish
Science Technology and Innovation Ministry (SAF2010-19579 and
SAF2013-42012-P) and a grant from the Instituto de Investigación
Valdecilla (IDIVAL) (APG/12).
References
|
1
|
Germain D: Estrogen carcinogenesis in
breast cancer. Endocrinol Metab Clin North Am. 40:473–484. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Pelekanou V and Leclercq G: Recent
insights into the effect of natural and environmental estrogens on
mammary development and carcinogenesis. Int J Dev Biol. 55:869–878.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Russo J and Russo IH: The role of estrogen
in the initiation of breast cancer. J Steroid Biochem Mol Biol.
102:89–96. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Pasqualini JR and Chetrite GS: Recent
insight on the control of enzymes involved in estrogen formation
and transformation in human breast cancer. J Steroid Biochem Mol
Biol. 93:221–236. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stanway SJ, Delavault P, Purohit A, Woo
LW, Thurieau C, Potter BV and Reed MJ: Steroid sulfatase: a new
target for the endocrine therapy of breast cancer. Oncologist.
12:370–374. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Pasqualini JR: The selective estrogen
enzyme modulators in breast cancer: a review. Biochim Biophys Acta.
1654:123–143. 2004.PubMed/NCBI
|
|
7
|
Suzuki T, Miki Y, Nakamura Y, Moriya T,
Ito K, Ohuchi N and Sasano H: Sex steroid-producing enzymes in
human breast cancer. Endocr Relat Cancer. 12:701–720. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Suzuki T, Miki Y, Nakata T, Shiotsu Y,
Akinaga S, Inoue K, Ishida T, Kimura M, Moriya T and Sasano H:
Steroid sulfatase and estrogen sulfotransferase in normal human
tissue and breast carcinoma. J Steroid Biochem Mol Biol.
86:449–454. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Cos S, González A, Álvarez-García V,
Alonso-González C and Martínez-Campa C: Melatonin and breast
cancer: selective estrogen enzyme modulator actions. Advances in
Cancer Drug Targets. Atta-ur-Rahman: 1. Bentham Science Publishers;
Sharjah, UAE: pp. 207–237. 2013
|
|
10
|
Sasano H, Suzuki T, Nakata T and Moriya T:
New development in intracrinology of breast carcinoma. Breast
Cancer. 13:129–136. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Cos S and Sánchez-Barceló EJ: Melatonin
and mammary pathological growth. Front Neuroendocrinol. 21:133–170.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Cos S and Sánchez-Barceló EJ: Melatonin,
experimental basis for a possible application in breast cancer
prevention and treatment. Histol Histopathol. 15:637–647.
2000.PubMed/NCBI
|
|
13
|
Hill SM and Blask DE: Effects of the
pineal hormone melatonin on the proliferation and morphological
characteristics of human breast cancer cells (MCF-7) in culture.
Cancer Res. 48:6121–6126. 1988.PubMed/NCBI
|
|
14
|
Blask DE, Sauer LA and Dauchy RT:
Melatonin as a chronobiotic/anticancer agent: Cellular,
biochemical, and molecular mechanisms of action and their
implications for circadian-based cancer therapy. Curr Top Med Chem.
2:113–132. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sánchez-Barceló EJ, Cos S, Fernández R and
Mediavilla MD: Melatonin and mammary cancer: a short review. Endocr
Relat Cancer. 10:153–159. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Cos S, González A, Martínez-Campa C,
Mediavilla MD, Alonso-González C and Sánchez-Barceló EJ:
Estrogen-signaling pathway: a link between breast cancer and
melatonin oncostatic actions. Cancer Detect Prev. 30:118–128. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Cos S, González A, Martínez-Campa C,
Mediavilla MD, Alonso-González C and Sánchez-Barceló EJ: Melatonin
as a selective estrogen enzyme modulator. Curr Cancer Drug Targets.
8:691–702. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
González A, Cos S, Martínez-Campa C,
Alonso-Gonzalez C, Sánchez-Mateos S, Mediavilla MD and
Sánchez-Barcelo EJ: Selective estrogen enzyme modulator actions of
melatonin in human breast cancer cells. J Pineal Res. 45:86–92.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Álvarez-García V, González A,
Martínez-Campa C, Alonso-González C and Cos S: Melatonin modulates
aromatase activity and expression in endothelial cells. Oncol Rep.
29:2058–2064. 2013.PubMed/NCBI
|
|
20
|
Cos S, Martínez-Campa C, González A,
Álvarez-García V, Alonso-González C, Mediavilla MD and
Sánchez-Barceló EJ: Melatonin and aromatase in breast cancer. Clin
Cancer Drug. 1:54–64. 2014. View Article : Google Scholar
|
|
21
|
Cos S, Alvarez-García V, González A,
Alonso-González C and Martínez-Campa C: Melatonin modulation of
crosstalk among malignant epithelial, endothelial and adipose cells
in breast cancer (Review). Oncol Lett. 8:487–492. 2014.PubMed/NCBI
|
|
22
|
Alvarez-García V, González A,
Alonso-González C, Martínez-Campa C and Cos S: Regulation of
vascular endo-thelial growth factor by melatonin in human breast
cancer cells. J Pineal Res. 54:373–380. 2013.
|
|
23
|
Alvarez-García V, González A,
Alonso-González C, Martínez-Campa C and Cos S: Antiangiogenic
effects of melatonin in endothelial cell cultures. Microvasc Res.
87:25–33. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Duncan L, Purohit A, Howarth NM, Potter BV
and Reed MJ: Inhibition of estrone sulfatase activity by
estrone-3-methylthio-phosphonate: A potential therapeutic agent in
breast cancer. Cancer Res. 53:298–303. 1993.PubMed/NCBI
|
|
25
|
Falany CN, Krasnykh V and Falany JL:
Bacterial expression and characterization of a cDNA for human liver
estrogen sulfotransferase. J Steroid Biochem Mol Biol. 52:529–539.
1995. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Barnabas O, Wang H and Gao XM: Role of
estrogen in angiogenesis in cardiovascular diseases. J Geriatr
Cardiol. 10:377–382. 2013.
|
|
27
|
Chakrabarti S, Morton JS and Davidge ST:
Mechanisms of estrogen effects on the endothelium: An overview. Can
J Cardiol. 30:705–712. 2014. View Article : Google Scholar
|
|
28
|
Khalil RA: Estrogen, vascular estrogen
receptor and hormone therapy in postmenopausal vascular disease.
Biochem Pharmacol. 86:1627–1642. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Bulun SE, Lin Z, Imir G, Amin S, Demura M,
Yilmaz B, Martin R, Utsunomiya H, Thung S, Gurates B, et al:
Regulation of aromatase expression in estrogen-responsive breast
and uterine disease: from bench to treatment. Pharmacol Rev.
57:359–383. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Bulun SE, Chen D, Lu M, Zhao H, Cheng Y,
Demura M, Yilmaz B, Martin R, Utsunomiya H, Thung S, et al:
Aromatase excess in cancers of breast, endometrium and ovary. J
Steroid Biochem Mol Biol. 106:81–96. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Chumsri S and Brodie A: Aromatase
inhibitors and breast cancer. Horm Mol Biol Clin Investig.
9:119–126. 2012.PubMed/NCBI
|
|
32
|
Chumsri S: Clinical utilities of aromatase
inhibitors in breast cancer. Int J Womens Health. 7:493–499. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Shields-Botella J, Bonnet P, Duc I,
Duranti E, Meschi S, Cardinali S, Prouheze P, Chaigneau AM, Duranti
V, Gribaudo S, et al: In vitro and in vivo models for the
evaluation of new inhibitors of human steroid sulfatase, devoid of
residual estrogenic activity. J Steroid Biochem Mol Biol.
84:327–335. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Santner SJ, Ohlsson-Wilhelm B and Santen
RJ: Estrone sulfate promotes human breast cancer cell replication
and nuclear uptake of estradiol in MCF-7 cell cultures. Int J
Cancer. 54:119–124. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hill SM, Spriggs LL, Simon MA, Muraoka H
and Blask DE: The growth inhibitory action of melatonin on human
breast cancer cells is linked to the estrogen response system.
Cancer Lett. 64:249–256. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Molis TM, Spriggs LL and Hill SM:
Modulation of estrogen receptor mRNA expression by melatonin in
MCF-7 human breast cancer cells. Mol Endocrinol. 8:1681–1690.
1994.PubMed/NCBI
|
|
37
|
Cos S, Martínez- Campa C, Mediavilla MD
and Sánchez-Barceló EJ: Melatonin modulates aromatase activity in
MCF-7 human breast cancer cells. J Pineal Res. 38:136–142. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Cos S, González A, Güezmes A, Mediavilla
MD, Martínez-Campa C, Alonso-González C and Sánchez-Barceló EJ:
Melatonin inhibits the growth of DMBA-induced mammary tumors by
decreasing the local biosynthesis of estrogens through the
modulation of aromatase activity. Int J Cancer. 118:274–278. 2006.
View Article : Google Scholar
|
|
39
|
González A, Martínez-Campa C, Mediavilla
MD, Alonso-González C, Alvarez-García V, Sánchez-Barceló EJ and Cos
S: Inhibitory effects of melatonin on sulfatase and
17β-hydroxysteroid dehydrogenase activity and expression in glioma
cells. Oncol Rep. 23:1173–1178. 2010.
|
|
40
|
González A, Alvarez-García V,
Martínez-Campa C, Mediavilla MD, Alonso-González C, Sánchez-Barceló
EJ and Cos S: In vivo inhibition of the estrogen sulfatase enzyme
and growth of DMBA-induced mammary tumors by melatonin. Curr Cancer
Drug Targets. 10:279–286. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Santner SJ and Santen RJ: Inhibition of
estrone sulfatase and 17β-hydroxysteroid dehydrogenase by
antiestrogens. J Steroid Biochem Mol Biol. 45:383–390. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Li Y, Xu Y, Li X, Qin Y and Hu R: Effects
of PPAR-α agonist and IGF-1 on estrogen sulfotransferase in human
vascular endo-thelial and smooth muscle cells. Mol Med Rep.
8:133–139. 2013.PubMed/NCBI
|
|
43
|
Xu Y, Yang X, Wang Z, Li M, Ning Y, Chen
S, Yin L and Li X: Estrogen sulfotransferase (SULT1E1) regulates
inflammatory response and lipid metabolism of human endothelial
cells via PPARγ. Mol Cell Endocrinol. 369:140–149. 2013. View Article : Google Scholar : PubMed/NCBI
|