Introduction
Ischemic heart disease, particularly acute
myocardial infarction, is a leading cause of morbidity and
mortality worldwide (1).
Immediate restoration of the interrupted blood supply to the heart
by thrombolysis or percutaneous transluminal coronary angioplasty
is the main mode of treatment (2–4).
However, abrupt reperfusion of ischemic myocardium can exacerbate
the damage to the ischemic tissue. This phenomenon has been termed
myocardial ischemia/reperfusion injury (MI/RI) (5,6).
MI/RI can lead to irreversible cell damage, ventricular
dysfunction, and heart failure, which can cause sudden cardiac
death (7).
Although the pathogenesis of reperfusion-induced
myocardial injury is multifactorial (8,9),
apoptosis plays an important role in reperfusion injury (10,11). Apoptosis is a highly regulated,
evolutionarily conserved and energy-dependent process. It is
triggered by a specific cascade of signaling pathways that
ultimately lead to cell death and can result in severe organ injury
(12,13). Cell biology studies have
demonstrated that blocking apoptosis may prevent the loss of
contractile cells, minimize ischemia/reperfusion-induced cardiac
injury, and, thus, retard or prevent the occurrence of heart
failure (14). Therefore, the
exploration of anti-apoptotic agents is a viable approach to the
optimization of reperfusion strategies.
Aralia taibaiensis Z. Z. Wang & H. C.
Zheng, a member of the Araliaceae family, is a widely distributed
species in the Qinba Mountains of Western China (15). The bark and root cortex are widely
used in traditional folk medicine for the treatment of diabetes
mellitus, hepatitis and stomach ulcers (16). In previous investigations, it was
identified that total saponins, particularly triterpenoid saponins,
extracted from Aralia taibaiensis (hereafter referred to as
sAT) are the main pharmaco logically active components of Aralia
taibaiensis (15,17). In addition, sAT had potent
antidiabetic activity (16,18,19) and anti-aging effect (18), which are associated with cardiac
disease, particularly ischemic disease. Thus sAT has potential
protective effects against MI/RI. However, the anti-MI/RI
properties of sAT have yet to be examined and the possible
molecular mechanisms involved to be determined.
The present study was therefore undertaken to
investigate the anti-MI/RI activities of sAT and to elucidate the
mechanisms underlying these effects in rats. The results provided
important insights into the effects of sAT in cardiac disease.
Materials and methods
Materials
Dulbecco's modified Eagle's medium (DMEM) and fetal
bovine serum (FBS) were purchased from Gibco-BRL (Grand Island, NY,
USA). The kits used for the determination of serum lactate
dehydrogenase (LDH) and creatine kinase isoenzyme-MB (CK-MB)
content were obtained from Jiancheng Bioengineering Institute
(Nanjing, China). MTT
[3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide], TTC
(2,3,5-triphenyltetrazolium chloride), Evans blue, and compound C
[AMP-activated protein kinase (AMPK) inhibitor] were purchased from
Sigma-Aldrich (St. Louis, MO, USA). The primary antibodies were as
follows: anti-AMPKα (Thr172; #2532), anti-phosphorylated AMPKα
(p-AMPKα) (Thr172; #2531), anti-caspase-3 (#9662), anti-Bcl-2
associated X protein (Bax; #2772), anti-cytochrome c
(#11940), anti-acetyl CoA carboxylase (ACC; #3676), anti-p-ACC
(#11818), β-actin (#4970), and anti-Bcl-2 (#2870; all from Cell
Signaling Technologies, Beverly, MA, USA). All these are rabbit
monoclonal antibodies. The secondary antibody (anti-rabbit IgG-B;
sc-53804) was purchased from Santa Cruz Biotechnology (Santa Cruz,
CA, USA). The caspase-3 assay kit was purchased from Chemicon
International, Inc. (Temecula, CA, USA). The fluorescent kit for
Hoechst 33258 was obtained from Roche Diagnostics (Mannheim,
Germany).
Plant materials
The root bark of Aralia taibaiensis Z. Z.
Wang & H. C. Zheng was collected at Mountain Taibai, Shaanxi,
China, in September 2008, and botanically identified by Dr Haifeng
Tang (Department of Pharmacy, Xijing Hospital, and Fourth Military
Medical University, Shaanxi, China). A voucher specimen
(FMMUDP-Voucher No. SAP012) was deposited in the Herbarium of the
Department of Pharmacy, Xijing Hospital, and the Fourth Military
Medical University, China.
Preparation of total saponins
A crude extract (total saponins) of Aralia
taibaiensis (sAT) was prepared as previously described, with
slight modifications (16). Dry,
powdered root bark (20 g) was extracted three times with 80% (v/v)
ethanol (herb:ethanol, 1:10, w/v) under reflux (80°C) for 60 min.
The alcohol extract was concentrated, suspended in distilled water,
and then partitioned successively with chloroform (ratio 1:3, v/v)
and n-butanol saturated with water (ratio 1:3, v/v, three times).
The n-butanol extracts were combined and evaporated using a rotary
evaporator at 60°C to produce a powdered residue. The yield was
0.78% (w/w). Oleanolic acid was used as a reference standard, and
the total saponins content was expressed as oleanolic acid
equivalents (i.e., 459.30 µg oleanolic acid equivalents/ mg
extract). The content of total saponins in sAT was >90%.
Animal treatment schedule
sAT was dissolved in physiological saline (0.9%
NaCl) prior to use. After a 1-week-long adaptation period, the
animals were randomized into five experimental groups of eight
animals each as follows: sham-operated group + saline, MI/RI group
+ saline, MI/RI + sAT (60 mg/kg/day, orally), MI/RI + sAT (120
mg/kg/day, orally), MI/RI + sAT (240 mg/kg/day, orally).
Pretreatment with sAT or saline was initiated 7 days prior to the
operation and administered once a day for 7 days.
Animal preparation for MI/RI
Experimental procedures for these animals were
conducted according to the National Institutes of Health Guidelines
for the Use of Laboratory Animals and were approved by the
Institutional Animal Care and Use Committee of the Fourth Military
Medical University. Male Sprague-Dawley rats (250±30 g; 10±0.5
weeks) were anesthetized with 10% chloral hydrate (0.35 ml/100 g
body weight) and ventilated using a positive-pressure respirator
for small animals (HX-100E, Chengdu Technology & Market Co.,
Chengdu, China). Electrodes were placed subcutaneously and
connected to an electrocardiograph (BL-420S; Taimeng Science
Technology, Ltd., Chengdu, China). During surgery, body temperature
was measured and maintained at 37°C by placing the rats on a
heating pad. After left thoracotomy, the left anterior descending
coronary artery was occluded with a 6-0 silk suture. After
occlusion for 30 min, the suture was loosened, and the myocardium
was reperfused for 3 h. Sham-operated rats underwent identical
surgery, but the suture was not tightened around the coronary
artery. ST-segment elevation in experimental animals was assessed
as a measure of myocardial ischemia as previously described
(20).
After 3 h of reperfusion, blood samples were
collected from the abdominal aorta to measure serum LDH and CK-MB
levels by using an enzyme-linked immunosorbent assay (Nanjing
Jiancheng Bioengineering Institute, Nanjing, China). The assay was
performed according to the manufacturer's instructions. Following
anesthesia, the rats were sacrificed by exsanguination (blood was
drawn from the abdominal aorta) and the hearts were harvested for
double-staining.
Measurement of myocardial infarct
size
Myocardial infarct size was measured using a
double-staining technique with 3% Evans blue and 4% TTC adapted as
previously described (21). After
6 h of reperfusion, the coronary artery was again occluded. To map
the areas at risk for ischemia, 3% Evans blue solution was infused
through the left jugular vein. The heart was quickly excised and
frozen at −20°C, and the ventricles were then cross-sectioned into
five sections. The sections were counterstained with 4% TTC (in
phosphate buffer, pH 7.8) for 15 min at 37°C and photographed after
overnight storage in 4% paraformaldehyde. The infarct size was
presented as the left ventricular infarct area as a percentage of
the ischemic area at risk.
TUNEL staining for apoptosis in vivo
To examine cardiac myocyte apoptosis, samples of
tissue from the ischemic zones were fixed in 4% paraformaldehyde,
embedded in paraffin, and cut into 5-mm transverse sections. The
terminal deoxynucleotidyl transferase-mediated dUTP nick
end-labeling (TUNEL) assay was carried out using an apoptosis
detection kit (Roche Applied Science, Mannheim, Germany) according
to the manufacturer's instructions. 4′,6-Diamidino-2-phenylindole
(DAPI) was used as a counterstain. The apoptotic cells were
analyzed by laser-scanning confocal microscopy (SP5-FCS; Leica
Microsystems, Wetzlar, Germany). The number of TUNEL-positive
nuclei was calculated by ImageJ software (NIH, Bethesda, MD, USA).
Quantitative analysis was carried out using the following formula:
percentage of TUNEL-positive cells = TUNEL-positive cells/total
cells.
H9c2 cell culture and treatment
Rat H9c2 cardiomyocytes were obtained from the
American Type Culture Collection (ATCC; Manassas, VA, USA). The
cells were cultured in DMEM with 10% fetal bovine serum (FBS), 100
U/ml penicillin, and 100 mg/ml streptomycin and maintained in a
humidified atmosphere of 5% CO2 at 37°C. The medium was
replaced every 2–3 days, and the cells were subcultured at 80–90%
confluence by detaching with 0.05%
trypsin-ethylenediaminetetraacetic acid and reseeding.
Anoxia/reoxygenation (A/R) injury
model
To mimic ischemic injury in vitro, ischemia
and reperfusion were performed in rat H9c2 cells based on a
previously described method (22). At 80% confluence, the cells were
incubated in an 'ischemic buffer' for 2 h at 37°C in a hypoxic
chamber (previously bubbled with 95% N2 and 5%
CO2). The buffer contained 137 mM NaCl, 12 mM KCl, 0.49
mM MgCl2, 0.9 mM CaCl2·2H2O, 4 mM
HEPES, and 20 mM sodium lactate (pH 6.2). For the A/R studies, the
medium was replaced by maintenance medium (DMEM with 2% FBS) during
the re-oxygenation period.
Cell viability
Cell viability was determined colorimetrically using
the MTT assay. Cells at the exponential phase were seeded at
1×104 cells/well in 96-well plates. After different
treatments, 20 µl of 5 mg/ml MTT solution was added to each
well (0.5 mg/ml final concentration), and the plates were incubated
for 4 h at 37°C. The supernatant was removed, the formazan crystals
were dissolved in 150 µl DMSO, and the optical density at
490 nm was read on a microplate reader (Tecan, Mannedorf,
Switzerland). The cell survival ratio was expressed as a percentage
of the control.
Determination of LDH and CK-MB release in
culture medium
The cell culture medium was collected after the A/R
procedure. The concentrations of LDH and CK-MB were determined by a
colorimetric method using commercial kits (Nanjing Jiancheng
Bioengineering Institute) according to the manufacturer's protocol.
The results were reported as U/l.
Hoechst 33258 staining assay
Briefly, H9c2 cardiomyocytes were washed with
ice-cold PBS, fixed in 10% neutral buffered formalin for 10 min at
room temperature, and washed again in ice-cold PBS. The
cardiomyocytes were then exposed to Hoechst 33258 (2 µg/ml
in PBS) and incubated for 20 min at room temperature. The cells
were then washed three times in PBS and examined under a
fluorescence microscope with an appropriate filter. Images were
captured at a magnification of ×200 using an Olympus microscope
(1X71; Olympus Corp., Tokyo, Japan).
Measurement of caspase-3 activity
Caspase-3 activity was determined in cytosolic
protein extracts using a colorimetric activity assay kit (Beyotime
Institute of Biotechnology, Haimen, China) according to the
manufacturer's protocol. Specifically, after determining the
protein concentration, the supernatant was incubated with the
caspase-3 substrate (Ac-DEVD-pNA) on a 96-well-plate. The activity
of caspase-3 was detected using a microplate reader (Tecan) at 405
nm.
Protein extraction and western blot
analysis
H9c2 cells cultured in 6-well plates
(3×105/well) were washed twice with ice-cold PBS and
lysed via incubation on ice for 30 min with lysis buffer (pH 7.5)
containing 20 mM Tris-HCl, 120 mM NaCl, 1.0% Triton X-100, 10%
glycerol, 2 mM ethylene-diaminetetraacetic acid, and protease
inhibitor cocktail (Roche GmbH). Following centrifugation of the
cell lysates at 4°C for 20 min at 10,000 × g, the supernatants
(total cell extracts) were frozen and stored at −80°C. The protein
concentration of each sample was determined using a bicinchoninic
acid assay protein assay kit (Pierce Chemical, Rockford, IL,
USA).
For the western blot analysis, equal amounts of
cytosolic protein lysates were separated by sodium dodecyl
sulfate-polyacrylamide gel electrophoresis on 10–15% gels. The gels
were transferred onto polyvinylidene fluoride membranes (PVDF;
Bio-Rad, Hercules, CA, USA), blocked for 30 min at 37°C with 5%
non-fat dry milk, and incubated with the primary antibodies
(1:1,000 dilution) overnight at 4°C. After three washes with
Tris-buffered saline plus Tween-20, the membranes were incubated
with secondary antibodies (1:200 dilution) in Tris-buffered saline
plus Tween-20 for 30 min at 37°C and washed as described above.
Subsequently, the blots were developed by enhanced
chemiluminescence according to the manufacturer's instructions.
Statistical analysis
Data were presented as mean ± standard deviation.
The statistical significance of differences between the means were
determined using SPSS 19 software (SPSS, Inc., Chicago, IL, USA)
for Windows using one-way analysis of variance followed by the
Bonferroni post hoc test or Student's t-test, as appropriate.
Differences with P-values of <0.05 were considered statistically
significant.
Results
sAT protected rats against apoptosis
induced by MI/RI
Myocardial infarct size, plasma CK, and plasma LDH
were the primary indicators used to assess myocardial injury after
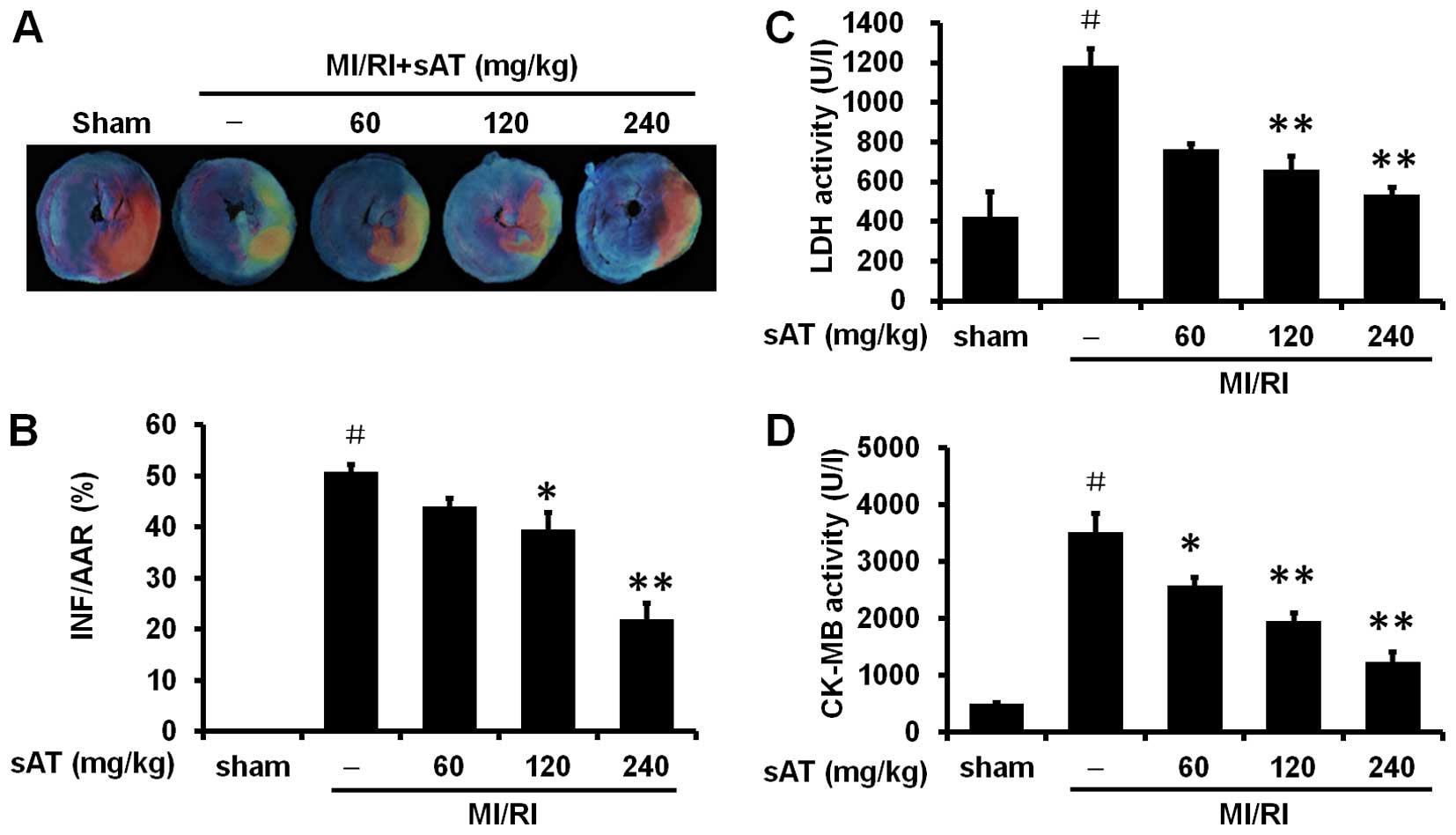
MI/RI. Representative areas at risk and infarct images are shown in
Fig. 1A and B. No myocardial
infarction was observed in hearts from the sham-operated group.
However, significant infarction was observed in rats in the MI/RI
group compared to rats in the sham group [50.8±1.5% (infarct area
as a percentage of area at risk), P<0.01]. Treatment with 120
and 240 mg/kg sAT significantly decreased infarct size compared to
the MI/RI-group (39.6±3.3%, P<0.05 and 22.0±3.0%, P<0.01,
respectively). There was no significant difference in the area at
risk between any of the groups.
Injury to the cardiomyocytes was determined by
measuring the levels of LDH and CK-MB in the serum at the end of
reperfusion (Fig. 1C and D).
MI/RI significantly increased LDH and CK-MB levels compared to
those in the sham group (1185.5±83.7%, P<0.01 and 3523.4±326.9%,
P<0.01, respectively), while 120 and 240 mg/kg sAT markedly
reduced the levels of LDH and CK-MB after MI/RI (P<0.01).
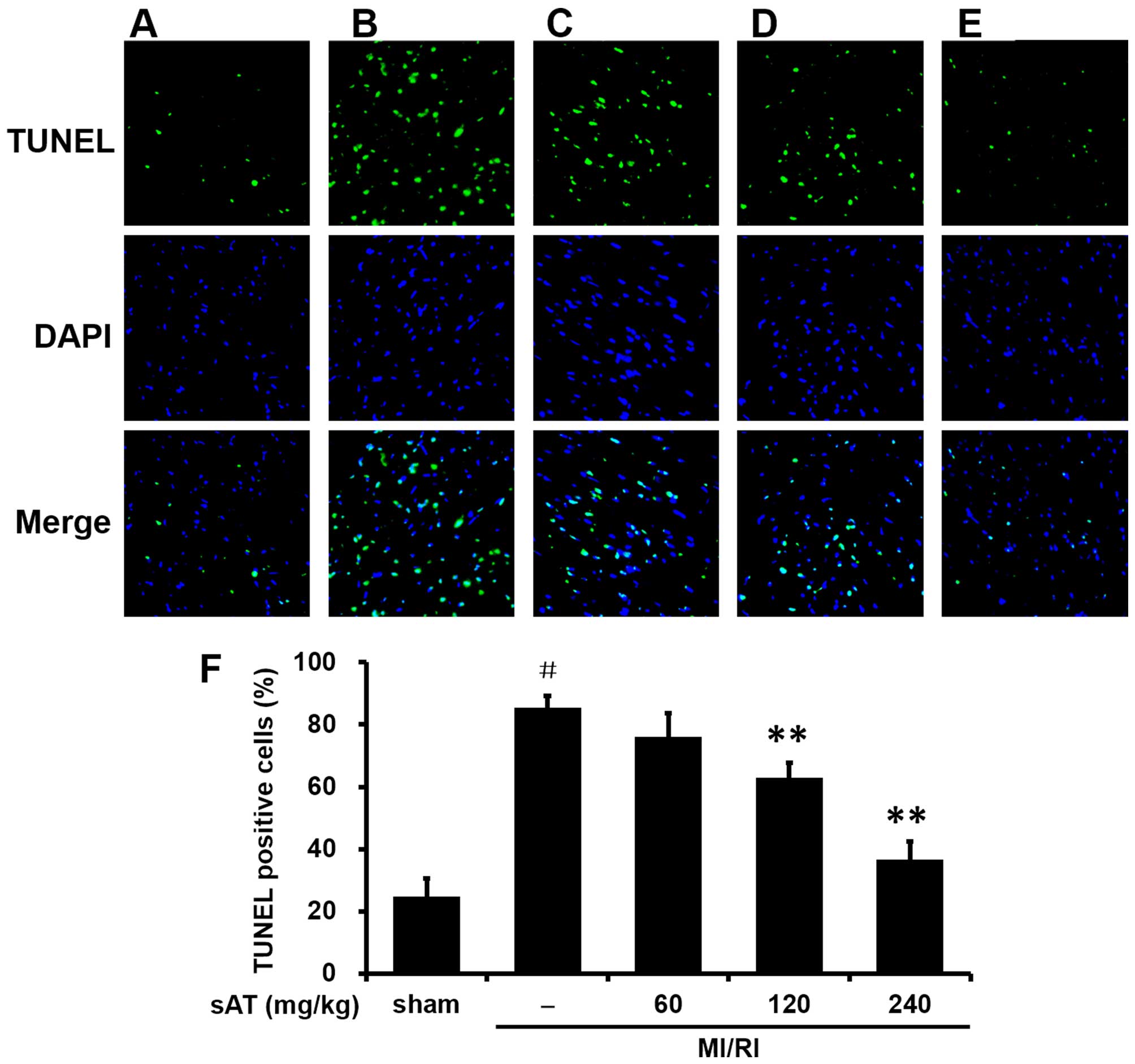
Consistent with these results, the MI/RI group
exhibited a significant increase in TUNEL-positive cells compared
with the sham-group (85.5±3.7%, P<0.01) (Fig. 2). Pretreatment with 120 or 240
mg/kg sAT substantially reduced the number of TUNEL-positive cells
(62.9±4.8%, P<0.01 and 36.6±5.7%, P<0.01, respectively).
Taken together, these data suggested that sAT decreased post-MI/RI
myocardial apoptosis in vivo.
sAT protects H9c2 cells following A/R
injury challenge
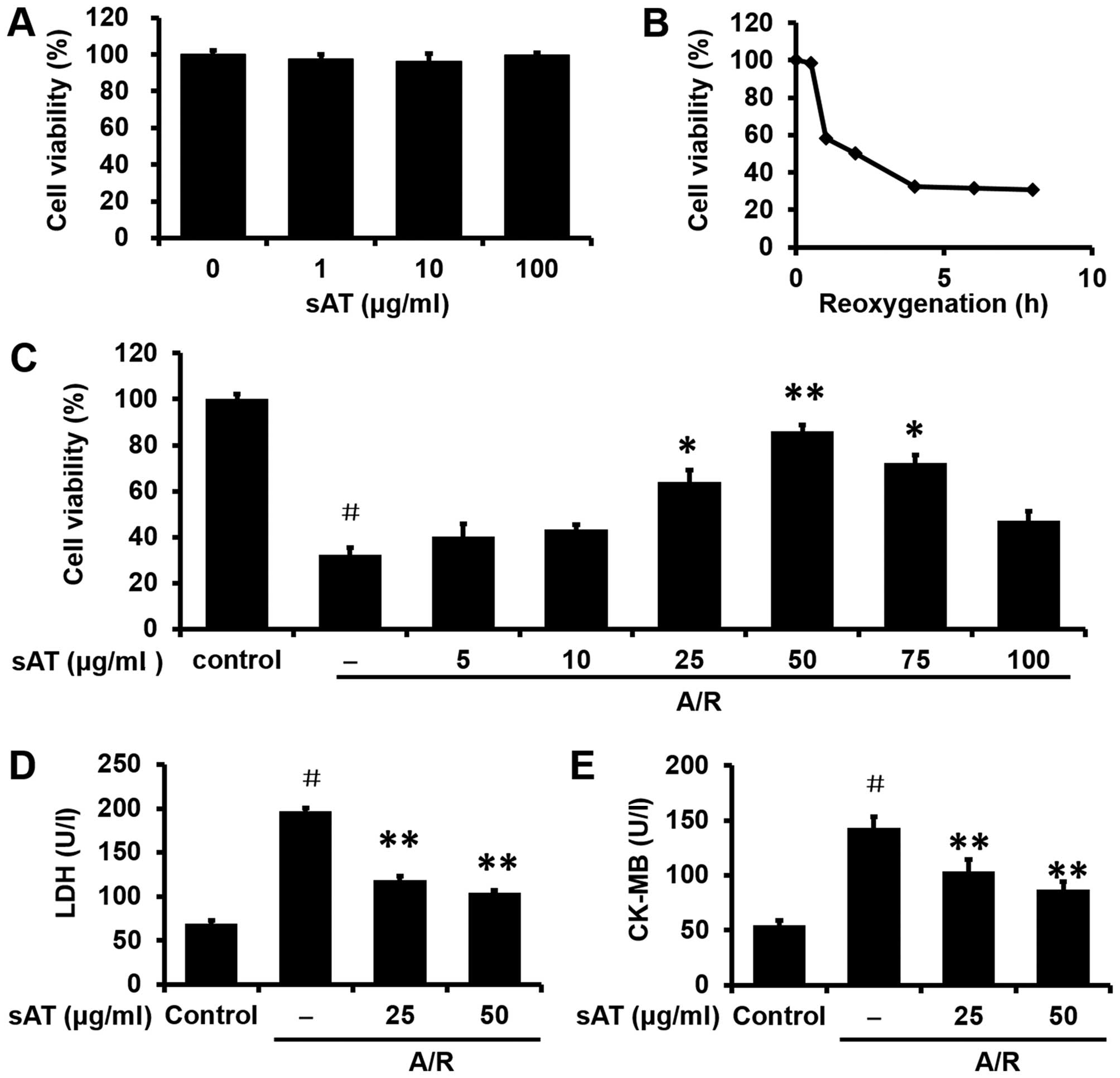
The toxicity of sAT was examined in cultured H9c2
cells incubated with sAT (1–100 µg/ml) for 24 h. As shown in
Fig. 3A, sAT did not alter cell
viability at concentrations reaching 100 µg/ml. The time
course of the loss of cell viability following A/R injury was also
assessed using MTT assays (Fig.
3B). Based on these data, 4 h was selected as the
re-oxygenation time to measure the markers of cell injury. Although
the A/R treatment significantly reduced cell survival, pretreatment
with sAT markedly increased survival in a concentration-dependent
manner. The 50 µg/ml concentration was selected for
subsequent investigation (Fig.
3C). LDH and CK-MB activities were measured in the supernatants
of culture media. An increase in LDH and CK-MB levels was observed
in the A/R group compared to the corresponding levels in the
control group (197.2±3.4%, P<0.01 and 143.5±9.7%, P<0.01,
respectively; Fig. 3D and E). The
levels of LDH and CK-MB were both decreased by sAT as compared to
the corresponding levels in the A/R group (P<0.01).
sAT protects H9C2 cardiomyocytes from
apoptosis induced by A/R injury
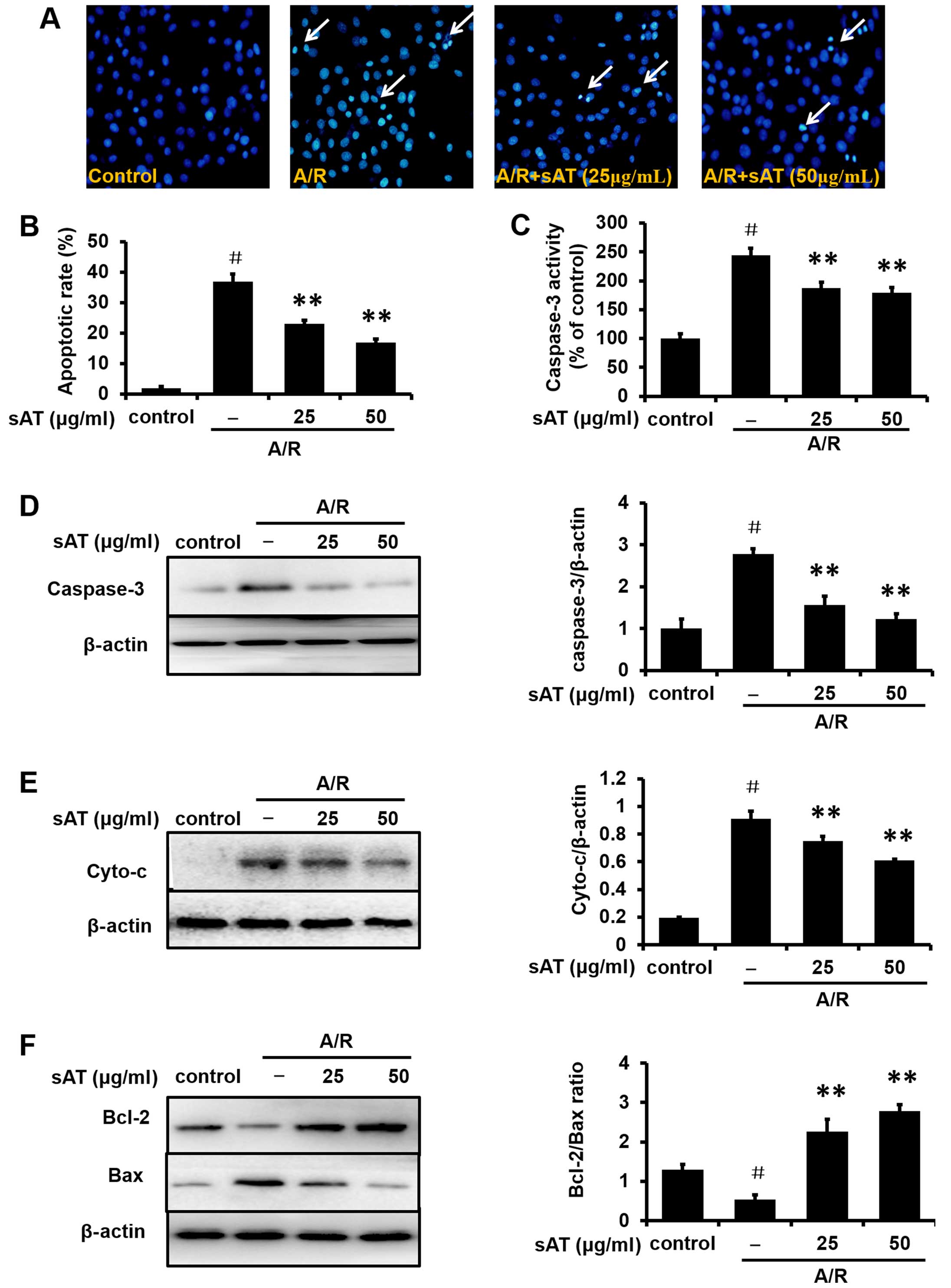
Subsequent to A/R injury, cardiomyocytes exhibited
typical characteristics of apoptosis, including shrinkage of the
nuclei, chromatin condensation, and the appearance of apoptotic
bodies. However, the number of cells with nuclear condensation and
fragmentation was significantly decreased in cardiomyocytes
incubated with sAT for 24 h following A/R injury (Fig. 4A). Quantitative analysis results
are shown in Fig. 4B. Similar
data were obtained from observation of the effects of sAT on
caspase-3 activity (Fig. 4C) and
protein expression of cleaved caspase-3 (Fig. 4D) and cytochrome c (Cyto-c;
Fig. 4E). The expression of Bax,
a pro-apoptotic protein, was markedly increased after A/R injury,
but was decreased in cells treated with sAT compared to A/R alone
(Fig. 4F). At the same time, the
expression of Bcl-2, an anti-apoptotic protein, decreased in the
A/R group and increased in the sAT-treated groups. The Bcl-2/Bax
ratio was also decreased in H9c2 cells following A/R injury
compared to normoxic control cells (0.54±0.12%, P<0.01), and
this decrease was reversed by treatment with sAT (2.27±0.31%,
P<0.01 and 2.78±0.17%, P<0.01, respectively).
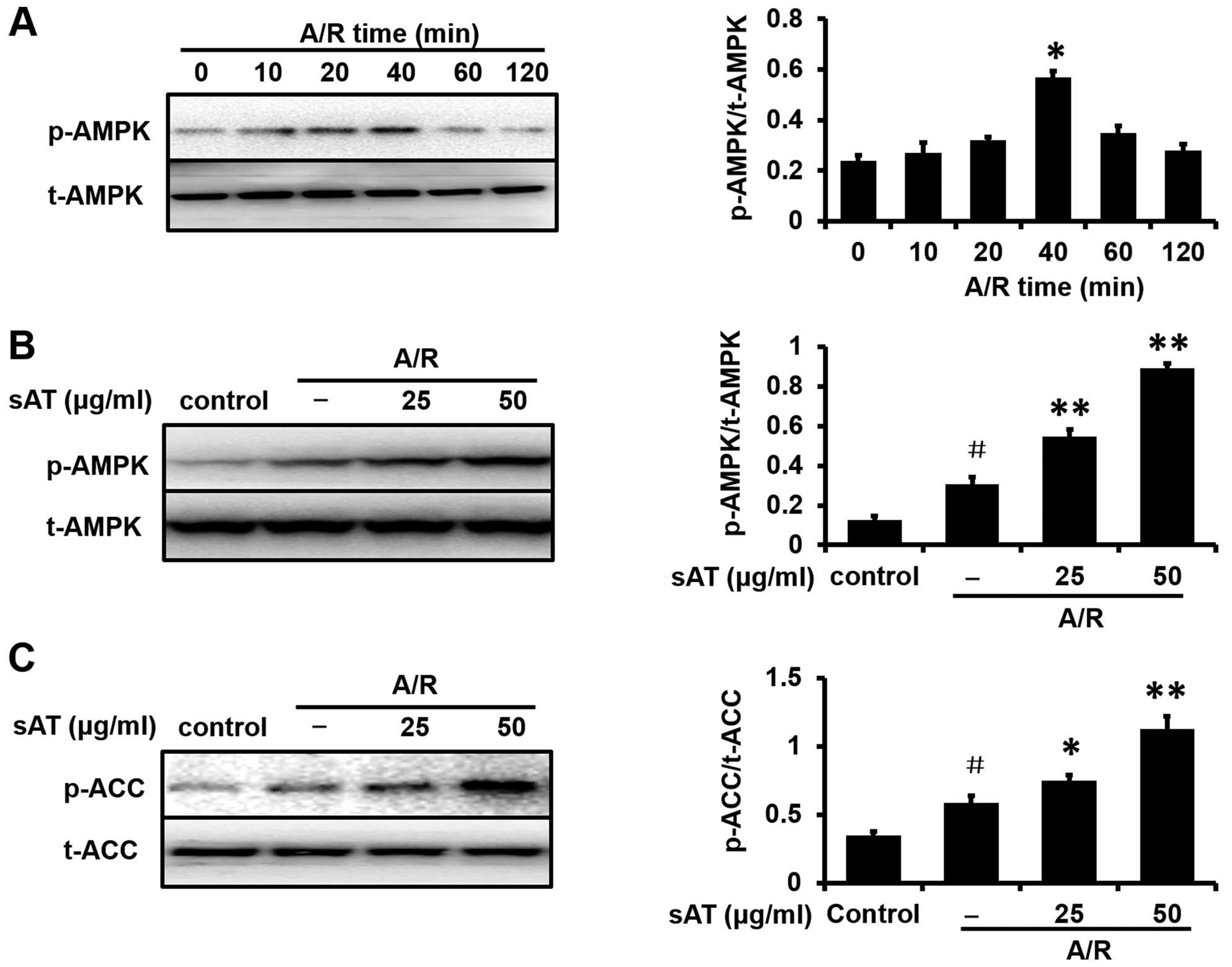
AMPK activation is involved in the
cardioprotective effects of sAT
To investigate the possible mechanisms involved in
the cardioprotective effects of sAT, the expression levels of
proteins associated with the AMPK pathway were determined by
western blot analysis. As shown in Fig. 5A, the phosphorylation of AMPK was
investigated in H9c2 cardiomyocytes exposed to A/R with different
re-oxygenation times (10, 20, 40, 60 and 120 min). Since the
highest level of p-AMPK (relative to total AMPK) was reached at 40
min, this time point was selected for subsequent mechanistic
studies. Enhancement of AMPK and ACC phosphorylation was observed
in the cells incubated with sAT after A/R injury compared with the
control group (Fig. 5B and
C).
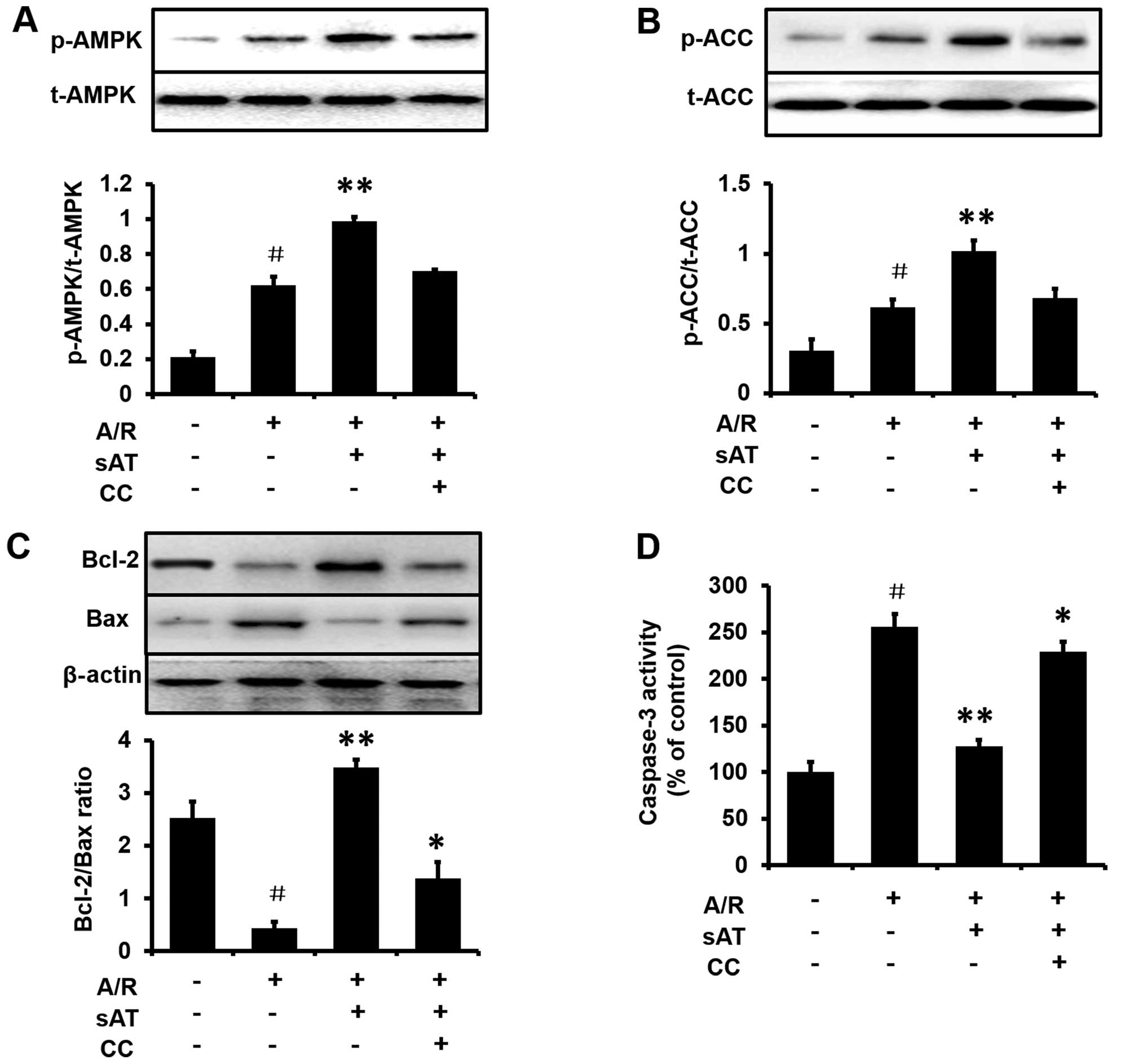
AMPK is required for the anti-apoptotic
effects of sAT in H9c2 cells
To investigate whether the AMPK pathway was required
for the protective effect of sAT in cardiomyocytes subjected to A/R
injury, cells were treated with the AMPK inhibitor, compound C (0.5
mM) for 1 h prior to incubation with sAT. Enhancement of AMPK
phosphorylation (Fig. 6A) and ACC
phosphorylation (Fig. 6B) by sAT
was eradicated by treatment with compound C. In addition, the
expression of Bax and Bcl-2 and the activity of caspase-3 were
measured following the incubation of cells under A/R with or
without compound C in the presence of sAT. As shown in Fig. 6C and D, compared with the
sAT-treated group, treatment with sAT plus compound C markedly
inhibited the increase in the Bcl-2/Bax ratio (1.38±0.31%,
P<0.05) and the reduction in caspase-3 activity (229.0±10.6%,
P<0.05).
Discussion
In the present study, we investigated the potential
protective effects of sAT against MI/RI in rats. We found that sAT
reduced the damage from MI/RI to the rat heart after left anterior
descending coronary artery occlusion. Rats treated with sAT had a
smaller infarct size compared with the MI/RI rats treated with
vehicle. Biochemical measures of cardiac damage (serum LDH and
CK-MB) also supported the protective effects of sAT. Furthermore,
the suppression of cardiomyocyte apoptosis and activation of the
AMPK pathway were observed in cardiomyocytes in vitro and
may contribute to the cardio-protective effects of sAT.
Acute ischemic injury resulting from coronary artery
disease leads to myocardial cell death, most likely due to ATP
depletion, increased calcium load, acidosis and oxidative stress
(23,24). In reperfusion injury, the
initiating event is the interruption and subsequent
re-establishment of blood supply to the tissue, which leads to
extensive secondary cardiac injury and tissue damage (25). In the past 20 years,
preconditioning and postconditioning treatments have been applied
to reduce reperfusion injury (26). However, the use of these
treatments in humans to reduce infarct size and improve
catheterization outcomes is impractical. Consequently, basic and
applied research should be directed at the identification of
pharmacological agents that can reproducibly mimic these techniques
and outcomes in humans.
Findings of previous studies have shown that sAT
exerts antidiabetic and anti-aging effects (16,18). Diabetes and aging are risk factors
for developing cardiovascular diseases, particularly ischemic heart
disease. In addition, it has been previously demonstrated that
Aralia species contain some ginseng-like triterpenoid
saponins. The effect produced by saponins from Aralia
species is similar to the effects of panaxosides from ginseng.
These effects may contribute to the ability of Aralia plants
to increase energy, strengthen the body, and improve the response
of the body to hypoxia with regard to the cardiovascular system
(27–30). In the present study, we identified
that sAT exhibited anti-MI/RI effects. Specifically, 120 and 240
mg/kg sAT significantly reduced infarct size in rat hearts. sAT
also caused increases in serum LDH and CK-MB, two cytosolic enzymes
that serve as diagnostic markers of ischemia injury to the
myocardium and leak out from damaged myocardial tissues to the
blood stream when the cell membrane becomes permeable or ruptures.
The two phenomena demonstrated that sAT may reduce and delay acute
MI/RI.
Apoptosis can lead to protein cleavage, DNA
breakdown and nuclear shrinkage (31). We found that pretreatment with sAT
minimized cardiac injury by preventing apoptosis induced by MI/RI
in vivo and in vitro. Notably, sAT pretreatment
markedly decreased the number of TUNEL-positive cells and prevented
morphological changes in the nucleus, as observed by Hoechst
staining, when compared with the vehicle-treated group. Thus, we
concluded that sAT had an anti-apoptotic role in the prevention of
MI/RI.
Mitochondrial dysfunction is a major cause of
cardiomyocyte death induced by MI/RI (32). ATP generation is the most
important function of mitochondria, particularly in the heart.
However, mitochondria also regulate apoptosis (33) through various pathways, such as
regulation of the activity of the Bcl-2 family, inhibition of
cytochrome c release and inhibition of caspase activation.
Thus, we examined the mechanism through which sAT protected against
apoptosis in cardiomyocytes. A discussion of each of these pathways
(i.e., regulation of Bcl-2 proteins, inhibition of Cyto-c release,
and suppression of caspase activation) is provided below.
The Bcl-2 family, key regulators of apoptosis,
comprises cell death promoters, such as Bax and Bad, and cell death
inhibitors, including Bcl-2 and Bcl-x (34). High Bax/Bcl-2 ratios are
associated with apoptotic activation. The results on early
apoptotic signaling following A/R showed that sAT impaired the
accumulation of Bax and upregulated the levels of Bcl-2 protein.
Thus, the Bcl-2/Bax ratio increased following treatment with sAT.
Therefore, Bcl-2 and Bax may be involved in mediating the
anti-apoptotic effects of sAT after A/R injury.
Cyto-c is a pro-apoptotic signaling molecule
localized within the mitochondrial cristae. Its release from the
mitochondria into the cytosol is a critical event in apoptosis
(35). In the present study,
pretreatment with sAT significantly suppressed the release of
Cyto-c from the mitochondria into the cytosol induced by A/R
injury. Of note, the release of Cyto-c from mitochondria
consequently activated caspase-3, which plays a pivotal role in
apoptosis (36). sAT markedly
suppressed the activity and expression of caspase-3, while A/R
increased the activity and expression of this critical apoptotic
regulator. Thus, the protective role of sAT may be mediated by
inhibition of the release of Cyto-c and suppression of the activity
of caspase-3.
AMPK is a serine/threonine kinase that senses energy
levels within the cell, and together with its downstream effector,
ACC, coordinates metabolic responses to maintain energy homeostasis
(37). Animals deficient in AMPK
exhibit increased cardiac hypertrophy, accelerated heart failure
and increased infarct sizes following coronary artery ligation
(38). The classic function of
AMPK is to maintain energy homeostasis by modulating metabolic
pathways (39,40). However, it has been demonstrated
that AMPK may protect against reperfusion injury by regulating cell
survival and limiting apoptosis rather than through its metabolic
actions (41). In the present
study, we observed a significant increase in the phosphorylation of
AMPK and its downstream target, ACC, during reperfusion. This
finding indicated that under stress conditions, sAT activated AMPK
to increase ATP production in order to maintain contractile
function and cell homeostasis. In addition, when the AMPK
inhibitor, compound C, was administered, the activation of AMPK and
ACC was suppressed. The anti-apoptotic effects of sAT were also
decreased by compound C. These results showed that pretreatment
with sAT conferred resistance to A/R injury-induced apoptosis by
activating the AMPK pathway in vitro. However, further
studies are needed to elucidate the underlying mechanism by using a
more specific animal model. The involvement of other signaling
pathways in the cardioprotective effects of sAT (different from
AMPK pathway) must also be addressed. In this manner, we could
powerfully assess the different curative effects between sAT and
positive control.
In summary, the results of the present study have
demonstrated that sAT pretreatment may reduce myocardial injury
in vitro and in vivo. The protective effects of sAT
may be due, in part, to the suppression of apoptosis and promotion
of survival via the AMPK pathway. Ongoing investigations of the
effects of sAT, including the active components and pharmacological
parameters, may lead to novel therapeutic strategies for the
treatment of myocardial ischemia and reperfusion injury.
Acknowledgments
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81470174 and
81001673).
Abbreviations:
|
AAR
|
ischemic-reperfused area-at-risk
|
|
ACC
|
acetyl CoA carboxylase
|
|
AMPK
|
AMP-activated protein kinase
|
|
Bax
|
Bcl-2-associated X protein
|
|
CK-MB
|
creatine kinase isoenzyme-MB
|
|
DMEM
|
Dulbecco's modified Eagle's medium
|
|
FBS
|
fetal bovine serum
|
|
INF
|
myocardial infarct size
|
|
LDH
|
lactate dehydrogenase
|
|
MI/RI
|
myocardial ischemia/reperfusion
injury
|
|
MTT
|
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl-tetrazolium bromide
|
|
sAT
|
total saponins extracted from Aralia
taibaiensis
|
|
TTC
|
2,3,5-triphenyltetrazolium
chloride
|
|
TUNEL
|
terminal deoxynucleotidyl
transferase-mediated dUTP nick end-labeling
|
References
|
1
|
Go AS, Mozaffarian D, Roger VL, Benjamin
EJ, Berry JD, Blaha MJ, Dai S, Ford ES, Fox CS, Franco S, et al:
American Heart Association Statistics Committee and Stroke
Statistics Subcommittee: Executive summary: heart disease and
stroke statistics–2014 update: a report from the American Heart
Association. Circulation. 129:399–410. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Fox KA, Steg PG, Eagle KA, Goodman SG,
Anderson FA Jr, Granger CB, Flather MD, Budaj A, Quill A, Gore JM,
et al: GRACE Investigators: Decline in rates of death and heart
failure in acute coronary syndromes, 1999–2006. JAMA.
297:1892–1900. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Lange RA and Hillis LD: Immediate
angioplasty for acute myocardial infarction. N Engl J Med.
328:726–728. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Simoons ML and Windecker S: Controversies
in cardiovascular medicine: Chronic stable coronary artery disease:
drugs vs. revascularization. Eur Heart J. 31:530–541. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Braunwald E and Kloner RA: Myocardial
reperfusion: A double-edged sword? J Clin Invest. 76:1713–1719.
1985. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Buja LM: Myocardial ischemia and
reperfusion injury. Cardiovasc Pathol. 14:170–175. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Yellon DM and Hausenloy DJ: Myocardial
reperfusion injury. N Engl J Med. 357:1121–1135. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Moens AL, Claeys MJ, Timmermans JP and
Vrints CJ: Myocardial ischemia/reperfusion-injury, a clinical view
on a complex pathophysiological process. Int J Cardiol.
100:179–190. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Mozaffari MS, Liu JY, Abebe W and Baban B:
Mechanisms of load dependency of myocardial ischemia reperfusion
injury. Am J Cardiovasc Dis. 3:180–196. 2013.PubMed/NCBI
|
|
10
|
Eefting F, Rensing B, Wigman J, Pannekoek
WJ, Liu WM, Cramer MJ, Lips DJ and Doevendans PA: Role of apoptosis
in reperfusion injury. Cardiovasc Res. 61:414–426. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
MacLellan WR and Schneider MD: Death by
design. Programmed cell death in cardiovascular biology and
disease. Circ Res. 81:137–144. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Abbate A, Bussani R, Amin MS, Vetrovec GW
and Baldi A: Acute myocardial infarction and heart failure: Role of
apoptosis. Int J Biochem Cell Biol. 38:1834–1840. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Fliss H and Gattinger D: Apoptosis in
ischemic and reperfused rat myocardium. Circ Res. 79:949–956. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Gill C, Mestril R and Samali A: Losing
heart: The role of apoptosis in heart disease–a novel therapeutic
target? FASEB J. 16:135–146. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Tang HF, Yi YH, Wang ZZ, Hu WJ and Li YQ:
Studies on the triterpenoid saponins of the root bark of Aralia
taibaiensis. Yao Xue Xue Bao. 31:517–523. 1996.In Chinese.
|
|
16
|
Xi M, Hai C, Tang H, Wen A, Chen H, Liu R,
Liang X and Chen M: Antioxidant and antiglycation properties of
triterpenoid saponins from Aralia taibaiensis traditionally used
for treating diabetes mellitus. Redox Rep. 15:20–28. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Tang HF, Yi YH, Wang ZZ, Jiang YP and Li
YQ: Oleanolic acid saponins from the root bark of Aralia
taibaiensis. Yao Xue Xue Bao. 32:685–690. 1997.In Chinese.
|
|
18
|
Li YN, Guo Y, Xi MM, Yang P, Zhou XY, Yin
S, Hai CX, Li JG and Qin XJ: Saponins from Aralia taibaiensis
attenuate D-galactose-induced aging in rats by activating FOXO3a
and Nrf2 pathways. Oxid Med Cell Longev. 2014:3205132014.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Weng Y, Yu L, Cui J, Zhu YR, Guo C, Wei G,
Duan JL, Yin Y, Guan Y, Wang YH, et al: Antihyperglycemic,
hypolipidemic and antioxidant activities of total saponins
extracted from Aralia taibaiensis in experimental type 2 diabetic
rats. J Ethnopharmacol. 152:553–560. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Li C, Liu Z, Tian J, Li G, Jiang W, Zhang
G, Chen F, Lin P and Ye Z: Protective roles of Asperosaponin VI, a
triterpene saponin isolated from Dipsacus asper Wall on acute
myocardial infarction in rats. Eur J Pharmacol. 627:235–241. 2010.
View Article : Google Scholar
|
|
21
|
Zhang JY, Chen ZW and Yao H: Protective
effect of urantide against ischemia-reperfusion injury via protein
kinase C and phosphtidylinositol 3′-kinase - Akt pathway. Can J
Physiol Pharmacol. 90:637–645. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Koyama T, Temma K and Akera T:
Reperfusion-induced contracture develops with a decreasing [Ca2+]i
in single heart cells. Am J Physiol. 261:H1115–H1122.
1991.PubMed/NCBI
|
|
23
|
Haunstetter A and Izumo S: Apoptosis:
Basic mechanisms and implications for cardiovascular disease. Circ
Res. 82:1111–1129. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Yue TL, Wang C, Gu JL, Ma XL, Kumar S, Lee
JC, Feuerstein GZ, Thomas H, Maleeff B and Ohlstein EH: Inhibition
of extracellular signal-regulated kinase enhances
Ischemia/Reoxygenation-induced apoptosis in cultured cardiac
myocytes and exaggerates reperfusion injury in isolated perfused
heart. Circ Res. 86:692–699. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Hausenloy DJ and Yellon DM:
Preconditioning and postconditioning: united at reperfusion.
Pharmacol Ther. 116:173–191. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Maxwell SR and Lip GY: Reperfusion injury:
a review of the pathophysiology, clinical manifestations and
therapeutic options. Int J Cardiol. 58:95–117. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang J, Lu S, Wang H and Zheng Q:
Protective role of Aralia elata polysaccharide on
mercury(II)-induced cardiovascular oxidative injury in rats. Int J
Biol Macromol. 59:301–304. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhang J, Wang H and Zheng Q:
Cardioprotective effect of Aralia elata polysaccharide on
myocardial ischemic reperfusion (IR) injury in rats. Int J Biol
Macromol. 59:328–332. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhang J, Wang H, Xue Y and Zheng Q:
Cardioprotective and antioxidant activities of a polysaccharide
from the root bark of Aralia elata (Miq.) Seem. Carbohydr Polym.
93:442–448. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang M, Xu X, Xu H, Wen F, Zhang X, Sun H,
Yao F, Sun G and Sun X: Effect of the total saponins of Aralia
elata (Miq) Seem on cardiac contractile function and intracellular
calcium cycling regulation. J Ethnopharmacol. 155:240–247. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mohanty IR, Maheshwari U, Joseph D and
Deshmukh Y: Bacopa monniera protects rat heart against
ischaemia-reperfusion injury: Role of key apoptotic regulatory
proteins and enzymes. J Pharm Pharmacol. 62:1175–1184. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Ertracht O, Malka A, Atar S and Binah O:
The mitochondria as a target for cardioprotection in acute
myocardial ischemia. Pharmacol Ther. 142:33–40. 2014. View Article : Google Scholar
|
|
33
|
Crow MT, Mani K, Nam YJ and Kitsis RN: The
mitochondrial death pathway and cardiac myocyte apoptosis. Circ
Res. 95:957–970. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xie Z, Koyama T, Suzuki J, Fujii Y,
Togashi H, Sawa H and Nagashima K: Coronary reperfusion following
ischemia: Different expression of bcl-2 and bax proteins, and
cardiomyocyte apoptosis. Jpn Heart J. 42:759–770. 2001. View Article : Google Scholar
|
|
35
|
Armstrong JS: Mitochondria: A target for
cancer therapy. Br J Pharmacol. 147:239–248. 2006. View Article : Google Scholar
|
|
36
|
Dorn GW II: Apoptotic and non-apoptotic
programmed cardiomyocyte death in ventricular remodelling.
Cardiovasc Res. 81:465–473. 2009. View Article : Google Scholar :
|
|
37
|
Steinberg GR and Kemp BE: AMPK in Health
and Disease. Physiol Rev. 89:1025–1078. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Zhang P, Hu X, Xu X, Fassett J, Zhu G,
Viollet B, Xu W, Wiczer B, Bernlohr DA, Bache RJ, et al: AMP
activated protein kinase-alpha2 deficiency exacerbates
pressure-overload-induced left ventricular hypertrophy and
dysfunction in mice. Hypertension. 52:918–924. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Horman S, Beauloye C, Vanoverschelde JL
and Bertrand L: AMP-activated protein kinase in the control of
cardiac metabolism and remodeling. Curr Heart Fail Rep. 9:164–173.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Lopaschuk GD: AMP-activated protein kinase
control of energy metabolism in the ischemic heart. Int J Obes.
32(Suppl 4): S29–S35. 2008. View Article : Google Scholar
|
|
41
|
Russell RR III, Li J, Coven DL, Pypaert M,
Zechner C, Palmeri M, Giordano FJ, Mu J, Birnbaum MJ and Young LH:
AMP-activated protein kinase mediates ischemic glucose uptake and
prevents postischemic cardiac dysfunction, apoptosis, and injury. J
Clin Invest. 114:495–503. 2004. View Article : Google Scholar : PubMed/NCBI
|