Introduction
Breast cancer is one of the most common malignancies
affecting women worldwide, and the second leading cause of
cancer-related mortality in women (1). The majority of breast cancer-related
deaths are caused by distant metastases from the primary tumor
site. Treatment failure mainly arises from cancer cell
proliferation, invasion and metastasis, which ultimately lead to
patient mortality. Invasion and metastasis are the fundamental
characteristics and major causes of morbidity and mortality in
patients with breast cancer. The metastatic process is initiated by
cancer cell invasion, which involves changes in cell adhesion, the
proteolytic degradation of the extracellular matrix (ECM) and the
migration of cancer cells through tissue (2). The ECM consists of type IV collagen,
laminin, heparan sulfate proteoglycan, nidogen, and fibronectin
(3). ECM degradation requires
extracellular proteinases, of which the matrix metalloproteinases
(MMPs) play an essential role in cancer metastasis (4,5).
The MMPs are a family of structurally conserved,
zinc-dependent endopeptidases, which are known to be involved in
the proteolytic degradation of the ECM. MMPs are divided into four
subclasses as follows: collagenases, gelatinases, stromelysins and
membrane-associated MMPs, based on their substrates (6). MMP-9 in particular, is considered to
be one of the critical MMPs involved in cancer cell invasion and
has been found to be directly associated with the invasion,
metastasis and poor prognosis of breast cancer (7,8).
Thus, inhibiting MMP-9 expression and/or its upstream regulatory
pathways, may prove to be critical in the treatment of malignant
tumors, including breast cancer. A variety of stimuli, including
growth factors (e.g., fibroblast growth factor-2, epidermal growth
factor and hepatocyte growth factor), cytokines (e.g., tumor
necrosis factor-α), oncogenes (e.g., Ras) and
12-O-tetradecanoylphorbol-13-acetate (TPA) are all known to
induce MMP-9 expression (9-13).
Among these stimuli, TPA is a well known, selective activator of
protein kinase C (PKC) (10)
which stimulates MMP-9 synthesis and secretion during breast cancer
cell invasion (14,15). Cytokine and TPA treatments have
been shown to induce MMP-9 expression through the activation of
transcription factors, such as nuclear factor (NF)-κB (16,17), which possess binding sites on the
MMP-9 gene promoter (18).
The NF-κB element is centrally involved in the inductoin of
MMP-9 gene expression by TPA (16,19).
Peucedanum japonicum Thunb. (PJT) is a
medicinal plant which belongs to the Umbelliferae family. PJT
leaves are traditionally consumed as a medicinal herb to treat
coughs in Japan and Korea. The PJT root has also been used as a
folk medicine for colds and neuralgia in Taiwan. Previous in
vitro studies have reported that PJT possesses antioxidant
activity (20), inhibits
tyrosinases (21), possesses
anti-obesity properties (22) and
opposes platelet aggregation (23). Furthermore, it has been
hypothesized that PJT may have anti-metastatic properties based on
its possible inhibition of cell invasion (24,25). However, the mechanisms through
which PJT exerts anti-invasive effets remain poorly understood.
In this study, we addressed this hypothesis by
assessing the potential effects of PJT on TPA-induced cell invasion
and MMP-9 expression in MCF-7 human breast cancer cells, and
exploring the related molecular mechanisms. Our findings
demonstrated that PJT suppressed TPA-induced MMP-9 expression by
blocking NF-κB through PKCα, and that the suppression of MMP-9
expression correlated with the inhibition of cell invasion.
Materials and methods
Cells and materials
MCF-7 cells were obtained from the American Type
Culture Collection (ATCC; Manassas, VA, USA). The cells were
cultured in Dulbecco's modified Eagle's medium (DMEM) supplemented
with 10% fetal bovine serum (FBS) and 1% antibiotics at 37°C in a
5% CO2 incubator. PJT, TPA,
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
and anti-β-actin antibody (Cat. no. A5441) were obtained from
Sigma-Aldrich (St. Louis, MO, USA). Antibodies against
phosphorylated (p)-IκBα (Cat. no. #2859) and p-IKKαβ (Cat. no.
#2697) and IKKα (Cat. no. #2682) and IKKβ (Cat. no. #2678) were
purchased from Cell Signaling Technology, Inc. (Beverly, MA, USA).
Antibodies against MMP-9 (Cat. no. SC-12759), p50 (Cat. no.
SC-7178), p65 (Cat. no. SC-372) and proliferating cell nuclear
antigen (PCNA; Cat. no. SC-7907) and horseradish peroxidase
(HRP)-conjugated IgG (Cat. no. SC-2004,2005) were purchased from
Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA).
Preparation of PJT extracts
Roots of PJT were purchased from Kwangmyungdang
Medicinal Herbs Co. Ltd. (Ulsan, Korea) and authenticated by
Professor G.S. Lee, one of the authors of this article. A voucher
specimen (WKU010107-PJ201305E) has been deposited at the Department
of Herbology, College of Korean Medicine (Wonkwang University,
Iksan, Korea). The powdered PJT (100 g) was extracted using
sonication with 1,000 ml of 70% aqueous ethanol for 1 h. The
extract was evaporated under 40 mmHg pressure using a rotary
evaporator and then freeze-dried. The yield of the final extract
was 12.02% (w/w).
Determination of cell viability
The effect of PJT on MCF-7 cell viability was
determined using an established MTT assay. Briefly,
3×104 cells were seeded in wells and incubated at 37°C
for 24 h to allow attachment. The attached cells were left
untreated or treated with 1, 10, 25, 50 and 100 µg/ml PJT
for 24 h at 37°C. The cells were then washed with PBS prior to the
addition of MTT (0.5 mg/ml in PBS) and incubated at 37°C for 30
min. Formazan crystals were dissolved with dimethylsulf-oxide (100
µl/well) and detected at 570 nm using a Model 3550
microplate reader (Bio-Rad, Richmond, CA, USA).
Western blot analysis
The MCF-7 cells (5×105) were treated with
PJT (25, 50 and 100 µg/ml) for 1 h and then incubated with
TPA for 24 h at 37°C. The cells were then lysed with ice-cold
M-PER® Mammalian Protein Extraction Reagent (Pierce
Biotechnology, Inc., Rockford, IL, USA). The protein concentration
in the lysate was determined using the Bradford method (26). Samples (20 µg) were
separated by sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) with 10% acrylamide and transferred onto
Hybond™-polyvinylidene fluoride membranes (GE Healthcare Life
Sciences, Little Chalfont, UK) using a western blot apparatus. Each
membrane was blocked for 2 h with 2% bovine serum albumin or 5%
skim milk, and then incubated overnight at 4°C with 1 µg/ml
of a 1:2,000 dilution of primary antibody. HRP-conjugated IgG
(1:2,000 dilution) was used as the secondary antibody. Protein
expression levels were determined by signal analysis using an image
analyzer (FujiFilm, Tokyo, Japan).
Gelatin zymography
Gelatin zymography was performed as previously
described (27). Conditioned
media were collected after 24 h of stimulation, mixed with
non-reducing sample buffer, and electrophoresed in a polyacrylamide
gel containing 0.1% (w/v) gelatin. The gel was washed at room
temperature for 30 min with 2.5% Triton X-100 solution, and
subsequently incubated at 37°C for 16 h in 5 mM CaCl2,
0.02% Brij and 50 mM Tris-HCl (pH 7.5). The gel was stained for 30
min with 0.25% (w/v) Coomassie Brilliant Blue (Sigma-Aldrich) in
40% (v/v) methanol and 7% (v/v) acetic acid, and photographed on an
image analyzer (FujiFilm). Proteolysis was imaged as a white zone
in a dark blue field. Densitometric analysis was performed using
Multi Gauge Image Analysis software (FujiFilm).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
RT-qPCR was performed as previously described
(27). Total RNA was extracted
from the MCF-7 breast cancer cells using a FastPure™ RNA kit
(Takara Bio, Otsu, Japan). The RNA concentration and purity were
determined by absorbance at 260/280 nm. cDNA was synthesized from 1
µg total RNA using a PrimeScript™ RT reagent kit (Takara
Bio). MMP-9 and glyceraldehyde 3-phosphate dehydrogenase
(GAPDH) mRNA expression were determined by quantitative
(real-time) PCR (qPCR) using the ABI PRISM 7900 sequence detection
system and SYBR®-Green (Applied Biosystems, Foster City,
CA, USA). The following primers were used: MMP-9 (NM 004994)
sense, CCTGGAGACCTGAGAACCAATCT and antisense,
CCACCCGAGTGTAACCATAGC; and GAPDH (NM 002046) sense,
ATGGAAATCCCATCACCATCTT and antisense, CGCCCCACTTGATTTTGG. To
control for variations in mRNA concentration, all results were
normalized to those of the housekeeping gene, GAPDH.
Relative quantification was performed using the comparative ΔΔCt
method according to the manufacturer's instructions.
Preparation of nuclear extract
Nuclear extract was prepared as previously described
(27). The MCF-7 cells
(2×106) were treated with PJT in the presence or absence
of TPA for 4 h. The cells were immediately washed twice, scraped
into 1.5 ml of ice-cold PBS (pH 7.5) and pelleted at 1,500 × g for
3 min. Cytoplasmic and nuclear extracts were prepared from the
cells using NE-PER® Nuclear and Cytoplasmic Extraction
reagents (Pierce Biotechnology, Inc.).
Membrane fractionation
The MCF-7 cells (5×107) were treated with
PJT (100 µg/ml) for 1 h and then incubated with TPA for 1 h
at 37°C. The cells were immediately washed twice, scraped into 1.5
ml of ice-cold PBS (pH 7.5), and pelleted at 4,000 rpm for 3 min.
Cell lysis was carried out in homogenization buffer (20 mM
Tris-HCl, 5 mM DTT, 2 mM EDTA, 5 mM EGTA, protease inhibitor,
phosphatase inhibitors, pH 7.5) with brief soni-cation (5 times for
10 sec each at 10% amplitude) after incubating on ice for 15-30
min. Cell debris was removed by centrifuging the sample at 3,000
rpm for 10 min at 4°C. Cell lysate was centrifuged at 13,000 rpm
for 30 min at 4°C to separate out soluble (cytosolic) and pellet
(membrane) fractions. The pellet fraction was incubated in
homogenization buffer containing 1% Triton X-100 for 30 min on ice,
centrifuged at 50,000 rpm for 1 h and the supernatant was collected
as the membrane fraction.
Electrophoretic mobility shift assay
(EMSA)
The activation of NF-κB was assayed using a gel
mobility shift assay using nuclear extracts. An oligonucleotide
containing the κ-chain (κB, 5′-CCGGTTAACAGAGGGGGCTTTCCGAG-3′)
binding site was synthesized and used as a probe for the gel
retardation assay. The two complementary strands were annealed and
labeled with [α-32P]dCTP. Labeled oligonucleotides
(10,000 cpm), 10 µg of nuclear extracts and binding buffer
[10 mM Tris-HCl, pH 7.6, 500 mM KCl, 10 mM EDTA, 50% v/v glycerol,
100 ng poly (dI·dC) and 1 mM dithiothreitol] were then incubated
for 30 min at room temperature in a final volume of 20 µl.
The reaction mixtures were analyzed by electrophoresis on 4%
polyacrylamide gels in 0.5X Tris-borate buffer. The gels were dried
and examined by autoradiography. Specific binding was controlled by
competition with 50-fold excess of cold κB oligonucleotide.
Invasion assay
Invasion assay was performed as previously described
(27). The invasion assay was
carried out in 24-well chambers (8-µm pore size) coated with
20 µl Matrigel diluted with DMEM. The Matrigel coating was
rehydrated in 0.5 ml DMEM for 30 min immediately before the
experiments. The MCF-7 cells (2×105) were added to the
upper chamber with chemoattractant in the bottom well. Conditioned
medium (0.5 ml) was added to the lower compartment of the invasion
chamber. The chambers were incubated for 24 h. Following
incubation, the cells on the upper side of the chamber were removed
using cotton swabs, and the cells that had migrated were fixed and
stained with Toluidine blue (Sigma-Aldrich) solution. Invading
cells were counted in 5 random areas of the membrane using a Leica
DM IL LED Inverted Lab Microscope (Leica, Wetzlar, Germany).
Analyzed data are the means ± SE from 3 individual experiments
performed in triplicate.
Statistical analysis
Statistical data analysis was performed using
analysis of variance (ANOVA) and Duncan's test. A value of
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effect of PJT on of MCF-7 cell
viability
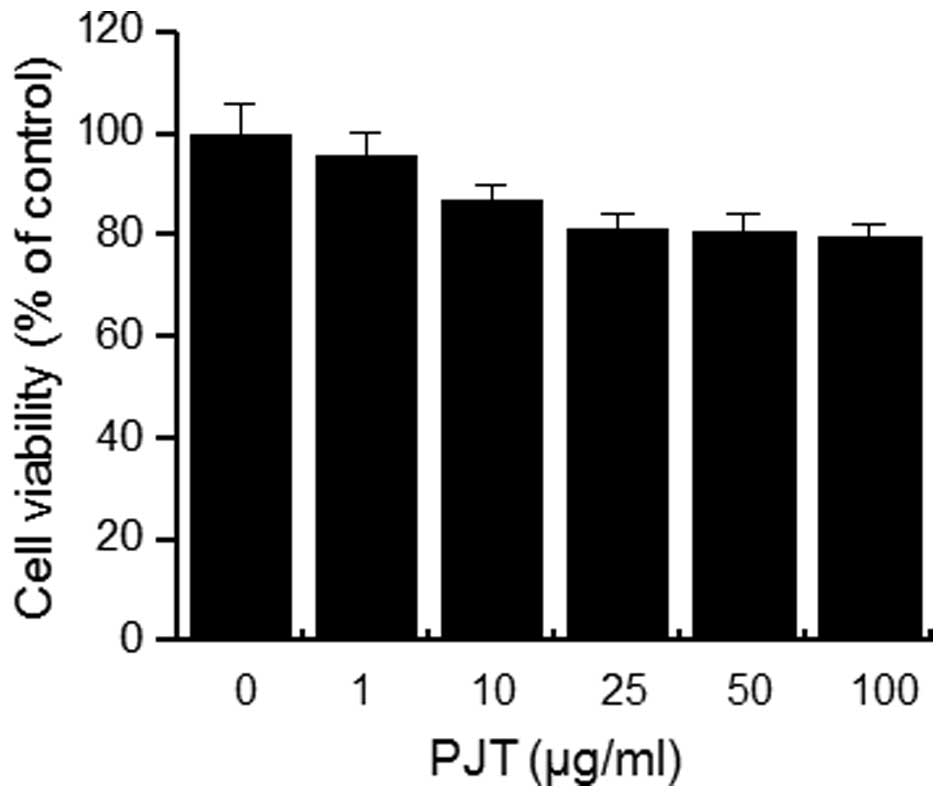
In order to investigate the cytotoxicity of PJT on
MCF-7 cells, the cells were seeded into wells of 96-well culture
plates at a density of 3×104 cells/well. The toxicity of
PJT on MCF-7 cells was determined by MTT assay. Treatment of the
MCF-7 cells with all concentrations of PJT for 24 h did not cause
any significant change in cell viability (Fig. 1). Thus, subsequent experiments
utilized the optimal non-toxic concentration (100 µg/ml) of
PJT.
PJT suppresses the mRNA and protein
expression as well as the secretion of MMP-9 induced by TPA in
MCF-7 cells
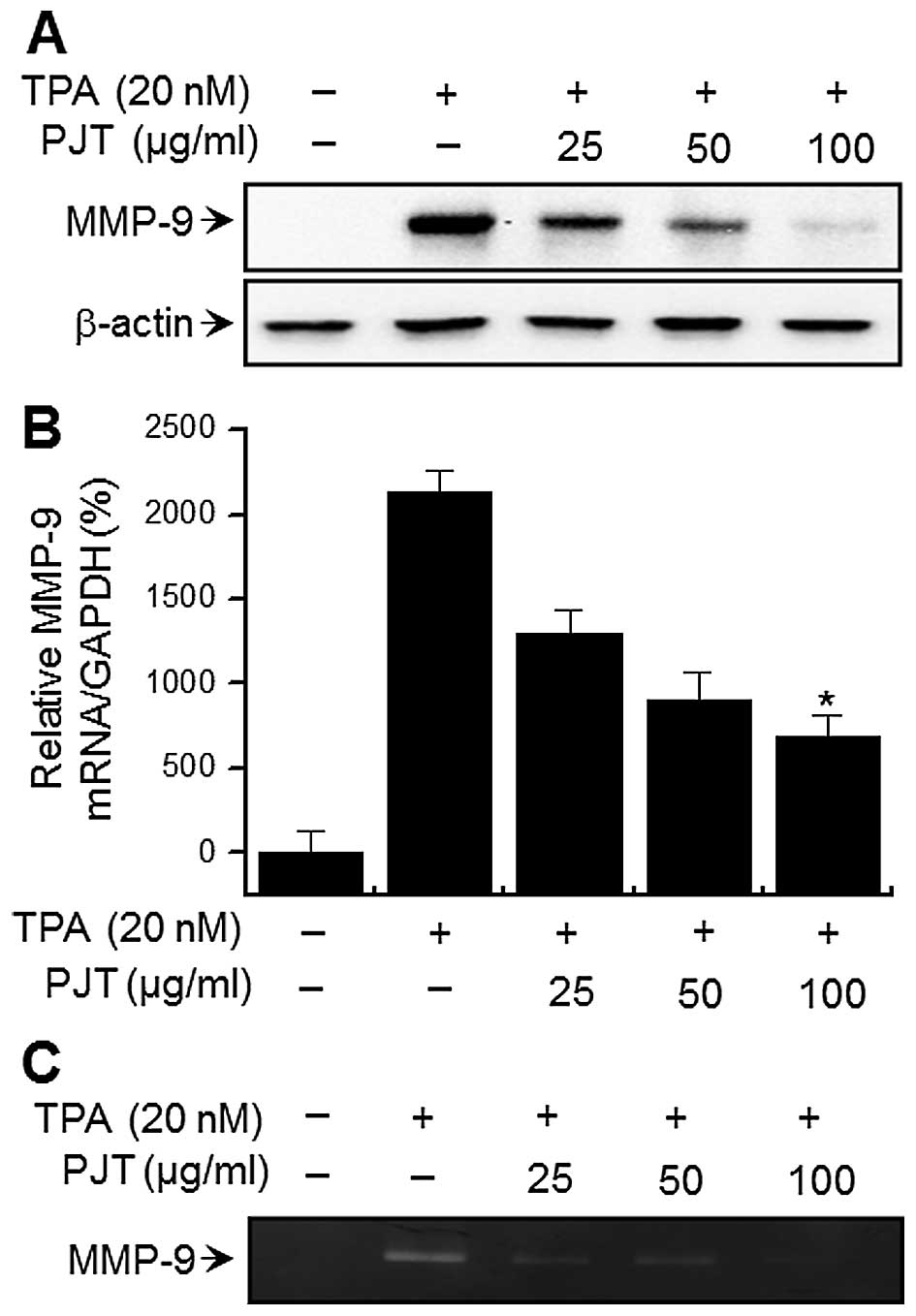
To examine the effects of PJT on TPA-induced MMP-9
expression, western blot analysis, RT-qPCR, and gelatin zymography
were performed on the MCF-7 cells. Western blot analysis revealed
that treatment of the MCF-7 cells with PJT blocked the upregulation
of TPA-induced MMP-9 protein expression (Fig. 2A). RT-qPCR revealed a TPA-induced
increase in MMP-9 expression levels in the MCF-7 cells, and
that PJT blocked the TPA-induced MMP-9 upregulation in a
dose-dependent manner (Fig. 2B).
To determine the effects of PJT on TPA-induced MMP-9 secretion,
gelatin zymography was performed, which demonstrated that treatment
of the MCF-7 cells with TPA increased MMP-9 secretion. However, PJT
significantly diminished TPA-induced MMP-9 secretion (Fig. 2C). These results demonstrated that
PJT exerts potent inhibitory effects on TPA-induced MMP-9
expression in MCF-7 cells.
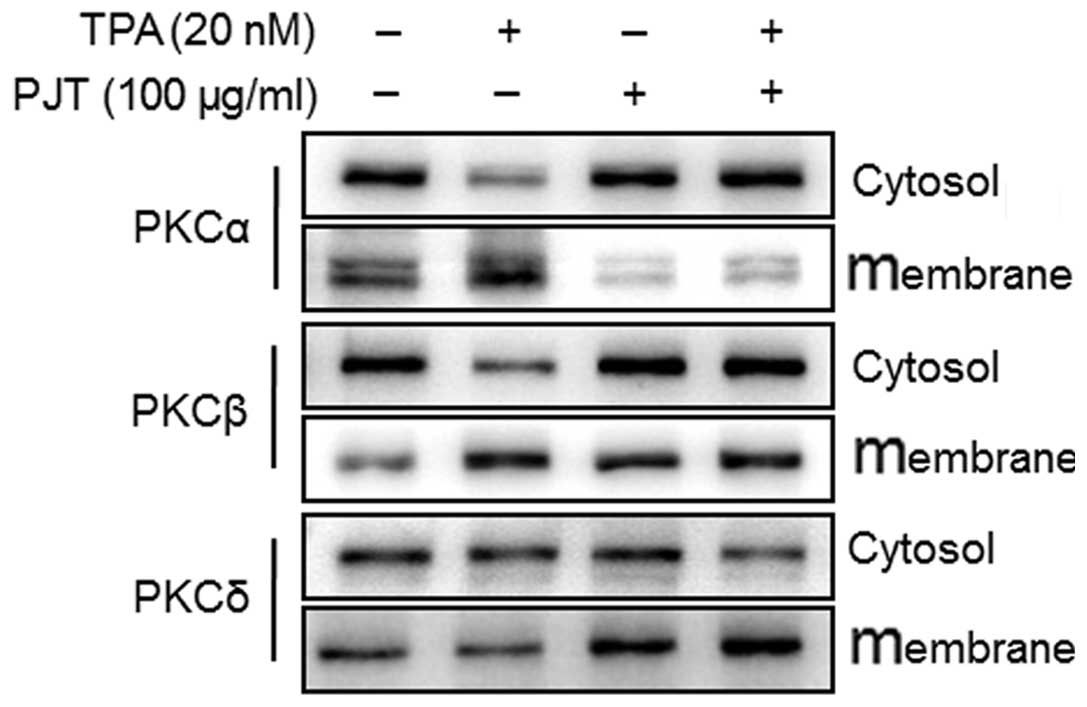
PJT inhibits the membrane localization of
PKCα induced by TPA
To determine whether PJT affects the levels of
various PKC isotypes in the TPA-treated MCF-7 cells, we measured
the levels of PKCα, PKCβ and PKCδ in the cytosolic and membrane
fractions. The TPA-induced membrane localization of PKCα was
blocked by pre-treatment with PJT for 1 h (Fig. 3). These results suggested that
TPA-induced MMP-9 expression involved PKCα activation in MCF-7
cells, and PJT inhibited TPA-induced PKCα activation.
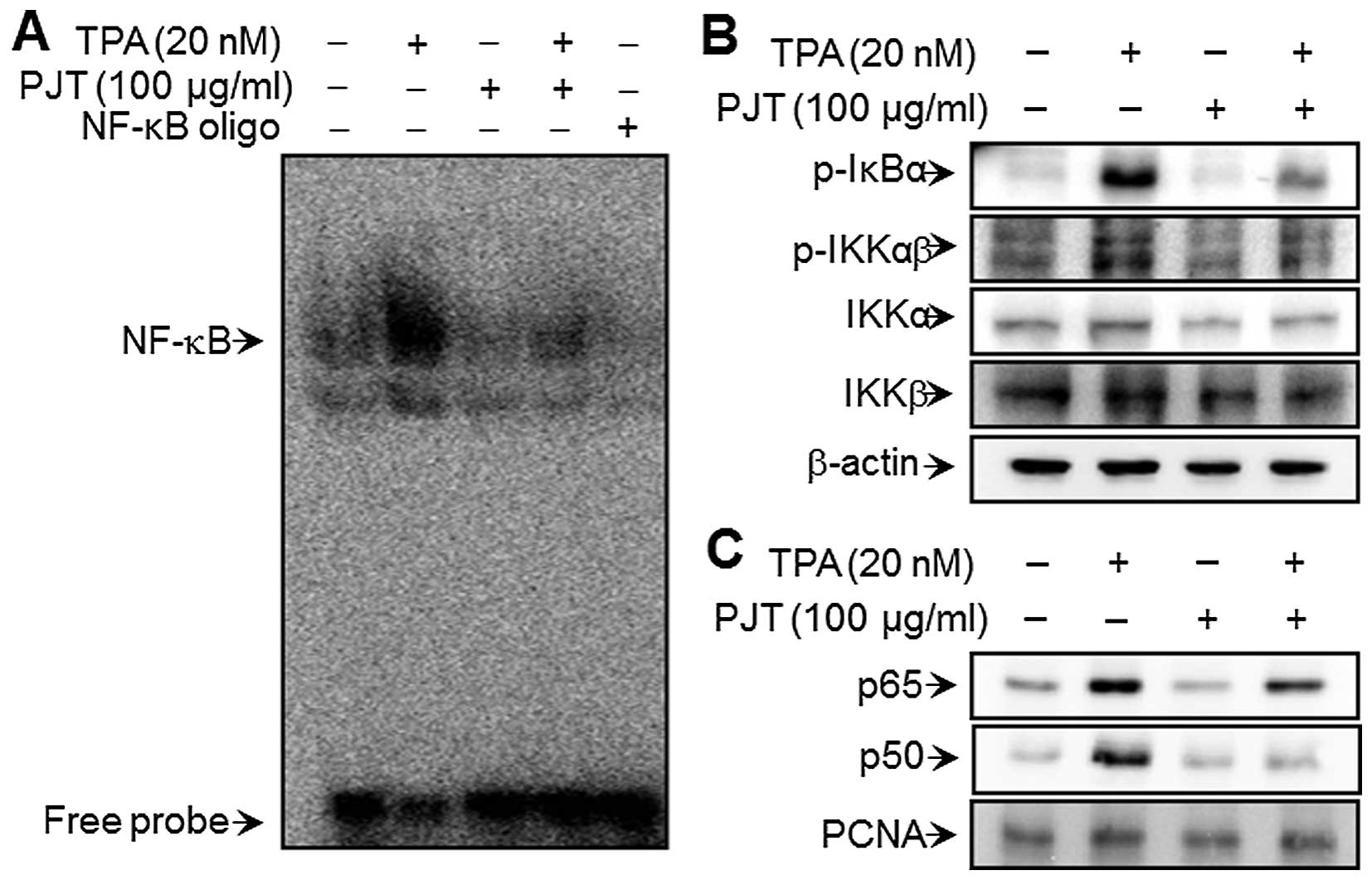
PJT inhibits TPA-induced NF-κB DNA
binding activity
To elucidate the mechanisms through which PJT
inhibits MMP-9 expression, the effect of PJT on the TPA-induced
activation of NF-κB was evaluated using EMSA. As shown in Fig. 4A, TPA substantially increased
NF-κB binding activity. Pre-treatment with PJT inhibited the
TPA-induced NF-κB binding activity; however, PJT alone had no
effect on NF-κB binding activity. These results are consistent with
the hypothesis that PJT specifically blocks NF-κB activation in
MCF-7 cells. Western blot analysis revealed that TPA induced the
phosphorylation of IKKαβ and IκBα, as well as that of IKKα and IKKβ
in the cytoplasm and, thereby, the nuclear translocation of NF-κB
subunits p50 and p65. These alterations were suppressed in the TPA
MCF-7 cells treated with that had also been treated with PJT
(Fig. 4B and C).
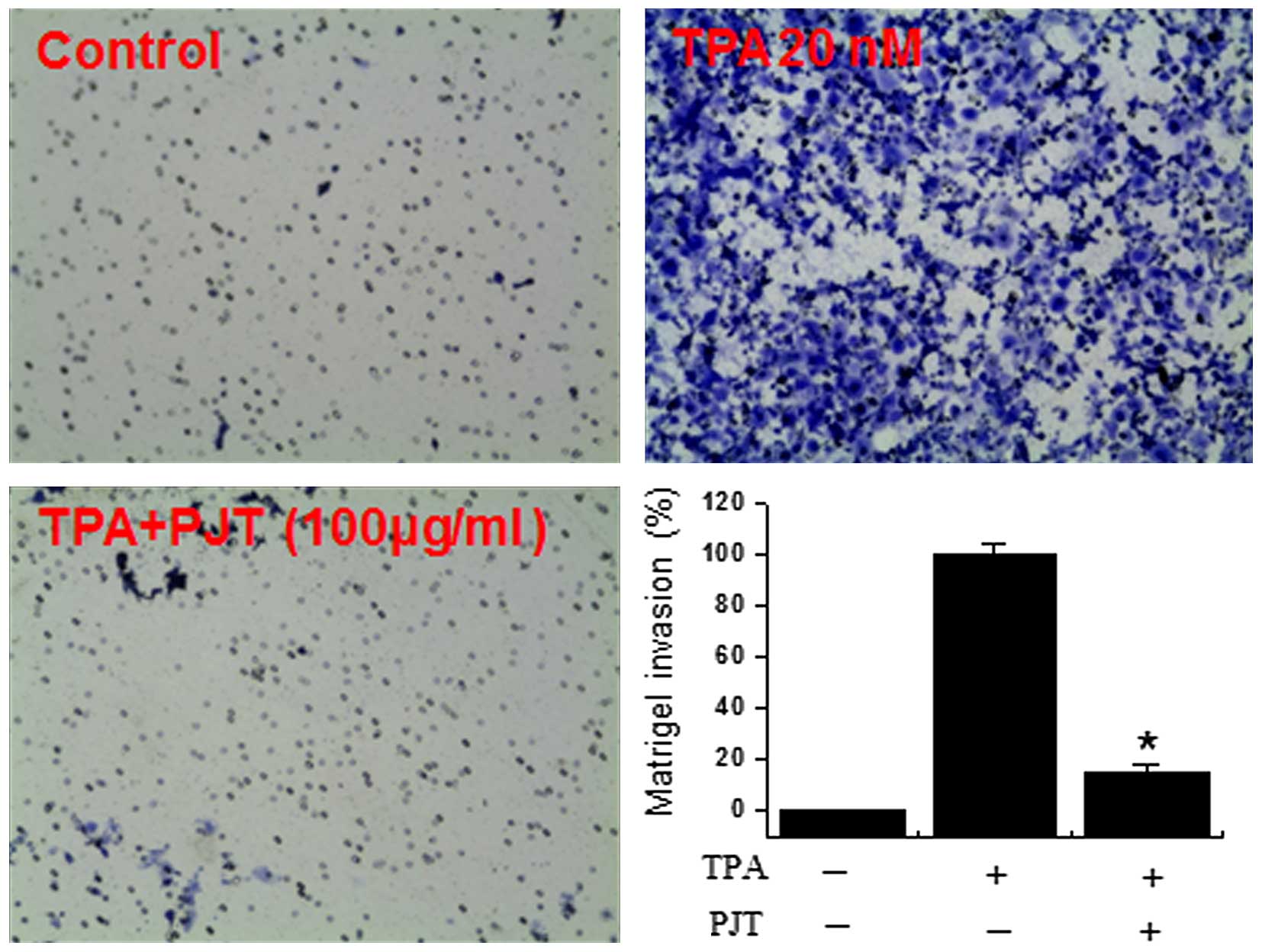
Effect of PJT on TPA-induced MCF-7 cell
invasion in vitro
It has been previously reported that the
upregulation of MMP-9 expression contributes to the invasive
ability of cancer cells (22,23). Thu, in this study, we investigated
the inhibitory effects of PJT on the invasive ability of MCF-7
breast cells by performing a Matrigel™ invasion assay. We treated
the cells with 100 µg/ml PJT prior to treatment with 20
µM TPA for 1 h and subsequently incubated the cells for 24
h. The level of TPA-induced cell invasion was significantly
increased compared with the untreated control cells. However,
TPA-induced cell invasion was suppressed by PJT treatment (Fig. 5).
Discussion
Metastasis is the primary cause of breast cancer
mortality. Tumor metastasis is a multistep process through which a
subset of individual cancer cells disseminate from a primary tumor
to distant secondary organs or tissues. This process involves cell
proliferation, ECM degradation, cell migration, and tumor growth at
metastatic sites (15,28). Cancer cell invasion is an early
step in the metastatic cascade, representing the beginning of the
transition from benign to malignant cancer. Tumor invasion is
morphologically associated with a distorted edge of the primary
tumor where individual or cohorts of cancer cells actively invade
the ECM surrounding the primary tumor (29). MMP-9 is important in tumor
metastasis as it is involved in basement membrane cleavage, which
allows cells with a migratory phenotype to be more invasive and
motile (23,25,30). In previous studies, inflammatory
cytokines, growth factors and phorbol esters have been shown to
stimulate MMP-9 by activating different intracellular signaling
pathways in breast cancer cells (31-33). The inhibition of MMP-9 expression
may thus prove to be an important therapeutic target the
development of therapies against tumor metastasis. This study
provides the first evidence, to the best of our knowledge, that PJT
inhibits TPA-induced cell invasion by suppressing MMP-9 expression
in MCF-7 breast cancer cells.
TPA increases the invasiveness of MCF-7 cells by
activating MMP-9 through PKCs. The activation of PKCs has been
shown to correlate significantly with an increased invasiveness in
breast carcinomas (34). TPA
activates conventional (α, βI, βII and γ) and novel (δ, ε, θ and η)
PKCs by binding to the C1 domains of these isoforms (35). The activation of PKCs by TPA
involves the translocation of PKC isoforms to the plasma membrane,
resulting in proliferation, differentiation and malignant
transformation, as well as tumor promotion and the progression in
cancer cells. In a previous study by our group, TPA stimulation
resulted in the translocation of PKCα and PKCδ from the cytosol to
the cell membrane, although the translocation of PKCβ was not
observed. Treatment with a non-toxic dose of a PKCδ inhibitor
(rottlerin), a broad PKC inhibitor (GF109203X) and a PKCα inhibitor
(Go6976) caused the marked inhibition of TPA-induced MMP-9
expression and secretion (36).
These data indicated that the TPA-induced activation of PKCα and
PKCδ mediated MMP-9 expression and secretion (36). In the present study, we
demonstrated that PJT inhibited the TPA-induced membrane
localization of PKCα, but not that of PKCδ (Fig. 3) in MCF-7 cells.
NF-κB, which has been reported to be regulated by
PKC isoforms (37,38) is a transcription factor that
regulates MMP-9 expression through binding sites on its
promoter (18). NF-κB is an
inducible, dimeric transcription factor that belongs to the
Rel/NF-κB family of transcription factors and consists of two major
polypeptides, p65 and p50 (39).
NF-κB is initially located in the cytoplasm in an inactive form
complexed with IκB, an inhibitory factor of NF-κB. Various
inducers, such as TPA, cytokines and stress cause the dissociation
of this complex, presumably through IκB phosphorylation, which
results in NF-κB being released from the complex. NF-κB is then
translocated to the nucleus, where it interacts with specific DNA
recognition sites to mediate gene transcription. NF-κB elements are
centrally involved in MMP-9 gene induction by TPA (16,40), and our results demonstrated that
PJT inhibited MMP-9 expression by suppressing NF-κB in breat cancer
cells treated with TPA.
Our experiments confirmed that TPA-induced cell
invasion was suppressed by PJT. This was demonstrated by the
Matrigel invasion assay showing the inhibition of the TPA-induced
invasion potential of MCF-7 cells by PJT (Fig. 5).
In conclusion, our results demonstrated that PJT is
a potent inhibitor of TPA-induced MMP-9 expression and strongly
blocked the action of the NF-κB signaling pathway through PKCα in
MCF-7 cells. This is the first study, to the best of our knowledge,
to demonstrate that TPA-induced MCF-7 cell invasion was suppressed
by PJT through the inhibition of MMP-9 expression and through the
suppression of PKCα/NF-κB in MCF-7 cells. The findings of this
study suggest that PJT may potentially be developed into a potent
chemopreventive agent for the prevention of breast cancer
metastasis.
Acknowledgments
The present study was supported by a grant from
Wonkwang University in 2013.
References
|
1
|
Siegel R, Ma J, Zou Z and Jemal A: Cancer
statistics, 2014. CA Cancer J Clin. 64:9–29. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Chambers AF and Matrisian LM: Changing
views of the role of matrix metalloproteinases in metastasis. J
Natl Cancer Inst. 89:1260–1270. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nakajima M, Welch DR, Belloni PN and
Nicolson GL: Degradation of basement membrane type IV collagen and
lung subendothelial matrix by rat mammary adenocarcinoma cell
clones of differing metastatic potentials. Cancer Res.
47:4869–4876. 1987.PubMed/NCBI
|
|
4
|
Egeblad M and Werb Z: New functions for
the matrix metalloproteinases in cancer progression. Nat Rev
Cancer. 2:161–174. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
5
|
Woessner JF Jr: Matrix metalloproteinases
and their inhibitors in connective tissue remodeling. FASEB J.
5:2145–2154. 1991.PubMed/NCBI
|
|
6
|
Stetler-Stevenson WG, Hewitt R and
Corcoran M: Matrix metal-loproteinases and tumor invasion: from
correlation and causality to the clinic. Semin Cancer Biol.
7:147–154. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Itoh Y and Nagase H: Matrix
metalloproteinases in cancer. Essays Biochem. 38:21–36. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Brinckerhoff CE and Matrisian LM: Matrix
metalloproteinases: a tail of a frog that became a prince. Nat Rev
Mol Cell Biol. 3:207–214. 2002. View
Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gum R, Wang H, Lengyel E, Juarez J and
Boyd D: Regulation of 92 kDa type IV collagenase expression by the
jun aminoterminal kinase- and the extracellular signal-regulated
kinase-dependent signaling cascades. Oncogene. 14:1481–1493. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Newton AC: Regulation of protein kinase C.
Curr Opin Cell Biol. 9:161–167. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zeigler ME, Chi Y, Schmidt T and Varani J:
Role of ERK and JNK pathways in regulating cell motility and matrix
metalloproteinase 9 production in growth factor-stimulated human
epidermal keratinocytes. J Cell Physiol. 180:271–284. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Hozumi A, Nishimura Y, Nishiuma T, Kotani
Y and Yokoyama M: Induction of MMP-9 in normal human bronchial
epithelial cells by TNF-alpha via NF-kappa B-mediated pathway. Am J
Physiol Lung Cell Mol Physiol. 281:L1444–L1452. 2001.PubMed/NCBI
|
|
13
|
Weng CJ, Chau CF, Hsieh YS, Yang SF and
Yen GC: Lucidenic acid inhibits PMA-induced invasion of human
hepatoma cells through inactivating MAPK/ERK signal transduction
pathway and reducing binding activities of NF-kappaB and AP-1.
Carcinogenesis. 29:147–156. 2008. View Article : Google Scholar
|
|
14
|
Lin CW, Hou WC, Shen SC, Juan SH, Ko CH,
Wang LM and Chen YC: Quercetin inhibition of tumor invasion via
suppressing PKC delta/ERK/AP-1-dependent matrix metalloproteinase-9
activation in breast carcinoma cells. Carcinogenesis. 29:1807–1815.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Lee SO, Jeong YJ, Kim M, Kim CH and Lee
IS: Suppression of PMA-induced tumor cell invasion by capillarisin
via the inhibition of NF-kappaB-dependent MMP-9 expression. Biochem
Biophys Res Commun. 366:1019–1024. 2008. View Article : Google Scholar
|
|
16
|
Hong S, Park KK, Magae J, Ando K, Lee TS,
Kwon TK, Kwak JY, Kim CH and Chang YC: Ascochlorin inhibits matrix
metallopro-teinase-9 expression by suppressing activator
protein-1-mediated gene expression through the ERK1/2 signaling
pathway: inhibitory effects of ascochlorin on the invasion of renal
carcinoma cells. J Biol Chem. 280:25202–25209. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Woo MS, Jung SH, Kim SY, Hyun JW, Ko KH,
Kim WK and Kim HS: Curcumin suppresses phorbol ester-induced matrix
metalloproteinase-9 expression by inhibiting the PKC to MAPK
signaling pathways in human astroglioma cells. Biochem Biophys Res
Commun. 335:1017–1025. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Eberhardt W, Huwiler A, Beck KF, Walpen S
and Pfeilschifter J: Amplification of IL-1 beta-induced matrix
metalloproteinase-9 expression by superoxide in rat glomerular
mesangial cells is mediated by increased activities of NF-kappa B
and activating protein-1 and involves activation of the
mitogen-activated protein kinase pathways. J Immunol.
165:5788–5797. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim HR, Kim JM, Kim MS, Hwang JK, Park YJ,
Yang SH, Kim HJ, Ryu DG, Lee DS, Oh H, et al: Saussurea lappa
extract suppresses TPA-induced cell invasion via inhibition of
NF-κB-dependent MMP-9 expression in MCF-7 breast cancer cells. BMC
Complement Altern Med. 14:1702014. View Article : Google Scholar
|
|
20
|
Hisamoto M, Kikuzaki H, Ohigashi H and
Nakatani N: Antioxidant compounds from the leaves of Peucedanum
japonicum thunb. J Agric Food Chem. 51:5255–5261. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hisamoto M, Kikuzaki H and Nakatani N:
Constituents of the leaves of Peucedanum japonicum Thunb. and their
biological activity. J Agric Food Chem. 52:445–450. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Nugara RN, Inafuku M, Iwasaki H and Oku H:
Partially purified Peucedanum japonicum Thunb extracts exert
anti-obesity effects in vitro. Nutrition. 30:575–583. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Chen IS, Chang CT, Sheen WS, Teng CM, Tsai
IL, Duh CY and Ko FN: Coumarins and antiplatelet aggregation
constituents from formosan Peucedanum japonicum. Phytochemistry.
41:525–530. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chiu WT, Shen SC, Chow JM, Lin CW, Shia LT
and Chen YC: Contribution of reactive oxygen species to
migration/invasion of human glioblastoma cells U87 via
ERK-dependent COX-2/PGE(2) activation. Neurobiol Dis. 37:118–129.
2010. View Article : Google Scholar
|
|
25
|
Noh EM, Park YJ, Kim JM, Kim MS, Kim HR,
Song HK, Hong OY, So HS, Yang SH, Kim JS, et al: Fisetin regulates
TPA-induced breast cell invasion by suppressing matrix
metalloproteinase-9 activation via the PKC/ROS/MAPK pathways. Eur J
Pharmacol. 764:79–86. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Bradford MM: A rapid and sensitive method
for the quantitation of microgram quantities of protein utilizing
the principle of protein-dye binding. Anal Biochem. 72:248–254.
1976. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Lee YR, Noh EM, Oh HJ, Hur H, Kim JM, Han
JH, Hwang JK, Park BH, Park JW, Youn HJ, et al:
Dihydroavenanthramide D inhibits human breast cancer cell invasion
through suppression of MMP-9 expression. Biochem Biophys Res
Commun. 405:552–557. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zwiefel K and Janni W: Current standards
in the treatment of breast cancer. Med Monatsschr Pharm.
34:280–288. 2011.In German.
|
|
29
|
Deryugina EI and Quigley JP: Matrix
metalloproteinases and tumor metastasis. Cancer Metastasis Rev.
25:9–34. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Scorilas A, Karameris A, Arnogiannaki N,
Ardavanis A, Bassilopoulos P, Trangas T and Talieri M:
Overexpression of matrix-metalloproteinase-9 in human breast
cancer: a potential favourable indicator in node-negative patients.
Br J Cancer. 84:1488–1496. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Cho HJ, Kang JH, Kwak JY, Lee TS, Lee IS,
Park NG, Nakajima H, Magae J and Chang YC: Ascofuranone suppresses
PMA-mediated matrix metalloproteinase-9 gene activation through the
Ras/Raf/MEK/ERK- and Ap1-dependent mechanisms. Carcinogenesis.
28:1104–1110. 2007. View Article : Google Scholar
|
|
32
|
Mo N, Li ZQ, Li J and Cao YD: Curcumin
inhibits TGF-β1-induced MMP-9 and invasion through ERK and Smad
signaling in breast cancer MDA- MB-231 cells. Asian Pac J Cancer
Prev. 13:5709–5714. 2012. View Article : Google Scholar
|
|
33
|
Lungu G, Covaleda L, Mendes O,
Martini-Stoica H and Stoica G: FGF-1-induced matrix
metalloproteinase-9 expression in breast cancer cells is mediated
by increased activities of NF-kappaB and activating protein-1. Mol
Carcinog. 47:424–435. 2008. View Article : Google Scholar
|
|
34
|
Stoll BA: Biological mechanisms in breast
cancer invasiveness: relevance to preventive interventions. Eur J
Cancer Prev. 9:73–79. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Fortino V, Torricelli C, Capurro E, Sacchi
G, Valacchi G and Maioli E: Antiproliferative and survival
properties of PMA in MCF-7 breast cancer cell. Cancer Invest.
26:13–21. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kim JM, Noh EM, Kwon KB, Kim JS, You YO,
Hwang JK, Hwang BM, Kim BS, Lee SH, Lee SJ, et al: Curcumin
suppresses the TPA-induced invasion through inhibition of
PKCα-dependent MMP-expression in MCF-7 human breast cancer cells.
Phytomedicine. 19:1085–1092. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Lallena MJ, Diaz-Meco MT, Bren G, Payá CV
and Moscat J: Activation of IkappaB kinase beta by protein kinase C
isoforms. Mol Cell Biol. 19:2180–2188. 1999. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Sun CM, Syu WJ, Don MJ, Lu JJ and Lee GH:
Cytotoxic sesquiterpene lactones from the root of Saussurea lappa.
J Nat Prod. 66:1175–1180. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Thanos D and Maniatis T: NF-kappa B: a
lesson in family values. Cell. 80:529–532. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Oh JH, Chung AS, Steinbrenner H, Sies H
and Brenneisen P: Thioredoxin secreted upon ultraviolet A
irradiation modulates activities of matrix metalloproteinase-2 and
tissue inhibitor of metalloproteinase-2 in human dermal
fibroblasts. Arch Biochem Biophys. 423:218–226. 2004. View Article : Google Scholar : PubMed/NCBI
|