Introduction
Vascular injury induces cellular responses which
lead to clinical events, including atherosclerosis, hypertension
and restenosis (1). One common
feature of vascular lesions is the migration of vascular smooth
muscle cells (VSMCs) (2). VSMCs
are mostly found in the media layer of normal arteries; however,
VSMCs migrate from the media towards the intima during the
development of atherosclerosis or vascular damage (3,4). A
number of factors, including matrix metalloproteinases (MMPs),
growth factors, cytokines and chemokines affect VSMC migration in
the microenvironment of atherosclerotic lesions (5). Therefore, the inhibition of VSMC
migration represents a potentially important therapeutic strategy
for the treatment of cardiovascular diseases.
Platelet-derived growth factor (PDGF), a potent
chemoattractant for VSMCs, is encoded by 4 genes: PDGF-A, -B, -C
and -D (6). Among these, only
PDGF-BB can bind to all homo- or heterodimeric PDGF receptors
(PDGF-Rs) (7). The binding of
PDGF-BB to PDGF-R activates various downstream signaling molecules,
including those in the phosphatidylinositol 3 kinase (PI3K)/Akt,
phospholipase C (PLC)-γ1, and extracellular signal-regulated kinase
(ERK)1/2 pathways (1,8,9).
Akt is a downstream target of PI3K and plays a pivotal role in cell
migration, growth and anti-apoptotic events in various cell types
(10,11). The mitogen-activated protein
kinase (MAPK) signaling pathway also plays an important role in the
regulation of the proliferation, migration and survival of
mammalian cells (8,12). Consequently, the upregulation of
PDGF signaling can cause the development and progression of
cardiovascular diseases, such as hypertension and atherosclerosis
(13). Therefore, modulating the
PDGF signaling pathway in VSMCs may be a favorable pharmacological
strategy for the prevention of atherosclerosis.
Various herbs and plants have traditionally been
used in oriental folk medicine for the treatment of various
diseases. The bark of Magnolia obovata Thunb. (Magnoliaceae;
M. obovata), also known as Hu-Bak, is one of the most
popular therapeutics in traditional Korean medicine and is widely
used to treat fever, headaches, diarrhea, anxiety and to relieve
asthma (14,15). It contains many types of secondary
metabolites, such as neolignans, sesquiterpene-neolignans,
aporphine alkaloids and essential oils (16,17). These compounds show anti-gastric
ulcer, anti-platelet, cytotoxic, anti-inflammatory, and
anti-complement activities (18,19). Recently, magnobovatol, a newly
identified neolignan from the fruits of M. obovata, has been
reported to inhibit the growth of a variety of human cancer cells,
including breast, colorectal, melanoma cancer cell lines (20). However, the effects of
magnobovatol on smooth muscle cell migration have not yet been
elucidated, at least to the best of our knowledge. Thus, the aim of
this study was to elucidate the anti-migratory activity and the
mechanistic target of magnobovatol in PDGF-BB-stimulated VSMCs.
Materials and methods
Cell culture
Human aortic smooth muscle cells (HASMCs) were
purchased from Applied Biological Materials Inc. (Cat. no. T4050)
and were maintained in Prigrow III medium (Applied Biological
Materials Inc.) (Cat. no. TM003) in the presence of 5% FBS (Cat.
no. SH30919.03; HyClone, Logan, UT, USA) and penicillin (100
U/ml)/streptomycin (100 μg/ml) (Invitrogen, Carlsbad, CA,
USA). MOVAS-1 murine primary aortic VSMCs were obtained from the
American Type Culture Collection (ATCC; Cat. no CRL-2797™) and were
cultured in Dulbecco's modified Eagle's medium (DMEM; Invitrogen)
supplemented with 10% FBS, G418 (0.2 mg/ml), penicillin (100
U/ml)/streptomycin (100 μg/ml) at 37°C with 5%
CO2 atmosphere in a humidified incubator.
Reagents
Magnobovatol was kindly provided by Dr Nam-In Baek
(Department of Oriental Medicine Biotechnology, Kyung Hee
University, Yongin, Korea) (16).
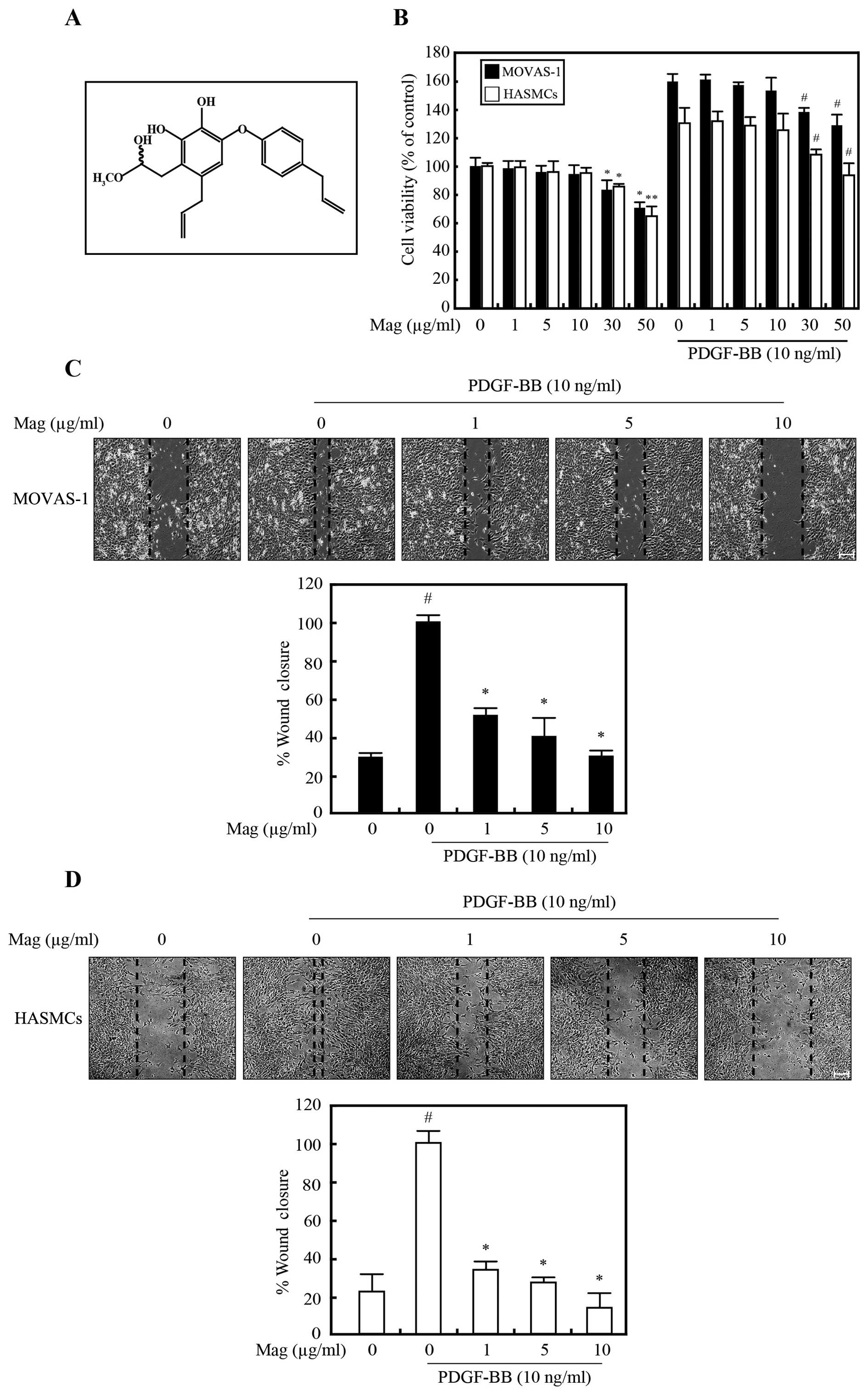
The structure of magnobovatol is illustrated in Fig. 1A.
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
was from Sigma (St. Louis, MO, USA). PDGF-BB was obtained from
Peprotech (Seoul, Korea). Gelatin was obtained from Difco
(Lexington, KY, USA). Lipofectamine 2000 reagent was purchased from
Invitrogen. Anti-p-PDGFRβ (Cat. no. 4549) and anti-p-Akt (Ser473;
Cat. no. 4058) antibodies were obtained from Cell Signaling
Technology, Inc. (Beverly, MA, USA). Anti-p-ERK (Cat. no. sc-7383),
anti-ERK (Cat. no. sc-93), anti-Akt (Cat. no. sc-5298) and
anti-α-tubulin (Cat. no. sc-5286) antibodies were from Santa Cruz
Biotechnology, Inc. (Santa Cruz, CA, USA). All the chemicals not
mentioned above were obtained from Sigma.
Cell viability
Cell viability was measured by MTT assay. The cells
were seeded in 96-well plates at a density of 2×105
cells/well. Following serum starvation for 24 h, the cells were
treated with PDGF-BB (10 ng/ml) in the presence of 1–50
μg/ml magnobovatol for 30 min. Following incubation for 24 h
at 37°C, 20 μl of 5 mg/ml MTT solution were added and the
cells were incubated for 1 h at 37°C and 100 μl dimethyl
sulfoxide was added to each well to dissolve the formazan. The
absorbance was measured at 550 nm using a microplate reader
(Magellan; Tecan). Cell viability is expressed as a percentage of
the absorbance value determined for the control cultures.
Western blot analysis
To examine the protein expression patterns, equal
amounts (15 μg) of total protein extracts were prepared. The
cell lysates were seperated on 9% SDS-polyacrylamide gels and
transferred onto nitrocellulose membranes. The blots were then
incubated with the antibodies (anti-p-PDGF-R, anti-p-ERK, anti-ERK,
anti-p-Akt and anti-Akt antibodies) and detected using the enhanced
chemiluminescence detection system (Amersham Pharmacia Biotech,
Piscataway, NJ, USA). The same blots were then stripped and
reprobed with anti-tubulin antibody for use as an internal control.
Quantitative analysis of the results of western blot analysis were
carried out using ImageJ software.
Wound healing assay
The cells were incubated until they reached 90 to
100% confluence in 12-well plates. Subsequently, a scratch was
gently made using a P-10 pipette tip, and the cells were then
treated with 1–10 μg/ml magnobovatol with PDGF-BB. The cells
were subsequently allowed to migrate [24 h (MOVAS-1) or 48 h
(HASMCs)] and incubated for various periods of time and images were
acquired. Phase contrast images were acquired using a Nikon
microscopy system (Nikon Instruments Inc., Melville, NY, USA). The
measurement of the wound healing gap distance was performed using
ImageJ software.
RT-PCR
Total RNA was extracted from the cells using TRIzol
reagent (Invitrogen) according to the manufacturer's instructions.
Approximately 2 μg of total RNA was used to prepare cDNA
using the SuperScript First Strand cDNA Synthesis kit (Bioneer
Corporation, Daejeon, South Korea). The following primers were used
in this study: mMMP-2 forward, 5′-AAGGATGGACTCCTGGCACATGCCTTT-3′
and reverse, 5′-ACCTGTGGGCTTGTCACGTGGTGT-3′; mGAPDH forward,
5′-GGAGCCAAAAGGGTCATCAT-3′ and reverse, 5′-GTGATGGCATGGACTGTGGT-3′;
hMMP-2 forward, 5′-ATGACAGCTGCACCACTGAG-3′ and reverse,
5′-ATTTGTTGCCCAGGAAAGTG-3′; and hGAPDH forward,
5′-CCATCACCATCTTCCAGGAG-3′ and reverse, 5′-CCTGCTTCACCACGTTCTTG-3′.
PCR was performed with Platinum Taq polymerase (Invitrogen) under
the following conditions: 30 cycles of 96°C for 40 sec, 55°C
(MMP-2) or 60°C (GAPDH) for 40 sec, 72°C for 1 min followed by 10
min at 72°C. The PCR products were electrophoresed on a 2% (w/v)
agarose gel in 1X Tris-acetate-EDTA (TAE) buffer, and stained with
ethidium bromide solution. All the PCR reactions were repeated at
least 3 times. GAPDH was amplified as an internal control. The
intensity of each band amplified by RT-PCR was analyzed using
MultiImage™ Light Cabinet (version 5.5; Alpha Innotech Corp., San
Leandro, CA, USA), and normalized to that of GAPDH mRNA in
corresponding samples.
Gelatin zymography
The presence of MMP-2 in the super-natants of
magnobovatol- and/or PDGF-BB-treated SMCs were analyzed using
gelatin zymograms. Briefly, the cells were incubated in serum-free
DMEM and the supernatants were collected following incubation for
24 h at 37°C, clarified by centrifugation (13,000 rpm for 5 min at
4°C), normalized to the total protein concentration of the cell
lysate, mixed with non-reducing Laemmli smaple buffer, and
separated by electrophoresis on 10% SDS-PAGE gels containing 1
mg/ml gelatin (Difco). Following electrophoresis, the gels were
re-natured by washing in 2.5% Triton X-100 solution twice for 30
min to remove all the SDS. The gels were then incubated in 50
mmol/l Tris-HCl (pH 7.4), 5 mmol/l CaCl2, and 1
μM ZnCl2 at 37°C overnight. Following incubation,
the gels were stained with 0.05% Coomassie brilliant blue R-250 for
30 min at room temperature and then destained in distilled water.
MMP-2 activities were visible as clear bands on a blue background
where the gelatin substrate had been hydrolyzed by enzyme
activity.
ELISA for MMP-2
The supernatants were collected for measuring the
amount of secreted MMP-2 protein. The total MMP-2 protein was
assayed according to the instructions provided with the Quantkine
ELISA kit (R&D Systems, Inc., Minneapolis, MN, USA). In brief,
samples and MMP-2 standards were added to microplates pre-coated
with antibody specifically recognizing both the pro- and active
forms of MMP-2. After washing, bound MMP-2 was measured using a
horseradish peroxidase-conjugated secondary anti-MMP-2 antibody,
developed with hydrogen peroxide and tetramethylbenzidine. The
optical density was measured at 450 nm using a Bio-Rad Model 550
microplate reader and associated Microplate Manager software
(Bio-Rad Laboratories, Mississauga, ON, Canada). The absorbance of
the samples was measured at 450 nm using a microplate reader.
Aortic ring assay
The ex vivo migration and proliferation of
VSMCs were measured by aortic ring assay using Matrigel. Male
C57BL/6J mice (n=6 in each group), which were 6–8 weeks of age,
were purchased from Orient Bio, Inc. (Seoul, Korea) and housed
under special pathogen-free conditions. All animals were treated in
accordance with the Animal Care Guidelines of Use of Laboratory
Animals and approved by the Laboratory of Animal Research in Asan
institute of Life Sciences. For the ex vivo migration and
proliferation of VSMCs, thoracic aortas were removed from the mice
which were sacrificed by prolonged exposure to a dose of isoflurane
followed by cervical dislocation and immediately transferred to a
culture dish containing serum-free DMEM. The peri-aortic
fibroadipose tissue was carefully removed with fine microdissecting
forceps and iridectomy scissors, paying special attention not to
damage the aortic wall. One millimeter-thick aortic rings,
approximately 12 per aorta, were sectioned and extensively rinsed
in 5 consecutive washes of serum-free DMEM. These rings were placed
and embedded in 48-well plates coated with Matrigel (BD
Biosciences, Franklin Lakes, NJ, USA). Simultaneously, magnobovatol
(0–10 μg/ml) and PDGF-BB (10 ng/ml) were added to the
culture medium for 3 days. The ring formation images were acquired
using a ZEISS microscope (Carl Zeiss, Oberkochen, Germany) and the
length of the sprouts was analyzed.
All animal procedures were reviewed and approved by
the Institutional Animal Care and Use Committee (IACUC) of
University of Ulsan College of Medicine (approval no. 2014-12-140)
and were performed in strict accordance with the Association for
Assessment and Accreditation of Laboratory Animal Care and the NIH
guidelines (Guide for the Care and Use of Laboratory Animals).
Statistical analysis
Statistical analysis was performed using the
computer program Prism (GraphPad Software, Inc., La Jolla, CA,
USA). The results are presented as the means ± SE. The statistical
significance of the differences between groups was analyzed by
repeated measures of one-way analysis of variance followed by a
Student's t-test. A P-value <0.05 was considered to indicate a
statistically significant difference.
Results
Magnobovatol inhibits PDGF-BB-induced
VSMC migration
To assess the effects of magnobovatol on cell
proliferation, mouse aortic smooth muscle cells (MOVAS-1 cells) and
HASMCs were treated with magnobovatol in the absence or presence of
10 ng/ml PDGF-BB for 24 h. Compared to the untreated control cells,
the MOVAS-1 cells (Fig. 1B, black
bar) and the HASMCs (Fig. 1B,
white bar) treated with magnobovatol at concentrations between 0
and 10 μg/ml exhibited no signs of cytotoxicity, regardless
of whether they were treated with PDGF-BB or not. Therefore, this
concentration range was used in all the subsequent experiments.
To determine the effects of magnobovatol on VSMC
migration, we performed a wound healing assay using both the HASMCs
and MOVAS-1 cells. After a scratch wound was made on the cells in
the plates, the effects of magnobovatol various concentrations on
PDGF-BB-induced cell migration were examined. The cells were
allowed to migrate for 24 h (MOVAS-1) or 48 h (HASMCs), and the
migration distances were then measured. As shown in Fig. 1C and D, magnobovatol suppressed
the PDGF-BB-induced migration of both the HASMCs (Fig. 1C) and the MOVAS-1 cells (Fig. 1D) in a dose-dependent manner,
demonstrating that magnobovatol suppressed VSMC migration.
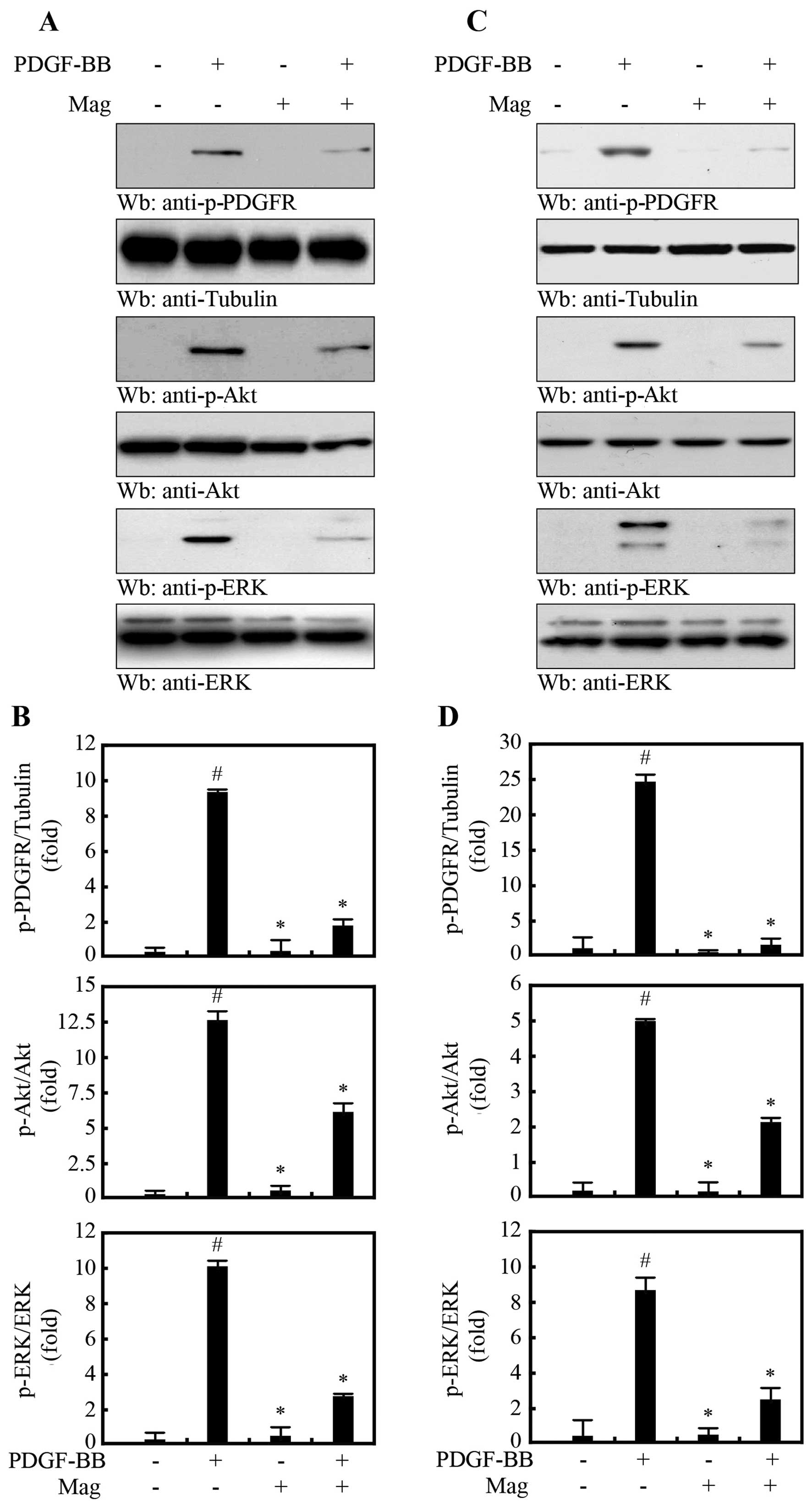
Magnobovatol inhibits the activation of
PDGF-R signaling and the ERK1/2 and Akt signaling pathways
PDGF triggers the PI3K/Akt and Ras/MAPK signaling
cascades by activating PDGF-R (8). Thus, in this study, to determine
whether magnobovatol inhibits PDGF-induced mitogenic effects, we
treated the HASMCs (Fig. 2A and
B) and MOVAS-1 cells (Fig. 2C and
D) with 10 μg/ml magnobovatol followed by treatment with
PDGF-BB. PDGF-BB markedly induced the phosphorylation of PDGF-R,
which was substantially attenuated by treatment with magnobovatol
(Fig. 2). In addition,
magnobovatol inhibited PDGF-induced ERK1/2 and Akt phosphorylation
in both the HASMCs (Fig. 2A and
B) and MOVAS-1 cells (Fig. 2C and
D). These results indicate that magnobovatol regulates VSMC
migration through the inhibition of PDGF-R activation.
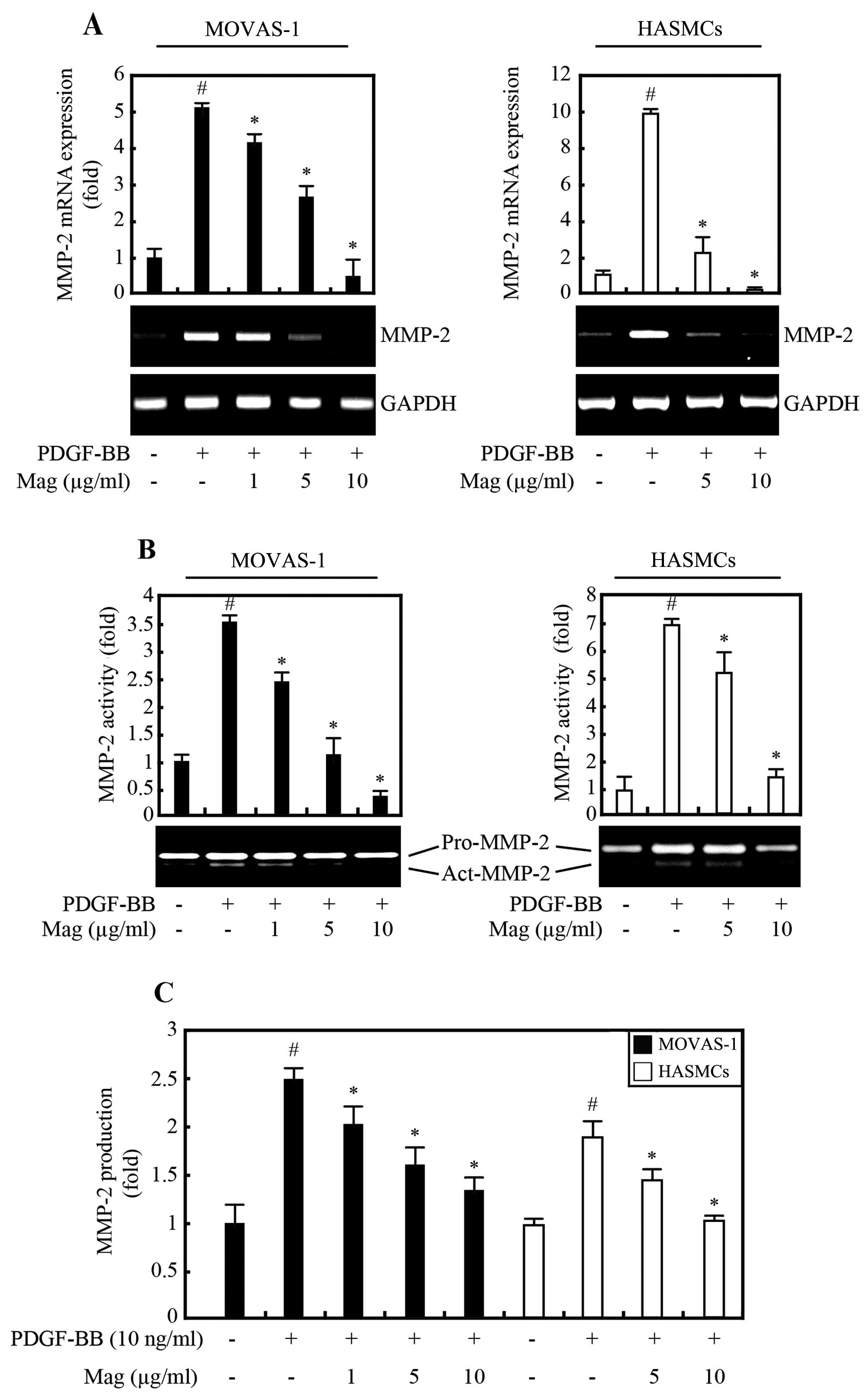
Magnobovatol suppresses PDGF-BB-induced
MMP-2 expression and proteolytic activity
MMPs involved in the breakdown of the extracellular
matrix (ECM) are associated with the development of atherosclerosis
(21). In particular, the
gelatinase MMP-2 is predominantly expressed in VSMCs in advanced
atherosclerotic lesions and in injury-induced neointimal lesions
(22). Since PDGF is a potent
inducer of MMP-2 (23), in this
study, we examined the effects of magnobovatol on the expression of
MMP-2 in VSMCs. Our results revealed that the basal levels of MMP-2
expression and activity were low in the both MOVAS-1 cells and
HASMCs, as indicated by RT-PCR (Fig.
3A), gelatin zymography (Fig.
3B), and ELISA (Fig. 3C).
However, MMP-2 mRNA expression and protein secretion were markedly
increased by stimulation with PDGF-BB (Fig. 3). Of note, magnobovatol inhibited
the increase in both MMP-2 expression and secretion, but it had no
effect on the expression of MMP-9 (data not shown). These results
suggested that the PDGF-BB-induced increase in MMP-2 expression and
secretion in VSMCs are inhibited by magnobovatol.
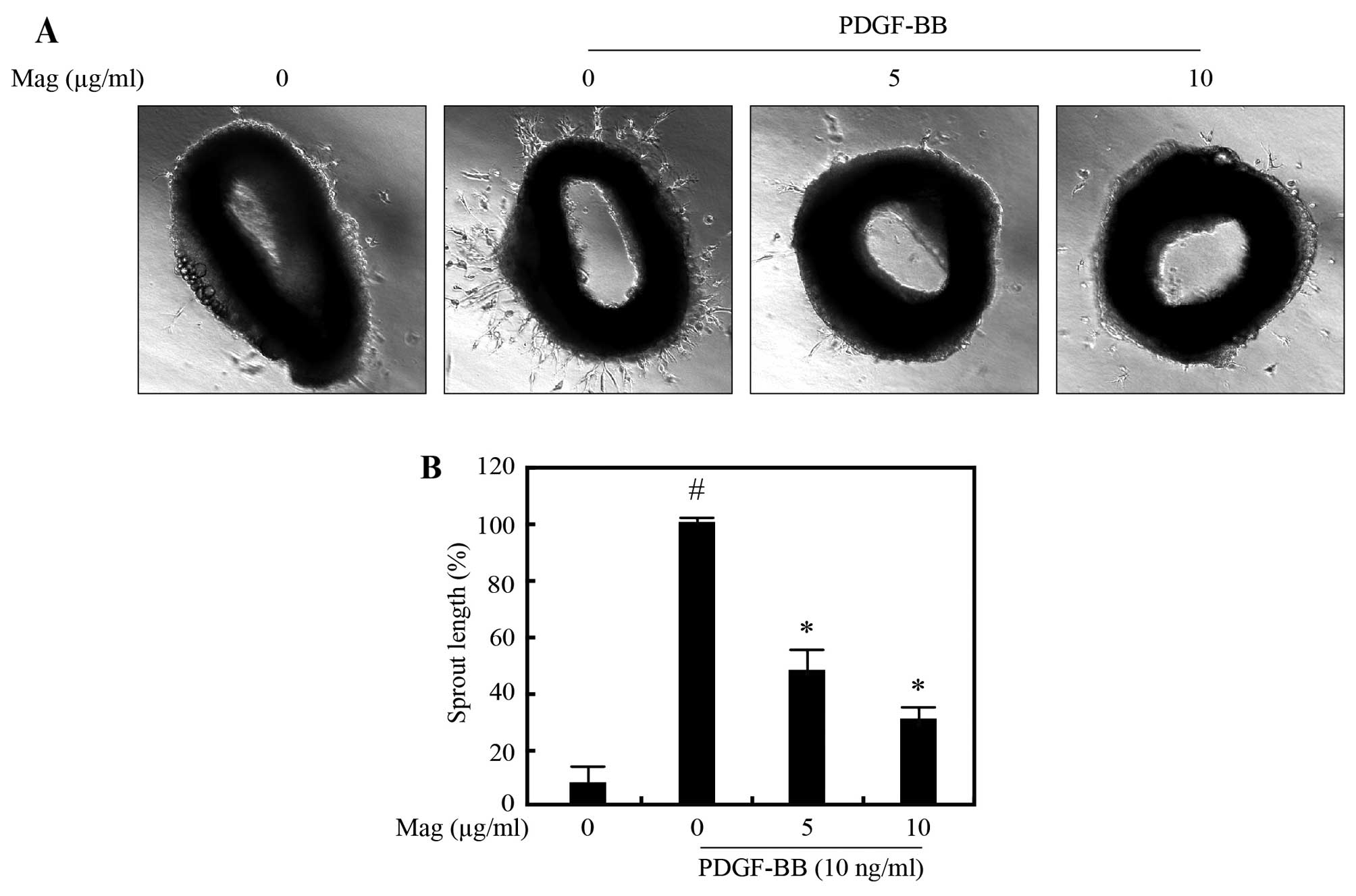
Magnobovatol attenuates PDGF-BB-induced
VSMC sprout outgrowth formation of aortic rings
To examine the effects of magnobovatol on VSMC
migration in an animal model of atherosclerosis, we performed an
ex vivo aortic ring assay. Aortic rings isolated from
C57BL/6 mice were embedded in Matrigel and the lengths of outgrowth
sprouts were measured after 3 days. As shown in Fig. 4, exposure to PDGF-BB (10 ng/ml)
increased sprouting, which was significantly attenuated by
magnobovatol treatment in a dose-dependent manner. These results
suggest that magnobovatol markedly attenuates PDGF-BB-induced VSMC
sprout outgrowth formation ex vivo.
Discussion
In the present study, we demonstrated the following:
i) magnobovatol inhibits PDGF-induced VSMC migration, ii)
magnobovatol inhibits the activation of PDGF-R downstream signaling
pathways, iii) magnobovatol regulates MMP-2 expression and
activity, and iv) consequently, magnobovatol inhibits PDGF-induced
migration through the suppression of PDGF-R-mediated MMP-2
expression and proteolytic activity in VSMCs.
PDGF acts as a potent inducer of VSMC proliferation
and migration through the phosphorylation of PDGF-R (24). Enhanced PDGF signaling has been
implicated in the pathophysiology of a variety of diseases
involving vascular wall remodeling (13,25). In the arteries, PDGF-R expression
is substantially induced following endothelial injury by
angioplasty or early-stage atherosclerosis (26). Conversely, inhibiting the PDGF
pathway using antisense oligonucleotides or a blocking antibody has
been successfully applied to suppress intimal thickening (27). Treatment with PDGF has previously
been demonstrated to induce the phosphorylation of Akt and ERK1/2
in both HASMCs and MOVAS-1 cells (4). In accordance with these findings, we
found that magnobovatol significantly attenuated PDGF-BB-induced
PDGF-R, Akt and ERK1/2 phosphorylation in VSMCs. These results
suggest that PDGF-R may be a direct target of magnobovatol in the
inhibition of VSMC migration. Furthermore, PI3K/Akt contributes to
ECM destruction by increasing the production of MMP-2 in VSMCs
(28). MMP-2 expression is
associated with the induction of SMC hyperplasia during
atherosclerosis and restenosis. In addition, MMP-2 plays a role in
the proliferation and migration of SMCs, which contributes to the
intimal thickening of vascular lesions in vivo (29,30). Our results revealed that
pre-treatment with magnobovatol inhibited the PDGF-induced increase
in MMP-2 expression and proteolytic activity in a dose-dependent
manner. Presumably, the suppressive effects of magnobovatol on the
PDGF-induced activation of Akt was associated with the subsequent
decrease in the mRNA expression and secretion of MMP-2.
In conclusion, the results of our study demonstrated
showed that magnobovatol inhibited the migration of cultured VSMCs,
which may be major inducers of atherosclerosis and restenosis,
in vitro. We demonstrate that magnobovatol regulates
molecules involved in the remodeling of the vessel wall and/or the
accumulation of cells and ECM. Therefore, our data may provide a
novel strategy with which to inhibit VSMC migration under
pathological conditions, thereby offering a novel therapeutic
approach for the treatment of restenosis and other vascular
diseases involving VSMC migration.
Acknowledgments
The present study was supported by the Basic Science
Research Program through the National Research Foundation of Korea
(NRF) funded by the Ministry of Education (NRF-2013R1A1A2008633)
and the Asan Institute for Life Sciences, Seoul, Korea (2014-570)
and the Student Research Grant (14–15) of the University of Ulsan College
of Medicine, Seoul, Korea.
References
|
1
|
Cidad P, Miguel-Velado E, Ruiz-McDavitt C,
Alonso E, Jiménez-Pérez L, Asuaje A, Carmona Y, García-Arribas D,
López J, Marroquín Y, et al: Kv1.3 channels modulate human vascular
smooth muscle cells proliferation independently of mTOR signaling
pathway. Pflugers Arch. 467:1711–1722. 2015. View Article : Google Scholar
|
|
2
|
Owens GK, Kumar MS and Wamhoff BR:
Molecular regulation of vascular smooth muscle cell differentiation
in development and disease. Physiol Rev. 84:767–801. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Newby AC and Zaltsman AB: Molecular
mechanisms in intimal hyperplasia. J Pathol. 190:300–309. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Hua Y, Dolence J, Ramanan S, Ren J and
Nair S: Bisdemethoxy-curcumin inhibits PDGF-induced vascular smooth
muscle cell motility and proliferation. Mol Nutr Food Res.
57:1611–1618. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Rudijanto A: The role of vascular smooth
muscle cells on the pathogenesis of atherosclerosis. Acta Med
Indones. 39:86–93. 2007.PubMed/NCBI
|
|
6
|
Heldin CH, Eriksson U and Ostman A: New
members of the platelet-derived growth factor family of mitogens.
Arch Biochem Biophys. 398:284–290. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Fredriksson L, Li H and Eriksson U: The
PDGF family: four gene products form five dimeric isoforms.
Cytokine Growth Factor Rev. 15:197–204. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Heldin CH and Westermark B: Mechanism of
action and in vivo role of platelet-derived growth factor. Physiol
Rev. 79:1283–1316. 1999.PubMed/NCBI
|
|
9
|
Millette E, Rauch BH, Kenagy RD, Daum G
and Clowes AW: Platelet-derived growth factor-BB transactivates the
fibroblast growth factor receptor to induce proliferation in human
smooth muscle cells. Trends Cardiovasc Med. 16:25–28. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Muto A, Fitzgerald TN, Pimiento JM,
Maloney SP, Teso D, Paszkowiak JJ, Westvik TS, Kudo FA, Nishibe T
and Dardik A: Smooth muscle cell signal transduction: implications
of vascular biology for vascular surgeons. J Vasc Surg. (Suppl A):
A15–A24. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Seo J, Lee HS, Ryoo S, Seo JH, Min BS and
Lee JH: Tangeretin, a citrus flavonoid, inhibits PGDF-BB-induced
proliferation and migration of aortic smooth muscle cells by
blocking AKT activation. Eur J Pharmacol. 673:56–64. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Zhan Y, Kim S, Izumi Y, Izumiya Y, Nakao
T, Miyazaki H and Iwao H: Role of JNK, p38, and ERK in
platelet-derived growth factor-induced vascular proliferation,
migration, and gene expression. Arterioscler Thromb Vasc Biol.
23:795–801. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Raines EW: PDGF and cardiovascular
disease. Cytokine Growth Factor Rev. 15:237–254. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Ho JW and Jie M: Pharmacological activity
of cardiovascular agents from herbal medicine. Cardiovasc Hematol
Agents Med Chem. 5:273–277. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Amblard F, Govindarajan B, Lefkove B, Rapp
KL, Detorio M, Arbiser JL and Schinazi RF: Synthesis, cytotoxicity,
and antiviral activities of new neolignans related to honokiol and
magnolol. Bioorg Med Chem Lett. 17:4428–4431. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Seo KH, Lee DY, Lee DS, Park JH, Jeong RH,
Jung YJ, Shrestha S, Chung IS, Kim GS, Kim YC and Baek NI:
Neolignans from the fruits of Magnolia obovata and their inhibition
effect on NO production in LPS-induced RAW 264.7 cells. Planta Med.
79:1335–1340. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Pyo MK, Yun-Choi HS and Hong YJ:
Antiplatelet activities of aporphine alkaloids isolated from leaves
of Magnolia obovata. Planta Med. 69:267–269. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Pyo MK, Lee Y and Yun-Choi HS:
Anti-platelet effect of the constituents isolated from the barks
and fruits of Magnolia obovata. Arch Pharm Res. 25:325–328. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Youn U, Chen QC, Lee IS, Kim H, Yoo JK,
Lee J, Na M, Min BS and Bae K: Two new lignans from the stem bark
of Magnolia obovata and their cytotoxic activity. Chem Pharm Bull
(Tokyo). 56:115–117. 2008. View Article : Google Scholar
|
|
20
|
Seo KH, Lee DY, Jeong RH, Yoo KH, Chung
IS, Kim GS, Seo WD, Kang HC, Ahn EM and Baek NI: Cytotoxicity of
Neolignans from Magnolia obovata Fruits. J Appl Biol Chem.
56:179–181. 2013. View Article : Google Scholar
|
|
21
|
Newby AC: Matrix metalloproteinases
regulate migration, proliferation, and death of vascular smooth
muscle cells by degrading matrix and non-matrix substrates.
Cardiovasc Res. 69:614–624. 2006. View Article : Google Scholar
|
|
22
|
Johnson C and Galis ZS: Matrix
metalloproteinase-2 and -9 differentially regulate smooth muscle
cell migration and cell-mediated collagen organization.
Arterioscler Thromb Vasc Biol. 24:54–60. 2004. View Article : Google Scholar
|
|
23
|
Risinger GM Jr, Updike DL, Bullen EC,
Tomasek JJ and Howard EW: TGF-beta suppresses the upregulation of
MMP-2 by vascular smooth muscle cells in response to PDGF-BB. Am J
Physiol Cell Physiol. 298:C191–C201. 2010. View Article : Google Scholar
|
|
24
|
Bornfeldt KE, Raines EW, Nakano T, Graves
LM, Krebs EG and Ross R: Insulin-like growth factor-I and
platelet-derived growth factor-BB induce directed migration of
human arterial smooth muscle cells via signaling pathways that are
distinct from those of proliferation. J Clin Invest. 93:1266–1274.
1994. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Ross R: The pathogenesis of
atherosclerosis: a perspective for the 1990s. Nature. 362:801–809.
1993. View
Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ferns GA, Raines EW, Sprugel KH, Motani
AS, Reidy MA and Ross R: Inhibition of neointimal smooth muscle
accumulation after angioplasty by an antibody to PDGF. Science.
253:1129–1132. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Sirois MG, Simons M and Edelman ER:
Antisense oligonucleotide inhibition of PDGFR-beta receptor subunit
expression directs suppression of intimal thickening. Circulation.
95:669–676. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Risinger GM Jr, Hunt TS, Updike DL, Bullen
EC and Howard EW: Matrix metalloproteinase-2 expression by vascular
smooth muscle cells is mediated by both stimulatory and inhibitory
signals in response to growth factors. J Biol Chem.
281:25915–25925. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Beaudeux JL, Giral P, Bruckert E,
Foglietti MJ and Chapman MJ: Matrix metalloproteinases and
atherosclerosis. Therapeutic aspects. Ann Biol Clin (Paris).
61:147–158. 2003.In French.
|
|
30
|
Kuzuya M and Iguchi A: Role of matrix
metalloproteinases in vascular remodeling. J Atheroscler Thromb.
10:275–282. 2003. View Article : Google Scholar
|