Introduction
The therapeutic use of cold atmospheric plasma (CAP)
constitutes an emerging interdisciplinary field that capitalizes on
the rapidly evolving technology of low-temperature plasma (1–4).
Plasma is an at least partially ionized gas and is defined as the
fourth state of matter. It is considered to be a mixture of
electrons, negative and positive ions, excited gas species, free
radicals and electromagnetic radiation. The properties of CAP can
be modified by changing various experimental conditions, such as
the types of set-ups, the voltage applied, the type of feed gas and
the gas flow rate (3–6). It has been demonstrated that CAP is
useful in potential applications, such as sterilization (7), wound healing (8), dentistry (9) and tissue regeneration (10). In addition, a recent medical
investigation focused on applying CAP to the treatment of cancer
(11).
CAP has been reported to effectively suppress cancer
cell growth in in vivo experimental models (12–14). There is also growing evidence that
the exposure of cancer cells to CAP or CAP-activated medium induces
apoptosis, and reactive oxygen species (ROS) and/or reactive
nitrogen species (RNS) are considered to be effective agents for
CAP-induced apoptosis (12–22). In a previous study, CAP produced
dose-dependent effects, ranging from increased cell proliferation
to apoptosis; these effects were the result of the production of
ROS (15). Air-CAP generates a
variety of ROS and RNS, and these radicals induce an apoptotic
signaling cascade by inducing mitogen-activated protein kinase
(MAPK) signaling in cancer cells (17). Adachi et al indicated that
hydrogen peroxide (H2O2) and other reactive
agents in plasma-activated medium induce cell injury via the
mitochondria-nuclear apoptotic cascade in cancer cells (21). More recently, we demonstrated that
much higher levels of hydroxyl (·OH) radicals were
produced using argon (Ar)-CAP in an aqueous solution than by using
X-irradiation, based on the analysis and quantification of electron
paramagnetic resonance (EPR) spectra. Under this condition, Ar-CAP
produced intracellular ROS, ·OH radicals,
H2O2, which is the recombination product of
·OH, and hypochlorite ion (OCl−) in human
lymphoma U937 cells (22).
Although the generation of ROS induced by CAP and the biological
effects of the thus generated ROS have been clarified gradually,
the mechanisms of CAP-mediated RNS-cell interactions are not yet
fully understood. In this study, we investigated the additional
effects of nitrogen (N2) in Ar gas on the apoptosis of
and changes in gene expression in U937 cells exposed to CAP.
Materials and methods
Cell culture
Human myelomonocytic lymphoma U937 cells were
obtained from the Human Science Research Resources Bank of the
Japan Health Sciences Foundation (Tokyo, Japan). The cells were
cultured in RPMI-1640 medium (Wako Pure Chemical Industries, Ltd.,
Osaka, Japan) supplemented with 10% heat-inactivated fetal bovine
serum, and maintained at 37°C in humidified air with 5%
CO2 and 95% air.
Exposure to CAP
CAP was produced using our previously described
method (22). The inert gases, Ar
and N2, were purchased from Hokusan Co., Ltd. (Toyama,
Japan). All gases were of pure grade (≥99.9%). The U937 cells in a
well of a 24-well plate were exposed to Ar-CAP or Ar containing
2.5% of N2 (Ar + N2-CAP) generated using a 20
kHz low frequency at 18 kV with a flow rate of 2 l/min for 0 to 3
min at room temperature. Following exposure to CAP, the cells were
cultured for 0–18 h at 37°C in a CO2 incubator.
Non-treated cells served as controls.
Optical emissions from CAP
Optical emissions from Ar-CAP were collected using
an optic fiber and a lens directed to a position at 8 mm ahead of
CAP. To examine the effects of N2 on the optical
emissions of Ar-CAP, N2 was added to the Ar gas at flow
rates from 0 to 50 (standard cubic centimeter per minute). The
emission was observed using a spectrometer (Shamrock SR-561-B1) and
an intensified charge-coupled-device camera (iStar DH734-25F-03)
(both from Andor Technology, Ltd., Belfast, UK), as described in a
previous study of ours (22).
EPR-spin trapping for the detection of
hydroxyl radicals
The detection of ·OH radicals following
exposure to Ar-CAP or Ar + N2-CAP was carried out using
the EPR-spin trapping with 5,5-dimethyl-1-pyrroline-N-oxide (DMPO;
Labotec Co., Ltd., Tokyo, Japan). An aqueous solution containing a
spin trap at a concentration of 10 mM was exposed to CAP for up to
120 sec. Immediately following exposure, a sample was transferred
to a glass capillary tube (VC-HO75P; Terumo, Tokyo, Japan) which
was then inserted into a special quartz tube in the cavity of an
EPR spectrometer (RFR-30; Radical Research Inc., Tokyo, Japan). The
EPR settings were as follows: microwave power, 4 mW; frequency,
9.425 GHz; center magnetic field, 329.5 mT; and modulation width,
0.1 mT. The yields of spin adducts were determined using the stable
nitroxide radical
3-carbamoyl-2,2,5,5-tetramethyl-1-pyrroline-1-oxide as a standard
at room temperature. The peak heights of the EPR signals were
expressed in relative units compared with those of the
Mn2+ internal standard, with one unit being equivalent
to approximately 7.7×106 M nitroxide radicals (22).
Measurement of nitrite
(NO2)/nitrate (NO3)
The U937 cells were exposed to Ar-CAP or Ar +
N2-CAP for 3 min and then the supernatant of the cells
was collected by centrifugation. The concentration of total
NO2/NO3 in the supernatant of the cells was
measured using an NO2/NO3 assay kit-C II
(Dojindo Laboratories, Kumamoto, Japan) according to the
manufacturer's instructions.
Measurements of cell viability and
apoptosis
For measuring cell viability, we used a
water-soluble tetrazolium salt WST-8
[2-(2-methoxy-4-nitrophenyl)-3-(4-nitrophenyl)-5-(2,4-disulfo-phenyl)-2H-tetrazolium,
monosodium salt]-based assay (Cell Counting kit-8; Dojindo
Laboratories). In brief, the cells were incubated in 110 µl
RPMI-1640 medium containing 9.1% (v/v) of WST-8 reagent in a
96-well cell culture plate at 37°C. Two hours later, the produced
formazan dye concentration was determined from the absorbance at
450 nm using a microplate reader (23). The level of apoptosis was
determined using an Annexin V-FITC kit (Immunotech, Marseille,
France). Fluorescein isothiocyanate (FITC)-labeled Annexin V and
propidium iodide were added to the cell suspension. Following
incubation for 20 min in the dark, the cells were analyzed using a
flow cytometer (Epics XL; Beckman Coulter K.K., Tokyo, Japan).
Apoptosis was expressed as the sum of early apoptotic and secondary
necrotic fractions.
RNA isolation
For global-scale gene expression and quantitative
polymerase chain reaction (qPCR) analyses, total RNA was extracted
from the cells using a NucleoSpin® RNA isolation kit
(Macherey-Nagel GmbH & Co., Düren, Germany) along with
on-column DNase I treatment. RNA quality was analyzed using a
Bioanalyzer 2100 (Agilent Technologies, Inc., Santa Clara, CA,
USA).
Global-scale gene expression
analysis
Global-scale gene expression analysis was carried
out using a GeneChip® microarray system with a Human
Genome U133-plus 2.0 array, which was spotted with 54,675 probe
sets (Affymetrix Inc., Santa Clara, CA, USA) according to the
manufacturer's instructions. The obtained hybridization intensity
data were analyzed using GeneSpring® GX (Agilent
Technologies, Inc.) to extract the significant genes. To examine
gene ontology, including biological processes, cellular components,
molecular functions and gene networks, the obtained data were
analyzed using Ingenuity® Pathway Analysis tools
(Ingenuity Systems Inc., Mountain View, CA, USA), as previously
described (23,24).
Measurement of mRNA levels by qPCR
The mRNA levels in the cells were determined
following exposure to CAP for 2 min followed by culture at 37°C for
3 h using an Mx3005P real-time PCR system (Agilent Technologies,
Inc.) with using SYBR Premix Ex Taq or Premix Ex Taq (for the use
of TaqMan probes) (both from Takara Bio Inc., Shiga, Japan). The
specific primers and probes for BCL2-associated athanogene 3
(BAG3), DnaJ [heat shock protein (HSP)40] homolog, subfamily B,
member 1 (DNAJB1), early growth response 1 (ERG1),
glyceraldehyde-3-phosphate dehydrogenase (GAPDH), heme oxygenase
(decycling) 1 (HMOX1), heat shock 70 kDa protein 1A/B (HSPA1A/B)
and heat shock 70 kDa protein 6 (HSPA6 or HSP70B′) were designed
based on the database. GAPDH was used as a control for
normalization, as previously described (24–26).
Statistical analysis
Data are presented as the means ± standard
deviations (SDs). Differences between pairs of data sets were
analyzed using Student's t-test, with values of P<0.05
considered to indicate statistically significant differences.
Results
Hydroxyl radical and
NO2/NO3 formations induced by exposure to
CAP
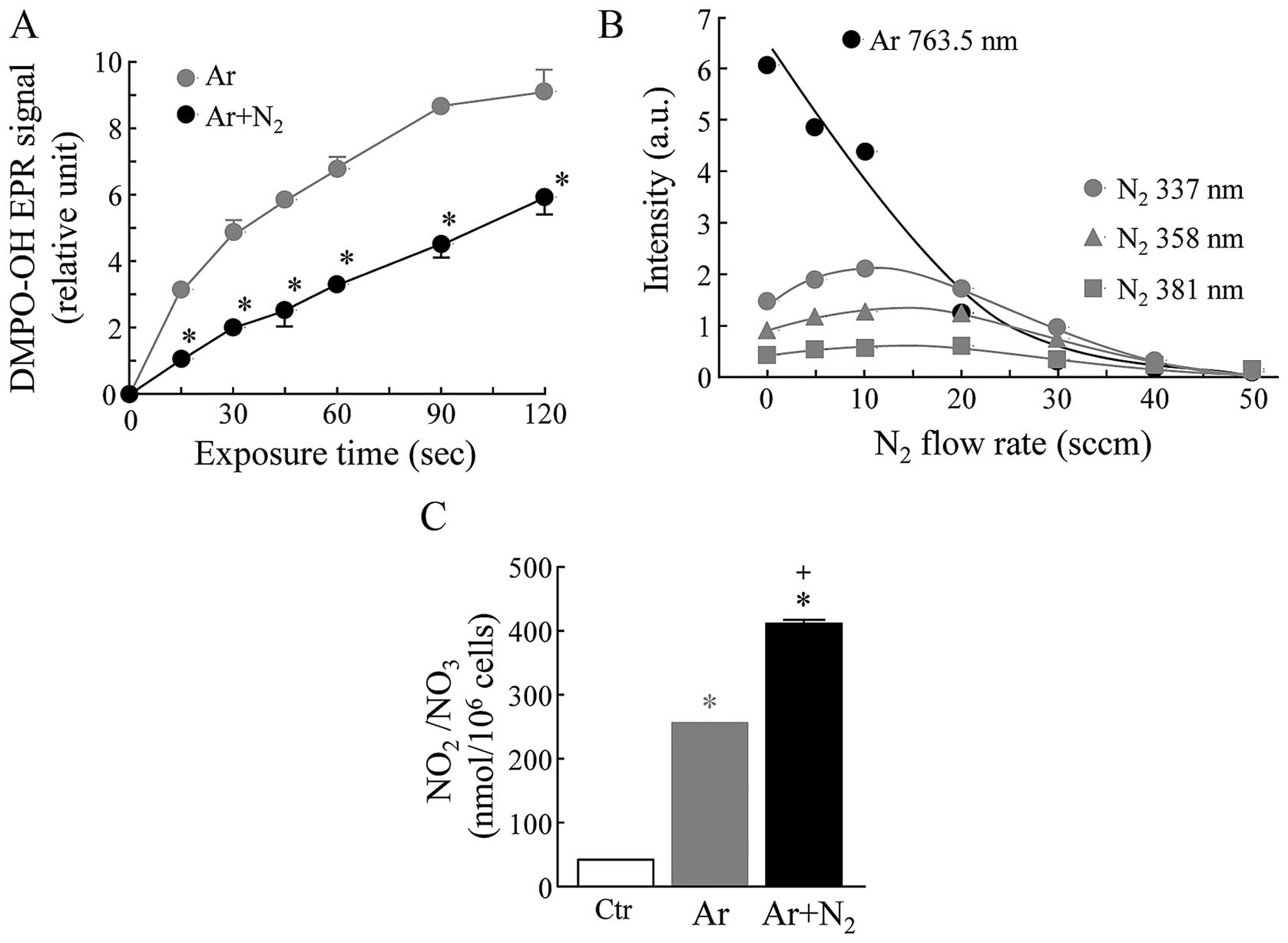
EPR-spin trapping experiments were carried out with
DMPO as a spin trap to detect ·OH radicals in aqueous
solution exposed to Ar-CAP or Ar + N2 (2.5%)-CAP
generated by using a 20 kHz low frequency at 18 kV with a flow rate
of 2 l/min at room temperature. The formations of ·OH
radicals as measured by the amount of DMPO-OH adducts were markedly
increased in the aqueous DMPO solutions exposed to Ar-CAP in an
exposure time-dependent manner. When the aqueous DMPO solutions
were exposed to Ar + N2-CAP, the levels of DMPO-OH
adducts were significantly reduced to approximately 50% of the
levels induced by Ar-CAP (Fig.
1A). The effects of N2 on the intensity of optical
emissions generated by Ar-CAP were also investigated. In our
previous study, we already confirmed that the emission lines and
bands from excited Ar and N2 were mainly observed at
696.5–852.1 and 316.0–416.0 nm, respectively (22). The intensity of emission spectra
from excited Ar at 763.5 nm was readily decreased depending on the
increase in the amount of N2 in Ar gas. On the other
hand, a slight and transient increase in the intensity of emission
spectra from excited N2 at 337.0, 358.0 and 381 nm was
observed in the N2-exposed samples (Fig. 1B). A much greater amount of total
NO2/NO3 (255.0±0.87 nmol/106
cells, mean ± SD) in the supernatant was detected in the human
lymphoma U937 cells exposed to Ar-CAP compared to the control cells
(41.7±0.49) (Fig. 1C). Moreover,
a further elevation in the amount of total
NO2/NO3 (410.3±4.0 nmol/106 cells,
mean ± SD) was observed in the Ar + N2-CAP-exposed
cells.
Effects of Ar- and Ar + N2-CAP
on cell viability and apoptosis
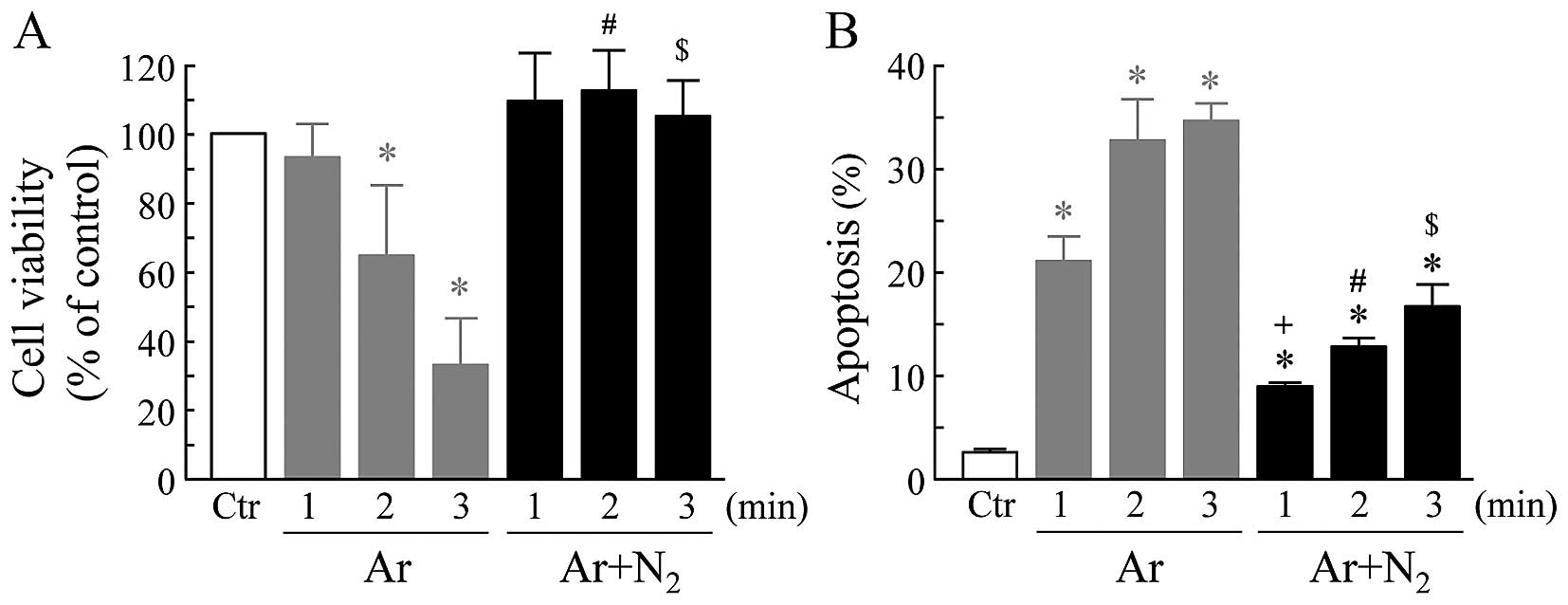
The viability or apoptosis of the cells following
exposure to CAP for 1 to 3 min followed by culture at 37°C for 18
or 6 h, respectively, was investigated. As demonstrated in Fig. 2, Ar-CAP significantly decreased
cell viability in an exposure-time dependent manner. By contrast,
cell viability was not affected by the exposure of the cells to Ar
+ N2-CAP (Fig. 2A). A
significant induction of apoptosis was observed in the cells
exposed to Ar-CAP in an exposure-time dependent manner. The
addition of N2 to the Ar gas markedly suppressed
Ar-CAP-induced apoptosis, and the percentage of suppression was
approximately 50% (Fig. 2B).
Gene expression analysis
The gene expression patterns in the cells following
exposure to CAP for 2 min followed by culture at 37°C for 3 h was
monitored using a GeneChip® microarray system. The
complete lists of probe sets from all samples are deposited at the
Gene Expression Omnibus, a public data base (accession no.
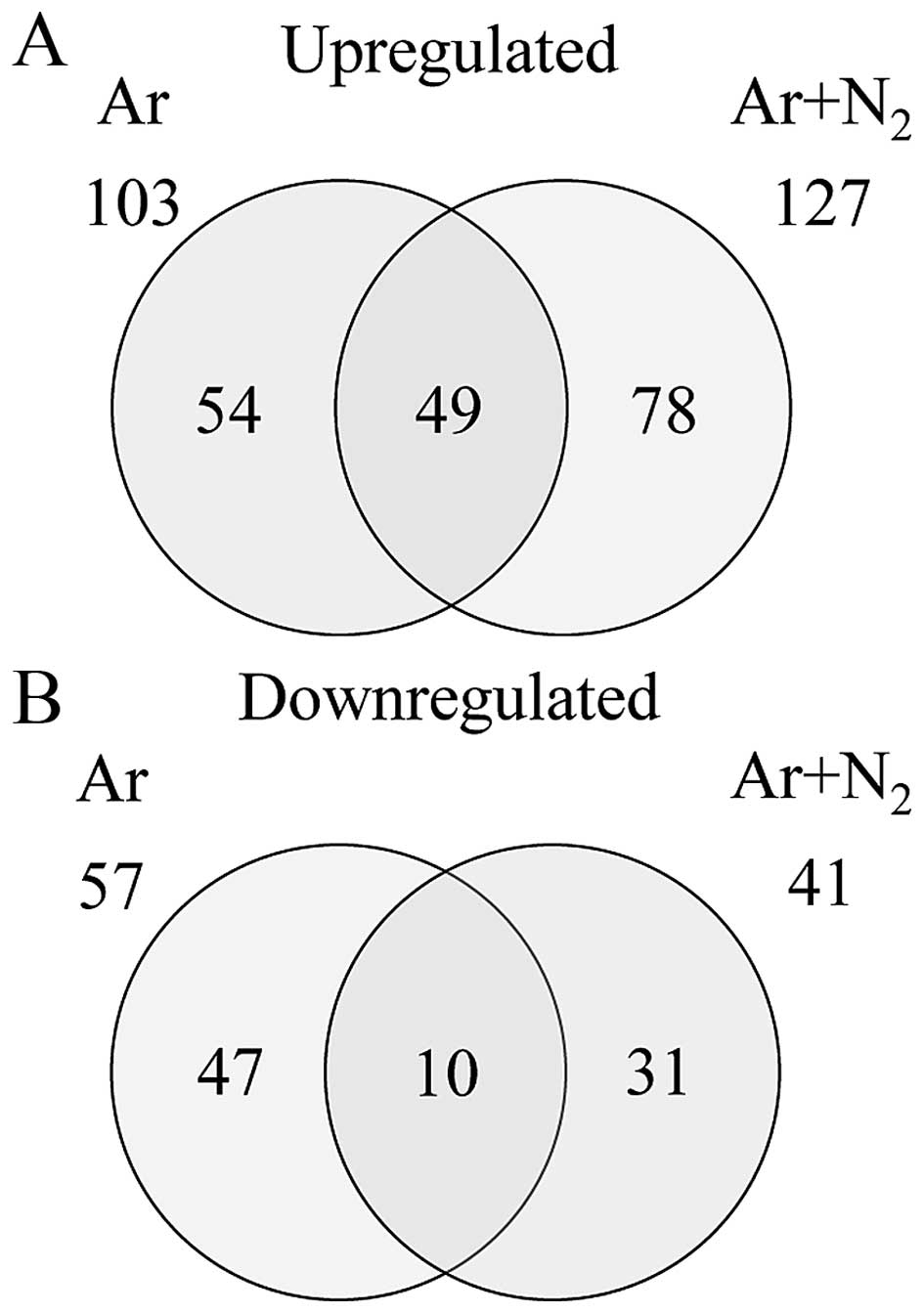
GSE76022). Gene expression analysis using GeneSpring®
software revealed that a number of genes were differentially
expressed by a factor of ≥2.0 between the cells exposed to Ar- or
Ar + N2-CAP and the control cells. The numbers of genes
expressed in either group or commonly in both groups are shown in
the Venn diagram in Fig. 3. The
total numbers of genes that were found to be differentially
expressed were 160 (103 up- and 57 downregulated genes) and 168
(127 up- and 41 downregulated genes) in the Ar- and Ar +
N2-CAP groups, respectively. In addition, the numbers of
commonly up- and downregulated genes were 49 and 10, respectively
(Fig. 3).
Identification of gene networks
associated with apoptosis
To identify gene networks associated with
CAP-induced apoptosis, functional category and pathway analyses
were conducted by using Ingenuity® Pathway Analysis
tools. A number of functionally annotated genes were identified
among both the upregulated and downregulated genes of the Ar- and
Ar + N2-CAP groups. We identified two gene networks,
designated as the pro-apoptosis gene network and the anti-apoptosis
gene network, in the functionally annotated and upregulated genes
of the Ar-CAP- and Ar + N2-CAP-exposed groups, and
showed that these were mainly associated with the biological
functions of the induction and prevention of apoptosis,
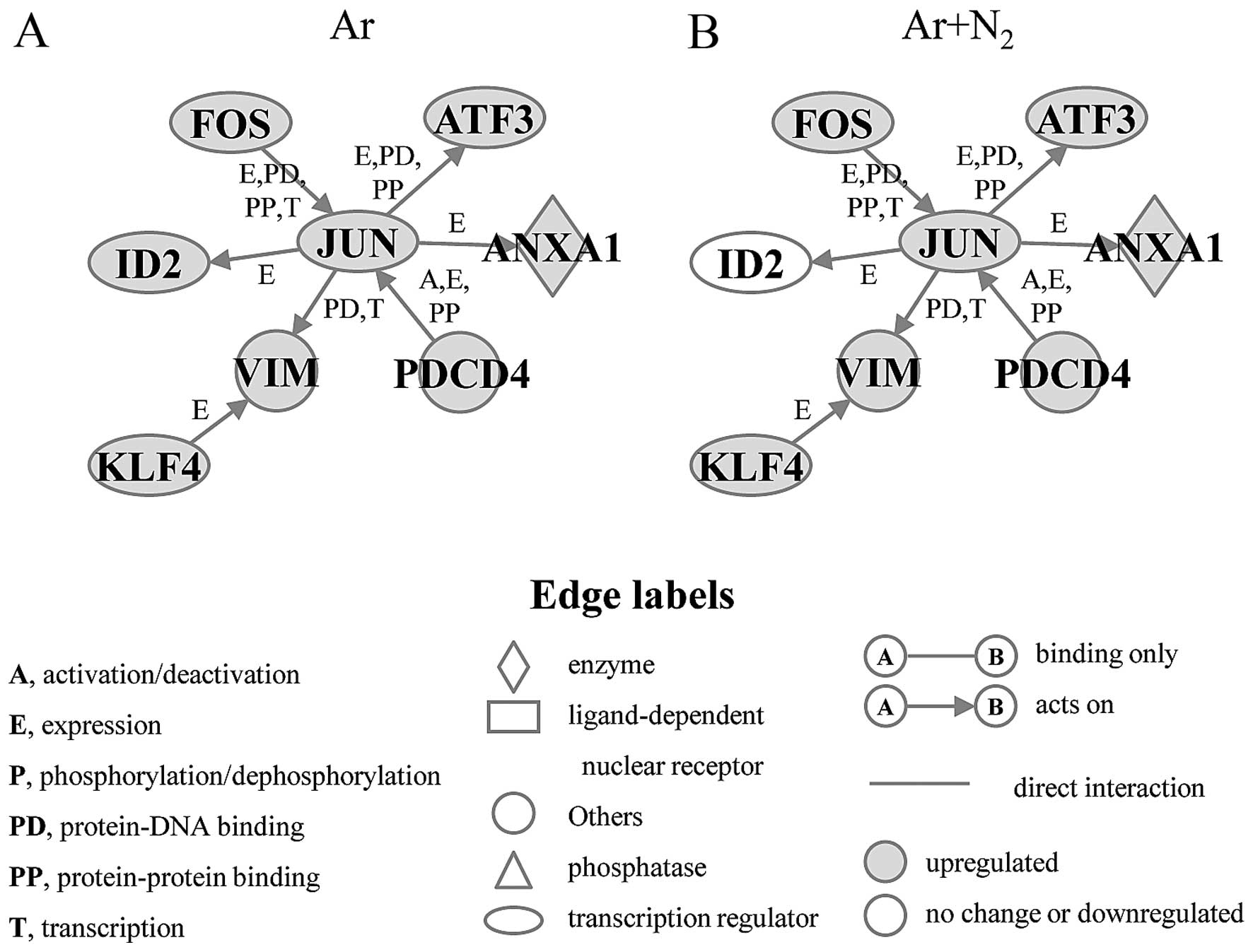
respectively. The pro-apoptosis gene network included 8 genes,
namely Annexin A1 (ANXA1), activating transcription factor 3
(ATF3), FBJ murine osteosarcoma viral oncogene homolog (FOS),
inhibitor of DNA binding 2, dominant negative helix-loop-helix
protein (ID2), jun proto-oncogene (JUN), Kruppel-like factor 4
(KLF4), programmed cell death 4 (PDCD4), and vimentin (VIM). The
expression levels of 7 of these 8 genes, with ID2 being the
exception, were increased under both the Ar-CAP- and Ar +
N2-CAP exposure conditions. A significant elevation in
the ID2 levels was only observed in the cells exposed to Ar-CAP
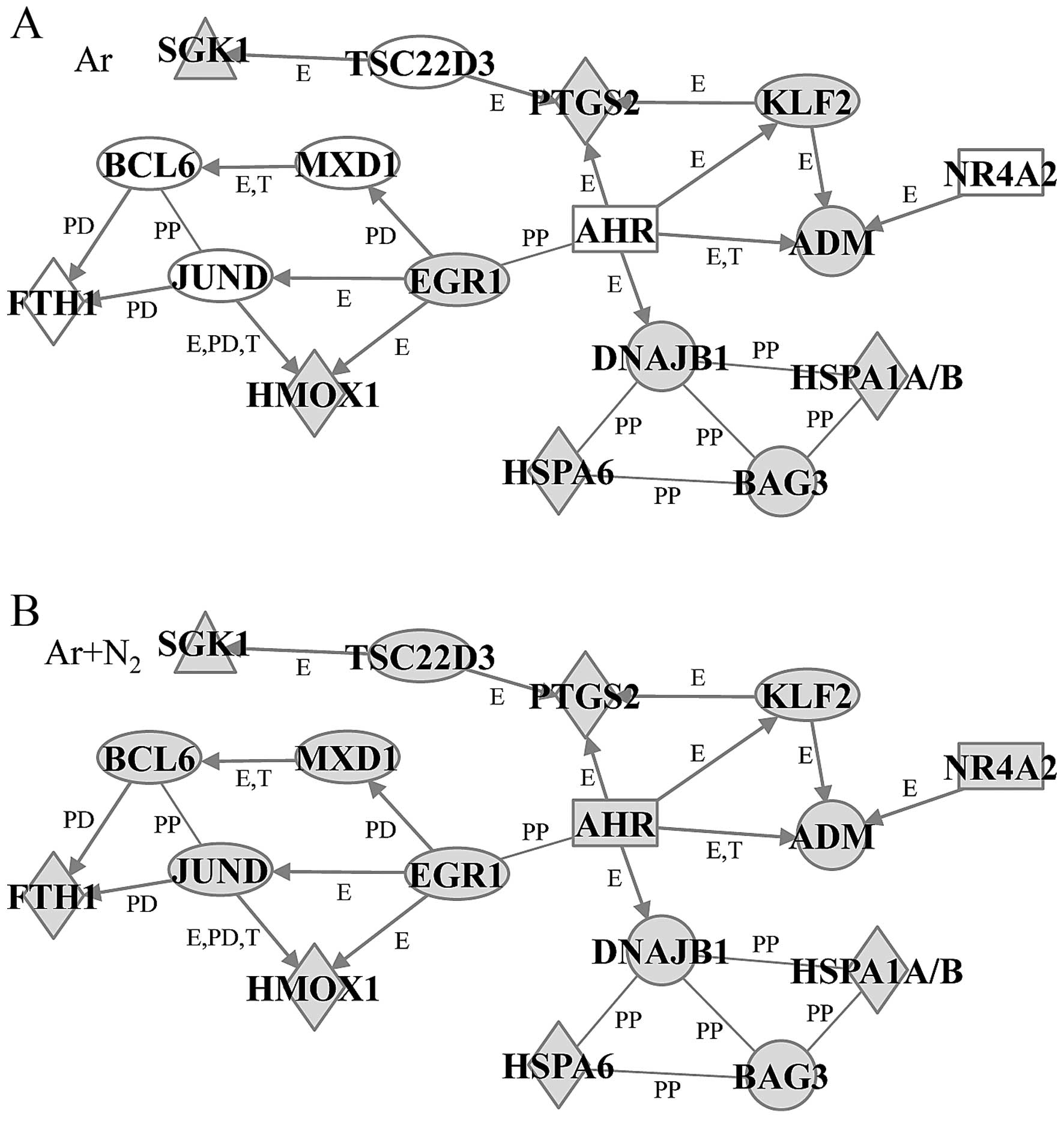
(Fig. 4). The anti-apoptosis gene
network contained 17 genes, including HSPs, DNAJB1, HMOX1, HSPA1A/B
and HSPA6, and adrenomedullin (ADM), aryl hydrocarbon receptor
(AHR), BAG3, B-cell CLL/lymphoma 6 (BCL6), EGR1, ferritin, heavy
polypeptide 1 (FTH1), jun D proto-oncogene (JUND), KLF2, MAX
dimerization protein 1 (MXD1), nuclear receptor subfamily 4 group A
member 2 (NR4A2), prostaglandin-endoperoxide synthase 2 (PTGS2),
serum/glucocorticoid regulated kinase 1 (SGK1) and TSC22 domain
family member 3 (TSC22D3). In this network, the expression levels
of 7 genes, AHR, BCL6, FTH1, JUND, MXD1, NR4A2 and TSC22D3, were
significantly lower in the Ar-CAP-exposed cells than in the Ar +
N2-CAP-exposed cells. However, the other 10 genes
remained significantly elevated under both conditions (Fig. 5).
Effects of Ar- and Ar + N2-CAP
on gene expression, as shown by qPCR
The mRNA levels in the cells following exposure to
CAP for 2 min followed by culture at 37°C for 3 h was monitored by
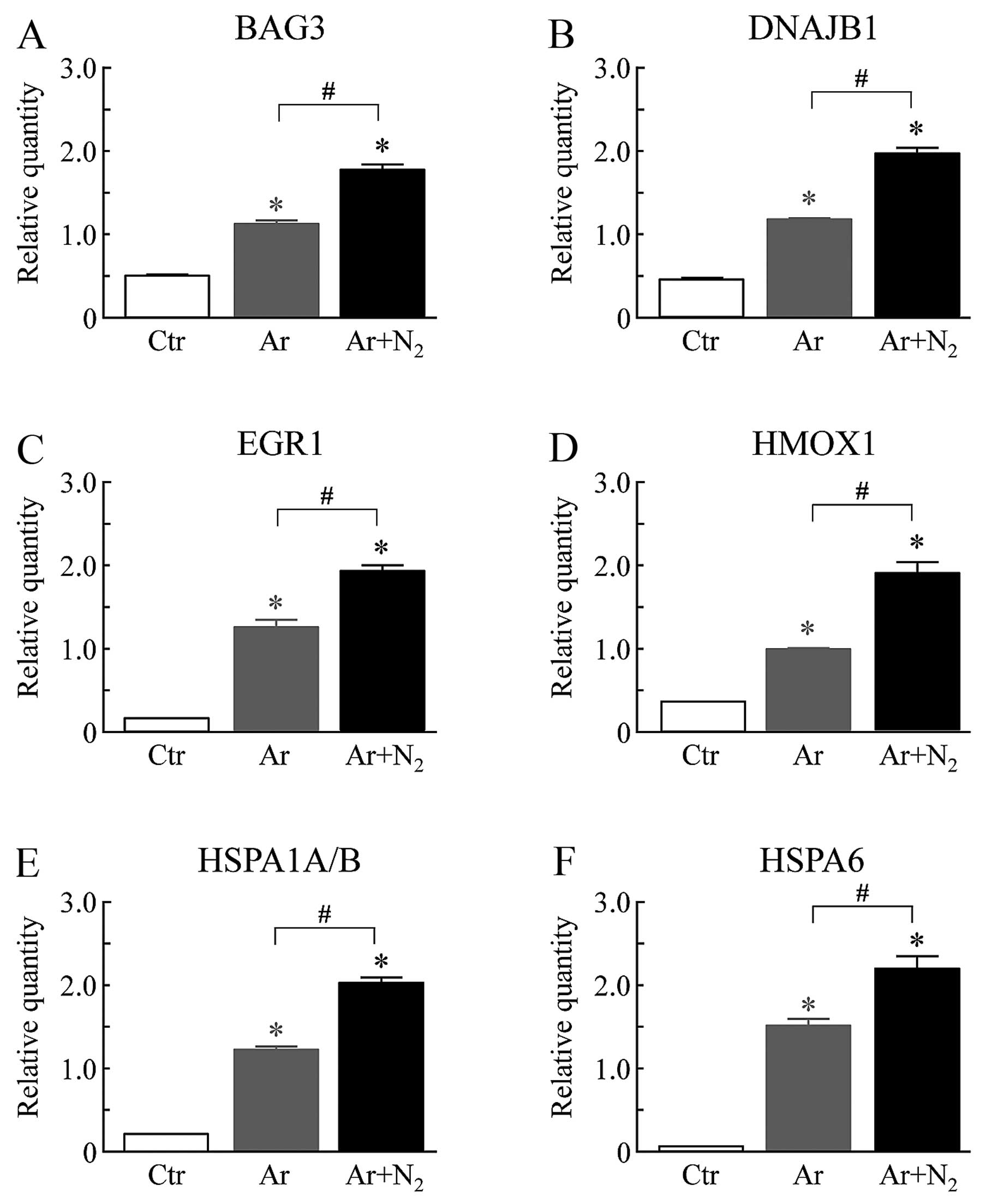
using real-time qPCR. We selected 6 genes, BAG3, DNAJB1, EGR1,
HMOX1, HSPA1A/B and HSPA6, from the anti-apoptosis gene network.
The expression levels of these 6 genes were significantly elevated
in the Ar-CAP-exposed cells compared to the control cells (Fig. 6). Under the Ar + N2-CAP
exposure conditions, a further increase in the expression levels of
these genes was observed in comparison with those under the Ar-CAP
exposure conditions.
Discussion
CAP is well known as a source of biologically active
agents, such as ROS and RNS (1–4),
and its characteristics can be modified by altering the
experimental conditions (3–6).
In the present study, enormous amounts of ·OH radicals
in aqueous solution were produced using an Ar-CAP-generating system
as reported previously (22).
Moreover, Ar gas modification by the addition of N2
resulted in decreased levels of ·OH radicals in the
aqueous solutions and increased levels of
NO2/NO3 in the supernatant of human lymphoma
U937 cells, indicating that N2 gas in Ar-CAP modified
the ratio of ROS to RNS. In the system used in this study and in
our previous study, a certain amount of
NO2/NO3 was detected even if Ar-CAP was used,
suggesting that the effluent Ar gas mixed with atmospheric air in
remote regions can influence the CAP characteristics (22).
There is accumulating experimental evidence that CAP
is an effective agent to suppress cancer cell growth and to induce
apoptosis under both in vivo and in vitro models, and
CAP-produced ROS and/or RNS play a principal role in its anticancer
effects (12–22). In general, relatively low levels
of ROS are essential to maintain the physiological homeostasis of
the cell. On the other hand, increased levels of ROS may be
detrimental and lead to cell death, including apoptosis (27). Kalghatgi et al (15) suggested that the dose-dependent
effects of CAP, which range from increased cell proliferation to
apoptosis, are related to the amount of ROS (15). In our previous studies using U937
cells, the degree of apoptosis was well associated with the degree
of produced ROS under a variety of experimental conditions
(28–30). In the present study, a marked
induction of apoptosis was observed in the U937 cells exposed to
Ar-CAP, whereas the addition of N2 to the Ar gas
significantly suppressed the Ar-CAP-induced increase in apoptosis.
These results suggest that these inhibitory effects of
N2 may be due to a decrease in ROS and/or increase in
RNS levels.
To elucidate the molecular mechanisms underlying
CAP-induced apoptosis, gene expression patterns were investigated
using a combination of high-density oligonucleotide microarray and
computational gene expression analysis tools. In the present study,
we identified a number of genes that exhibited a ≥2.0-fold
difference in expression between the cells exposed to Ar- or Ar +
N2-CAP and the control cells. In addition, by using
Ingenuity® pathway analysis, we discovered a significant
gene network, herein designated as the pro-apoptosis gene network,
that was associated with the biological function of the induction
of apoptosis. In this network, ANXA1 (31), ATF3 (32), FOS (33), JUN (34) and KLF4 (35) have been reported to function as
pro-apoptotic molecules in a wide variety of cell types, including
cancer cells. JUN, ATF3 and FOS belong to the basic-region leucine
zipper (bZIP) transcription factor family, and the homo- and
heterodimeric bZIP protein complexes act as activators and
suppressors of transcription. For example, the JUN and FOS proteins
form the heterodimer of the activating protein-1 (AP-1) complex
(36). These three transcription
factors have been shown to be markedly upregulated in U937 cells
undergoing heat stress-induced apoptosis (24). Previous findings have suggested
that the overexpression of JUN in combination with FOS enhances the
sensitivity of keratinocytes to apoptosis (37). In this network, almost all genes
were upregulated under both the Ar- and Ar + N2-CAP
conditions, with the single exception being ID2, whose expression
was observed only under the Ar-CAP conditions (Fig. 4). The elevation of gene expression
in the pro-apoptosis gene network may have been closely associated
with the induction of apoptosis by CAP in U937 cells.
In addition, we successfully identified the
anti-apoptosis gene network, which was associated with the
biological function of the prevention of apoptosis from upregulated
genes in Ar + N2-CAP-exposed cells (Fig. 5). In this network, 7 genes,
including AHR (38), FTH1
(39) and NR4A2 (40), which have been reported to inhibit
apoptosis, were expressed at lower levels under the Ar-CAP
conditions compared to the Ar + N2-CAP conditions. The
downregulation of these genes may contribute to the ability to
induce apoptosis induced by Ar-CAP. The anti-apoptosis gene network
also contained several HSPs, DNAJB1, HMOX1, HSPA1A/B and HSPA6, and
BAG3. Of note, qPCR clearly demonstrated that the expression levels
of these genes were significantly higher under either the Ar- or Ar
+ N2-CAP conditions than under the control conditions
(Fig. 6). HSPs and BAG3, a
co-chaperone of HSP70, are primarily regulated by heat shock
transcription factor 1 (HSF1), and these proteins function as
anti-apoptotic molecules against various types of stress,
particularly heat (41,42). DNAJB1 (43), HMOX1 (44), HSPA1A/B (43), HSPA6 (45) and BAG3 (46) are known to participate in the
prevention of apoptosis. Previous findings have indicated that very
weak but nonetheless significant heat-inducible heat shock
element-binding activity of HSF1 was observed when cells were
incubated at 39°C for 20 min (47). In the same CAP system as used
herein and in our previous study, temperature increases of
1.7±0.2°C (mean ± SD) and 2.3±0.2°C were observed following 2 and 5
min of exposure at room temperature, respectively (22). It is also known that HSF1 can be
activated directly by oxidative stress (48). Therefore, we considered that the
induction of HSP-related genes may have been due to the activation
of HSF1 by ROS, rather than to an increase in temperature under
both CAP conditions. In the present study, further elevations in
the total amount of NO2/NO3 and the
overexpression of HSP-related genes were detected in the cells
exposed to Ar + N2-CAP (Figs. 1C and 6). Previous studies have indicated that
nitric oxide (NO) is generated by CAP with atmospheric
N2 and is then rapidly converted to other species
including NOX (49). In previous
studies, NO has been reported to induce Hsp70 expression and lead
to cytoprotection in cells (50,51) and the induction of HSP70
expression by NO has shown to be regulated by HSF1 activation
(52). Under our CAP conditions,
the induction of HSPs via NO generation may have participated in
the suppression of Ar-CAP-induced apoptosis by N2.
The findings of the present study provide insight
toward the eventual elucidation of the molecular mechanisms
underlying the CAP-induced apoptosis of cancer cells. The
modulation of gaseous conditions in CAP may be useful for future
clinical applications, such as when switching from a sterilizing
mode to a cytocidal effect for the treatment of cancer.
Abbreviations:
|
ADM
|
adrenomedullin
|
|
AHR
|
aryl hydrocarbon receptor
|
|
ANXA1
|
annexin A1
|
|
ATF3
|
activating transcription factor 3
|
|
BAG3
|
BCL2-associated athanogene 3
|
|
BCL6
|
B-cell CLL/lymphoma 6
|
|
bZIP
|
basic-region leucine zipper
|
|
CAP
|
cold atmospheric plasma
|
|
DNAJB1
|
DnaJ (HSP40) homolog, subfamily B,
member 1
|
|
DMPO
|
5,5-dimethyl-1-pyrroline-N-oxide
|
|
EPR
|
electron paramagnetic resonance
|
|
ERG1
|
early growth response 1
|
|
FITC
|
fluorescein isothiocyanate
|
|
FOS
|
FBJ murine osteosarcoma viral oncogene
homolog
|
|
FTH1
|
ferritin, heavy polypeptide 1
|
|
GAPDH
|
glyceraldehyde-3-phosphate
dehydrogenase
|
|
HMOX1
|
heme oxygenase (decycling) 1
|
|
HSF1
|
heat shock transcription factor 1
|
|
HSPs
|
heat shock proteins
|
|
HSPA1A/B
|
heat shock 70 kDa protein 1A/B
|
|
HSPA6
|
heat shock 70 kDa protein 6
|
|
ID2
|
inhibitor of DNA binding 2, dominant
negative helix-loop-helix protein
|
|
JUN
|
jun proto-oncogene
|
|
JUND
|
jun D proto-oncogene
|
|
KLF
|
Kruppel-like factor
|
|
MXD1
|
MAX dimerization protein 1
|
|
NR4A2
|
nuclear receptor subfamily 4 group A
member 2
|
|
PDCD4
|
programmed cell death 4
|
|
PTGS2
|
prostaglandin-endoperoxide synthase
2
|
|
qPCR
|
quantitative polymerase chain
reaction
|
|
RNS
|
reactive nitrogen species
|
|
ROS
|
reactive oxygen species
|
|
SDs
|
standard deviations
|
|
SGK1
|
serum/glucocorticoid regulated kinase
1
|
|
TSC22D3
|
TSC22 domain family member 3
|
|
VIM
|
vimentin
|
Acknowledgments
This study was supported in part by a Grant-in-Aid
for Scientific Research on Innovative Areas, grant nos. 25108503
and 15H00892 from the Ministry of Education, Culture, Sports,
Science and Technology of Japan.
References
|
1
|
Hoffmann C, Berganza C and Zhang J: Cold
Atmospheric Plasma: Methods of production and application in
dentistry and oncology. Med Gas Res. 3:212013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mai-Prochnow A, Murphy AB, McLean KM, Kong
MG and Ostrikov KK: Atmospheric pressure plasmas: Infection control
and bacterial responses. Int J Antimicrob Agents. 43:508–517. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Arjunan KP, Sharma VK and Ptasinska S:
Effects of atmospheric pressure plasmas on isolated and cellular
DNA-a review. Int J Mol Sci. 16:2971–3016. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Isbary G, Shimizu T, Li YF, Stolz W,
Thomas HM, Morfill GE and Zimmermann JL: Cold atmospheric plasma
devices for medical issues. Expert Rev Med Devices. 10:367–377.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Joh HM, Choi JY, Kim SJ, Chung TH and Kang
TH: Effect of additive oxygen gas on cellular response of lung
cancer cells induced by atmospheric pressure helium plasma jet. Sci
Rep. 4:66382014. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hertwig C, Steins V, Reineke K, Rademacher
A, Klocke M, Rauh C and Schlüter O: Impact of surface structure and
feed gas composition on Bacillus subtilis endospore inactivation
during direct plasma treatment. Front Microbiol. 6:7742015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Brun P, Brun P, Vono M, Venier P,
Tarricone E, Deligianni V, Martines E, Zuin M, Spagnolo S,
Cavazzana R, et al: Disinfection of ocular cells and tissues by
atmospheric-pressure cold plasma. PLoS One. 7:e332452012.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Schmidt A, Dietrich S, Steuer A, Weltmann
KD, von Woedtke T, Masur K and Wende K: Non-thermal plasma
activates human keratinocytes by stimulation of antioxidant and
phase II pathways. J Biol Chem. 290:6731–6750. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Duske K, Koban I, Kindel E, Schröder K,
Nebe B, Holtfreter B, Jablonowski L, Weltmann KD and Kocher T:
Atmospheric plasma enhances wettability and cell spreading on
dental implant metals. J Clin Periodontol. 39:400–407. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Steinbeck MJ, Chernets N, Zhang J, Kurpad
DS, Fridman G, Fridman A and Freeman TA: Skeletal cell
differentiation is enhanced by atmospheric dielectric barrier
discharge plasma treatment. PLoS One. 8:e821432013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Hirst AM, Frame FM, Maitland NJ and
O'Connell D: Low temperature plasma: A novel focal therapy for
localized prostate cancer? BioMed Res Int. 2014:8783192014.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Partecke LI, Evert K, Haugk J, Doering F,
Normann L, Diedrich S, Weiss FU, Evert M, Huebner NO, Guenther C,
et al: Tissue tolerable plasma (TTP) induces apoptosis in
pancreatic cancer cells in vitro and in vivo. BMC Cancer.
12:4732012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Walk RM, Snyder JA, Srinivasan P, Kirsch
J, Diaz SO, Blanco FC, Shashurin A, Keidar M and Sandler AD: Cold
atmospheric plasma for the ablative treatment of neuroblastoma. J
Pediatr Surg. 48:67–73. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Utsumi F, Kajiyama H, Nakamura K, Tanaka
H, Mizuno M, Ishikawa K, Kondo H, Kano H, Hori M and Kikkawa F:
Effect of indirect nonequilibrium atmospheric pressure plasma on
anti-proliferative activity against chronic chemo-resistant ovarian
cancer cells in vitro and in vivo. PLoS One. 8:e815762013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kalghatgi S, Kelly CM, Cerchar E, Torabi
B, Alekseev O, Fridman A, Friedman G and Azizkhan-Clifford J:
Effects of non-thermal plasma on mammalian cells. PLoS One.
6:e162702011. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhao S, Xiong Z, Mao X, Meng D, Lei Q, Li
Y, Deng P, Chen M, Tu M, Lu X, et al: Atmospheric pressure room
temperature plasma jets facilitate oxidative and nitrative stress
and lead to endoplasmic reticulum stress dependent apoptosis in
HepG2 cells. PLoS One. 8:e736652013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ahn HJ, Kim KI, Hoan NN, Kim CH, Moon E,
Choi KS, Yang SS and Lee JS: Targeting cancer cells with reactive
oxygen and nitrogen species generated by atmospheric-pressure air
plasma. PLoS One. 9:e861732014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Thiyagarajan M, Anderson H and Gonzales
XF: Induction of apoptosis in human myeloid leukemia cells by
remote exposure of resistive barrier cold plasma. Biotechnol
Bioeng. 111:565–574. 2014. View Article : Google Scholar
|
|
19
|
Ma Y, Ha CS, Hwang SW, Lee HJ, Kim GC, Lee
KW and Song K: Non-thermal atmospheric pressure plasma
preferentially induces apoptosis in p53-mutated cancer cells by
activating ROS stress-response pathways. PLoS One. 9:e919472014.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kaushik N, Kumar N, Kim CH, Kaushik NK and
Choi EH: Dielectric barrier discharge plasma efficiently delivers
an apoptotic response in human monocytic lymphoma. Plasma Process
Polym. 11:1175–1187. 2014. View Article : Google Scholar
|
|
21
|
Adachi T, Tanaka H, Nonomura S, Hara H,
Kondo S and Hori M: Plasma-activated medium induces A549 cell
injury via a spiral apoptotic cascade involving the
mitochondrial-nuclear network. Free Radic Biol Med. 79:28–44. 2015.
View Article : Google Scholar
|
|
22
|
Uchiyama H, Zhao QL, Hassan MA, Andocs G,
Nojima N, Takeda K, Ishikawa K, Hori M and Kondo T: EPR-spin
trapping and flow cytometric studies of free radicals generated
using cold atmospheric argon plasma and X-ray irradiation in
aqueous solutions and intracellular milieu. PLoS One.
10:e01369562015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Tabuchi Y, Yunoki T, Hoshi N, Suzuki N and
Kondo T: Genes and gene networks involved in sodium
fluoride-elicited cell death accompanying endoplasmic reticulum
stress in oral epithelial cells. Int J Mol Sci. 15:8959–8978. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Furusawa Y, Tabuchi Y, Wada S, Takasaki I,
Ohtsuka K and Kondo T: Identification of biological functions and
gene networks regulated by heat stress in U937 human lymphoma
cells. Int J Mol Med. 28:143–151. 2011.PubMed/NCBI
|
|
25
|
Kariya A, Tabuchi Y, Yunoki T and Kondo T:
Identification of common gene networks responsive to mild
hyperthermia in human cancer cells. Int J Mol Med. 32:195–202.
2013.PubMed/NCBI
|
|
26
|
Kariya A, Furusawa Y, Yunoki T, Kondo T
and Tabuchi Y: A microRNA-27a mimic sensitizes human oral squamous
cell carcinoma HSC-4 cells to hyperthermia through downregulation
of Hsp110 and Hsp90. Int J Mol Med. 34:334–340. 2014.PubMed/NCBI
|
|
27
|
Finkel T and Holbrook NJ: Oxidants,
oxidative stress and the biology of ageing. Nature. 408:239–247.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li FJ, Kondo T, Zhao QL, Tanabe K, Ogawa
R, Li M and Arai Y: Enhancement of hyperthermia-induced apoptosis
by a free radical initiator, 2,2′-azobis (2-amidinopropane)
dihydrochloride, in human histiocytic lymphoma U937 cells. Free
Radic Res. 35:281–299. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Honda H, Kondo T, Zhao QL, Feril LB Jr and
Kitagawa H: Role of intracellular calcium ions and reactive oxygen
species in apoptosis induced by ultrasound. Ultrasound Med Biol.
30:683–692. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li P, Zhao QL, Wu LH, Jawaid P, Jiao YF,
Kadowaki M and Kondo T: Isofraxidin, a potent reactive oxygen
species (ROS) scavenger, protects human leukemia cells from
radiation-induced apoptosis via ROS/mitochondria pathway in
p53-independent manner. Apoptosis. 19:1043–1053. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Dalli J, Jones CP, Cavalcanti DM, Farsky
SH, Perretti M and Rankin SM: Annexin A1 regulates neutrophil
clearance by macrophages in the mouse bone marrow. FASEB J.
26:387–396. 2012. View Article : Google Scholar :
|
|
32
|
Hua B, Tamamori-Adachi M, Luo Y, Tamura K,
Morioka M, Fukuda M, Tanaka Y and Kitajima S: A splice variant of
stress response gene ATF3 counteracts NF-kappaB-dependent
anti-apoptosis through inhibiting recruitment of CREB-binding
protein/p300 coactivator. J Biol Chem. 281:1620–1629. 2006.
View Article : Google Scholar
|
|
33
|
Zhang X, Zhang L, Yang H, Huang X, Otu H,
Libermann TA, DeWolf WC, Khosravi-Far R and Olumi AF: c-Fos as a
proapoptotic agent in TRAIL-induced apoptosis in prostate cancer
cells. Cancer Res. 67:9425–9434. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Ruiter GA, Zerp SF, Bartelink H, van
Blitterswijk WJ and Verheij M: Alkyl-lysophospholipids activate the
SAPK/JNK pathway and enhance radiation-induced apoptosis. Cancer
Res. 59:2457–2463. 1999.PubMed/NCBI
|
|
35
|
Li Z, Zhao J, Li Q, Yang W, Song Q, Li W
and Liu J: KLF4 promotes hydrogen-peroxide-induced apoptosis of
chronic myeloid leukemia cells involving the bcl-2/bax pathway.
Cell Stress Chaperones. 15:905–912. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Newman JR and Keating AE: Comprehensive
identification of human bZIP interactions with coiled-coil arrays.
Science. 300:2097–2101. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mils V, Piette J, Barette C, Veyrune J,
Tesnière A, Escot C, Guilhou JJ and Basset-Séguin N: The
proto-oncogene c-fos increases the sensitivity of keratinocytes to
apoptosis. Oncogene. 14:1555–1561. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Marlowe JL and Puga A: Aryl hydrocarbon
receptor, cell cycle regulation, toxicity, and tumorigenesis. J
Cell Biochem. 96:1174–1184. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Pham CG, Bubici C, Zazzeroni F, Papa S,
Jones J, Alvarez K, Jayawardena S, De Smaele E, Cong R, Beaumont C,
et al: Ferritin heavy chain upregulation by NF-kappaB inhibits
TNFalpha-induced apoptosis by suppressing reactive oxygen species.
Cell. 119:529–542. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Wang J, Yang J, Zou Y, Huang GL and He ZW:
Orphan nuclear receptor nurr1 as a potential novel marker for
progression in human prostate cancer. Asian Pac J Cancer Prev.
14:2023–2028. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Beere HM: 'The stress of dying': The role
of heat shock proteins in the regulation of apoptosis. J Cell Sci.
117:2641–2651. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kabbage M and Dickman MB: The BAG
proteins: A ubiquitous family of chaperone regulators. Cell Mol
Life Sci. 65:1390–1402. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Evert BO, Wüllner U and Klockgether T:
Cell death in polyglutamine diseases. Cell Tissue Res. 301:189–204.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Bindu S, Pal C, Dey S, Goyal M, Alam A,
Iqbal MS, Dutta S, Sarkar S, Kumar R, Maity P and Bandyopadhyay U:
Translocation of heme oxygenase-1 to mitochondria is a novel
cytoprotective mechanism against non-steroidal anti-inflammatory
drug-induced mitochondrial oxidative stress, apoptosis, and gastric
mucosal injury. J Biol Chem. 286:39387–39402. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Yang Z, Zhuang L, Szatmary P, Wen L, Sun
H, Lu Y, Xu Q and Chen X: Upregulation of heat shock proteins
(HSPA12A, HSP90B1, HSPA4, HSPA5 and HSPA6) in tumour tissues is
associated with poor outcomes from HBV-related early-stage
hepatocellular carcinoma. Int J Med Sci. 12:256–263. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Yunoki T, Tabuchi Y, Hayashi A and Kondo
T: BAG3 protects against hyperthermic stress by modulating NF-κB
and ERK activities in human retinoblastoma cells. Graefes Arch Clin
Exp Ophthalmol. 253:399–407. 2015. View Article : Google Scholar
|
|
47
|
Tanabe M, Nakai A, Kawazoe Y and Nagata K:
Different thresholds in the responses of two heat shock
transcription factors, HSF1 and HSF3. J Biol Chem. 272:15389–15395.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Ahn SG and Thiele DJ: Redox regulation of
mammalian heat shock factor 1 is essential for Hsp gene activation
and protection from stress. Genes Dev. 17:516–528. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Liebmann J, Scherer J, Bibinov N,
Rajasekaran P, Kovacs R, Gesche R, Awakowicz P and Kolb-Bachofen V:
Biological effects of nitric oxide generated by an atmospheric
pressure gas-plasma on human skin cells. Nitric Oxide. 24:8–16.
2011. View Article : Google Scholar
|
|
50
|
Byrne CR and Hanson PJ: Induction of heat
shock protein 72 by a nitric oxide donor in guinea-pig gastric
mucosal cells. Eur J Pharmacol. 353:117–122. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Manucha W and Vallés PG: Cytoprotective
role of nitric oxide associated with Hsp70 expression in neonatal
obstructive nephropathy. Nitric Oxide. 18:204–215. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Manucha W and Vallés P: Hsp70/nitric oxide
relationship in apoptotic modulation during obstructive
nephropathy. Cell Stress Chaperones. 13:413–420. 2008. View Article : Google Scholar : PubMed/NCBI
|