Introduction
The prevalence of diabetes mellitus (DM) is expected
to rise significantly, and DM is viewed as a public health problem
of epidemic proportions (1,2).
DM-induced vascular diseases are major contributors to morbidity
and mortality in the aging population (3).
The endothelium is composed of a monolayer of cells
covering the vascular lumen and it provides a continuous barrier
between the elements of blood and the arterial wall. The
endothelium plays an important role in the regulation of vascular
tone, by balancing the release of endothelium-derived vasodilatory
factors, such as nitric oxide (NO) and prostacyclin, with
endothelium-derived vasoconstrictory factors including endothelin-1
(ET-1), angiotensin II and reactive oxygen species (ROS) (4,5).
In diabetes, the disruption of the balance between vasodilatory and
vasoconstrictor actions in the endothelium is considered as a
crucial event in the initiation of the atherosclerotic process.
The incretin hormone, glucagon-like peptide-1
(GLP-1), is secreted postprandially from the L-cells of the lower
gut and has a short half-life of minutes before it is rapidly
degraded by dipeptidyl peptidase-4 (DPP-4) (6). Sitagliptin is a stable, orally
effective DPP-4 inhibitor, which increases the active concentration
and duration of action of GLP-1 (7,8).
GLP-1 plays a decisive role in glucose-stimulated insulin release
and the preservation of plasma glucose homeostasis. Notably, GLP-1
has been reported to exert cardioprotective and vasodilatory
effects and to attenuate vascular endothelial dysfunction in
vivo and in vitro (9–13).
Most studies have focused on vasodilatory actions, which are
principally mediated by NO, a vasodilator and anti-inflammatory
molecule produced by the action of endothelial nitric oxide
synthase (eNOS) (14,15). However, little is known regarding
vasoconstrictor actions on the vascular endothelium which are
partly mediated by ET-1, a potent vasoconstrictor and
pro-inflammatory molecule secreted by the endothelium in animal
models.
Thus, in the present study, we established a rat
model of diabetes-associated arteriosclerosis in order to determine
whether sitagliptin attenuates endothelial dysfunction and inhibits
ET-1 expression in the aortic endothelium and to explore the
underlying mechanisms responsible for these effects.
Materials and methods
Animal experimental procedures
Twenty-five male Sprague- Dawley rats (6 weeks old,
weighing approximately 150–180 g) were obtained from the
Experimental Animal Centre of Anhui Medical University (Hefei,
China). The rats were randomly divided into the following four
groups: i) control group (n=7); ii) DM group (n=6); iii) DM +
low-dose sitagliptin (10 mg/kg) treatment group (n=6); and iv) DM +
high-dose sitagliptin (30 mg/kg) treatment group (n=6). All groups
were subjected to a 12:12 h light-dark cycle (lights on at 06:00)
under controlled conditions of temperature (22±1°C) and humidity
(50–60%). With the exception of the control group, all of the rats
received a high-fat (HF) diet (2% cholesterol, 10% lard, and 88%
normal diet) and sufficient water for 8 weeks, after which time,
they were subjected to an intraperitoneal glucose tolerance test
(IPGTT) and an intraperitoneal insulin tolerance test (IPITT). The
rats in the DM and the sitagliptin treatment groups were injected
once intraperitoneally with a dose of streptozotocin (25 mg/kg;
Sigma-Aldrich, St. Louis, MO, USA) to induce diabetes and blood
glucose levels were tested 1 week after the streptozotocin
injection. The animals with glucose levels ≥11.1 mmol/l were
considered diabetic. The rats in the control and DM groups were
given normal saline. The rats in the low-dose (10 mg/kg) and
high-dose (30 mg/kg) sitagliptin (Merck Serono Co., Ltd.,
Guangzhou, China) groups received sitagliptin once daily by oral
gavage for 12 weeks. The rats were weighed every 3 days and the
dosage was adjusted accordingly. All the rats were euthanized at
the end of week 21. All experiments were approved by the Ethics
Committee of Anhui Medical University (Hefei, China).
Non-invasive, transcutaneous ultrasound
measurement of blood flow velocity
Non-invasive, transcutaneous ultrasound evaluation
of the blood flow velocity was performed two days prior to the end
of the experiment as previously described (16,17). Briefly, the rats were
anaesthetized using 10% chloral hydrate solution and placed in the
dorsal decubitus position for the shaving of their abdomens. After
10 min of rest in the supine position, ultrasonic examination of
the abdominal aorta was performed using a 13-MHz ultrasound probe
(GES6 two-dimensional Color Doppler Ultrasound Diagnostic
Apparatus; GE Healthcare, Piscataway, NJ, USA). The transducer was
lubricated with ultrasound gel and placed at the level of the
abdominal aorta with minimal pressure. The probe was placed 1.0 cm
precisely below the renal artery to obtain a longitudinal axis view
of the abdominal aorta. Image settings were optimized to allow the
clearest definition of the abdominal aorta. Images of the abdominal
aorta were recorded continuously throughout the entire procedure.
The ultrasound probe remained in the same position for the duration
of the measurement period. The end-diastolic velocity (EDV) and the
peak systolic velocity (PSV) were recorded.
Tissue collection
Following the withdrawal of food overnight, the rats
were deeply anesthetized using 10% chloral hydrate solution (0.3
ml/100 g body weight). Blood samples were obtained from the
abdominal aorta. The aortic arch and the thoracic aorta were
removed. The aortic arch was longitudinally cut and fixed in 4%
paraformaldehyde for subsequent pathological and
immunohistochemical assays. Part of the thoracic aortas were placed
in a dish containing ice-cold Krebs solution (composition in
mmol/l: NaCl, 120; KCl,4.7; KH2PO4, 1.18;
CaCl2, 2.25; NaHCO3, 24.5;
MgSO4•7H2O, 1.2; glucose, 11.1; EDTA, 0.03)
and continuously aerated with 95% O2 and 5%
CO2. The samples were then cut into rings (3 mm in
length) for the measurement of isometric contractile tension. The
remaining thoracic aortas were frozen in liquid nitrogen and stored
at −80°C for reverse transcription-quantitative polymerase chain
reaction (RT-qPCR) and western blot analysis.
Analysis of markers in blood
Blood samples were taken by cardiac puncture, and
the levels of serum fasting blood glucose (FBG), total cholesterol
(Tch), low-density lipoprotein cholesterol (LDL-c) and triglyceride
(TG) were measured using commercially available spectrophotometric
assay kits (Beijing BHKT Clinical Reagent Co., Ltd., Beijing,
China). Fasting and postprandial serum GLP-1 levels were measured
using an enzyme-linked immunosorbent assay kit (Cusabio, Wuhan,
China). Serum levels of ET-1 and insulin were measured using a
radioimmunoassay kit (Beifang Biology, Beijing, China). Serum NO
levels were measured using a commercial kit (Jiancheng, Nanjing,
China). The insulin sensitivity index was calculated using the
[1/(FBG × fasting insulin)] formula (18). All analyses were performed in
duplicate.
Hematoxylin and eosin (H&E)
staining
Fixed aortic specimens were dehydrated, embedded in
paraffin, sectioned (4 µm thickness) and stained with
H&E as previously described (19). The artery wall was observed using
a microscope (Leica, Wetzlar, Germany) and the mean artery wall
thickness was calculated.
Immunohistochemical analysis
Briefly, the sections of the thoracic aorta were
deparaffinised, rehydrated, and fixed with methanol-0.3%
H2O2 solution for 20 min at room temperature.
The antigens were unmasked by cooling for 30 min at room
temperature after high compression heating. Primary antibodies
against ET-1 (Santa Cruz Biotechnology, Inc., Santa Cruz, CA, USA)
were added and incubated with the tissue sections overnight
followed by sequential incubation with a biotinylated antibody
(anti-rat IgG) and horseradish streptavidin for 30 min at 37°C.
Finally, the samples were incubated with diaminobenzidine for
coloration and counterstained with haematoxylin for 2 min to stain
the nuclei. Positively-stained cells appeared brown in colour.
Images were captured using a microscope (Leica), and the integral
optical density values were measured using the JD-801 pathological
imaging analysis system (Jiangsu JEDA Science-Technology
Development Co., Ltd., Jiangsu, China).
Measurement of isometric contractile
tension
Individual aortic rings were vertically suspended
between two stainless steel wire hooks in a jacketed organ bath
containing 25 ml Krebs solution. The Krebs solution was replaced at
15-min intervals. The bathing solution was bubbled continuously
with a mixture of 95% O2 and 5% CO2 at 37°C.
The isometric contractile tension was recorded continuously using a
BL-420F biological function experimental system (Chengdu Taimeng
Science and Technology Co., Ltd., Chengdu, China). The resting
tension applied to the aortic rings was increased incrementally to
a final tension of 2 g. The rings were then equilibrated for 45
min. Following equilibration, the rings were precontracted with 1
µmol/l phenylephrine (Phe); once a stable contraction
plateau was obtained, either 10−8 to 10−4
mol/l acetylcholine (Ach) or 10−8 to 10−4
mol/l sodium nitroprusside (SNP) was added to the organ bath
cumulatively until a maximal vasodilator response was achieved.
Cumulative vasodilator response data are expressed as the
percentage of relaxation relative to the Phe-induced
precontraction.
RT-qPCR
SYBR-Green RT-qPCR was performed in order to detect
the mRNA levels of ET-1. Total RNA was extracted from the aortic
tissue of rats using TRIzol reagent (Invitrogen, Grand Island, NY,
USA). RNA was reverse transcribed to cDNA using the PrimeScript RT
reagent kit (Takara Bio, Otsu, Japan). qPCR was performed with
PikoReal (Thermo Fisher Scientific, Waltham, MA, USA) using SYBR
Premix Ex Taq (Takara Bio). The relative quantification values for
the expression of these genes were calculated using the ΔΔCT method
and corrected using a housekeeping gene. The following primer
sequences for the analysis of ET-1 and GAPDH mRNA were used: ET-1
forward, 5′-CAACCAGACACCGTCCTCTT-3′ and reverse,
5′-CTTGGAAAGCCACAAACAGC-3′; and GAPDH forward,
5′-TCATTGACCTCAACTACA-3′ and reverse,
5′-CAAAGTTGTCATGGATGACC-3′.
Western blot analysis
The aortas were washed three times in TBS, and then
lysed in RIPA buffer. The lysates were centrifuged at 14,000 × g
for 30 min at 4°C. The total protein concentration of each sample
was measured using a Micro BCA Protein Assay reagent kit (Pierce,
Rockford, IL, USA). The same amount of lysate from each line in SDS
sample buffer was resolved by 10% SDS-polyacrylamide gel
electrophoresis and electroblotted onto a PVDF membrane, which was
then blocked with 5% fat-free milk in TBST (TBS, 0.1% Tween-20) for
1 h at room temperature. The ET-1 Ab (1:250 dilution; sc-21625-R;
Santa Cruz Biotechnology, Inc.); phosphorylated (p-)AMP-activated
protein kinase (AMPK) (Thr172) Ab (1:500 dilution; no.
2535) and p-nuclear factor (NF)-κB p65 Ab (1:500 dilution; no.
3033) (both from Cell Signaling Technology, Inc., Danvers, MA,
USA); and p-IκBα Ab (1:250 dilution; sc-8404; Santa Cruz
Biotechnology, Inc.) were incubated overnight at 4°C, followed by
incubation with secondary antibodies conjugated with horseradish
peroxidase (Beijing Zhongshan Jinqiao Biotechnology Co., Ltd.,
Beijing, China). Specific bands of the target proteins were stained
with chemiluminescence reagent (Pierce). Densitometric scanning of
the exposed X-ray film was used for semi-quantitative measurement
of the protein bands. β-actin levels were used to normalise the
relative protein expression levels.
Statistical analysis
All statistical analyses were performed using SPSS
software, version 18.0. All data are presented as the means ±
standard deviation (SD). Comparisons between the groups were
conducted using one-way analysis of variance (ANOVA) and the
Student-Newman-Keuls (SNK) method. A P-value <0.05 was
considered to indicate a statistically significant difference.
Results
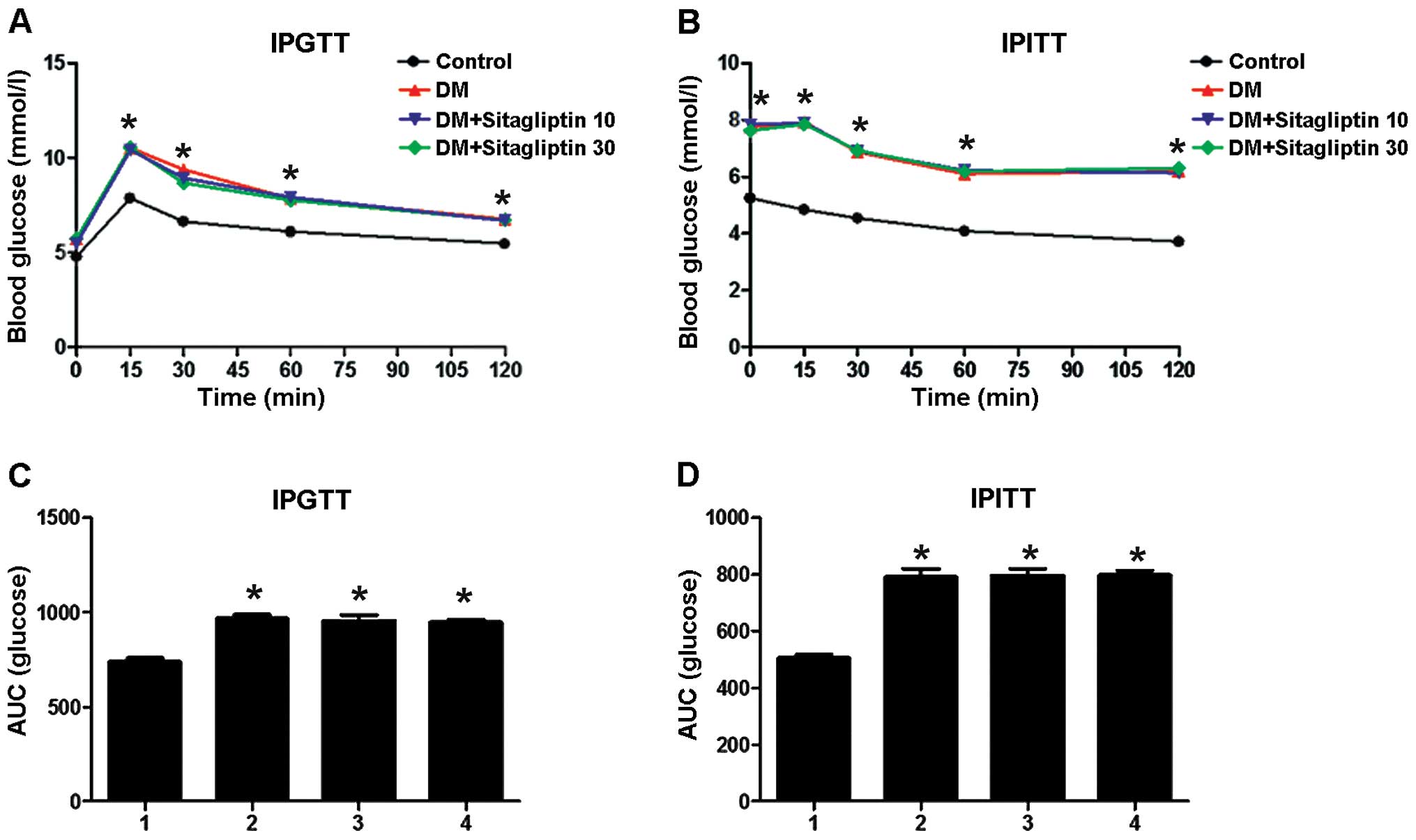
IPGTT and IPITT
Following 8 weeks of HF feeding, IPGTT and IPITT
results revealed that the blood glucose levels in the rats fed an
HF diet were significantly higher than those in the control rats at
baseline (IPITT only) and at 15, 30, 60 and 120-min time-points
(P<0.05). The area under the curve (AUC) for the glucose level
increased significantly in the rats fed an HF diet for 8 weeks
(P<0.05) (Fig. 1). These
results suggested that the rats fed an HF diet were insulin
resistant.
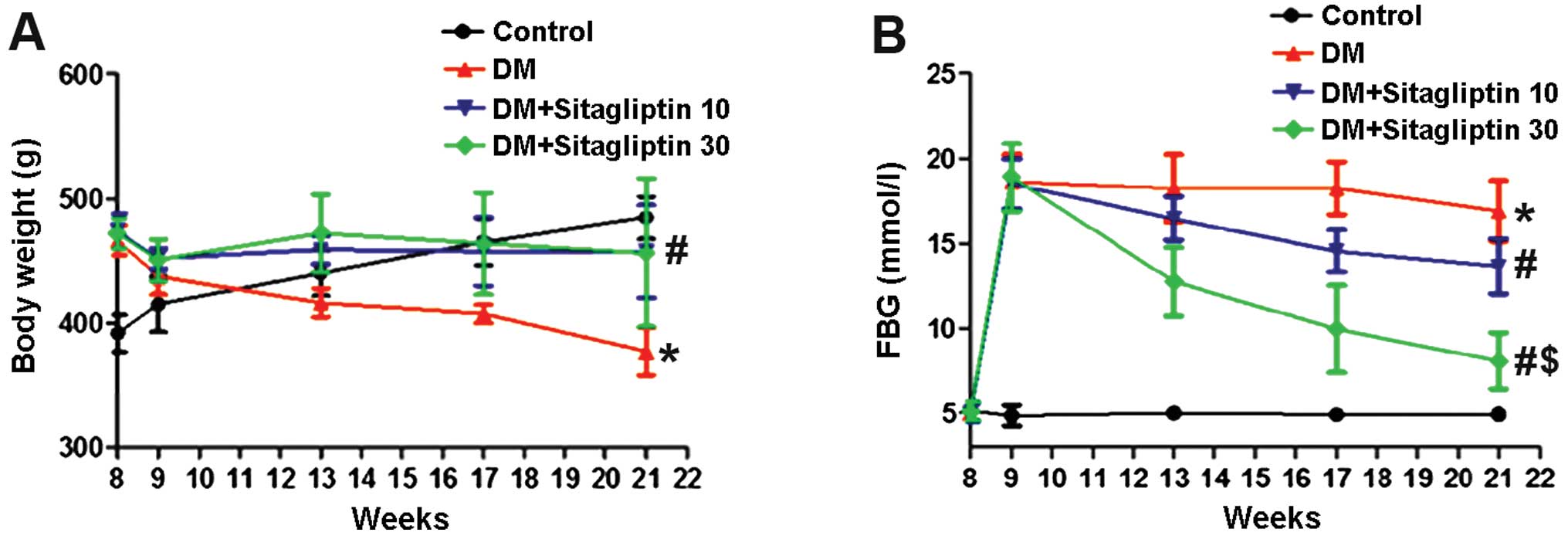
Metabolic parameters in diabetic
rats
Statistical analysis showed that the body weight and
the blood glucose levels were significantly changed in the DM
group, and that sitagliptin reduced the extent of weight loss as
well as the high FBG levels during the experimental period
(Fig. 2). Following 12 weeks of
treatment, FBG, Tch, TG, LDL-c and insulin resistance were
significantly higher in the diabetic rats compared with the control
group (P<0.05). The administration of high-dose sitagliptin
significantly reduced FBG levels and insulin resistance
(P<0.05). However, the levels of Tch, TG and LDL-c were not
significantly affected by low dose of sitagliptin (P>0.05)
(Table I).
 | Table IBasic parameters of rats in different
experimental groups following 12 weeks of treatment. |
Table I
Basic parameters of rats in different
experimental groups following 12 weeks of treatment.
| Controls (n=7) | DM (n=6) | DM + Sitagliptin 10
(n=6) | DM + Sitagliptin 30
(n=6) |
|---|
| Weight (g) |
485.00±17.15a | 377.33±19.07 |
457.83±37.03a |
456.67±59.54a |
| FBG (mmol/l) | 4.93±0.27a | 16.92±1.79 | 13.67±1.61a |
7.1±2.15a,b |
| Tch (mmol/l) | 2.77±0.14a | 6.58±1.6 | 6.56±1.28 | 6.28±1.06 |
| TG (mmol/l) | 1.14±0.07a | 5.01±1.39 | 4.71±1.00 | 4.48±0.51a |
| LDL-c (mmol/l) | 1.08±0.05a | 4.65±1.5 | 4.58±0.97 | 4.29±0.75 |
| Fasting insulin
(uIU/ml) | 12.07±3.36a | 44.95±11.07 | 30.68±7.06a |
16.90±4.17a,b |
| lnISI | −4.05±0.30a | −6.60±0.21 | −6.00±0.24 |
−4.87±0.26a,b |
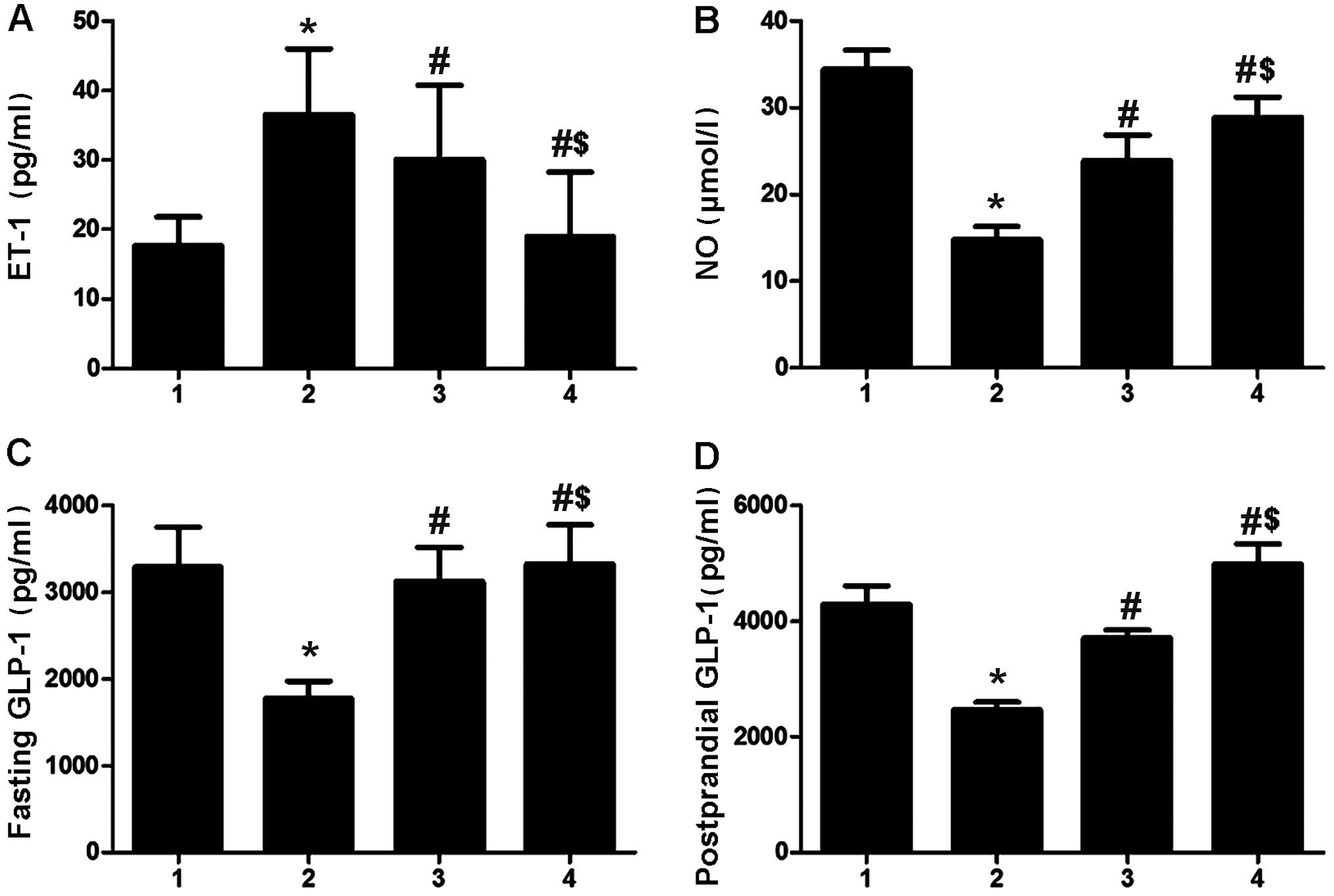
Effects of sitagliptin on serum levels of
GLP-1, ET-1 and NO
The induction of diabetes significantly increased
the serum levels of ET-1 and reduced the levels of fasting and
postprandial GLP-1 as well as NO (P<0.05). The adminstration of
high- and low-dose sitagliptin significantly reduced the serum
levels of ET-1 and elevated the serum levels of GLP-1 as well as NO
to varying extents (P<0.05) (Fig.
3).
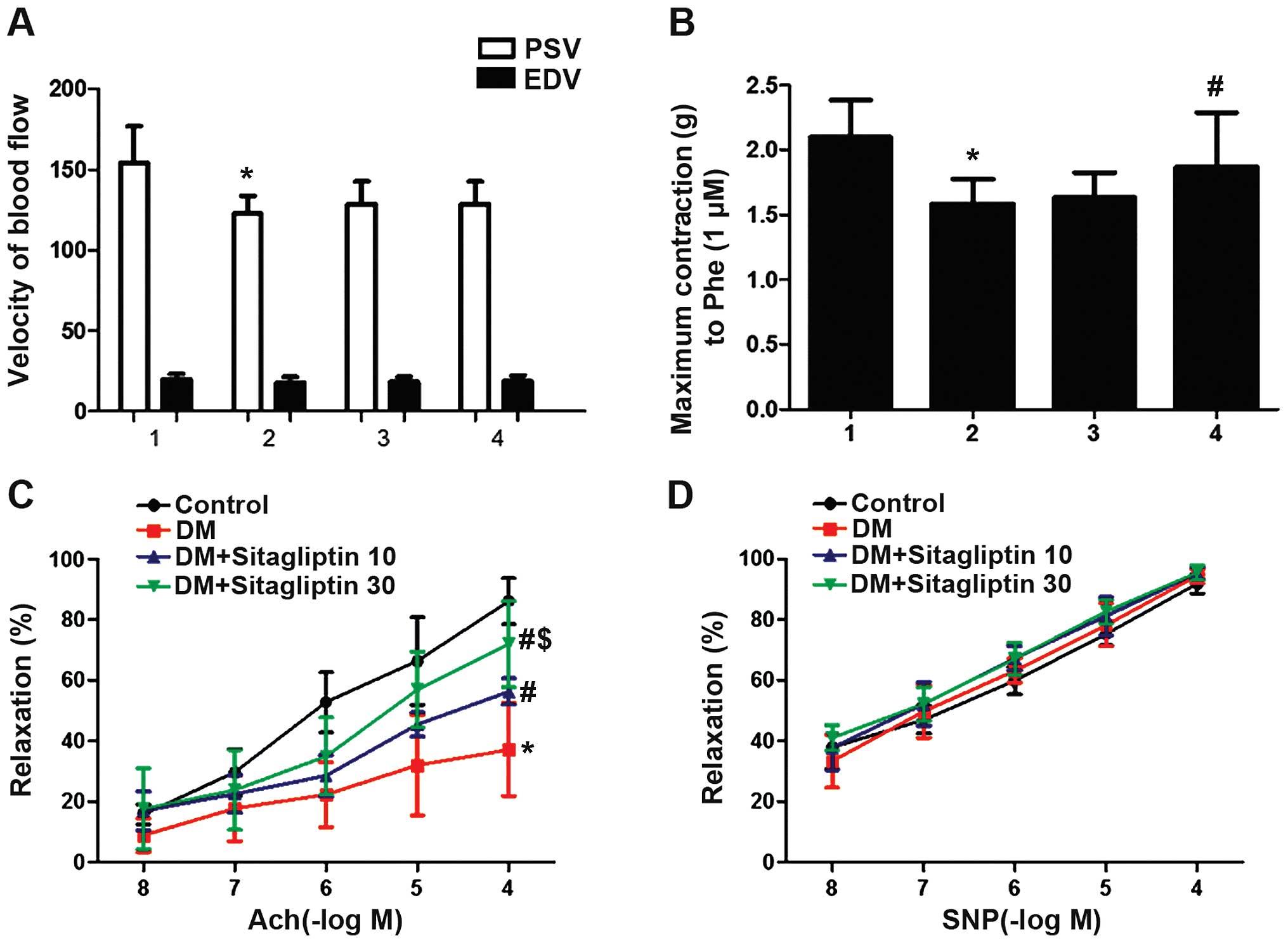
Endothelial function and velocity of
blood flow
We then examined the effects of sitagliptin on
vascular endothelial function in the diabetic rats. Isometric
contractile tension experiments on isolated aortic rings were
performed. As illustrated in Fig.
4, in the DM group, we observed inhibition of the cumulative
contraction-relaxation response to Ach in the thoracic aortic rings
compared with the controls (P<0.05), whereas there were no
significant changes in the cumulative contraction-relaxation
response to SNP (P>0.05). The administration of different doses
of sitagliptin partly reversed the diabetes-induced impairment of
the cumulative concentration-vasodilator responses to Ach in the
thoracic aortic rings precontracted with Phe (P<0.05) and had no
effect on the SNP-induced cumulative contraction-relaxation
responses (Fig. 4C and D). The
contractile response to Phe in the untreated diabetic rats was
lower than that in the control rats and the rats treated with
high-dose sitagliptin (Fig. 4B).
In addition, performing non-invasive transcutaneous ultrasound
measurement of blood flow velocity in the abdominal aorta showed
that PSV was significantly reduced in the DM group, and was not
significantly increased by the administration of sitagliptin
(P<0.05) (Fig. 4A).
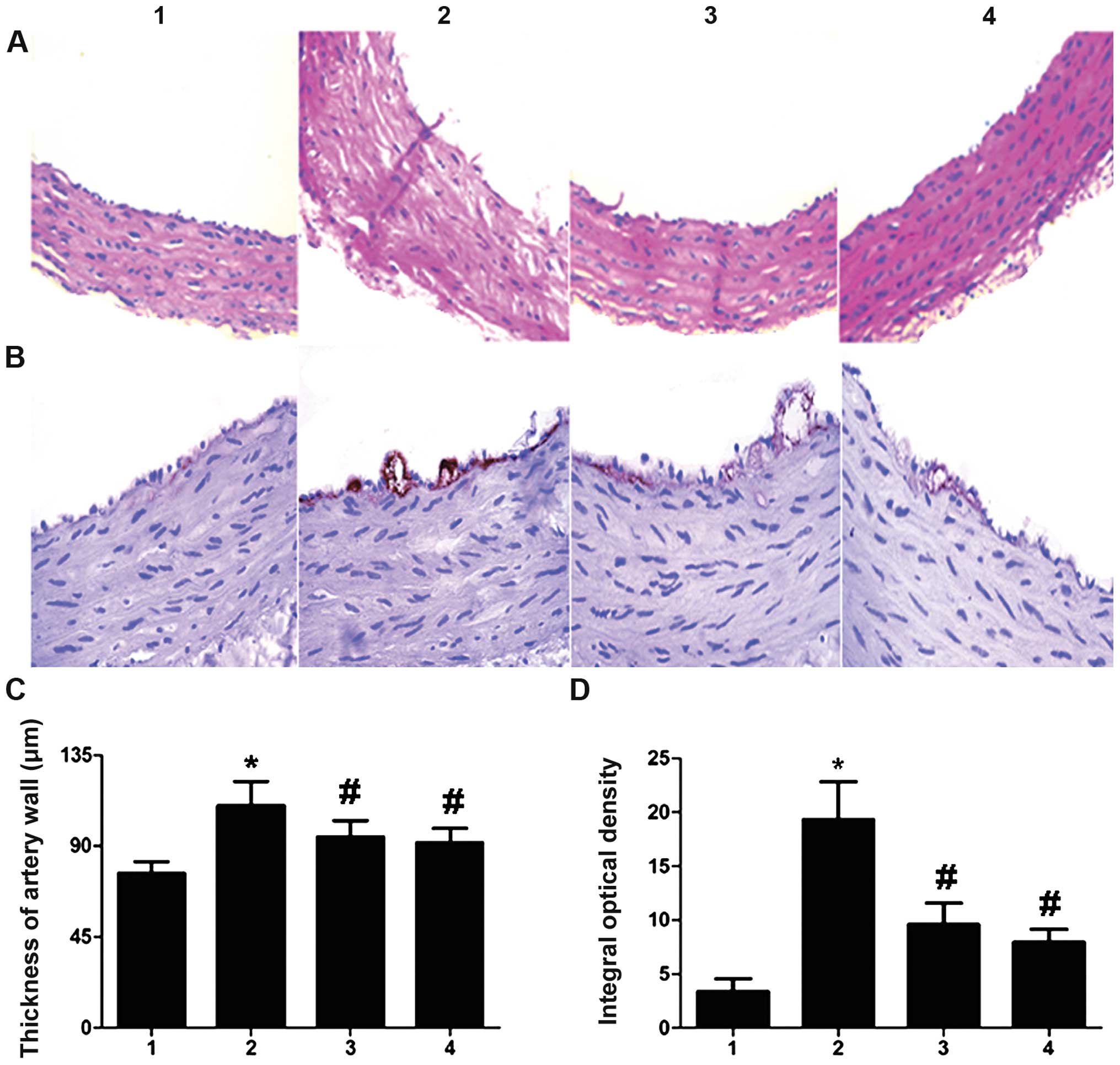
Morphological characteristics of rat
aortic tissue
We examined the morphological characteristics of the
arterial lesions in the H&E-stained tissue sections. The
endothelial cells adhering to inner elastic plates showed
structural changes in the DM group compared with those in the
control group. Indeed, the vascular smooth muscle cells (VSMCs) of
the tunica media were arranged irregularly, as shown by irregular
nuclei and uneven staining of the cytoplasm, and the media layer
thickness of the aorta was also markedly increased. However, the
administration of sitagliptin attenuated the remodeling of the
aortic intima, corrected the disarrangement of the VSMCs and
reduced the media thickness of the aorta (Fig. 5A and C).
As shown in Fig. 5B
and D, a high ET-1 expression level was found in the intima of
the DM rats, in comparison with that in the control rats
(P<0.05). High- and low-dose administration of sitagliptin led
to a significant downregulation of ET-1 expression in the intima of
the arteries (P<0.05).
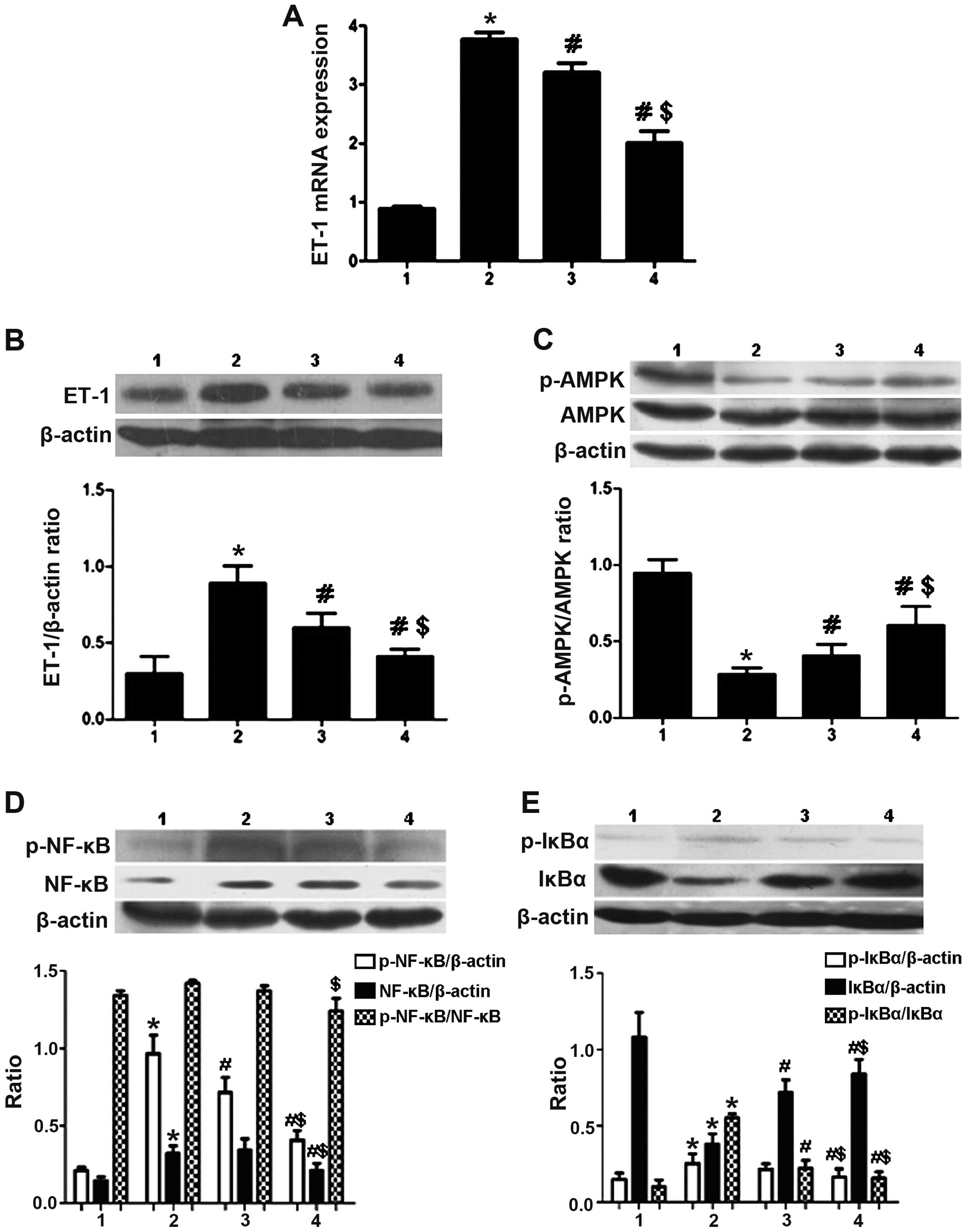
Relative gene and protein expression of
signaling pathway components
As shown in Fig. 6A
and B, hyperglycaemia and dyslipidaemia stimulated ET-1
expression at the transcriptional and translational levels, whereas
the administration of sitagliptin significantly inhibited ET-1
expression (P<0.05). Western blot analysis also revealed that
the rats in the DM group exhibited markedly enhanced expression of
p-NF-κB, NF-κB and p-IκBα (P<0.05), and reduced expression of
p-AMPK as well as IκBα (P<0.05) (Fig. 6C–E). Notably, the administration
of sitagliptin reversed these effects on protein expression
(P<0.05), thus, reversing the detrimental effects of
hyperglycaemia and dyslipidaemia in the rat aorta.
Discussion
In the present study, we utilised streptozotocin
injection to induce a defect in insulin secretion and a HF diet to
induce an insulin-resistant state in a rat model of type 2
diabetes. We have demonstrated that different doses of the DPP-4
inhibitor sitagliptin ameliorated glucose metabolism, insulin
resistance and hemodynamic dysfunction to varying extents. Most
importantly, sitagliptin markedly inhibited ET-1 expression at the
transcriptional and translational levels, as well as ET-1-mediated
endothelial dysfunction in the thoracic aorta samples isolated from
rats with streptozotocin-induced diabetes. We suggest that the
mechanism responsible for these effects potentially involves the
activation of the AMPK signaling pathway through the elevation of
GLP-1 levels, which subsequently suppresses the NF-κB/IκBα
system.
Several studies have indicated that DPP-4 inhibitors
and GLP-1 analogues may exert beneficial effects on glucose
metabolism and insulin resistance (20,21). Our results showed that sitagliptin
decreased FBG levels and alleviated insulin resistance in a
dose-dependent manner, as GLP-1, which was elevated by the
administration of sitagliptin to the diabetic rats, preserved
plasma glucose homeostasis and protected β-cells in the pancreatic
islets. Due to the enhancement of glucose metabolism, the weights
of the rats in the treatment groups were also increased compared
with the untreated rats. The lipid data were roughly similar to the
findings of previous studies (12,14), which reported that GLP-1 had
minimal effects on the TG, Tch and LDL levels induced by a HF diet.
The minimal changes in the lipid profiles following the
administration of sitagliptin to diabetic rats, suggest that the
vasoprotective and cardioprotective effects of sitagliptin may not
be directly associated with its effects on lipid metabolism.
Emerging evidence (4,22)
has shown that diabetes, a coronary heart disease (CHD) equivalent,
is closely associated with endothelial dysfunction, specifically
with regard to the inability of the endothelium to regulate
vascular homeostasis, which is a hallmark of vascular disease and
atherosclerosis. The proposed mechanisms which are responsible for
the endothelial damage occurring in diabetes, dependently and
independently of the damage due to other cardiovascular risk
factors, include hyperglycemia, insulin resistance, excessive free
fatty acids (FFAs), oxidative stress and low-grade systemic
inflammation (4,5). Endothelial dysfunction is
characterized by subdued vasodilation through reduced NO synthesis
which induces platelet aggregation, smooth muscle cell
proliferation and the nuclear transcription of leukocyte-adhesion
molecules, as well as augmented vasoconstriction (4,5).
The endothelium secretes the potent vasoconstrictor ET-1 which
promotes the development of cardiovascular disease through its
pro-inflammatory and mitogenic effects (23). Current evidence links endothelial
dysfunction with type 1 and type 2 diabetes, by demonstrating
impaired endothelial-dependent vasorelaxation (4). Indeed, data from the present study
has demonstrated that the serum NO levels were markedly reduced and
the serum ET-1 levels were significantly increased in the diabetic
rats. Particularly, diabetes promotes ET-1 expression in the aorta.
Furthermore, aortic rings were used to prove that Ach-induced
endothelium-dependent vasodilation was severely impaired in
diabetes and the contractile response to Phe was also much smaller
as previously reported (24),
indicating that arterial remodeling in the diabetic rats caused
decreased elasticity and compliance of the aorta as well as reduced
endothelial function.
As a member of a new class of anti-diabetic drugs,
sitagliptin enhances glucose metabolism by increasing the active
concentration and duration of action of GLP-1. It has been
previously reported that GLP-1 reduces the progression of
atherosclerosis by attenuating endothelial dysfunction (8–10,12–15) and we have further verified that
sitagliptin enhances endothelial-dependent vasorelaxation in a
dose-dependent manner. Previous studies have emphasized that GLP-1
stimulated NO production by enhancing the expression of eNOS
(14,15). In the present study, it is the
first time to the best of our knowledge that a study has mainly
focused on the effect of GLP-1 on ET-1 expression in diabetic rats.
Firstly, the results revealed that sitagliptin markedly enhanced
serum GLP-1 levels and inhibited serum ET-1 levels. Furthermore, as
ET-1 expression is mainly modulated at a transcriptional level, we
examined ET-1 expression at both the transcriptional and
translational levels in the rat aortas, and the data showed that
ET-1 expression was inhibited by sitagliptin, which partly
accounted for the attenuation of endothelial dysfunction. These
findings provide additonal evidence for the cardioprotective effect
of GLP-1.
Previous investigations have identified the factors
which regulate ET-1, which may be partly mediated by NF-κB
(25,26). Elevated levels of advanced
glycation end products (AGEs) have been demonstrated to induce ET-1
expression (26). In addition,
high concentrations of glucose have been shown to induce NF-κB
activation in human umbilical vein endothelial cells (HUVECs)
(27). Quehenberger et al
(26) have found that the
transient transfection of bovine aortic endothelial cells (BAECs)
with plasmids overexpressing IκBα led to a partial inhibition of
AGE-mediated ET-1 expression, and transient transfection with
plasmids overexpressing NF-κB p50 and p65 resulted in the induction
of ET-1 expression, which suggested that AGE-mediated ET-1
induction is controlled by NF-κB. In animal models, Piechota and
Goraca (25) have also
demonstrated that NF-κB inhibition plays a protective role in
ET-1-induced oxidative lung injury, highlighting the important role
of the NF-κB pathway in the ET-1-induced overproduction of ROS. As
a transcriptional regulator of NF-κB, IκB kinase binds to and
stabilizes NF-κB. Following phosphorylation, IκB kinase is degraded
which results in the stimulation of NF-κB phosphorylation (28). The involvement of the
NF-κB/IκB/ET-1 pathway is consistent with our data regarding the
enhancement of the phosphorylation of IκBα and NF-κB, as well as
the increased expression of the downstream target ET-1 in diabetic
rat aortas.
It is well documented that exendin-4 inhibits
monocyte adhesion to endothelial cells, and ameliorates
atherosclerotic lesions in apolipoprotein E-deficient mice, by
suppressing NF-κB activation in macrophages (29). Additionally, it has been
previously reported that liraglutide exerted an anti-inflammatory
effect on HUVECs by increasing NO generation and inhibiting NF-κB
activation, which was blocked by an AMPK inhibitor, suggesting that
the action of liraglutide on NF-κB may occur partly through AMPK
activation (30). In vivo,
it has been reported that sitagliptin attenuates the progression of
atherosclerosis in apolipoprotein-E knockout mice through
AMPK-dependent mechanisms (31).
Thus, we hypothesized that sitagliptin enhances serum GLP-1 levels,
which may inhibit NF-κB activation by regulating the AMPK signaling
pathway in the aorta. To verify this hypothesis in vivo, we
established a rat model of streptozotocin-induced diabetes and
administered sitagliptin for 12 weeks. The suppression of IκBα and
NF-κB phosphorylation as well as the activation of AMPK in rat
aortas were observed in the high-dose sitagliptin group. Taking
into account the finding that ET-1 expression is inhibited, we
suggest that the downregulation of ET-1 expression by sitagliptin
in the aortic endothelium is closely associated with the
AMPK/NF-κB/IκBα signaling pathway. This conclusion is partly
supported by a previous in vitro study, which demonstrated
that liraglutide activated GLP-1 receptors and suppressed ET-1
expression in high glucose-cultured HUVECs by inhibiting NF-κB
phosphorylation and NF-κB translocation from the cytoplasm to the
nucleus (32). However, the
downregulation of ET-1 expression in the aortic endothelium may be
associated with the decreased glucose levels in the treatment
groups. However, an in vitro study by Dai et al
(32) has noted that the effects
of GLP-1 on ET-1 expression and upstream signaling pathways may
occur independently of glucose metabolism, to a certain extent.
In summary, the present study has confirmed that
sitagliptin apparently attenuates endothelial dysfunction and
delays the progression of atherosclerosis in a dose-dependent
manner in an animal model of diabetes. It is noteworthy that the
findings of our study indicate that sitagliptin inhibits ET-1
expression in the aortic endothelium by suppressing the NF-κB/IκBα
system through the activation of the AMPK pathway. By determining
the effect of sitagliptin on ET-1 expression in vivo we have
identified a novel mechanism which contributes to our current
understanding of the way in which GLP-1 attenuates endothelial
dysfunction.
Acknowledgments
The present study was funded by the National Natural
Science Foundation of China (nos. 81470568, 81270372 and 81070232),
the Anhui Provincial Natural Science Foundation (no. 1608085MH168)
and the Research Project for Practice Development of National TCM
Clinical Research Bases (no. JDZX2015133).
References
|
1
|
Wang J, Luben R, Khaw KT, Bingham S,
Wareham NJ and Forouhi NG: Dietary energy density predicts the risk
of clinically incident type 2 diabetes: the EPIC-Norfolk study.
Diabetes Care. 31:2120–2125. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Wilmot EG, Edwardson CL, Achana FA, Davies
MJ, Gorely T, Gray LJ, Khunti K, Yates T and Biddle SJ: Sedentary
time in adults and the association with diabetes, cardiovascular
disease and death: systematic review and meta-analysis.
Diabetologia. 55:2895–2905. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li J, Dong Y, Wu T and Tong N: Differences
between western and Asian type 2 diabetes patients in the incidence
of vascular complications and mortality: A systematic review of
randomized controlled trials on lowering blood glucose. J Diabetes.
Dec 11–2015.Epub ahead of print. View Article : Google Scholar
|
|
4
|
Sena CM, Pereira AM and Seiça R:
Endothelial dysfunction - a major mediator of diabetic vascular
disease. Biochim Biophys Acta. 1832:2216–2231. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Roberts AC and Porter KE: Cellular and
molecular mechanisms of endothelial dysfunction in diabetes. Diab
Vasc Dis Res. 10:472–482. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Drucker DJ: Glucagon-like peptides.
Diabetes. 47:159–169. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Luft VC, Schmidt MI, Pankow JS, Hoogeveen
RC, Couper D, Heiss G and Duncan BB; Atherosclerosis Risk in
Communities Investigators: Dipeptidyl peptidase IV and incident
diabetes: the Atherosclerosis Risk in Communities (ARIC) study.
Diabetes Care. 33:1109–1011. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Matheeussen V1, Waumans Y, Martinet W, Van
Goethem S, Van der Veken P, Scharpé S, Augustyns K, De Meyer GR and
De Meester I: Dipeptidyl peptidase in atherosclerosis: expression
and role in macrophage differentiation, activation and apoptosis.
Basic Res Cardiol. 108(350)2013. View Article : Google Scholar
|
|
9
|
Ban K, Noyan-Ashraf MH, Hoefer J, Bolz SS,
Drucker DJ and Husain M: Cardioprotective and vasodilatory actions
of glucagon-like peptide 1 receptor are mediated through both
glucagon-like peptide 1 receptor-dependent and -independent
pathways. Circulation. 117:2340–2350. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Nyström T1, Gutniak MK, Zhang Q, Zhang F,
Holst JJ, Ahrén B and Sjöholm A: Effects of glucagon-like peptide-1
on endothelial function in type 2 diabetes patients with stable
coronary artery disease. Am J Physiol Endocrinol Metab.
287:E1209–E1215. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Noyan-Ashraf MH, Shikatani EA, Schuiki I,
Mukovozov I, Wu J, Li RK, Volchuk A, Robinson LA, Billia F, Drucker
DJ and Husain M: A glucagon-like peptide-1 analog reverses the
molecular pathology and cardiac dysfunction of a mouse model of
obesity. Circulation. 127:74–85. 2013. View Article : Google Scholar
|
|
12
|
Vittone F, Liberman A, Vasic D, Ostertag
R, Esser M, Walcher D, Ludwig A, Marx N and Burgmaier M:
Sitagliptin reduces plaque macrophage content and stabilises
arteriosclerotic lesions in Apoe (−/−) mice.
Diabetologia. 55:2267–2275. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shah Z1, Kampfrath T, Deiuliis JA, Zhong
J, Pineda C, Ying Z, Xu X, Lu B, Moffatt-Bruce S, Durairaj R, et
al: Long-term dipeptidyl-peptidase 4 inhibition reduces
atherosclerosis and inflammation via effects on monocyte
recruitment and chemotaxis. Circulation. 124:2338–2349. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Matsubara J, Sugiyama S, Sugamura K,
Nakamura T, Fujiwara Y, Akiyama E, Kurokawa H, Nozaki T, Ohba K,
Konishi M, et al: A dipeptidyl peptidase-4 inhibitor,
des-fluoro-sitagliptin, improves endothelial function and reduces
atherosclerotic lesion formation in apolipoprotein E-deficient
mice. J Am Coll Cardiol. 59:265–276. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Han L, Yu Y, Sun X and Wang B: Exendin-4
directly improves endothelial dysfunction in isolated aortas from
obese rats through the cAMP or AMPK-eNOS pathways. Diabetes Res
Clin Pract. 97:453–460. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Chen YX, Wang XQ, Fu Y, Yao YJ, Kong MY,
Nie RQ and Wang JF: Pivotal role of inflammation in vascular
endothelial dysfunction of hyperlipidemic rabbit and effects by
atorvastatin. Int J Cardiol. 146:140–144. 2011. View Article : Google Scholar
|
|
17
|
Drolet MC, Plante E, Battistini B, Couet J
and Arsenault M: Early endothelial dysfunction in cholesterol-fed
rabbits: a non-invasive in vivo ultrasound study. Cardiovasc
Ultrasound. 2(10)2004. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Tang ST, Su H, Zhang Q, Tang HQ, Wang CJ,
Zhou Q, Wei W, Zhu HQ and Wang Y: Melatonin attenuates aortic
endothelial permeability and arteriosclerosis in
streptozotocin-induced diabetic rats: possible role of MLCK- and
MLCP-dependent MLC phosphorylation. J Cardiovasc Pharmacol Ther.
21:82–92. 2016. View Article : Google Scholar
|
|
19
|
Zhou B1, Pan Y, Hu Z, Wang X, Han J, Zhou
Q, Zhai Z and Wang Y: All-trans-retinoic acid ameliorated high fat
diet-induced atherosclerosis in rabbits by inhibiting platelet
activation and inflammation. J Biomed Biotechnol. 2012(259693)2012.
View Article : Google Scholar
|
|
20
|
Picatoste B1, Ramírez E, Caro-Vadillo A,
Iborra C, Ares-Carrasco S, Egido J, Tuñón J and Lorenzo O:
Sitagliptin reduces cardiac apoptosis, hypertrophy and fibrosis
primarily by insulin-dependent mechanisms in experimental type-II
diabetes. Potential roles of GLP-1 isoforms. PLoS One.
8:e783302013. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Zhao HJ, Xu Q, Zhang GY, Liu Y, Yu T, Li
HY, Yang LN, Wang HY and Xie LB: Therapeutic effects of GLP-1
analog on rats with experimental diabetes mellitus. J Jilin
University. 33:474–479. 2007.
|
|
22
|
Pasterkamp G: Methods of accelerated
atherosclerosis in diabetic patients. Heart. 99:743–749. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Böhm F and Pernow J: The importance of
endothelin-1 for vascular dysfunction in cardiovascular disease.
Cardiovasc Res. 76:8–18. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Cicek FA, Kandilci HB and Turan B: Role of
ROCK upregulation in endothelial and smooth muscle vascular
functions in diabetic rat aorta. Cardiovasc Diabetol. 12(51)2013.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Piechota A and Goraca A: Influence of
nuclear factor-κB inhibition on endothelin-1 induced lung edema and
oxidative stress in rats. J Physiol Pharmacol. 62:183–188.
2011.PubMed/NCBI
|
|
26
|
Quehenberger P, Bierhaus A, Fasching P,
Muellner C, Klevesath M, Hong M, Stier G, Sattler M, Schleicher E,
Speiser W and Nawroth PP: Endothelin 1 transcription is controlled
by nuclear factor-kappaB in AGE- stimulated cultured endothelial
cells. Diabetes. 49:1561–1570. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Ho FM, Lin WW, Chen BC, Chao CM, Yang CR,
Lin LY, Lai CC, Liu SH and Liau CS: High glucose-induced apoptosis
in human vascular endothelial cells is mediated through NF-kappaB
and c-Jun NH2-terminal kinase pathway and prevented by PI3K/Akt
eNOS pathway. Cell Signal. 18:391–399. 2006. View Article : Google Scholar
|
|
28
|
Suzuki J, Ogawa M, Muto S, Itai A, Isobe
M, Hirata Y and Nagai R: Novel IkB kinase inhibitors for treatment
of nuclear factor-kB-related diseases. Expert Opin Investig Drugs.
20:395–405. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Arakawa M, Mita T, Azuma K, Ebato C, Goto
H, Nomiyama T, Fujitani Y, Hirose T, Kawamori R and Watada H:
Inhibition of monocyte adhesion to endothelial cells and
attenuation of atherosclerotic lesion by a glucagon-like peptide-1
receptor agonist, exendin-4. Diabetes. 59:1030–1037. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hattori Y, Jojima T, Tomizawa A, Satoh H,
Hattori S, Kasai K and Hayashi T: A glucagon-like peptide-1 (GLP-1)
analogue, liraglutide, upregulates nitric oxide production and
exerts anti-inflammatory action in endothelial cells. Diabetologia.
53:2256–2563. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zeng Y, Li C, Guan M, Zheng Z, Li J, Xu W,
Wang L, He F and Xue Y: The DPP-4 inhibitor sitagliptin attenuates
the progress of atherosclerosis in apolipoprotein-E-knockout mice
via AMPK-and MAPK-dependent mechanisms. Cardiovasc Diabetol.
13(32)2014. View Article : Google Scholar
|
|
32
|
Dai Y, Mehta JL and Chen M: Glucagon-like
peptide-1 receptor agonist liraglutide inhibits endothelin-1 in
endothelial cell by repressing nuclear factor-kappa B activation.
Cardiovasc Drugs Ther. 27:371–380. 2013. View Article : Google Scholar : PubMed/NCBI
|