Introduction
Inflammation is one of most important and ubiquitous
defensive reactions to stimuli, such as toxins and pathogens, and
it is characterized by redness, pain, swelling, and a sensation of
heat at the site of infection (1). During the inflammatory process, a
number of inflammatory cells infiltrate the damaged tissue and
produce inflammatory mediators that exaggerate inflammatory
responses. Nitric oxide (NO), prostaglandin (PG)E2,
interleukins (ILs) and tumor necrosis factor-α (TNF-α), produced by
the infiltrated cells or damaged tissue, act as pleiotropic
effector molecules to amplify acute inflammation and lead to the
activation of adaptive immune responses (2,3).
Thus, it is essential that the inflammatory process be controlled
spatiotemporally to prevent unwanted tissue damage and the
development of inflammation-associated disorders.
Macrophages play a central role in inflammatory
diseases and produce many pro-inflammatory cytokines, such as IL-6,
TNF-α and IL-1β. Macrophages also produce inflammatory mediators,
including NO and PGE2 (4). As the major component of the outer
membrane of Gram-negative bacteria cell walls, lipopolysaccharide
(LPS) is capable of stimulating macrophages to produce
pro-inflammatory cytokines, including TNF-α and IL-6, and
inflammatory mediators such as NO and PGE2 (5). NO is produced by inducible nitric
oxide synthase (iNOS) and mediates many disease processes including
atherosclerosis, inflammation, carcinogenesis, hypertension,
obesity and diabetes (6–8). PGE2 is a major
cyclooxygenase (COX) product at inflammatory sites, where it
contributes to local increases in blood flow, edema formation and
pain sensitization (9). Thus, the
inhibition of these pro-inflammatory mediators may be an effective
strategy for the treatment of inflammatory diseases.
Lagerstroemia ovalifolia Teijsm. &
Binn
(LO) is a member of the Lythraceae family (10,11). This tree species has commercial
value. It is used for medium-heavy construction (door, window
frames), bridge and boat building, various agriculture implements
and tool handles. LO has traditionally been used as an herbal
medicine in Java without distinction of related species (e.g.,
L. speciosa) without distinction of species; the bark for
diarrhea, the leaves for malaria and dermatosis (12,13). The effects and biological activity
of the methanolic extract of LO (LOME) on inflammation, as well as
the mechanisms of action responsible for these effects remain
largely unknown. To date, this plant has shown few and unclear
effects to the best of our knowledge. In the present study, we
examined the anti-inflammatory effects of LOME in various
experiments. We used an LPS-stimulated, murine macrophage cell
line, RAW264.7, as a model of inflammation to determine the effects
of LOME. The binding of LPS to downstream signaling cascades,
including mitogen-activated protein kinases (MAPKs) and nuclear
factor-κB (NF-κB) pathways (14),
led to the production of inflammatory mediators from macrophages,
such as TNF-α, IL-1β, IL-6, PGE2 and NO. Our results
indicated that LOME inhibited NO, PGE2 and the
pro-inflammatory cytokines in LPS-stimulated macrophages through
NF-κB signaling, as well as the MAPK, extracellular
signal-regulated kinase (ERK)1/2, p38 and c-Jun N-terminal kinase
(JNK)1/2 pathways. These results provide evidence for the use of
LOME as an anti-inflammatory treatment.
Materials and methods
Materials
LO was collected from the Ujung Kulon National Park,
in the province of Banten, Indonesia. Plant samples were collected
and identified by staff at the Center for Pharmaceutical and
Medical Technology (PTFM; Tangerang, Indonesia), and verified at
the Herbarium Bogoriense (LIPI; Bogor, Indonesia). Voucher
specimens recorded as KRIB 0038535 and PMT 537, have been deposited
in the herbarium (KRIB) of the Korea Research Institute of
Bioscience and Biotechnology (Daejeon, Korea) as well as in the
Center for Pharmaceutical and Medical Technology (PTFM) and the
Herbarium Bogoriense. After drying and grinding the leaves of LO,
the powder (10.8 kg) was added to methanol (100 liters). The
extraction was performed using the method of repercolation at room
temperature. The extract was filtered and concentrated by a rotary
evaporator (Rotavapor 4000; Heidolph, PT AbadiNusa Usahasemesta,
Jakarta) under reduced pressure, to obtain 1,030 g LOME. In
subsequent experiments, LOME was dissolved in dimethyl sulfoxide
(DMSO) at a concentration of 20 mg/ml, and then diluted to various
concentrations prior to use.
Cell culture
The murine macrophage cell line RAW264.7 was
obtained from the American Type Culture Collection (ATCC;
Rockville, MD, USA). The cells were grown in Dulbecco's modified
Eagle's medium (DMEM; Gibco-BRL, Grand Island, NY, USA)
supplemented with 10% heat-inactivated fetal bovine serum (FBS;
HyClone, Logan, UT, USA) and 1% antibiotic-antimycotic solution
(Invitrogen, Grand Island, NY, USA). The RAW264.7 cells were
maintained by weekly passage, and the cells were utilized for
experimentation at 60–80% confluence. Following the incubation of
the RAW264.7 cells for 4 h, 0–30 µg/ml LOME was added.
Determination of cell viability
Cell viability was determined by the
mitochondrial-dependent reduction of
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT;
Amresco, LLC, Solon, OH, USA) to purple formazan. Briefly, the
RAW264.7 cells were seeded into each well of 96-well plates at a
density of 1×104 cells/well. After 4 h, the cells were
treated with LOME for 24 h. MTT solution (5 µl; 5 mg/ml) was
then added to each well. After a 4 h incubation at 37°C with 5%
CO2, the supernatant was removed and the formazan
crystals formed by the viable cells were dissolved in 100 µl
DMSO. The absorbance of each well was read at 570 nm using a
microplate reader (Benchmark; Bio-Rad Laboratories, Inc., Hercules,
CA, USA). Based on the results of the cytotoxicity assay, we used a
nontoxic concentration of LOME on the RAW264.7 cells in subsequent
experiments.
NO assay
Nitrite concentration in the medium was measured as
an indicator of NO production according to the Griess reaction
method, as previously described (15). The RAW264.7 cells were plated at a
density of 2.5×105 cells/ml in 96-well plates, and then
incubated with or without LPS (0.5 µg/ml) in the absence or
presence of various concentrations of LOME for 24 h. The nitrite
accumulation in the supernatant was assessed by a Griess reaction.
Each 100 µl of the culture supernatant was mixed with an
equal volume of the Griess reagent [0.1%
N-(1-naphthyl)-ethylenediamine and 1% sulfanilamide in 5%
phosphoric acid] and incubated at room temperature for 10 min. The
absorbance was measured at 540 nm using a microplate reader
(Benchmark; Bio-Rad Laboratories, Inc.) and a series of known
concentrations of sodium nitrite were used as a standard.
PGE2 assay
The RAW264.7 cells were plated at a density of
2.5×105 cells/ml in a 96-well culture plate. The cells
were incubated with various concentrations (5, 10, 20 or 30
µg/ml) of LOME for 1 h, followed by LPS treatment (0.5
µg/ml) for 24 h. Following a 24 h incubation, the
PGE2 concentration in the culture media was quantified
using an enzyme-linked immunosorbent assay (ELISA) kit (Cayman
Chemical Co., Ann Arbor, MI, USA) according to manufacturer's
instructions.
Cytokine assays
The levels of IL-6, IL-1β and TNF-α were determined
using commercial ELISA kits. IL-6, IL-1β and TNF-α were purchased
from R&D Systems, Inc. (Minneapolis, MN, USA). The assay
procedure for each kit was conducted according to the
manufacturer's instructions. The concentrations of the mediators
were determined at 450 nm using a microplate reader (Benchmark;
Bio-Rad Laboratories, Inc.).
Semi-quantitative reverse transcription
(RT)-PCR
RT-PCR was performed to detect the mRNA expression
of iNOS, COX-2, IL-6, IL-1β, TNF-α and β-actin. Briefly, after LPS
(0.5 µg/ml) stimulation of the RAW264.7 cells for 6 h, total
RNA was isolated using TRIzol™ reagent (Invitrogen, Carlsbad, CA,
USA) according to the manufacturer's instructions. An RT reaction
was perormed using a kit for producing cDNA (Qiagen GmbH, Hilden,
Germany). PCR was performed using specific forward and reverse
primers, and a premix according to the manufacturer's instructions
(Bioneer Corporation, Daejeon, Korea). The PCR conditions for each
PCR reaction were as follows: 94°C for 5 min (1 cycle), 94°C for 30
sec, 60°C for 30 sec, and 72°C for 45 sec (for 30 cycles); and then
a final extension phase at 72°C for 10 min. The primer sequences
were as follows: TNF-α sense, 5′-CAT CTT CTC AAA ATT CGA GTG ACA
A-3′ and antisense, 5′-TGG GAG TAG ACA AGG TAC AAC CC-3′; IL-1β
sense, 5′-GTG TCT TTC CCG TGG ACC TT-3′ and antisense; 5′-TCG TTG
CTT GGT TCT CCT TG-3′; IL-6 sense, 5′-AAC GAT GAT GCA CTT GCA GA-3′
and antisense, 5′-GAG CAT TGG AAA TTG GGG TA-3′; iNOS sense, 5′-CAA
GAG TTT GAC CAG AGG ACC-3′ and antisense, 5′-TGG AAC CAC TCG TAC
TTG GGA-3′; COX-2 sense, 5′-GAA GTC TTT GGT CTG GTG CCT G-3′ and
anti-sense, 5′-GTC TGC TGG TTT GGA ATA GTT GC-3′; and β-actin
sense, 5′-TGT TTG AGA CCT TCA ACA CC-3′ and antisense, 5′-CGC TCA
TTG CCG ATA GTG AT-3′. β-actin expression was included as an
internal, housekeeping gene control. The reaction products were
separated by electrophoresis on a 1.5% agarose gel, stained with
EtBr, and visualized by UV transillumination (CoreBioSystem, Seoul,
Korea). The images were captured using an Olympus C4000 zoom camera
system (C4000; Olympus America Inc., Melville, NY, USA).
Western blot analysis
The RAW264.7 macrophages were pretreated with the
indicated concentrations of LOME or vehicle for 1 h and stimulated
with LPS (0.5 µg/ml) for 20 min (phosphor) and 18 h (COX-2
and iNOS). Proteins in each sample (30 µg of total protein)
were resolved on a 10% sodium dodecyl sulfate (SDS)-polyacrylamide
gel by SDS-polyacrylamide gel electrophoresis (PAGE). The proteins
were transferred to a polyvinylidene difluoride (PVDF) membrane and
exposed to the appropriate antibodies. Each membrane was incubated
for 1 h in 5% skim milk in TBS-T buffer (0.1 M Tris-HCl, pH 7.4,
0.9% NaCl and 0.1% Tween-20) to block non-specific binding, and
then incubated with primary antibodies that recognized p65 (cat.
no. sc-372, 1:1,000; Santa Cruz Biotechnology, Inc., Santa Cruz,
CA, USA); iκBα (cat. no. 9242, 1:1,000; Cell Signaling Technology,
Inc., Danvers, MA, USA), iNOS (cat. no. ADI-905-431, 1:1,000;
obtained from Enzo Life Sciences, Inc., Farmingdale, NY, USA);
COX-2 (cat. no. sc-1747, 1:1,000; Santa Cruz Biotechnology, Inc.);
β-actin (cat. no. 4967, 1:2,000) and PARP (cat. no. 9542) (both
from Cell Signaling Technology, Inc.); the total forms of ERK2
(sc-154), p38 MAPK (sc-7149), JNK1/3 (sc-474) (1:1,000, all from
Santa Cruz Biotechnology, Inc.); the phosphorylated forms of p38
MAPK (ADI-KAP-MA022) and JNK1/2 (KAP-SA011 (1:1,000; both from Enzo
Life Sciences, Inc.); and the phosphorylated forms of ERK (9106,
1:1,000; Cell Signaling Technology, Inc.). Each protein was
detected using an enhanced chemiluminescence (ECL) detection system
(Bio-Rad Laboratories, Inc.).according to manufacturer's
instructions
Immunocytochemistry
The RAW264.7 cells were cultured on Permanox plastic
chamber slides (Nalge Nunc International, Rochester, NY, USA) and
fixed in methanol at 4°C for 20 min. The slides were washed three
times with phosphate-buffered saline (PBS) and blocked with 3%
(w/v) BSA in PBS for a further 30 min. The slides were then
incubated for 24 h at 4°C with anti-iNOS (rabbit polyclonal IgG,
1:200 dilution), and anti-NF-κB p65 subunit (rabbit polyclonal IgG,
1:200 dilution) (both from Santa Cruz Biotechnology, Inc.)
antibodies. After washing to remove excess primary antibody, the
slides were further incubated with anti-rabbit Alexa Fluor
488-conjugated antibody (Invitrogen, Carlsbad, CA, USA) for 2 h at
room temperature, washed with PBS, and mounted using ProLong Gold
Antifade reagent containing 4′,6-diamidino-2-phenylin-dole (DAPI;
Invitrogen, Carlsbad, CA, USA) for 5 min, prior to the localization
and quantification of the nuclei. Subsequently, the slides were
cover-slipped and visualized using a confocal laser scanning
microscope (LSM510m; Carl Zeiss, Inc., Oberkochen, Germany). Images
of the samples were captured under the same exposure conditions,
and the nuclei were quantified from the images obtained.
Statistical analysis
For the statistical analyses, the values are
expressed as the means ± SEM of the sample determinations. The
statistical significance was determined using a two-tailed
Student's t-test for independent means, and a P-value <0.05 was
considered to indicate a statistically significant difference.
Results
Evaluation of the cytotoxicity of LOME in
RAW264.7 cells
A prerequisite to studying the biological activity
of LOME was to ensure that it did not exert a detrimental effect on
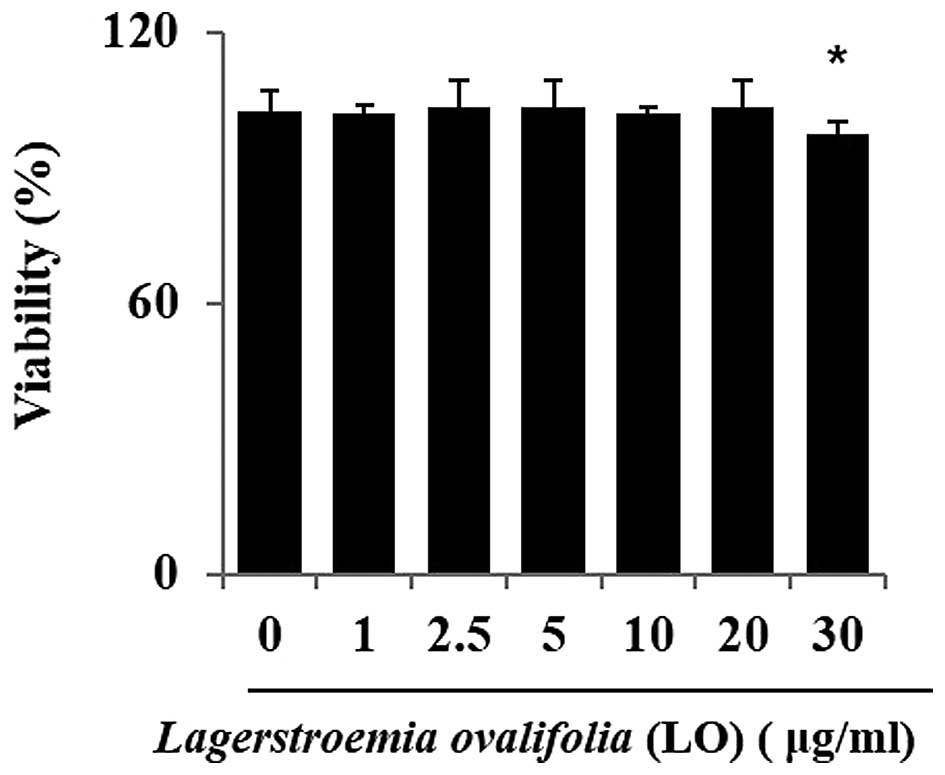
cell metabolism. To determine whether LOME affected cell viability,
the RAW264.7 cells were incubated for 24 h with the extract at a
wide range of concentrations (0–30 µg/ml), and cell
viability was evaluated using an MTT assay. A statistically
significant decrease in cell survival was observed at a
concentration of 30 µg/ml (Fig. 1).
LOME reduces the production of NO by
suppressing iNOS expression in LPS-stimulated RAW264.7 cells
As it is important to regulate the overproduction of
NO, which induces pro-inflammatory responses in inflammatory
disorders (16), we examined the
effect of LOME on the regulation of NO production in LPS-stimulated
macrophages. Prior to evaluating the regulatory effect of LOME in
LPS-stimulated macrophages, we first confirmed that LOME did not
induce NO production in the RAW264.7 cells. However, the
pre-incubation of cells with LOME for 1 h prior to LPS stimulation
resulted in reduced NO production compared with that in the
LPS-stimulated RAW264.7 cells. Furthermore, NO production was
gradually attenuated by increasing the concentration of LOME up to
30 µg/ml (Fig. 2A). To
determine whether LOME reduces NO production by regulating protein
expression, we measured the expression of iNOS in the RAW264.7
cells by RT-PCR and western blot analysis. According to this
analysis, iNOS expression was downregulated in a dose-dependent
manner (Fig. 2C and D). We
further examined the effect of LOME, which inhibits iNOS, using
immunocytochemistry. The iNOS levels were markedly increased in
LPS-stimulated cells (Fig. 2D);
however, pre-incubation with the LOME prevented the expression of
these proteins. Thus, we suggest that LOME inhibits iNOS
production.
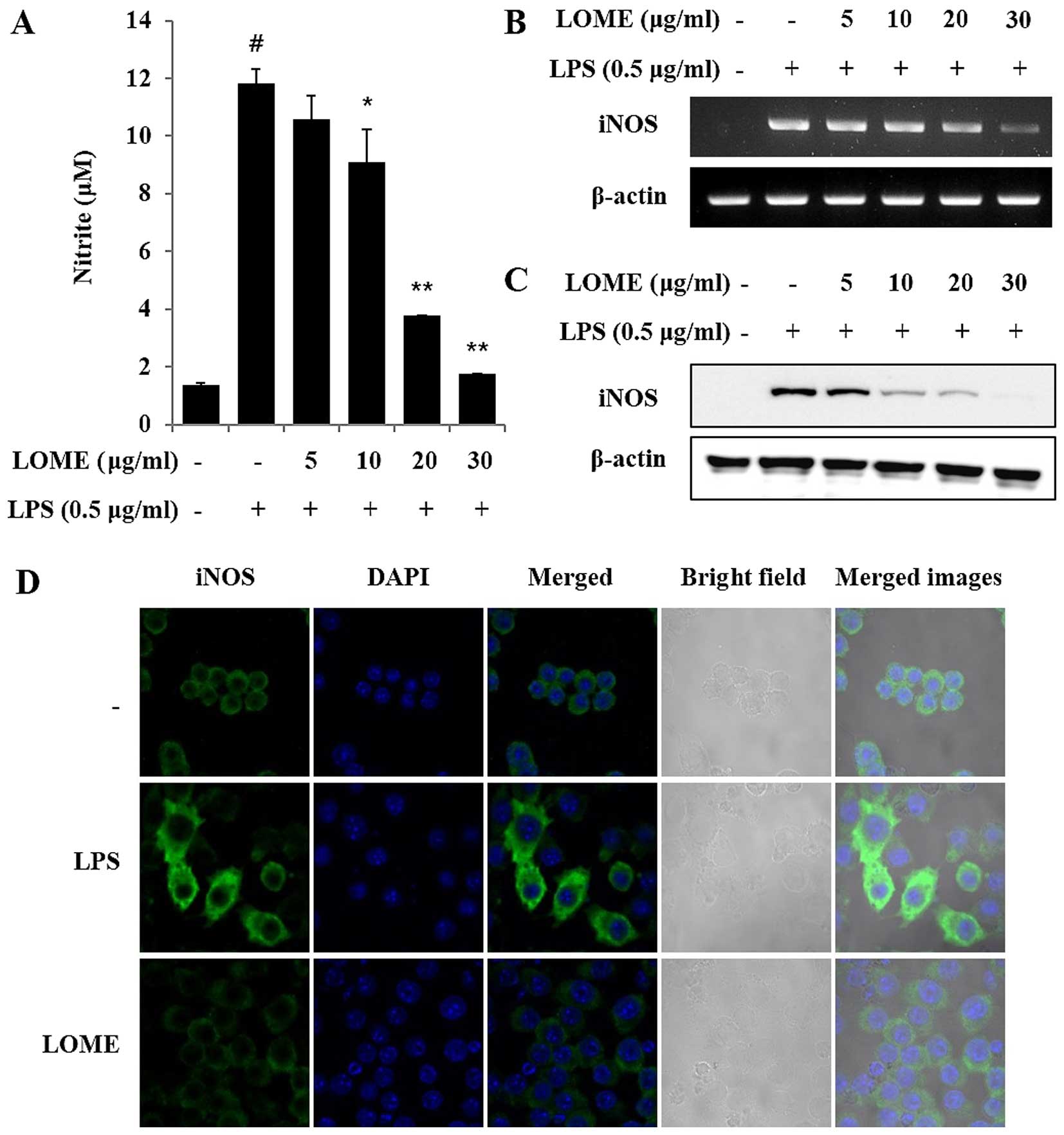 | Figure 2Methanolic extract of L.
ovalifolia (LOME) inhibits lipopolysaccharide (LPS)-induced
nitric oxide (NO) production and inducible nitric oxide synthase
(iNOS) expression in RAW264.7 cells. (A) RAW264.7 cells were
treated with the LOME (5, 10, 20 or 30 µg/ml) for 1 h and
then stimulated with LPS (0.5 µg/ml). The culture media were
collected at 24 h, and the NO concentrations were measured using
the Griess reaction. Three independent experiments were performed,
and the data are presented as the means ± SEM.
*P<0.05 and **P<0.01, compared with the
cells treated with LPS alone. The hash symbol (#) is used to
indicate the negative control, error range. (B) RAW264.7 cells were
treated with LOME for 1 h and stimulated with LPS for 6 h. β-actin
expression was used as an internal control for RT-PCR. (C) RAW264.7
cells were treated with LOME for 1 h and stimulated with LPS for 18
h. Cell lysates were obtained, the iNOS protein levels were
evaluated using western blot analysis, and β-actin expression was
used as an internal control. (D) The cells were treated with LOME
for 1 h and stimulated with LPS for a further 20 h. Cellular
localization of iNOS was determined by immunocytochemistry.
Following fixation, the cells were stained with blue and green
labels (magnification, x400). Control, DMSO (0.1%); LPS, LPS only
(0.5 µg/ml); LOME, LOME (30 µg/ml) and LPS. |
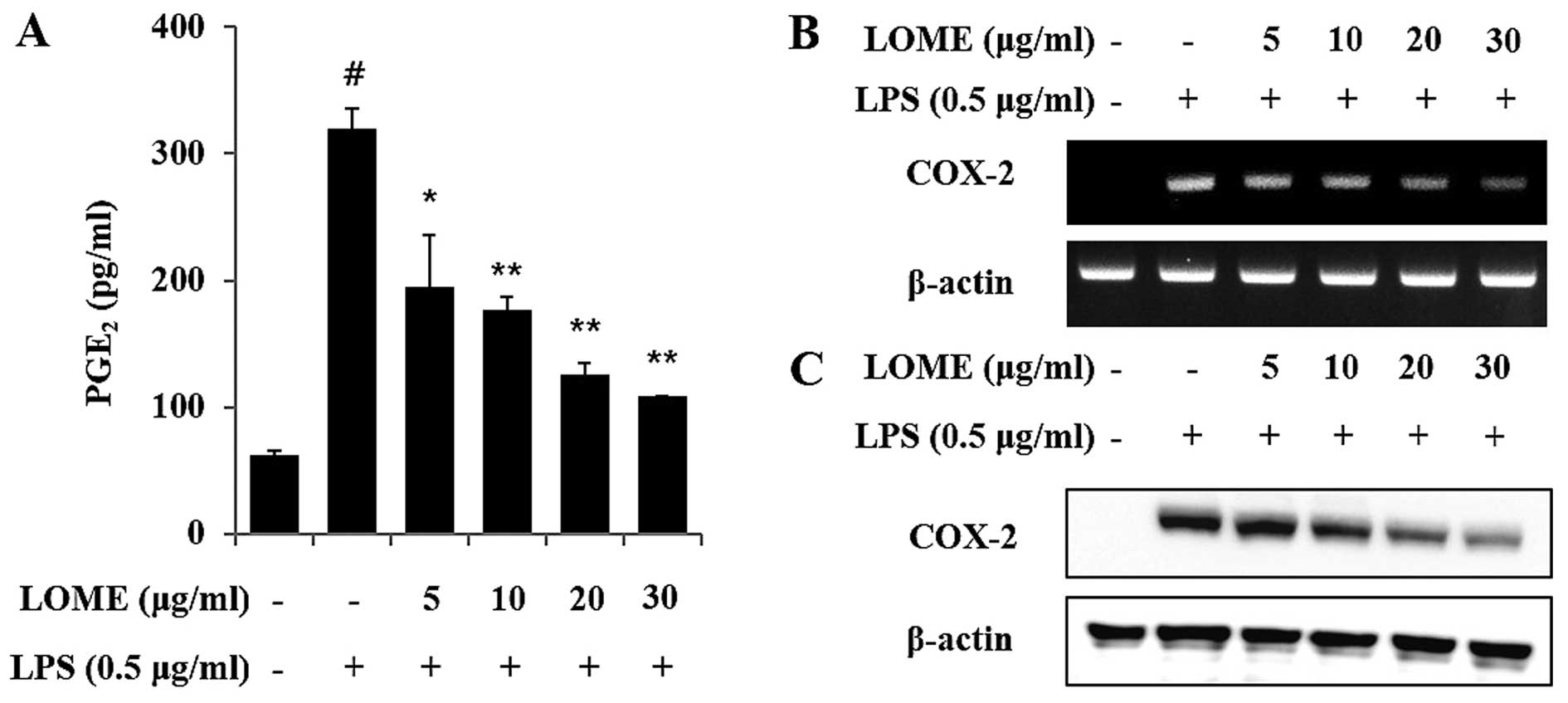
LOME decreases the production of
PGE2 by suppressing COX-2 expression in LPS-stimulated
RAW264.7 cells
The unstimulated RAW264.7 cells secreted basal
levels of PGE2 and LPS stimulation induced an increase
in PGE2 production (Fig.
3A). In a dose-response study, LOME treatment significantly
diminished LPS induction of COX-2, which corresponded with a
decreased accumulation of PGE2 in the culture media
(Fig. 3). The LPS-stimulated
RAW264.7 cells overexpressed COX-2 mRNA, whereas the LOME-treated
RAW264.7 cells stimulated with LPS, exhibited decreased expression
of COX-2 mRNA. COX-2 protein expression was consistent with the
mRNA results (Fig. 3B and C).
LOME inhibited COX-2 protein expression in the RAW264.7 cells
stimulated with LPS.
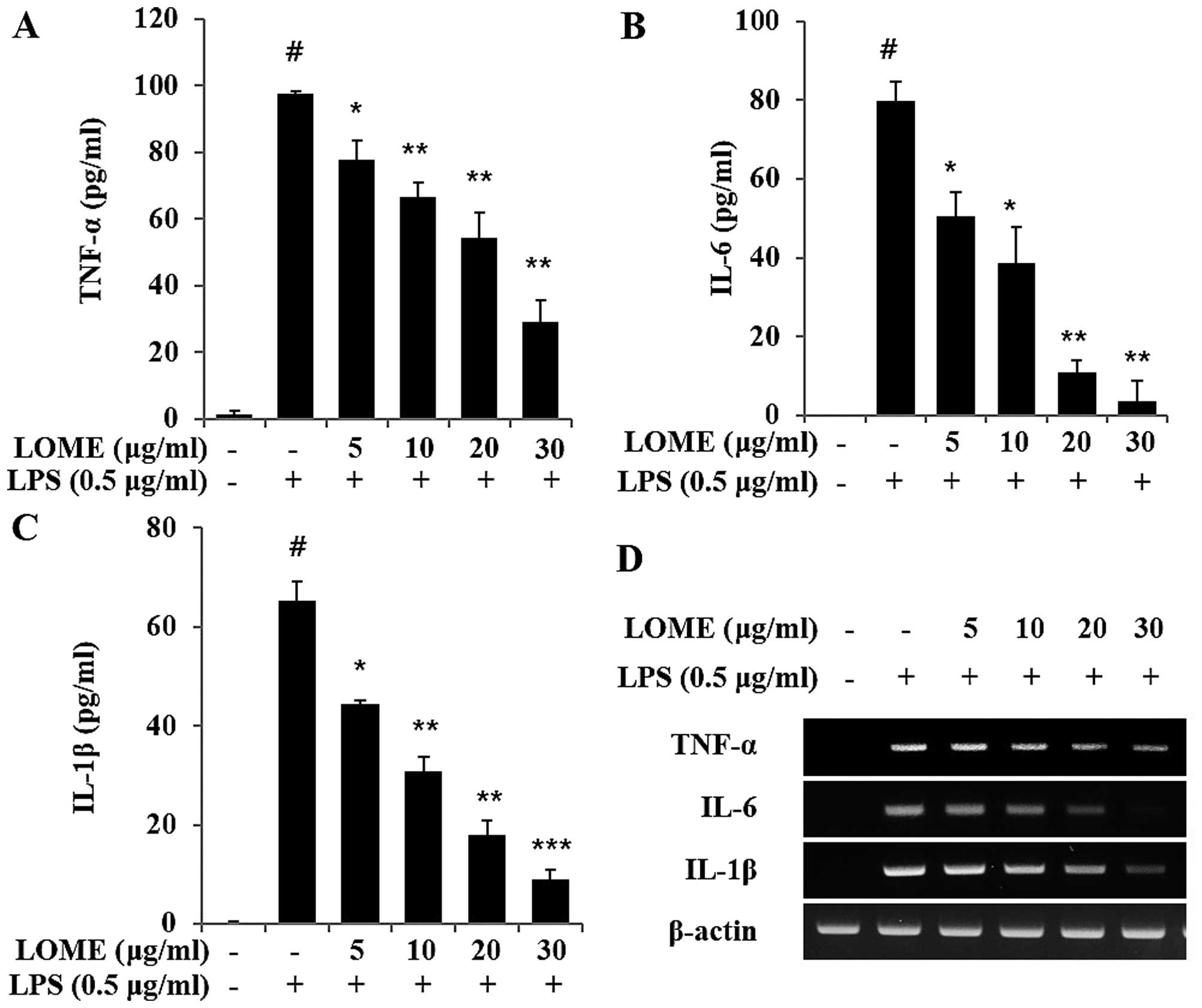
LOME reduces the release of
pro-inflammatory cytokines in LPS-stimulated RAW264.7 cells
The levels of TNF-α, IL-1β and IL-6 were measured in
the media of RAW264.7 cells that were treated with varying
concentrations of LOME, and then treated with 0.5 µg/ml of
LPS (Fig. 4A–C). It was found
that LOME significantly reduced the production of TNF-α, IL-1β and
IL-6 in a dose-dependent manner (Fig.
4A–C). LOME alone had no effect on TNF-α, IL-1β and IL-6
production in the RAW264.7 cells (data not shown). Additionally,
the mRNA levels of TNF-α, IL-1β and IL-6 were evaluated using
RT-PCR (Fig. 4D). Changes in the
mRNA expression of TNF-α, IL-1β and IL-6 were similar to those
observed in their protein expression levels.
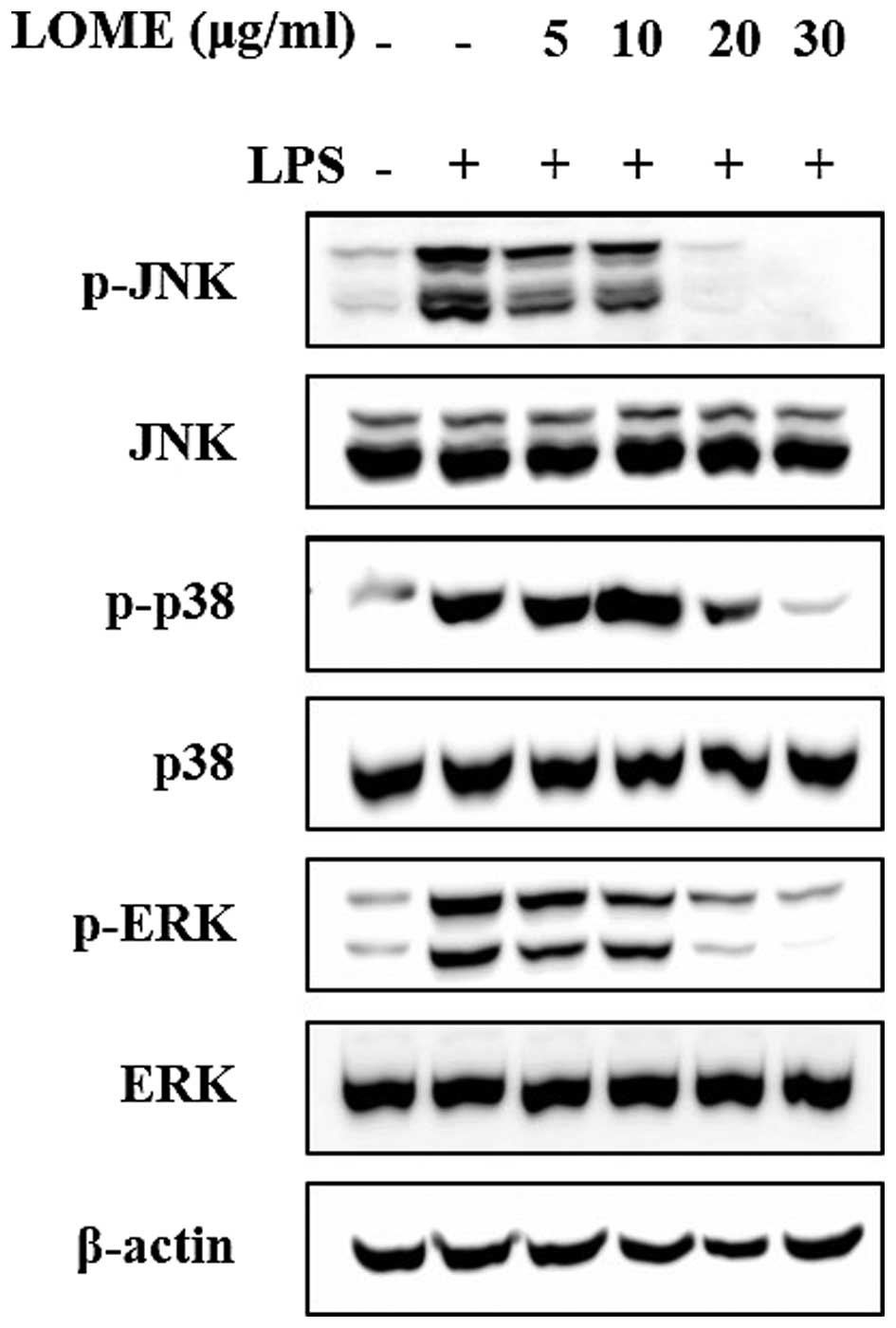
Effects of LOME on the activation of
MAPKs in LPS-stimulated RAW264.7 cells
Three MAPKs, namely ERK, p38 and JNK, are known to
be activated by LPS. MAPKs play an important role in the
transcriptional regulation of the LPS-induced expression of iNOS
and COX-2 by activating the transcription factor NF-κB (17). As shown in Fig. 5, LOME clearly decreased the
phosphorylation of ERK1/2, JNK and p38. These results indicate that
the inhibitory effect of LOME on TNF-α, IL-1β, IL-6, NO and
PGE2 may have been mediated by the downstream MAPK
pathways.
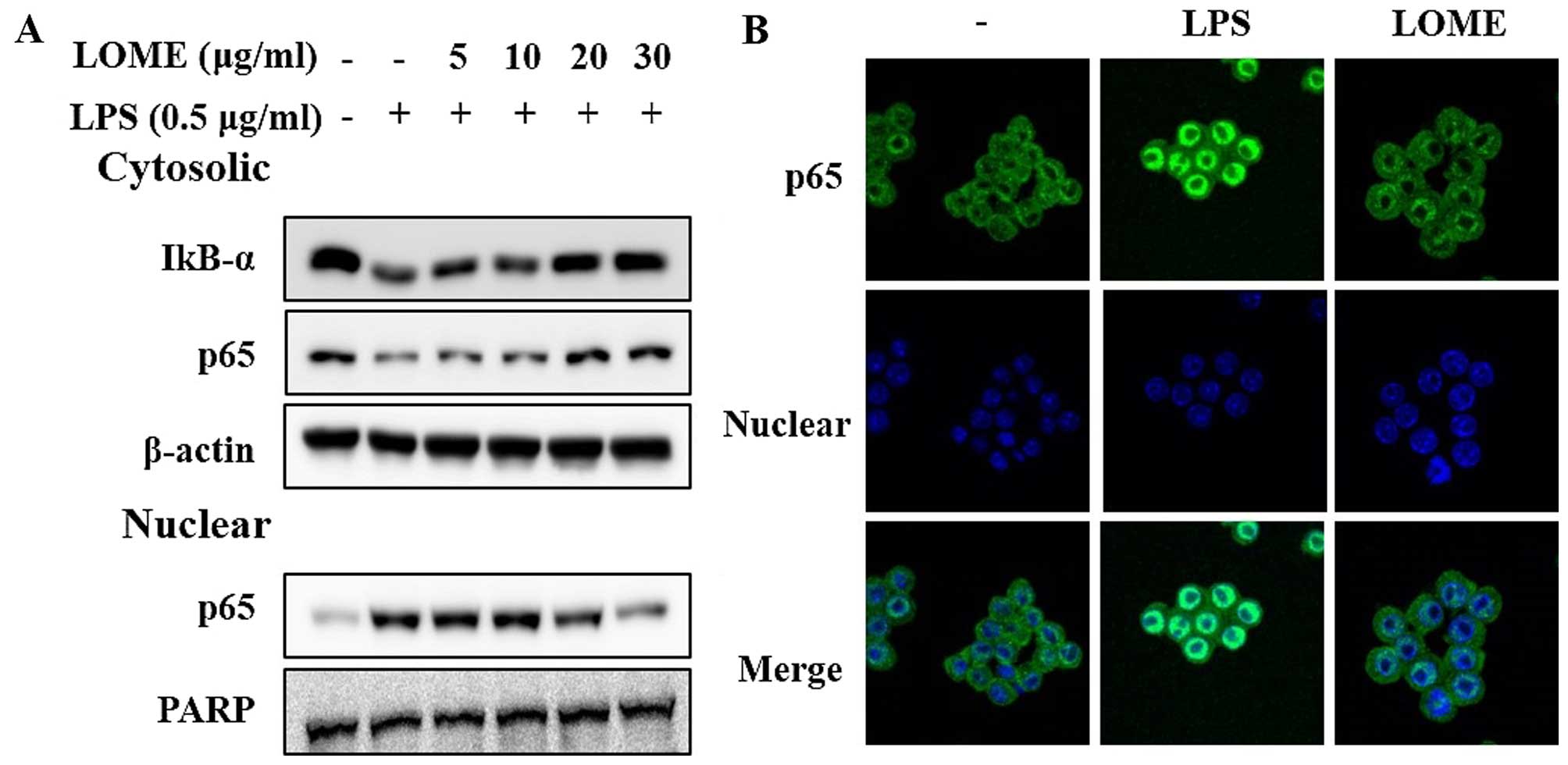
Effects of LOME on LPS-stimulated NF-κB
transcriptional activity through the suppression and nuclear
translocation of the p65 subunit in RAW264.7 cells
NF-κB plays a pivotal role in regulating the
expression of iNOS, COX-2 and inflammatory cytokines such as TNF-α,
IL-1β and IL-6 (18). As a
plausible molecular mechanism for the inhibition of the
inflammatory response, the effect of LOME on the NF-κB signaling
pathway was explored (Fig. 6A).
LPS exposure (0.5 µg/ml) for 30 min facilitated the
degradation of IκB-α, the phosphorylation of IκB-α, and the nuclear
accumulation of NF-κB. However, pre-treatment with LOME inhibited
the degradation of IκB-α. Furthermore, LOME decreased the
LPS-mediated phosphorylation of IκB-α and nuclear NF-κB
accumulation in a dose-dependent manner. We further investigated
the effect of the LOME on NF-κB activation using
immunocytochemistry. The LPS-stimulated translocation of the NF-κB
p65 subunit from the cytosol to the nucleus in the RAW264.7 cells
(Fig. 6B), was inhibited by LOME.
Thus, the anti-inflammatory effect of LOME may be due to inhibition
of the NF-κB signaling pathway.
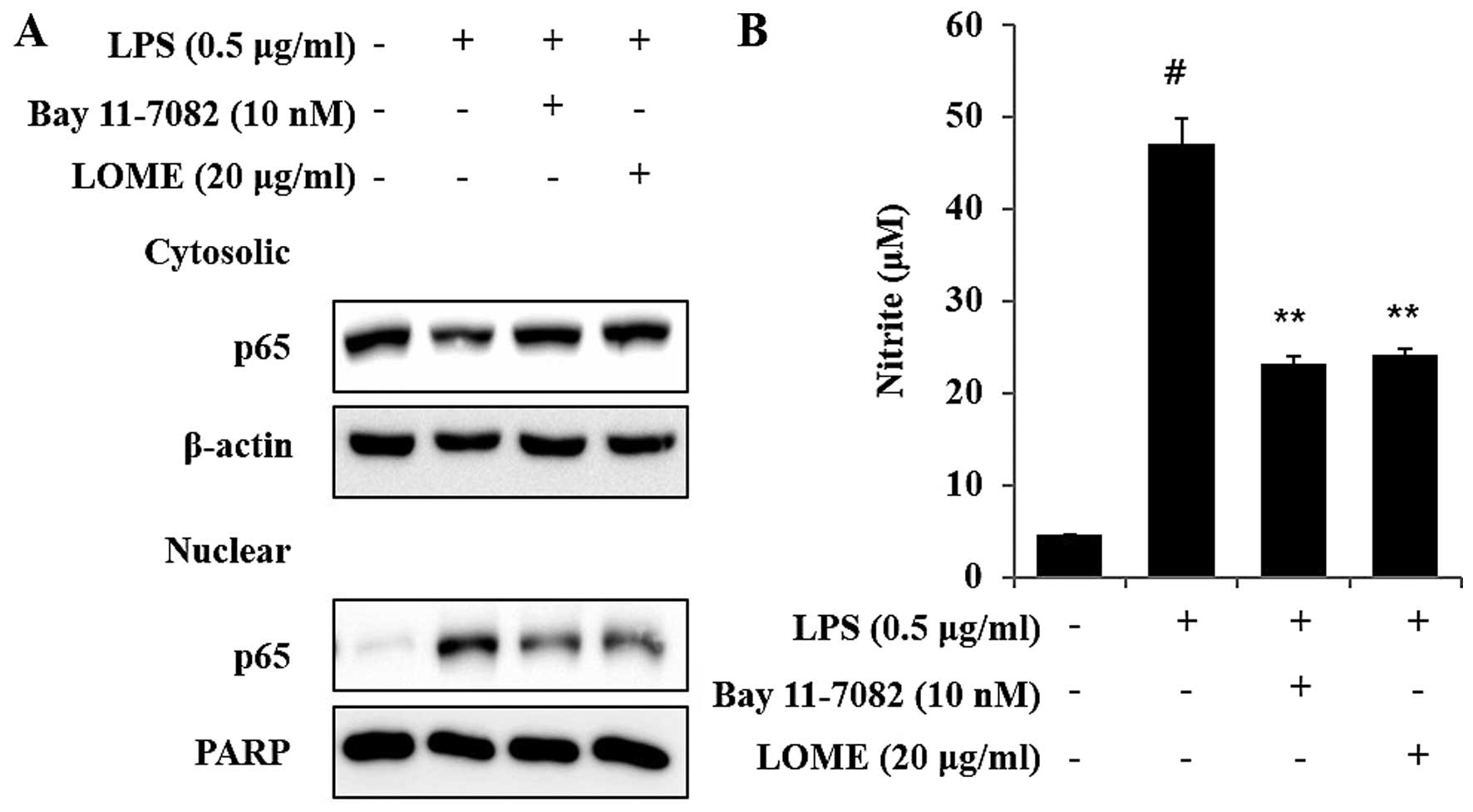
LOME and an inhibitor of IκB-α
phosphorylation (Bay 11-7082) inhibit NO expression and the
activation of NF-κB in LPS-stimulated RAW264.7 cells
In the present study, Bay 11-7082 was used to
inhibit the NF-κB pathway. Bay 11-7082, also known as
(E)-3-(4-methylphenylsulfonyl)-2-propenenenitrile, has a molecular
weight of 207.25 kDa and was provided at ≥98% purity (HPLC). It is
an irreversible inhibitor of IκB-α phosphorylation, which increases
the stabilization of IκB-α and specifically blocks NF-κB signaling.
Bay 11-7082 has been shown to be a potent inducer of apoptosis in a
number of cancer cell lines. Although Bay 11-7082 has not undergone
clinical development, prior in vivo animal experiments have
demonstrated its limited toxicity and therapeutic effectiveness
(19,20).
LOME significantly suppressed the LPS-induced
activation of NF-κB in the RAW264.7 cells, as shown by western blot
analysis. Bay 11-7082 (Calbiochem-Merck, Darmstadt, Germany), a
potential anti-inflammatory agent, is an irreversible inhibitor of
cytokine-inducible IκB-α phosphorylation (IC50=10
µM) (21). Bay 11-7082
blocked the LPS-induced increases in the activation of NF-κB. As
expected, LOME (20 µg/ml) inhibited the LPS-induced
activation of NF-κB (Fig. 7A).
These results suggest that LOME reduces inflammation by inhibiting
the LPS-induced activation of NF-κB in the RAW264.7 cells. In
addition, LOME and Bay 11-7082 also affected NO expression. The
inhibition of NO expression by each of the compounds was clearly
effective (Fig. 7B).
Discussion
Inflammation not only participates in host defense
but also controls the pathogenesis of numerous diseases, including
type II diabetes mellitus (22),
atherosclerosis (23), obesity
(24), silicosis, Alzheimer's
disease (25), gout (26) and kidney disease (27). An elevated level of circulating
inflammatory cytokines, in particular, and a combined elevation of
IL-1 and IL-6, constitute risk factors for the pathogenesis of type
II diabetes mellitus. Thus, the reduction of inflammation may be an
important therapeutic target for controlling various diseases. In
the present study, LOME significantly inhibited the production of
NO, PGE2, IL-6, IL-1β and TNF-α in the LPS-stimulated
RAW264.7 cells, and suppressed the mRNA or protein expression of
iNOS, COX-2, and MAPKs (p38 MAPK, JNK and ERK). These results were
accompanied by a reduction in the translocation of NF-κB to the
nucleus in the LPS-stimulated RAW264.7 cells.
Among the many pro-inflammatory mediators, NO and
PGE2 play key roles in inflammatory reactions (28,29). NO is generated in different types
of cell by at least three isoforms of NOS, among which iNOS is an
inducible and Ca2+-independent isoform of NOS. Massive
amounts of NO produced by iNOS under inflammatory conditions, are
potentially harmful as NO is capable of activating pro-inflammatory
signaling to produce oxidative stress. Thus, the inhibition of NO
production in response to inflammatory stimuli may be a useful
therapeutic strategy in treating inflammatory diseases (30). In addition, prostanoids are a
group of lipid mediators that regulate numerous processes in the
body. These processes include the regulation of blood pressure,
blood clotting, sleep, labor and inflammation. PGH2 is
the common substrate for a number of different synthases that
produce the major prostanoids, including PGD2,
PGE2, prostacyclin (PGI2), and thromboxane
(TXA2). Among these, PGE2 plays crucial roles
in various biological events including neuronal function, female
reproduction, vascular hypertension, tumorigenesis, kidney function
and inflammation (31). During
inflammatory responses, pain is produced through complex
interactions between various inflammatory mediators, one of which
is PGE2. Fever is a nearly universal sign of
inflammation and infectious processes. Taken together, these
findings indicate that NO and PGE2 are involved in the
regulation of various processes of acute and chronic inflammation.
Thus, in this study, we examined the effects of LOME on the
production of NO and PGE2. In this analysis, we found
that LOME inhibited the LPS-induced production of NO and
PGE2 in the RAW264.7 cells. In addition, the mRNA
expression of iNOS and COX-2 was inhibited by LOME. These results
indicate that LOME exerts anti-inflammatory effects by
down-regulating the expression of iNOS and COX-2, and suggests that
LOME may act as an anti-inflammatory agent.
The MAPKs are a group of signaling molecules that
also appear to play critical roles in inflammatory processes
(32). Evidence suggests that the
activation of MAPKs is important in the regulation of NO
production, through activation of NF-κB (33). In agreement with these previous
observations, the LPS stimulation of the RAW264.7 cells caused
phosphorylation of the ERK, JNK, and p38 kinases, whereas LOME
significantly inhibited ERK and p38 phosphorylation in these same
cells (Fig. 5). With a reduction
in the phosphorylation of MAPKs, LOME suppressed NF-κB
translocation into the nucleus. These findings indicate that the
anti-inflammatory effects of LOME may be closely associated with
blocking NF-κB translocation into the nucleus, caused by a
reduction in the phosphorylation of MAPKs.
NF-κB is an important factor that regulates the
expression of inflammation-associated mediators, including iNOS,
COX-2 and TNF-α; which contain NF-κB binding motifs within their
promoters. Many anti-inflammatory agents, such as a methanolic
extract of Werklea insignis, exhibit their potency by
suppressing NF-κB signaling (34). In particular, natural products
targeting the NF-κB pathway are considered to be effective in
treating inflammatory diseases due to the inhibition of one or more
activation steps in the signaling pathway. NF-κB is a
redox-sensitive transcription factor that regulates a multitude of
inflammatory genes, including cytokines, chemokines, adhesion
molecules and acute phase proteins. In the present study, after
treatment with LOME, the level of nuclear NF-κB p65 was reduced
while the cytoplasmic fraction accumulated. Although the reduction
in iNOS and COX-2 may be the result of NF-κB inactivation, other
mechanisms responsible for the effects of LOME effects may not be
excluded due to the wide contribution of NF-κB. Thus, LOME may
affect the upstream molecular components of IκB-NF-κB signaling. To
clarify the underlying molecular networks, more experiments are
warranted. LOME exerts anti-inflammatory effects on RAW264.7 cells
and macrophages; however, the detailed molecular mechanisms remain
unclear. LOME suppresses LPS-stimulated macrophage inflammatory
responses through the blockade of NF-κB; the detailed molecular
mechanism involves the inhibition of COX-2 and iNOS expression in
RAW264.7 macrophages, through the inactivation of NF-κB by blocking
the degradation of IκB. These studies, and our data, extend our
understanding of the molecular mechanisms underlying the biological
activity and pharmacological uses of LOME. In addition, the Bay
11-7082 inhibitor and LOME, attenuated NF-κB activity, thereby
inhibiting the expression of NO (Fig.
7B). The ability to inhibit COX-2 and iNOS expression may
account, in part, for the anti-inflammatory properties of LOME.
Furthermore, to elucidate clearly the mechanism responsible for the
anti-inflammatory effects of LOME, we examined whether NF-κB was
involved in exerting its anti-inflammatory effects using Bay
11-7082 (an NF-κB inhibitor). Based on these results, we concluded
that Bay 11-7082 inhibits NO secretion through its effects on the
NF-κB pathway, not by inhibiting the inflammatory response through
another mechanism. Taken together, these findings show that LOME
has the potential to be used as a potential therapeutic treatment
for inflammatory disorders.
In conclusion, this study demonstrated that LOME
inhibited the production of NO and PGE2 as well as
pro-inflammatory mediators, in RAW264.7 cells. In addition, LOME
reduced the mRNA expression of iNOS and COX-2 by inhibiting the
NF-κB and MAPK (ERK, JNK and p38) signaling pathways. These
findings suggest that LOME may be used as both a soothing agent and
for the treatment of inflammatory diseases.
Acknowledgments
The present study was supported by grants from the
Ministry of Science, ICT and Future Planning (FGC1011534), and from
the Korea Research Institute of Bioscience and Biotechnology
(KGM1221622) of the Republic of Korea.
References
|
1
|
Lee AK, Sung SH, Kim YC and Kim SG:
Inhibition of lipopolysaccharide-inducible nitric oxide synthase,
TNF-alpha and COX-2 expression by sauchinone effects on
I-kappaBalpha phosphorylation, C/EBP and AP-1 activation. Br J
Pharmacol. 139:11–20. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Curran AD: The role of nitric oxide in the
development of asthma. Int Arch Allergy Immunol. 111:1–4. 1996.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Ketteler M, Cetto C, Kirdorf M, Jeschke
GS, Schafer JH and Distler A: Nitric oxide in sepsis-syndrome:
Potential treatment of septic shock by nitric oxide synthase
antagonists. Kidney Int Suppl. 64:S27–S30. 1998.PubMed/NCBI
|
|
4
|
Heo SJ, Yoon WJ, Kim KN, Ahn GN, Kang SM,
Kang DH, Affan A, Oh C, Jung WK and Jeon YJ: Evaluation of
anti-inflammatory effect of fucoxanthin isolated from brown algae
in lipopolysaccharide-stimulated RAW 264.7 macrophages. Food Chem
Toxicol. 48:2045–2051. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Lu Y, Suh SJ, Kwak CH, Kwon KM, Seo CS, Li
Y, Jin Y, Li X, Hwang SL, Kwon O, et al: Saucerneol F, a new
lignan, inhibits iNOS expression via MAPKs, NF-kappaB and AP-1
inactivation in LPS-induced RAW264.7 cells. Int Immunopharmacol.
12:175–181. 2012. View Article : Google Scholar
|
|
6
|
Mordan LJ, Burnett TS, Zhang LX, Tom J and
Cooney RV: Inhibitors of endogenous nitrogen oxide formation block
the promotion of neoplastic transformation in C3H 10T1/2
fibroblasts. Carcinogenesis. 14:1555–1559. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ochoa JB, Udekwu AO, Billiar TR, Curran
RD, Cerra FB, Simmons RL and Peitzman AB: Nitrogen oxide levels in
patients after trauma and during sepsis. Ann Surg. 214:621–626.
1991. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hunt J and Gaston B: Airway nitrogen oxide
measurements in asthma and other pediatric respiratory diseases. J
Pediatr. 137:14–20. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Giuliano F and Warner TD: Origins of
prostaglandin E2: involvements of cyclooxygenase (COX)-1
and COX-2 in human and rat systems. J Pharmacol Exp Ther.
303:1001–1006. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Graham A, Nowicke JW, Skvarla JJ, Graham
SA, Patel V and Lee S: Palynology and systematics of the
Lythraceae. II. Genera Haitia through Peplis. Am J Bot. 74:829–850.
1987. View
Article : Google Scholar
|
|
11
|
de Wilde WJJO and Duyfjes BEE:
Lagerstroemia (Lythraceae) in Malesia. Blumea - Biodiversity,
Evolution and Biogeography of Plants. 59:113–122. 2014. View Article : Google Scholar
|
|
12
|
Priyadi H, Takao G, Rahmawati I,
Supriyanto B, Ikbal Nursal W and Rahman I: Five hundred plant
species in Gunung Halimun Salak National Park, West Java: a
checklist including Sundanese names, distribution and use. Cifor.
p182010.
|
|
13
|
Burkill IH: A Dictionary of the Economic
Products of the Malay Peninsula. II. Crown Agents; London: pp.
p13001935
|
|
14
|
Su YW, Chiou WF, Chao SH, Lee MH, Chen CC
and Tsai YC: Ligustilide prevents LPS-induced iNOS expression in
RAW 264.7 macrophages by preventing ROS production and
down-regulating the MAPK, NF-kappaB and AP-1 signaling pathways.
Int Immunopharmacol. 11:1166–1172. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Park JW, Kwon OK, Kim JH, Oh SR, Kim JH,
Paik JH, Marwoto B, Widjhati R, Juniarti F, Irawan D and Ahn KS:
Rhododendron album Blume inhibits iNOS and COX-2 expression in
LPS-stimulated RAW264.7 cells through the downregulation of NF-κB
signaling. Int J Mol Med. 35:987–994. 2015.PubMed/NCBI
|
|
16
|
Cirino G, Distrutti E and Wallace JL:
Nitric oxide and inflammation. Inflamm Allergy Drug Targets.
5:115–119. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Guha M and Mackman N: LPS induction of
gene expression in human monocytes. Cell Signal. 13:85–94. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Lawrence T, Willoughby DA and Gilroy DW:
Anti-inflammatory lipid mediators and insights into the resolution
of inflammation. Nat Rev Immunol. 2:787–795. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Keller SA, Hernandez-Hopkins D, Vider J,
Ponomarev V, Hyjek E, Schattner EJ and Cesarman E: NF-kappaB is
essential for the progression of KSHV- and EBV-infected lymphomas
in vivo. Blood. 107:3295–3302. 2006. View Article : Google Scholar
|
|
20
|
Zheng X, Chang RL, Cui XX, Avila G, Huang
MT, Liu Y, Kong AN, Rabson AB and Conney AH: Inhibition of NF-κB by
(E)3-[(4-methylphenyl)-sulfonyl]-2-propenenitrile (BAY11-7082; BAY)
is associated with enhanced
12-O-tetradecanoylphorbol-13-acetate-induced growth suppression and
apoptosis in human prostate cancer PC-3 cells. Int J Oncol.
32:257–264. 2008.
|
|
21
|
Mori N, Yamada Y, Ikeda S, Yamasaki Y,
Tsukasaki K, Tanaka Y, Tomonaga M, Yamamoto N and Fujii M: Bay
11-7082 inhibits transcription factor NF-kappaB and induces
apoptosis of HTLV-I-infected T-cell lines and primary adult T-cell
leukemia cells. Blood. 100:1828–1834. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Schroder K, Zhou R and Tschopp J: The
NLRP3 inflammasome: a sensor for metabolic danger? Science.
327:296–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Duewell P, Kono H, Rayner KJ, Sirois CM,
Vladimer G, Bauernfeind FG, Abela GS, Franchi L, Nunez G, Schnurr
M, et al: NLRP3 inflammasomes are required for atherogenesis and
activated by cholesterol crystals. Nature. 464:1357–1361. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Vandanmagsar B, Youm YH, Ravussin A,
Galgani JE, Stadler K, Mynatt RL, Ravussin E, Stephens JM and Dixit
VD: The NLRP3 inflammasome instigates obesity-induced inflammation
and insulin resistance. Nat Med. 17:179–188. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Halle A, Hornung V, Petzold GC, Stewart
CR, Monks BG, Reinheckel T, Fitzgerald KA, Latz E, Moore KJ and
Golenbock DT: The NALP3 inflammasome is involved in the innate
immune response to amyloid-beta. Nat Immunol. 9:857–865. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Martinon F, Petrilli V, Mayor A, Tardivel
A and Tschopp J: Gout-associated uric acid crystals activate the
NALP3 inflammasome. Nature. 440:237–241. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Tsai PY, Ka SM, Chang JM, Chen HC, Shui
HA, Li CY, Hua KF, Chang WL, Huang JJ, Yang SS and Chen A:
Epigallocatechin-3-gallate prevents lupus nephritis development in
mice via enhancing the Nrf2 antioxidant pathway and inhibiting
NLRP3 inflammasome activation. Free Radic Biol Med. 51:744–754.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Goldring MB and Berenbaum F: The
regulation of chondrocyte function by proinflammatory mediators:
prostaglandins and nitric oxide. Clin Orthop Relat Res.
427:S37–S46. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kim HS, Park JW, Kwon OK, Kim JH, Oh SR,
Lee HK, Bach TT, Quang BH and Ahn KS: Anti-inflammatory activity of
a methanol extract from Ardisia tinctoria on mouse macrophages and
paw edema. Mol Med Rep. 9:1388–1394. 2014.PubMed/NCBI
|
|
30
|
Sautebin L: Prostaglandins and nitric
oxide as molecular targets for anti-inflammatory therapy.
Fitoterapia. 71(Suppl 1): S48–S57. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kobayashi T and Narumiya S: Prostanoids in
health and disease; lessons from receptor-knockout mice. Adv Exp
Med Biol. 507:593–597. 2002. View Article : Google Scholar
|
|
32
|
Kim JY, Kim HJ, Kim SM, Park KR, Jang HJ,
Lee EH, Jung SH and Ahn KS: Methylene chloride fraction of the
leaves of Thuja orientalis inhibits in vitro inflammatory
biomarkers by blocking NF-kappaB and p38 MAPK signaling and
protects mice from lethal endotoxemia. J Ethnopharmacol.
133:687–695. 2011. View Article : Google Scholar
|
|
33
|
Ji LL, Gomez-Cabrera MC and Vina J: Role
of nuclear factor kappaB and mitogen-activated protein kinase
signaling in exercise-induced antioxidant enzyme adaptation. Appl
Physiol Nutr Metab. 32:930–935. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Park JW, Kwon OK, Jang HY, Jeong H, Oh SR,
Lee HK, Han SB and Ahn KS: A leaf methanolic extract of Wercklea
insignis attenuates the lipopolysaccharide-induced inflammatory
response by blocking the NF-kappaB signaling pathway in RAW 264.7
macrophages. Inflammation. 35:321–331. 2012. View Article : Google Scholar
|