Introduction
Bone is actively maintained by the coordinated
balance between osteoblasts and osteoclasts (1–3).
Osteoblasts mediate bone formation, while osteoclasts are
responsible for bone resorption (1,2).
The activities of osteoblasts can be regulated at the level of
differentiation by various regulatory signals. The differentiation
and activity of osteoblasts are regulated by various anabolic
factors, such as insulin, members of the transforming growth factor
(TGF)-β family [including bone morphogenetic proteins (BMPs)] and
Wnt proteins, and by intracellular kinases, such as Akt (4). In particular, BMPs initiate
osteoblast differentiation through the induction of the expression
and post-translational modification of various osteogenic
transcription factors, including Runx2 (Cbfa1), Osterix and several
homeodomain-containing Dlx proteins (5–10).
Subsequently, these osteogenic transcription factors regulate the
differentiation of osteoblasts (10–16).
The Runt domain transcription factors are composed
of a larger DNA-binding α subunit and a smaller non-DNA-binding β
subunit. There are three mammalian genes (Runx1, Runx2, and Runx3)
encoding the α subunit. Runx2 and Runx3 are essential for
chondrocyte maturation, a prerequisite for endochondral
ossification. In addition, Runx2 is essential for the commitment of
multipotent mesenchymal cells into the osteoblastic lineage, and it
inhibits adipocyte differentiation (17). Runx2 DNA binding sequences have
been identified in the enhancer/promoter regions of many osteoblast
specific genes, and Runx2 can bind to the osteoblast-specific
cis-acting element (OSE) present in the promoter
regions of collagen type I, α1 (COL1A1), osteocalcin
(OC), alkaline phosphatase (ALP), bone sialoprotein
(BSP) and osteopontin (OPN) (18,19). The function of Runx2 is regulated
by osteogenic signaling factors, such as BMP2, a member of the
TGF-β superfamily (20). BMP2
induces Runx2 expression through Smad1/5/8, and BMP2-activated
Smads physically interact with Runx2 to induce osteoblast
differentiation (7,21–23).
Osterix (also known as Sp7) is a zinc
finger-containing osteoblast-specific transcription factor and it
is essential for the differentiation and proliferation of
osteoblasts (24–27). The DNA-binding domain of Osterix
is located at the C-terminus and it contains three C2H2-type zinc
finger domains that share a high degree of identity with similar
motives in Sp1, Sp3 and Sp4. N-terminal proline-rich region (PRR)
mediates the protein-protein interaction. Osterix acts downstream
of Runx2 and regulates the expression of many osteoblast
differentiation markers including ALP, OC, osteonectin, OPN and
Runx2 (15,24,28).
Bone mass in adult humans decreases with age,
leading to an increased risk of fractures. Bone mineral density
(BMD) and bone metabolism are affected by genetic, endocrine,
mechanical and nutritional factors, with interactions among the
different factors (29).
Nutritional factors are particularly important for bone health as
they are modifiable (30).
Natural products of plant origin are still a major part of
traditional medicinal systems in Korea.
The root of Lithospermum erythrorhizon Sieb.
et Zucc. (LR), an herbal medicine, is known to possess various
antiviral and biological activities, including the inhibition of
human immunodeficiency virus type 1 (HIV-1), and it is extensively
used in traditional medicine due to the known functions of herbs
described in the literature of traditional Korean and Chinese
medicines (31,32). Ethanol (EtOH) extract from LR
(LES) has long been used in traditional Asian medicine for the
treatment of skin cancer. It has been reported that the extracts
from LR attenuates immunosuppression induced by cyclophosphamide,
an antitumor agent (33,34). The majority of studies on the
bioactivities of LR have been carried out mainly with
naphthoquinone pigments, including shikonin and its derivatives,
which are extracted using non-polar solvents, such as hexane or by
supercritical extraction (35,36). Shikonin, a major active component
of LR, possesses numerous pharmacological properties, including
anti-inflammatory properties. Shikonin plays a dual role in the
regulation of the early and late stages of collagen type II
arthritis (37). Shikonin exerts
protective effects on cartilage in rheumatoid arthritis (38), but also induces osteoclast
differentiation in vitro (39). However, the function of LR in
osteoblast differentiation remains unknown. Moreover, to the best
of our knowledge, there are only a few studies available to date on
the bioactivities of water-soluble and EtOH extracts of LR, even
though they are more suitable materials for the development of
health functional food from LR.
In this study, we examined the effects of LES on
osteoblastogenesis and we aimed to determine whether the osteoblast
transcription factors, Runx2 and Osterix, play a role in these
effects. We demonstrated that treatment with LES promoted
osteoblast differentiation induced by BMP4 and enhanced the
osteogenic functions of Runx2 and Osterix by increasing their
protein levels and transcriptional activity.
Materials and methods
Plant materials and extraction
Dried 1-year-old Lithospermi radix (LR) was
purchased from Jacheon, Chungbuk, South Korea. A voucher specimen
(MPS000071) has been deposited at the Herbarium of the Department
of Herbal Crop Research, National Institute of Horticultural and
Herbal Science, RDA, Eumseong, Korea. The powder of LR (1.5 kg) was
extracted with 70% EtOH (15 l) at 80°C for 2.5 h using a natural
substance extractor (EG-BE1; TIPBio, Siheung, Korea) to obtain 70%
EtOH extract. The EtOH extract was concentrated under a vacuum
using a rotary evaporator (N-1200B; Eyela, Tokyo, Japan) and dried
in a freeze dryer (LP20; ilShinBioBase Co., Ltd., Dongducheon,
Korea) to yield the final test samples (LES, 424 g).
Plasmids, antibodies and reagents
Plasmids for Myc-tagged Osterix and Myc-tagged Runx2
were constru cted in a CMV promoter-derived mammalian expression
vector (pCS4-3Myc; obtained from Dr C.Y. Yeo, Ehwa University,
Korea). Anti-Myc (no. 9E10, diluted 1:1,000) was purchased from
Roche Applied Science (Seokyung Bldg. Seoul, Korea). Anti-α-tubulin
antibody (no. B-5-1-2, diluted 1:5,000) was purchased from
Sigma-Aldrich (St. Louis, MO, USA). Anti-Osterix (no. A-13, diluted
1:1,000) and Dlx5 antibodies (no. C-20, diluted 1:1,000) were
purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA).
Anti-Runx2 antibody (no. ab76956, diluted 1:1,000) was purchased
from Abcam (Boston, MA, USA). Recombinant human BMP4 was purchased
from R&D Systems (Minneapolis, MN, USA).
Cell culture and transient
transfection
The cells (293 cells and C2C12 mouse myoblasts) were
cultured at 37°C, 5% CO2 in Dulbecco's modified Eagle's
medium (DMEM) supplemented with 5% (for 293 cells) or 10% (for
C2C12 cells) fetal bovine serum (FBS), 100 U/ml penicillin, and 100
g/ml streptomycin. All cells were purchased from the American Type
Culture Collection (ATCC, Manassas, VA, USA). The C2C12 cells were
treated with BMP4 (30 ng/ml) and the EtOH extract from
Lithospermum erythrorhizon Sieb. et Zicao (LES) (each 30 or
60 mg/ml) for 3 days. DMEM, FBS, and the antibiotics were purchased
from Life Technologies (Grand Island, NY, USA). Transient
transfection was performed using a polyethylenimine (PEI)
(Polysciences, Inc., Warrington, PA, USA)-mediated method. Total
amounts of transfected plasmids in each group were equalized by
adding the empty vector (pCS4+; obtained from Dr Yeo CY).
Cell lysate preparation and immunoblot
analysis
The cells were lysed in ice-cold lysis buffer [25 mM
HEPES (pH 7.5), 150 mM NaCl, 1% NP-40, 0.25% sodium deoxycholate,
10% glycerol, 1 mM EDTA] containing phosphatase inhibitors (25 mM
NaF, 1 mM Na3VO4) and protease inhibitors
(250 µM phenylmethylsulfonyl fluoride, 10 µg/ml
leupeptin, 10 µg/ml aprotinin, and 10 µg/ml pepstatin
A). Lysates were cleared by centrifugation at 16,000 × g for 15 min
at 4°C, and the supernatants were used as cell lysates. Cell
lysates containing 30 µg of total proteins were subjected to
sodium dodecyl sulfate-polyacrylamide gel electro phoresis
(SDS-PAGE) and the protein content was estimated using a bovine
serum albumin protein assay. Proteins were mixed with sample buffer
containing β-mercaptoethanol and heated at 100°C for 3 min. A total
of 30 µg of each cell lysate was fractionated by SDS-PAGE on
a 10% polyacrylamide gel and transferred onto polyvinylidene
difluoride (PVDF) membranes. After blocking with 5% skim milk in
Tris-buffered saline (TBS) containing 0.02% Tween-20 at room
temperature for 40 min, the proteins were visualized with
appropriate primary antibodies (diluted 1:1,000) at room
temperature for 1 h. Tubulin (diluted 1:5,000) was used as a
loading control. This was followed by the addition of anti-rabbit
or anti-mouse horseradish peroxidase (HRP)-conjugated secondary
antibodies diluted 1:25,000 in TBS/Tween-20. The blots were
visualized by enhanced ECL-chemiluminescence reagent (GE Healthcare
Life Sciences, Logan, UT, USA). Signals were detected and analyzed
using the LAS-4000 luminescent image analyzer (Fuji Photo Film Co.,
Ltd., Tokyo, Japan).
Luciferase reporter assay
The C2C12 cells were seeded on 24-well plates 1 day
prior to transfection. ALP-Luc (obtained from H.M. Ryoo, Seoul
University, Korea) and BSP-Luc (obtained from J.T. Kho, Chonnam
National University, Korea) luciferase reporters contained the
regulatory sequence of the osteoblast differentiation markers, ALP
or BSP. The cells were transfected with a CMV promoter-driven
β-galactosidase reporter (pCMV-β-gal), luciferase reporter and the
indicated combinations of the expression plasmids. Thirty-six hours
later, luciferase activities were measured using the Luciferase
Reporter Assay kit (E1501; Promega, Madison, WI, USA) and a
luminometer, and normalized with the corresponding β-galactosidase
activities for transfection efficiency. Experiments were performed
in triplicate and repeated at least 3 times. The averages and
standard deviations (SD) of representative experiments are
shown.
RNA preparation and semi-quantitative
RT-PCR
Total cellular RNA was prepared using TRIzol reagent
(Life Technologies) according to the manufacturer's instructions.
Random hexamer-primed cDNA was synthesized from 1 µg of
total RNA using the SuperScript III First-Strand Synthesis System
(Life Technologies). The following conditions were used for the
amplification by PCR: initial denaturation at 94°C for 1 min;
followed by 23–30 cycles of denaturation at 94°C for 30 sec,
annealing at a temperature optimized for each primer pair for 30
sec, and extension at 72°C for 30 sec; final extension at 72°C for
5 min. The following PCR primers were used: ALP forward,
5′-GGGTGGACTACCTCTTAGGTC-3′ and reverse,
5′-ATGATGTCCGTGGTCAATCCTG-3′ (30 cycles); BSP forward,
5′-CAGAAGTGGATGAAAACGAG-3′ and reverse, 5′-CGGTGGCGAGGTGGTCCCAT-3′
(25 cycles); COL1A1 forward, 5′-TCTCCACTCTTCTAGGTTCCT-3′ and
reverse, 5′-TTGGGTCATTTCCACATGC-3′ (23 cycles); Runx2
forward, 5′-AGCAACAGCAACAGCAG-3′ and reverse,
5′-GTAATCTGACTCTGTCCTTG-3′ (35 cycles); Osterix forward,
5′-GGGTTAAGGGGAGCAAAGTCAGAT-3′ and reverse,
5′-CTGGGGAAAGGAGGCACAAAGAAG-3′ (35 cycles); GAPDH forward,
5′-ACCACAGTCCATGCCATCAC-3′ and reverse, 5′-TCCACCACCCTGTTGCTGTA-3′
(25 cycles).
ALP staining and activity assay
The C2C12 cells were treated with BMP4 (30 ng/ml)
for 3 days, fixed in 4% paraformaldehyde for 15 min at room
temperature, rinsed with phosphate-buffered saline (PBS), and
stained with 5-bromo-4-chloro-3-indolyl phosphate/nitro blue
tetrazolium (BCIP/NBT) solution (Sigma-Aldrich) for 15 min at room
temperature. For ALP assay, the cells were washed with PBS and
lysed in 0.5% Triton X-100. ALP enzymatic activity was measured
using the SensoLyte pNPP ALP assay kit (AnaSpec, Inc., Fremont, CA,
USA) according to the manufacturer's instructions.
Alizarin Red S staining
The C2C12 cells in 24-well plates were transfected
using the PEI-mediated method. The C2C12 cells were stimulated with
BMP4. The cells were pre-treated with BMP4 for 10 days. These cells
were cultured at 5% CO2, 37°C. The transfected C2C12
cells were then fixed in 4% paraformaldehyde for 15 min at room
temperature and washed with PBS. They were then exposed to Alizarin
Red S solution (no. A5533; Sigma-Aldrich) adjusted to pH 4.1–4.3
using 0.5% ammonium hydroxide for 30 min at room temperature. The
mineralization-positive cells were stained red.
Statistical analysis
All experiments were performed with triplicate
independent samples and were repeated at least 3 times yielding
qualitatively identical results. The results are expressed as the
means ± standard error of the mean. Data were analyzed using the
Student's t-test (SPSS version 17.0 software; SPSS, Inc., Chicago,
IL, USA). A value of p<0.05 was considered to indicate a
statistically significant difference.
Results
LES enhances osteoblast differentiation
induced by BMP4
Recently, it was reported that LES regulates
osteoclastogenesis in vitro (39). However, the signaling mechanisms
of LES are less well known in osteoblastogenesis. The precise
molecular signaling mechanisms of action of LES as regards
osteoblast differentiation are not yet fully understood. Therefore,
in this study, we examined the effects of LES on the osteoblast
differentiation of C2C12 cells. We examined whether LES affects
BMP4-induced osteoblast differentiation. Stimulation with BMP4
induced the osteoblast differentiation of the C2C12 myoblasts. The
C2C12 cells were cultured and treated with low and high
concentrations (30 or 60 µg/ml) of LES for 3 days. LES
increased the expression of ALP, an osteoblast marker, in the cells
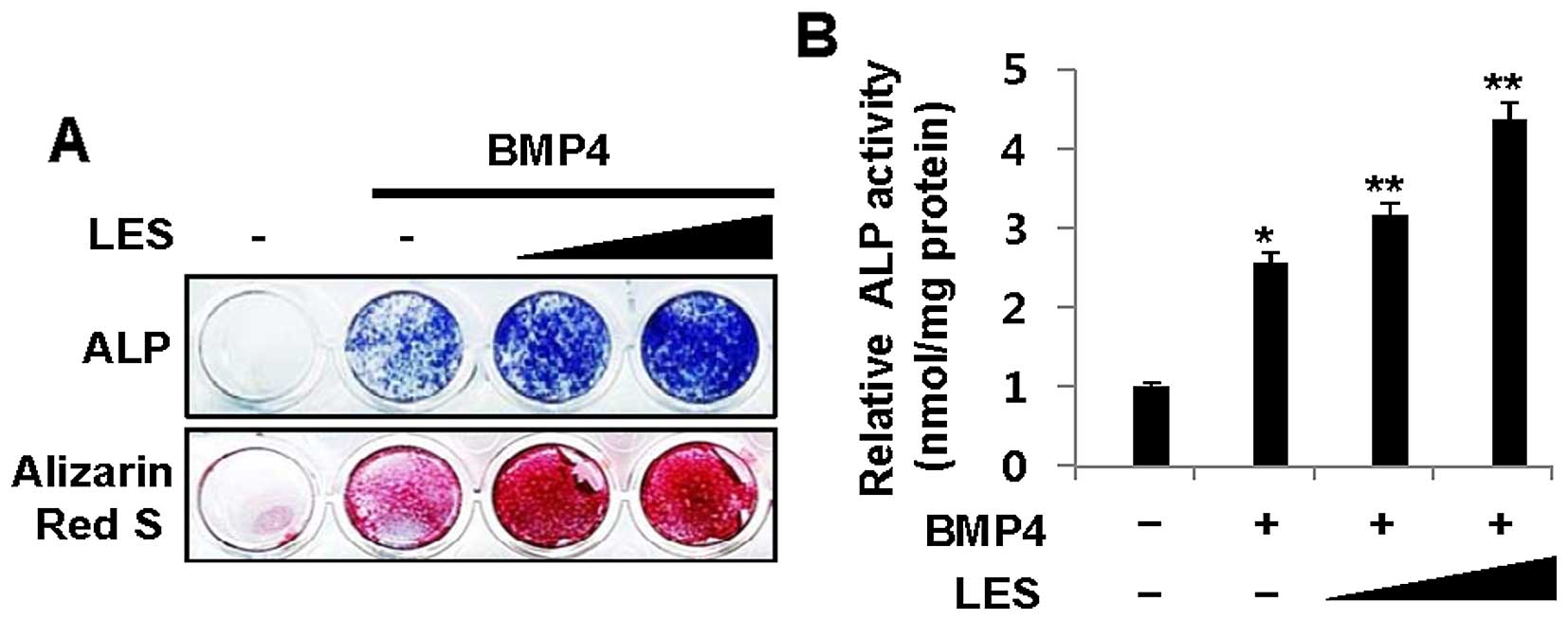
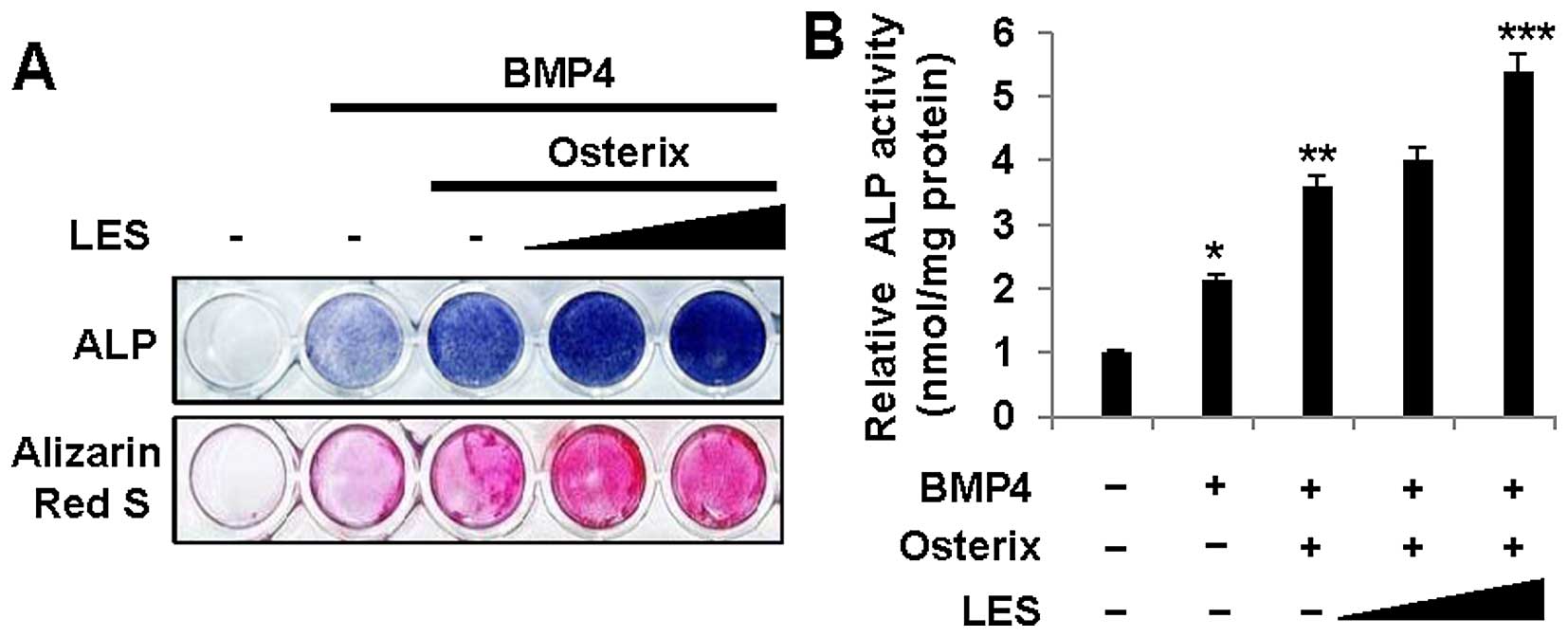
stimulated with BMP4, as shown by ALP staining (Fig. 1A, top panel), and also increased
ALP activity (Fig. 1B).
Similarly, LES increased mineralization, as observed by Alizarin
Red S staining (Fig. 1A, bottom
panel). These results suggest that LES enhances osteoblast
differentiation induced by BMP4.
LES affects the expression levels and
transcriptional activity of osteoblast markers in BMP4-induced
osteoblast differentiation
Previous studies have demonstrated that BMP4 gene
transfer into rodent muscle promotes bone formation (40–43). In this study, we confirmed that
LES enhances BMP4-induced osteoblast differentiation (Fig. 1). Treatment with LES increased the
expression levels of the osteoblast transcription factors, Runx2,
Osterix and Dlx5 (Fig. 2A), and
those of the specific markers, ALP, BSP and
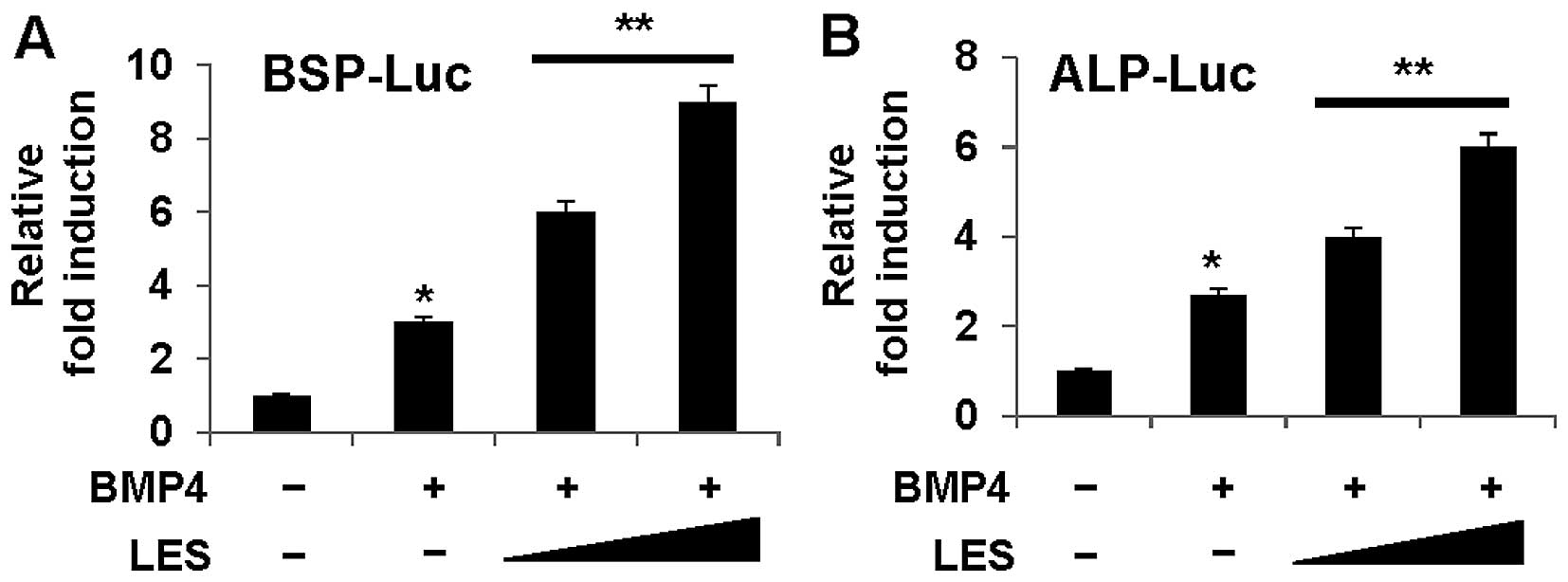
COL1A1, in the cells also treated with BMP4 (Fig. 2B). Moreover, we examined whether
LES can modulate the transcriptional activity using the
osteoblast-specific luciferase reporters, BSP-Luc (Fig. 3A) and ALP-Luc (Fig. 3B). LES significantly enhanced the
expression of the reporters. These results indicate that LES
regulates the transcriptional activity of osteoblast transcription
factors and is critical for the osteoblast transcription
factor-induced expression, of at least a subset of osteoblast
markers.
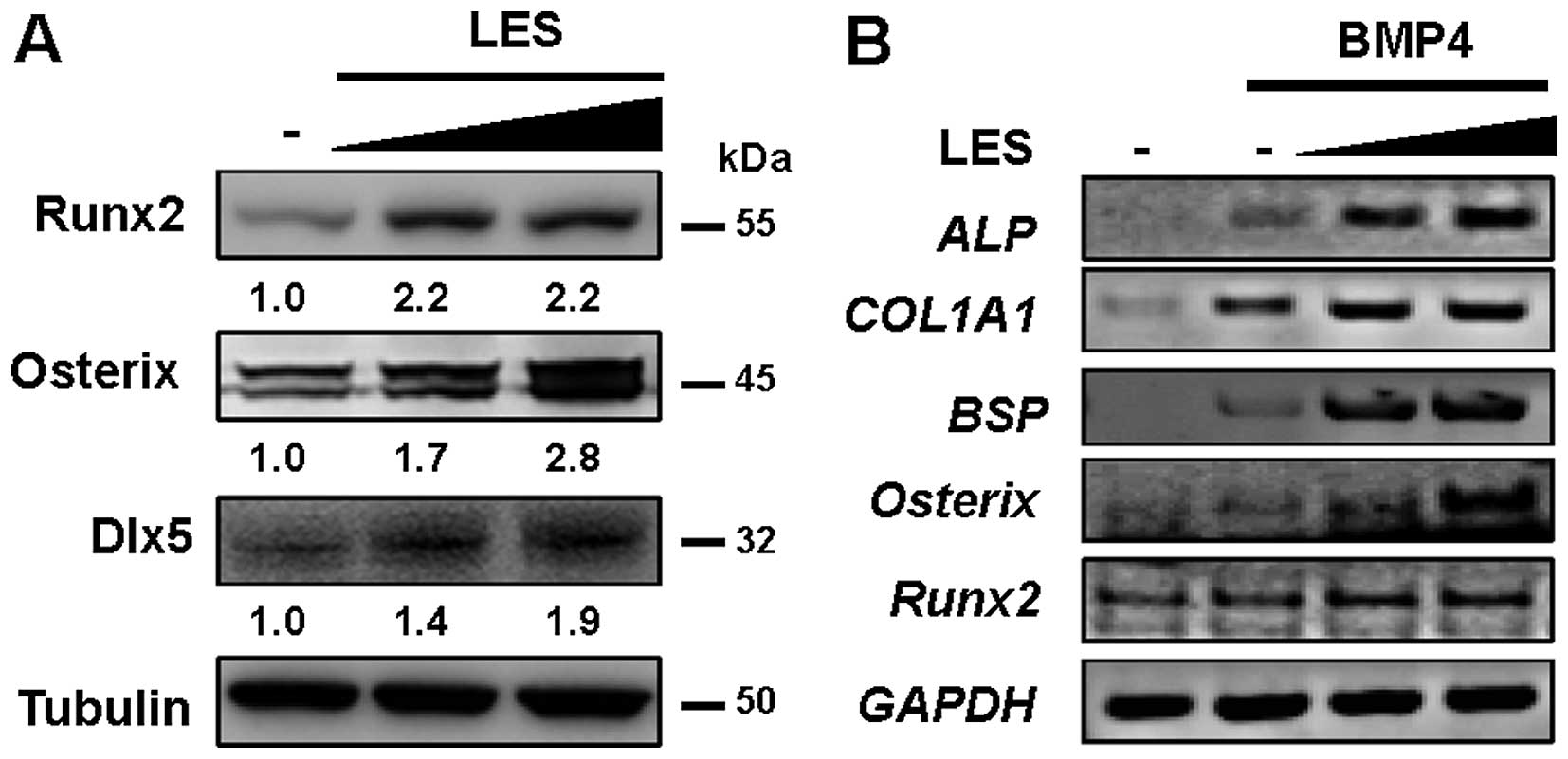 | Figure 2LES increases the expression of
osteoblast target genes during osteoblast differentiation. C2C12
cells were treated with bone morphogenetic protein 4 (BMP4) (30
ng/ml) and increasing amounts of LES (30 or 60 μg/ml) for 3
days. (A) The endogenous expression levels of osteoblast target
genes (Runx2, Osterix and Dlx5) in cell lysates compared by
immunoblotting using antibody against Runx2, Osterix or Dlx5,
respectively. Tubulin was used as a loading control. (B) The
expression levels of the osteoblast-specific markers, alkaline
phosphatase (ALP), bone sialoprotein (BSP), collagen
type I, α1 (COL1A1), Osterix, Runx2 compared
by RT-PCR. GAPDH was used as a loading control. LES, ethanol (EtOH)
extract from Lithospermum erythrorhizon Sieb. et Zucc. |
LES regulates the functions of the
osteoblast-specific transcription factor, Runx2, in BMP4-induced
osteoblast differentiation
The runt-related transcription factor, Runx2,
regulates the expression of bone and cartilage-related genes and is
required for bone formation. The regulatory mechanisms control both
the activation and repression of Runx2 gene transcription during
osteoblast differentiation and skeletal development. Runx2 is
essential for the proper execution of the osteogenic program
(7,19,44). In this study, in order to
elucidate the mechanisms of action of the major transcription
factor, Runx2, in cells treated with LES, we examined whether LES
affects Runx2 in cells stimulated with BMP4 to induce osteoblast
differentiation. The cells stimulated with BMP and treated with LES
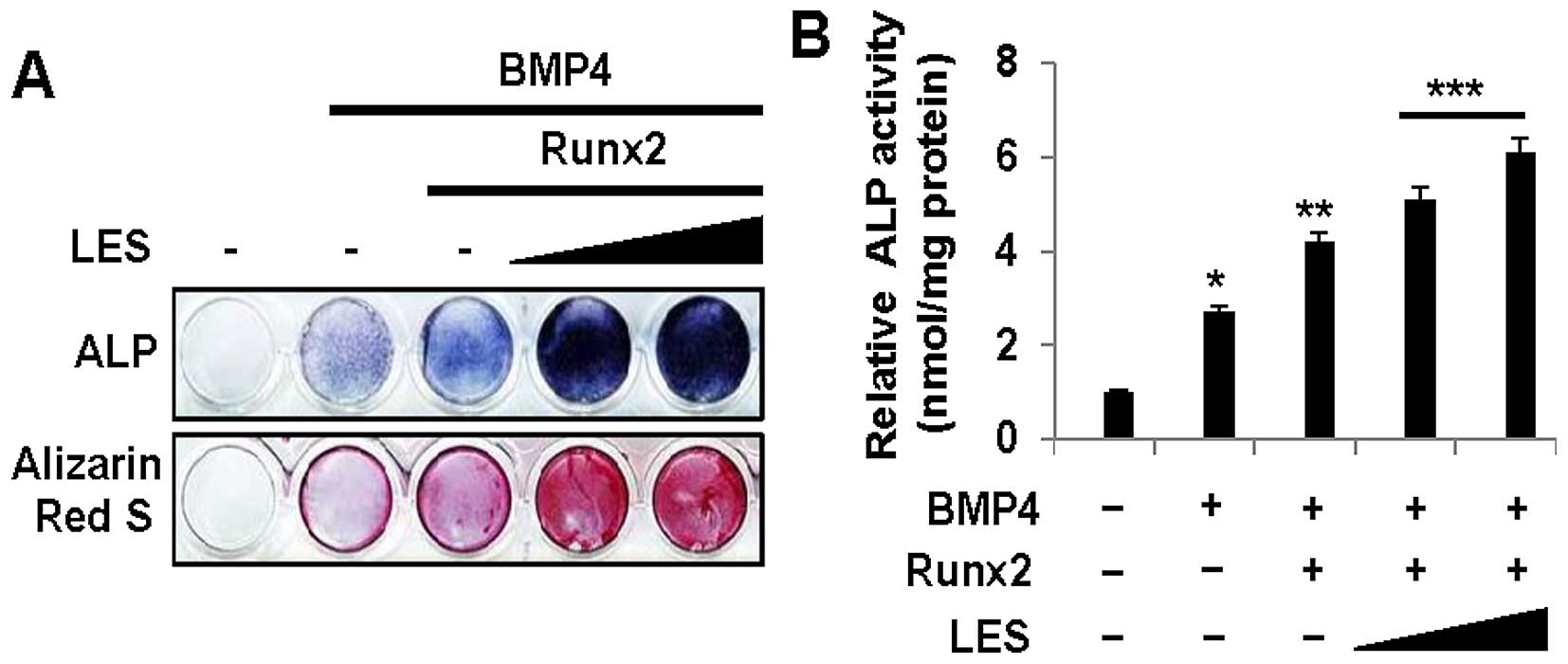
were transfected with a Runx2 expression plasmid. First, we found
that LES increased ALP activity, as shown by ALP staining (Fig. 4A, top panel), and also increased
ALP activity (Fig. 4B) in the
cells overexpressing Runx2 in the presence of BMP4. Similarly, LES
increased mineralization, as shown by Alizarin Red S staining
(Fig. 4A, bottom panel). We then
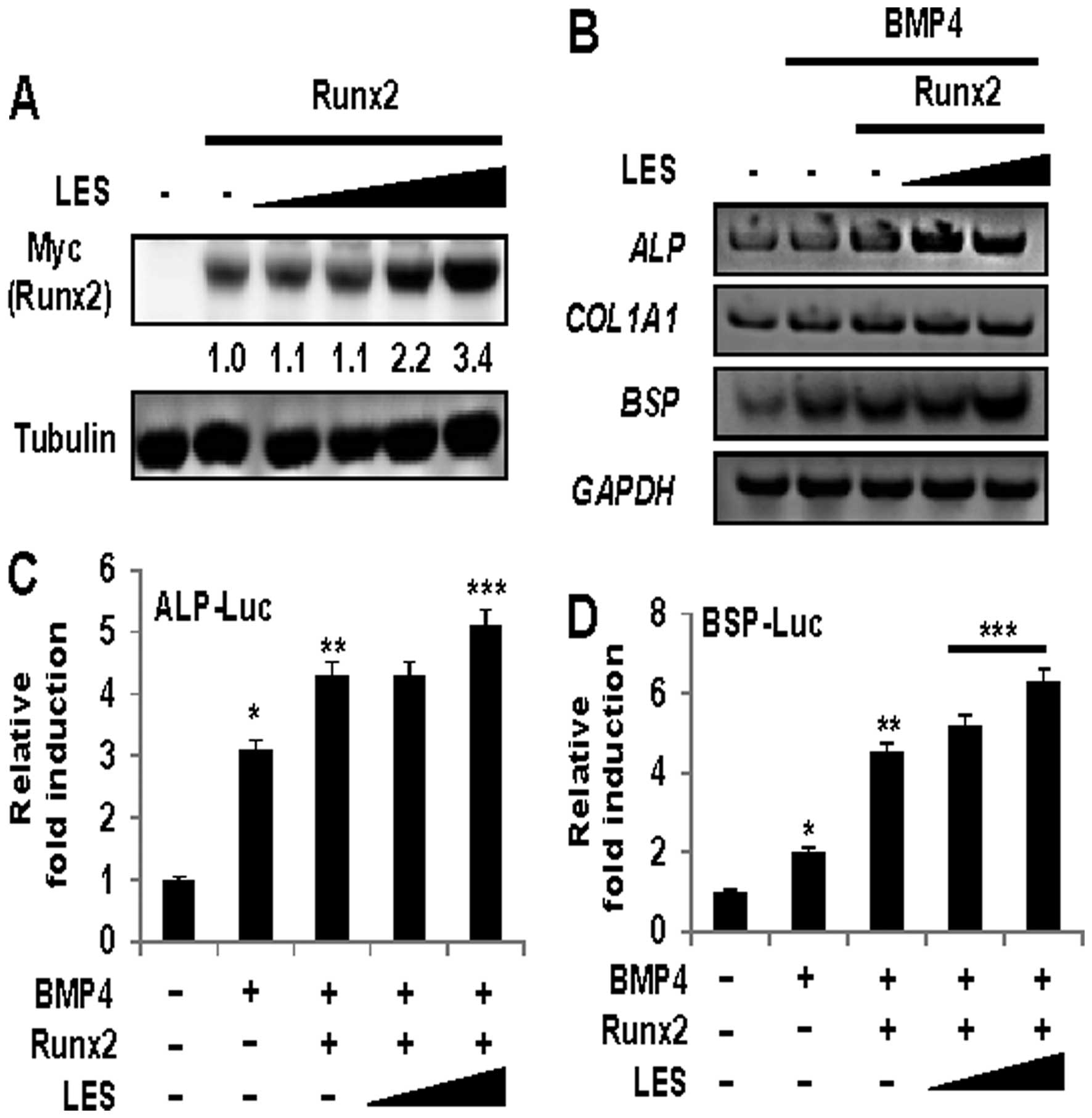
investigated whether LES affects the protein level of Runx2. The
protein levels of Runx2 were increased in the cells treated with
LES (Fig. 5A). In addition, LES
increased the expression of osteoblast specific markers in the
cells stimulated with BMP4 (Fig.
2B). We then examined the effects of LES on the BMP4- and
Runx2-induced expression of osteoblast marker genes (ALP,
BSP and COL1A1). As shown in Fig. 5B, not only ALP and
COL1A1, the early-stage osteogenic differentiation markers,
but also BSP, a late-stage osteogenic differentiation
marker, was positively affected by LES. The expression of
osteoblast markers in the cells stimulated with BMP4 to induce
osteoblast differentiation was increased in the presence of Runx2
and LES. Finally, to examine whether LES can modulate the
transcriptional activity of Runx2, we examined the effects of LES
on the transcriptional activity of Runx2 using the
osteoblast-specific luciferase reporters, ALP-Luc (Fig. 5C) and BSP-Luc (Fig. 5D). LES significantly enhanced the
transcriptional activity of the Runx2-induced expression of the
reporters. Taken together, these results suggest that LES regulates
the expression levels and transcriptional activity of Runx2 during
osteoblast differentiation.
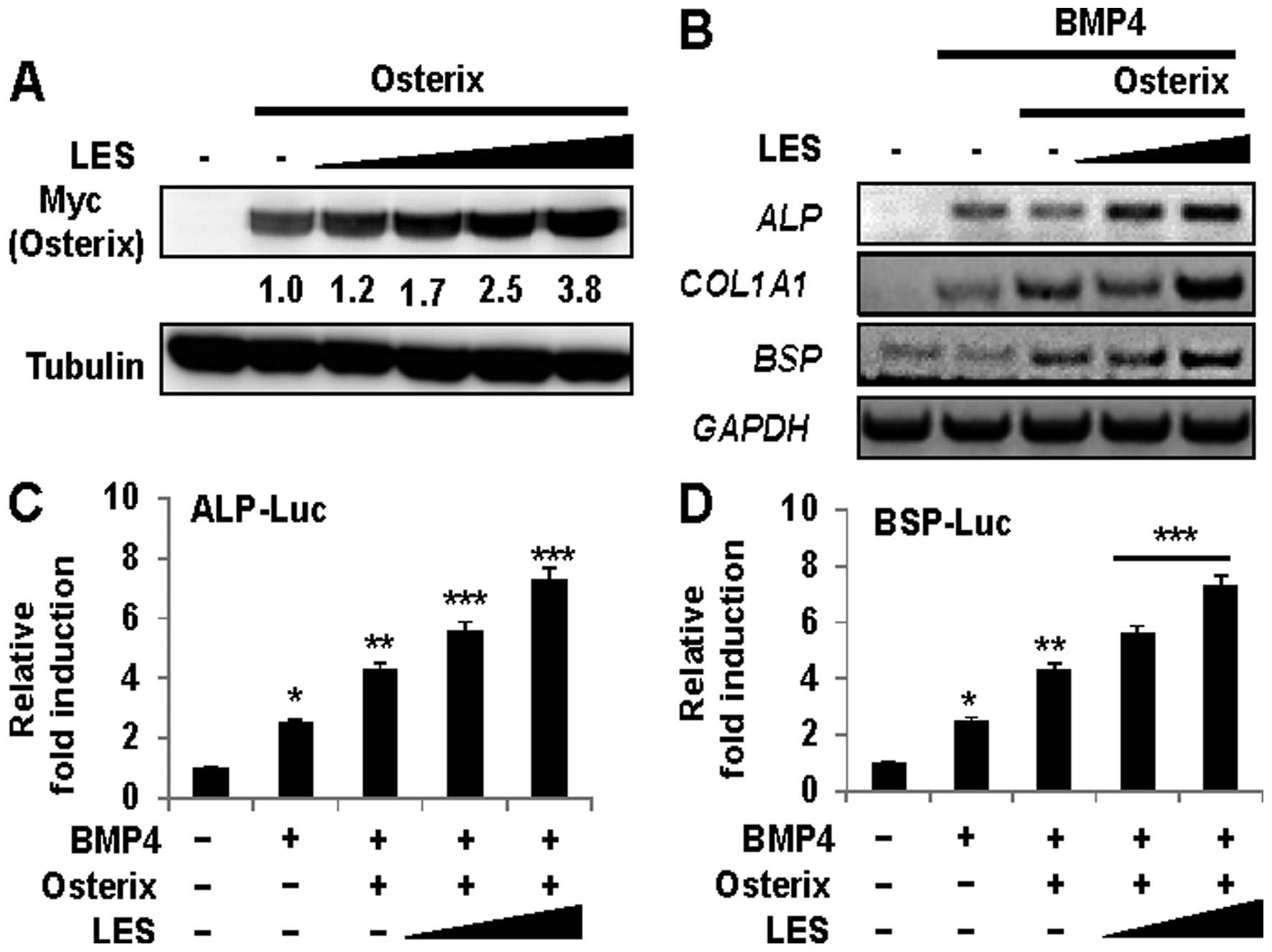
LES regulates the functions of the
osteoblast-specific transcription factor, Osterix, in BMP4-induced
osteoblast differentiation
Osterix has been identified as a zinc
finger-containing transcription factor. It is required for
osteoblast differentiation and bone formation (15). BMP4 induces osteoblast
differentiation and promotes bone formation (40–43). In the present study, we examined
whether LES affects the BMP4-induced osteoblast differentiation of
C2C12 cells. Osteoblast differentiation and mineralization were
measured by ALP staining and Alizarin Red S. LES increased ALP
activity, as shown by ALP staining (Fig. 6A, top panel) and increased ALP
activity (Fig. 6B) in the
presence of Osterix and BMP4. Similarly, LES increased
mineralization, as shown by Alizarin Red S staining (Fig. 6A, bottom panel). We also examined
whether LES regulates the level of ectopically expressed Osterix
protein in non-osteogenic 293 cells. LES significantly enhanced the
level of overexpressed Osterix protein in a dose-dependent manner
(Fig. 7A). In addition, we
examined whether LES affects the transcriptional activity of
Osterix using the osteoblast-specific luciferase reporters, ALP-Luc
(Fig. 7C) and BSP-Luc (Fig. 7D). LES significantly enhanced the
transcriptional activity of the Osterix-induced expression of the
reporters. LES increased the expression of the osteoblast-specific
marker genes ALP, BSP, and COL1A1, in the
cells stimulated with BMP4 (Fig.
7B). Taken together, these results suggest that LES regulates
the expression levels and transcriptional activity of Osterix
during osteoblast differentiation.
Discussion
In this study, we examined the effects of LES, as a
novel osteogenic material, on osteoblast differentiation. In
previous studies, shikonin, from the root of Lithospermum
erythrorhizon, has been shown to protect cartilage in
rheumatoid arthritis, but also to induce osteoclastogenesis in
vitro (38,39). However, hydrophobic shikonin and
its derivatives are not extracted in the water-soluble EtOH
extract, LES.
In our results, we found that LES promoted
osteoblast differentiation and increased the expression levels and
transcriptional activity of Runx2 and Osterix, the master genes of
osteoblast differentiation. Even though the functions of LES in
osteoblastogenesis are not yet well understood, our results
indicate that the water-soluble components of LR could be the main
active components which enhance osteoblastogenesis, instead of
shikonin and its derivative. In other studies, lithospermic acid, a
water-soluble phenolic compound, which is a standardized major
bioactive component at a concentration of 2 mg/g (45), and polysaccharides such as
lithospermans A, B and C (46)
have also been identified in the EtOH extracts of LR. Therefore, a
strong osteoblast differentiation effect by LES may be induced
directly or indirectly by lithospermic acid and lithospermans A, B
and C. However, further studies are required in order to evaluate
the effects of the hydrophilic compounds from LES on
osteoblastogenesis. Lithospermic acid is known to have antioxidant
activity (47) and hypouricemic
activity, and to exert anti-inflammatory effects (48), and anti-diabetic effects (49). However, there has been no study to
date on the effects of lithospermic acid on osteoblast
differentiation, at least to the best of our knowledge. In other
studies, the 95% EtOH extract of LR was shown to exert a
moisturizing effect (50) and the
70% EtOH extract of LR enhanced the epidermal level of ceramides
(51).
The specific master gene of osteoblast
differentiation, Runx2, plays a key role in bone formation. A
loss-of-function mutation of Runx2 in mice results in no
mineralized bone and the lack of mature osteoblasts and osteogenic
differentiation markers (12,52,53). The function of Runx2 is regulated
at several levels, such as transcription, translation,
post-translational modification and protein-protein interactions by
multiple signal transduction pathways. It is known that various
kinases phosphorylate Runx2. The phosphorylation of Runx2 is an
important mechanism that regulates its activity during
osteoblastogenesis. Bone formation is stimulated by the
phosphorylation of Runx2 via the MAPK/ERK signaling pathway
(54,55) or protein kinase C (PKC)-δ
(56). Thus, the regulation of
Runx2 activity by the alteration of its phosphorylation status is
important in bone formation.
Osterix is a novel zinc finger-containing
transcription factor that is essential for the differentiation of
pre-osteoblasts into functional osteoblasts (15). The function of Osterix can be
regulated via post-translational modification by protein
kinase-mediated osteogenesis. In a previous study, p38 was shown to
regulate the expression of osteoblast-specific genes by the
phosphorylation of Osterix (57).
It has also been suggested that Akt induces the phosphorylation of
threonine residue(s) on Osterix during osteoblast differentiation
(58). Tyrosine kinase Src
enhances osteogenic differentiation through the phosphorylation of
Osterix (59).
Previous studies have revealed that shikonin, a
major component of LES is involved in the regulation of ERK1/2 and
Akt phosphorylation (60,61). Therefore, studies determining the
phosphorylation by several protein kinases are warranted in order
to understand the significance of the regulation of LES and the
osteoblast-specific master genes, Runx2 and Osterix. The
identification of protein kinase mechanisms underlying the effects
of LES on osteoblast differentiation will enhance our understanding
of the regulatory mechanisms of Osterix and Runx2 osteogenic
function. Our study provides a basis for understanding the effects
of LES on osteoblast differentiation.
In conclusion, in this study, we demonstrate that
LES enhances osteoblast differentiation. LES modulates the function
of the master genes, Runx2 and Osterix, through the regulation of
their protein expression and transcriptional activity. Thus, LES
may be a potential therapeutic agent for bone diseases, including
osteoporosis. In addition, LR has been approved as a food source by
the Ministry of Food and Drug safety, South Korea (MFDS), and the
EtOH extract of LR (LES) is recognized as a safe food material for
humans. Our data indicate that LES has potential for development as
a potential candidate material for health functional food for bone
health.
Acknowledgments
This study was supported by a grant from the
Cooperative Research Program for Agriculture Science and Technology
Development, Rural Development Administration, Republic of Korea
(project no. PJ01122302).
References
|
1
|
Katagiri T and Takahashi N: Regulatory
mechanisms of osteoblast and osteoclast differentiation. Oral Dis.
8:147–159. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Heino TJ and Hentunen TA: Differentiation
of osteoblasts and osteocytes from mesenchymal stem cells. Curr
Stem Cell Res Ther. 3:131–145. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Teitelbaum SL: Bone remodeling and the
osteoclast. J Bone Miner Res. 8(Suppl 2): S523–S525. 1993.
View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Kawamura N, Kugimiya F, Oshima Y, Ohba S,
Ikeda T, Saito T, Shinoda Y, Kawasaki Y, Ogata N, Hoshi K, et al:
Akt1 in osteo-blasts and osteoclasts controls bone remodeling. PLoS
One. 2:e10582007. View Article : Google Scholar
|
|
5
|
Blum B, Moseley J, Miller L, Richelsoph K
and Haggard W: Measurement of bone morphogenetic proteins and other
growth factors in demineralized bone matrix. Orthopedics. 27(Suppl
1): s161–s165. 2004.PubMed/NCBI
|
|
6
|
Yamaguchi A, Komori T and Suda T:
Regulation of osteoblast differentiation mediated by bone
morphogenetic proteins, hedgehogs, and Cbfa1. Endocr Rev.
21:393–411. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ohba S, Chung UI and Tei Y: Osteoblast
differentiation induced by BMP signaling and Runx2 through Cbfb
regulation. Nihon Rinsho. 65(Suppl 9): 71–74. 2007.In Japanese.
|
|
8
|
Matsubara T, Kida K, Yamaguchi A, Hata K,
Ichida F, Meguro H, Aburatani H, Nishimura R and Yoneda T: BMP2
regulates Osterix through Msx2 and Runx2 during osteoblast
differentiation. J Biol Chem. 283:29119–29125. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Ulsamer A, Ortuño MJ, Ruiz S, Susperregui
AR, Osses N, Rosa JL and Ventura F: BMP-2 induces Osterix
expression through up-regulation of Dlx5 and its phosphorylation by
p38. J Biol Chem. 283:3816–3826. 2008. View Article : Google Scholar
|
|
10
|
Komori T: Regulation of osteoblast
differentiation by transcription factors. J Cell Biochem.
99:1233–1239. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Lee MH, Kwon TG, Park HS, Wozney JM and
Ryoo HM: BMP-2-induced Osterix expression is mediated by Dlx5 but
is independent of Runx2. Biochem Biophys Res Commun. 309:689–694.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Komori T, Yagi H, Nomura S, Yamaguchi A,
Sasaki K, Deguchi K, Shimizu Y, Bronson RT, Gao YH, Inada M, et al:
Targeted disruption of Cbfa1 results in a complete lack of bone
formation owing to maturational arrest of osteoblasts. Cell.
89:755–764. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bendall AJ and Abate-Shen C: Roles for Msx
and Dlx homeoproteins in vertebrate development. Gene. 247:17–31.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Hassan MQ, Javed A, Morasso MI, Karlin J,
Montecino M, van Wijnen AJ, Stein GS, Stein JL and Lian JB: Dlx3
transcriptional regulation of osteoblast differentiation: temporal
recruitment of Msx2, Dlx3, and Dlx5 homeodomain proteins to
chromatin of the osteocalcin gene. Mol Cell Biol. 24:9248–9261.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nakashima K, Zhou X, Kunkel G, Zhang Z,
Deng JM, Behringer RR and de Crombrugghe B: The novel zinc
finger-containing transcription factor osterix is required for
osteoblast differentiation and bone formation. Cell. 108:17–29.
2002. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li H, Marijanovic I, Kronenberg MS, Erceg
I, Stover ML, Velonis D, Mina M, Heinrich JG, Harris SE, Upholt WB,
et al: Expression and function of Dlx genes in the osteoblast
lineage. Dev Biol. 316:458–470. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Komori T: Regulation of skeletal
development by the Runx family of transcription factors. J Cell
Biochem. 95:445–453. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cheng A and Genever PG: SOX9 determines
RUNX2 transactivity by directing intracellular degradation. J Bone
Miner Res. 25:2680–2689. 2010. View
Article : Google Scholar : PubMed/NCBI
|
|
19
|
Stein GS, Lian JB, van Wijnen AJ, Stein
JL, Montecino M, Javed A, Zaidi SK, Young DW, Choi JY and Pockwinse
SM: Runx2 control of organization, assembly and activity of the
regulatory machinery for skeletal gene expression. Oncogene.
23:4315–4329. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee KS, Kim HJ, Li QL, Chi XZ, Ueta C,
Komori T, Wozney JM, Kim EG, Choi JY, Ryoo HM and Bae SC: Runx2 is
a common target of transforming growth factor beta1 and bone
morphogenetic protein 2, and cooperation between Runx2 and Smad5
induces osteoblast-specific gene expression in the pluripotent
mesenchymal precursor cell line C2C12. Mol Cell Biol. 20:8783–8792.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mukherjee A and Rotwein P: Akt promotes
BMP2-mediated osteoblast differentiation and bone development. J
Cell Sci. 122:716–726. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Phimphilai M, Zhao Z, Boules H, Roca H and
Franceschi RT: BMP signaling is required for RUNX2-dependent
induction of the osteoblast phenotype. J Bone Miner Res.
21:637–646. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Javed A, Afzal F, Bae JS, Gutierrez S,
Zaidi K, Pratap J, van Wijnen AJ, Stein JL, Stein GS and Lian JB:
Specific residues of RUNX2 are obligatory for formation of
BMP2-induced RUNX2-SMAD complex to promote osteoblast
differentiation. Cells Tissues Organs. 189:133–137. 2009.
View Article : Google Scholar :
|
|
24
|
Fu H, Doll B, McNelis T and Hollinger JO:
Osteoblast differentiation in vitro and in vivo promoted by
Osterix. J Biomed Mater Res A. 83:770–778. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tu Q, Valverde P and Chen J: Osterix
enhances proliferation and osteogenic potential of bone marrow
stromal cells. Biochem Biophys Res Commun. 341:1257–1265. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kim YJ, Kim HN, Park EK, Lee BH, Ryoo HM,
Kim SY, Kim IS, Stein JL, Lian JB, Stein GS, et al: The
bone-related Zn finger transcription factor Osterix promotes
proliferation of mesenchymal cells. Gene. 366:145–151. 2006.
View Article : Google Scholar
|
|
27
|
Hatta M, Yoshimura Y, Deyama Y, Fukamizu A
and Suzuki K: Molecular characterization of the zinc finger
transcription factor, Osterix. Int J Mol Med. 17:425–430.
2006.PubMed/NCBI
|
|
28
|
Zhang C, Cho K, Huang Y, Lyons JP, Zhou X,
Sinha K, McCrea PD and de Crombrugghe B: Inhibition of Wnt
signaling by the osteoblast-specific transcription factor Osterix.
Proc Natl Acad Sci USA. 105:6936–6941. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
New SA, Robins SP, Campbell MK, Martin JC,
Garton MJ, Bolton-Smith C, Grubb DA, Lee SJ and Reid DM: Dietary
influences on bone mass and bone metabolism: further evidence of a
positive link between fruit and vegetable consumption and bone
health? Am J Clin Nutr. 71:142–151. 2000.PubMed/NCBI
|
|
30
|
Tucker KL, Hannan MT, Chen H, Cupples LA,
Wilson PW and Kiel DP: Potassium, magnesium, and fruit and
vegetable intakes are associated with greater bone mineral density
in elderly men and women. Am J Clin Nutr. 69:727–736.
1999.PubMed/NCBI
|
|
31
|
Chen X, Yang L, Zhang N, Turpin JA,
Buckheit RW, Osterling C, Oppenheim JJ and Howard OM: Shikonin, a
component of chinese herbal medicine, inhibits chemokine receptor
function and suppresses human immunodeficiency virus type 1.
Antimicrob Agents Chemother. 47:2810–2816. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Gao H, Liu L, Qu ZY, Wei FX, Wang SQ, Chen
G, Qin L, Jiang FY, Wang YC, Shang L and Gao CY: Anti-adenovirus
activities of shikonin, a component of Chinese herbal medicine in
vitro. Biol Pharm Bull. 34:197–202. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Jin R, Wan LL, Mitsuishi T, Kodama K and
Kurashige S: Immunomodulative effects of Chinese herbs in mice
treated with anti-tumor agent cyclophosphamide. Yakugaku Zasshi.
114:533–538. 1994.In Japanese. PubMed/NCBI
|
|
34
|
Chen HM, Wang PH, Chen SS, Wen CC, Chen
YH, Yang WC and Yang NS: Shikonin induces immunogenic cell death in
tumor cells and enhances dendritic cell-based cancer vaccine.
Cancer Immunol Immunother. 61:1989–2002. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang XC, Feng J, Huang F, Fan YS, Wang YY,
Cao LY and Wen CP: Effects of shikonin isolated from zicao on lupus
nephritis in NZB/W F1 mice. Biol Pharm Bull. 32:1565–1570. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Kim SJ, Kim JM, Shim SH and Chang HI:
Shikonin induces cell cycle arrest in human gastric cancer (AGS) by
early growth response 1 (Egr1)-mediated p21 gene expression. J
Ethnopharmacol. 151:1064–1071. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Dai Q, Fang J and Zhang FS: Dual role of
shikonin in early and late stages of collagen type II arthritis.
Mol Biol Rep. 36:1597–1604. 2009. View Article : Google Scholar
|
|
38
|
Kim YO, Hong SJ and Yim SV: The efficacy
of shikonin on cartilage protection in a mouse model of rheumatoid
arthritis. Korean J Physiol Pharmacol. 14:199–204. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Youn YN, Lim E, Lee N, Kim YS, Koo MS and
Choi SY: Screening of Korean medicinal plants for possible
osteoclastogenesis effects in vitro. Genes Nutr. 2:375–380. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Kotajima S, Kishimoto KN, Watanuki M,
Hatori M and Kokubun S: Gene expression analysis of ectopic bone
formation induced by electroporatic gene transfer of BMP4. Ups J
Med Sci. 111:231–241. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Li G, Peng H, Corsi K, Usas A, Olshanski A
and Huard J: Differential effect of BMP4 on NIH/3T3 and C2C12
cells: implications for endochondral bone formation. J Bone Miner
Res. 20:1611–1623. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Ou M, Zhao Y, Zhang F and Huang X: Bmp2
and Bmp4 accelerate alveolar bone development. Connect Tissue Res.
56:204–211. 2015. View Article : Google Scholar
|
|
43
|
Zhang Z, Song Y, Zhang X, Tang J, Chen J
and Chen Y: Msx1/Bmp4 genetic pathway regulates mammalian alveolar
bone formation via induction of Dlx5 and Cbfa1. Mech Dev.
120:1469–1479. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Drissi H, Luc Q, Shakoori R, Chuva De
Sousa Lopes S, Choi JY, Terry A, Hu M, Jones S, Neil JC, Lian JB,
et al: Transcriptional autoregulation of the bone related
CBFA1/RUNX2 gene. J Cell Physiol. 184:341–350. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Thuong PT, Kang KW, Kim JK, Seo DB, Lee
SJ, Kim SH and Oh WK: Lithospermic acid derivatives from
Lithospermum erythrorhizon increased expression of serine
palmitoyltransferase in human HaCaT cells. Bioorg Med Chem Lett.
19:1815–1817. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Konno C, Mizuno T and Hikino H: Isolation
and hypoglycemic activity of lithospermans A, B and C, glycans of
Lithospermum erythrorhizon roots. Planta Med. 51:157–158. 1985.
View Article : Google Scholar
|
|
47
|
Jin CJ, Yu SH, Wang XM, Woo SJ, Park HJ,
Lee HC, Choi SH, Kim KM, Kim JH, Park KS, et al: The effect of
lithospermic acid, an antioxidant, on development of diabetic
retinopathy in spontaneously obese diabetic rats. PLoS One.
9:e982322014. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Liu X, Chen R, Shang Y, Jiao B and Huang
C: Lithospermic acid as a novel xanthine oxidase inhibitor has
anti-inflammatory and hypouricemic effects in rats. Chem Biol
Interact. 176:137–142. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Kang ES, Lee GT, Kim BS, Kim CH, Seo GH,
Han SJ, Hur KY, Ahn CW, Ha H, Jung M, et al: Lithospermic acid B
ameliorates the development of diabetic nephropathy in OLETF rats.
Eur J Pharmacol. 579:418–425. 2008. View Article : Google Scholar
|
|
50
|
Chang MJ, Huang HC, Chang HC and Chang TM:
Cosmetic formulations containing Lithospermum erythrorhizon root
extract show moisturizing effects on human skin. Arch Dermatol Res.
300:317–323. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Kim J and Cho Y: Gromwell (Lithospermum
erythrorhizon) supplementation enhances epidermal levels of
ceramides, glucosylceramides, β-glucocerebrosidase, and acidic
sphingomyelinase in NC/Nga mice. J Med Food. 16:927–933. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Mundlos S, Otto F, Mundlos C, Mulliken JB,
Aylsworth AS, Albright S, Lindhout D, Cole WG, Henn W, Knoll JH, et
al: Mutations involving the transcription factor CBFA1 cause
cleidocranial dysplasia. Cell. 89:773–779. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Otto F, Thornell AP, Crompton T, Denzel A,
Gilmour KC, Rosewell IR, Stamp GW, Beddington RS, Mundlos S, Olsen
BR, et al: Cbfa1, a candidate gene for cleidocranial dysplasia
syndrome, is essential for osteoblast differentiation and bone
development. Cell. 89:765–771. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Qiao M, Shapiro P, Kumar R and Passaniti
A: Insulin-like growth factor-1 regulates endogenous RUNX2 activity
in endothelial cells through a phosphatidylinositol
3-kinase/ERK-dependent and Akt-independent signaling pathway. J
Biol Chem. 279:42709–42718. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Xiao G, Jiang D, Gopalakrishnan R and
Franceschi RT: Fibroblast growth factor 2 induction of the
osteocalcin gene requires MAPK activity and phosphorylation of the
osteoblast transcription factor, Cbfa1/Runx2. J Biol Chem.
277:36181–36187. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Kim BG, Kim HJ, Park HJ, Kim YJ, Yoon WJ,
Lee SJ, Ryoo HM and Cho JY: Runx2 phosphorylation induced by
fibroblast growth factor-2/protein kinase C pathways. Proteomics.
6:1166–1174. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Ortuño MJ, Ruiz-Gaspà S,
Rodríguez-Carballo E, Susperregui AR, Bartrons R, Rosa JL and
Ventura F: p38 regulates expression of osteoblast-specific genes by
phosphorylation of osterix. J Biol Chem. 285:31985–31994. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Choi YH, Jeong HM, Jin YH, Li H, Yeo CY
and Lee KY: Akt phosphorylates and regulates the osteogenic
activity of Osterix. Biochem Biophys Res Commun. 411:637–641. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Choi YH, Han Y, Lee SH, Cheong H, Chun KH,
Yeo CY and Lee KY: Src enhances osteogenic differentiation through
phosphorylation of Osterix. Mol Cell Endocrinol. 407:85–97. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
60
|
Öberg AI, Yassin K, Csikasz RI, Dehvari N,
Shabalina IG, Hutchinson DS, Wilcke M, Östenson CG and Bengtsson T:
Shikonin increases glucose uptake in skeletal muscle cells and
improves plasma glucose levels in diabetic Goto-Kakizaki rats. PLoS
One. 6:e225102011. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Gwon SY, Ahn JY, Jung CH, Moon BK and Ha
TY: Shikonin suppresses ERK 1/2 phosphorylation during the early
stages of adipocyte differentiation in 3T3-L1 cells. BMC Complement
Altern Med. 13:2072013. View Article : Google Scholar : PubMed/NCBI
|