Introduction
Pulmonary hypertension (PH) is a complication of
numerous pulmonary conditions. PH is currently classified into five
categories according to different etiologies of the disease as
follows: pulmonary arterial hypertension, PH due to left heart
disease, PH due to lung disease, chronic thromboembolic PH and PH
due to unclear multifactorial mechanisms (1). If left untreated, PH can lead to
right-sided heart failure and even mortality (2,3).
Prolonged exposure to alveolar hypoxia, at a high
altitude or secondary to chronic pulmonary or cardiovascular
diseases, is an important cause of PH (4). Acute hypoxia induces
vasoconstriction that causes a reversible increase in pulmonary
vascular resistance, whereas prolonged hypoxia stimulates vascular
remodeling and persistent vasoconstriction in hypoxic PH by
inducing the proliferation and migration of pulmonary arterial
smooth muscle cells (PASMCs), altering the behavior of pulmonary
arterial endothelial cells (PAECs) and myofibroblasts, and
accumulating the extracellular matrix proteins secreted by the
resident cells (5). Moreover, the
interactions between different types of cells are also critical in
the pathophysiology of PH. For example, PAECs have been shown to
release a variety of factors that stimulate PASMC proliferation
(6,7). A prominent pathological change in
the blood vessels of patients with PH is the muscularization of
distal pulmonary arteries, as evidenced by an in increase in the
number of α-smooth muscle actin (α-SMA)-positive cells in in
situ immunohistochemical analysis (8), implying that PASMCs act as an
essential player in the pathogenesis of PH.
Hypoxia-inducible factor-1 (HIF-1) is a nuclear
transcription factor that functions as a master regulator of
adaptive responses to hypoxia (9). HIF-1 is a heterodimer composed of an
O2-regulated HIF-1α subunit and a constitutively
expressed HIF-1β subunit. Under hypoxic conditions, HIF-1 is
stabilized and activates the transcription of numerous genes, the
products of which are involved in angiogenesis, erythropoiesis,
energy metabolism and cell survival (10). HIF-1 has been implicated in the
pathogenesis of PH based on both experimental and clinical data.
Mice with heterozygous knockout of HIF-1α
(Hif1a+/−) have been shown to display attenuated
manifestations of chronic hypoxia-induced PH, such as
hypoxia-induced muscularization of pulmonary arterioles and medial
wall thickening (11). Recently,
using a mouse model with smooth muscle-specific knockout of HIF-1α,
Ball et al demonstrated that HIF-1α in the smooth muscle
cells played a crucial role in pulmonary vascular remodeling and
the development of PH in response to chronic hypoxia (12). In addition, clinical studies have
revealed a marked elevation in HIF-1α expression in proliferating
PAECs of plexiform lesions within the lungs of patients with severe
PH (13). These findings
implicate HIF-1α in the pathologic alterations in both PASMCs and
PAECs in PH, suggesting that HIF-1α, as a master regulator of
hypoxic responses, is a promising therapeutic candidate for hypoxic
PH. In addition to HIF-1α, a study on HIF-2α, a HIF-1α homologue,
demonstrated that the heterozygous deletion of HIF-2α attenuated
hypoxia-induced PH in mice (14),
suggesting that HIF-2α also contributes to the development of
hypoxic PH; however, whether HIF-3 is involved in the pathogenesis
of PH remains unknown.
In this study, an animal model of hypoxia-induced PH
was established by exposing adult rats to 10% O2 for 3
weeks. The effects of the lentivirus-mediated delivery of HIF-1α
short hairpin RNA (shRNA), which was administered by intratracheal
instillation prior to exposure to hypoxia, on the manifestations of
hypoxia-induced PH were assessed. In addition, rat primary PASMCs
were cultured in vitro and transduced with HIF-1α shRNA in
order to examine the anti-proliferative effects of HIF-1α shRNA on
PASMCs.
Materials and methods
Lentiviruses
An EGFP-encoding lentiviral strains carrying
the shRNA oligonucleotides that target 5′-GACCAACAACUUGAAGAUG-3′ or
5′-GCAGCAGGAATTGGAACGT-3′ on HIF-1α mRNA, namely HIF-1α shRNA #1
and HIF-1α shRNA #2, respectively, were ordered from HanBio
(Shanghai, China). A scramble shRNA with the sequence of
5′-GATCCCCGTTCTCCGAACGTGTCACGTTTCAAGAGAACGTGACACGTTCGGAGAACTTTTT-3′
was cloned in parallel to create a control lentiviral strain.
Animal model of chronic hypoxia-induced
PH and delivery of lentiviruses to the lungs
All animal experimental procedures were approved by
the Animal Care and Ethics Committee of China Medical University,
Shenyang, China and performed in accordance with the Guidelines for
the Care and Use of Laboratory Animals. The development of the
animal model of chronic hypoxia-induced PH and shRNA delivery to
the pulmonary vessels were conducted based on a previously
described method (15). Briefly,
6–8-week-old Sprague-Dawley (SD) rats, purchased from Liaoning
Changsheng Biotech Biotech Co., Ltd. (Benxi, China), were assigned
to the following 5 groups (n=6/group): i) normoxia (untreated); ii)
hypoxia; iii) hypoxia + scramble; iv) hypoxia + HIF-1α shRNA #1;
and v) hypoxia + HIF-1α shRNA #2. Following anesthesia,
1.5×108 U of lentiviruses harboring HIF-1α shRNA or
scramble shRNA was administered to each rat by intratracheal
instillation through the mouth (in a total of 300 µl, and 50
µl was instilled per day for 6 days). Following 6 days of
lentiviral treatment, the rats, apart from those in the normoxia
group, were exposed to hypoxia (10% O2 environment) for
3 weeks to induce PH, and the rats in the normoxia group were
maintained under the normoxic conditions (21% O2) during
the same period.
Determination of right ventricular
systolic pressure
Following 3 weeks of exposure to hypoxia or
normoxia, right ventricular systolic pressure (RVSP) was measured
based on a previously described method (16) using the BL420S animal performance
analyzing system (Thaimeng, Chengdu, China). Briefly, the rats were
anesthetized and placed on their backs. An incision was made on the
neck to expose the trachea, which was then connected to a
respirator. The respiratory parameters of the mechanical
ventilation were set as follows: respiratory rate, 80 breaths/min;
tidal volume, 6–10 ml/kg/min. A catheter was punched in the right
external jugular vein and pushed along the vein to reach the right
ventricle. The other end of the catheter was connected to the
BL420S system for the measurement of RVSP.
Fluorescence microscopy for the
determination of lentiviral transduction to the lungs
Following the determination of RVSP, the rats were
euthanized by an overdose of anesthesia (10% chloral hydrate at 5
ml/kg body weight), and the lungs were excised. A fraction of the
lung tissues was fixed with 4% paraformaldehyde, dehydrated with
sucrose and embedded in optimal cutting temperature (OCT) compound,
and 10-µm-thick cryosections were obtained using the Leica
CM1860 cryosectioning station (Leica, Wetzlar, Germany). The
intensity of green fluorescence protein (GFP) in the lung sections
was examined under an Olympus BX53 fluorescence microscope
(Olympus, Tokyo, Japan) at ×200 magnification.
Isolation of pulmonary arteries from SD
rats
The isolation of the pulmonary arteries was
performed as previously described (17). Briefly, the rats were sacrificed,
and the lungs were excised from the chest cavity and rinsed with
phosphate-buffered saline (PBS). The superficial tissue and the
bronchus artery were discarded with fine micro-scissors, and the
adventitia was removed from the isolated arteries under a
dissecting microscope (AE31; Motic Electric, Xiamen, China). The
isolated arteries were then frozen in liquid nitrogen for later
experiments.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from the rat pulmonary
arteries or the cultured PASMCs using the RNAsimple total RNA kit
(Tiangen, Beijing, China), and then reverse transcribed into cDNA
using Super M-MLV reverse transcriptase (BioTeke, Beijing, China).
The mRNA levels of HIF-1α and β-actin were determined by
quantitative PCR (qPCR) in an Exicycler 96 quantitative
fluorescence analyzer (Bioneer, Daejeon, Korea) using SYBR-Green
Master Mix (Solarbio, Beijing, China) and the following primers:
HIF-1α forward, 5′-CCTACTATGTCGCTTTCTTGG-3′, and HIF-1α reverse,
5′-GTTTCTGCTGCCTTGTATGG-3′; β-actin forward, 5′-GGAGA
TTACTGCCCTGGCTCCTAGC-3′, and β-actin reverse,
5′-GGCCGGACTCATCGTACTCCTGCTT-3′.
Western blot analysis
For total protein extraction, the pulmonary arteries
were physically homogenized and lysed with RIPA lysis buffer
containing 1% v/v PMSF, and the cultured PASMCs were lysed with
NP-40 lysis buffer (both from Beyotime, Haimen, China). Proteins
were separated by sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE), and transferred onto a PVDF membrane
(Millipore, Bedford, MA, USA). The membrane was blocked with 5%
milk and incubated with anti-HIF-1α antibody (1:400; Cat. no.
PB0245; Boster, Wuhan, China) overnight at 4°C, followed by
incubation with horseradish peroxidase (HRP)-conjugated goat
anti-rabbit IgG secondary antibody (1:5000; Cat. no. A0208;
Beyotime) for 45 min at room temperature. Thereafter, the immune
complexes were visualized using the ECL system (Qihai
Biotechnology, Shanghai, China). To verify equal loading and
transfer, the membrane was re-probed with anti-β-actin antibody
(1:1000; Cat.no. sc-47778; Santa Cruz Biotechnology, Inc., Dallas,
TX, USA) and goat anti-mouse IgG (1:5000; Cat. no. A0216;
Beyotime). The film was scanned and analyzed using Gel-Pro-Analyzer
software for densitometric measurement of the target bands using
β-actin as an internal control.
Morphometric and immunohistochemical
analyses
A fraction of the excised lungs was fixed in 4%
paraformaldehyde and embedded in paraffin. Paraffin blocks of the
lung tissues were sliced into 5-µm-thick sections. Some
sections were stained with hematoxylin and eosin (H&E)
following the standard protocol for morphological examination as in
previous studies (18), and
vascular remodeling was assessed based on a previously described
method (15). Briefly, 60–80
intra-acinar vessels per rat accompanying either alveolar ducts or
alveoli were evaluated by two researchers in a blinded manner. The
external diameter and internal diameter of these arteries were
measured by the use of ImageJ software (NIH, Bethesda, MD, USA).
The arterial wall thickness was calculated as follows: percentage
wall thickness = [(external diameter - internal diameter)/external
diameter] ×100.
For immunohistochemical analysis, following antigen
retrieval, the lung sections were treated with 3%
H2O2, blocked with goat serum and incubated
with anti-α-SMA antibody (Abcam, Cambridge, MA, USA) overnight at
4°C, followed by serial incubations with biotin-conjugated goat
anti-mouse IgG secondary antibody and HRP-labeled streptavidin
(both from Beyotime). Diaminobezidine (Sigma-Aldrich, St. Louis,
MO, USA) was added to initiate the chromogenic reaction, and
hematoxylin was used to stain the nuclei. The sections were mounted
and examined under a microscope (DP73; Olympus) at ×400
magnification.
Preparation and culture of PASMCs,
lentiviral transduction and exposure to hypoxia
Another group of 6–8-week-old healthy male SD rats
(purchased from Liaoning Changsheng Biotech Biotech Co., Ltd.) was
used for the isolation of PASMCs by an explant method as previously
described (19). Briefly, the
thoracic aorta was removed and washed with PBS. A longitudinal
incision was made, and the inner endothelial layer was scraped off
gently with a scalpel. The aorta was cut into 1–3 mm2
pieces. The aortic fragments from 2 rats were placed with the inner
surface down in one FBS-rinsed 35 mm2 culture dish.
After 12 h of incubation at 37°C, approximately 5×105
attached cells were obtained. The cells were cultured in Dulbecco's
modified Eagle's medium (DMEM) (Gibco, Carlsbad, CA, USA)
supplemented with 20% fetal bovine serum (FBS; HyClone, Logan, UT,
USA) at 37°C in an atmosphere of 21% O2 and 5%
CO2. The PASMCs were identified by immunofluorescence
staining with anti-α-SMA antibody (1:50), and the images were
captured under an Olympus BX53 fluorescence microscope at ×600
magnification. Cells between passage 3 and 8 were infected with
lentiviruses harboring HIF-1α shRNA #1 or scramble shRNA, and
cultured at 37°C in a normoxic environment for 24 h. Thereafter,
the medium was changed to fresh culture medium and the cells were
placed in a 37°C chamber of 5% O2 and 5% CO2
for exposure to hypoxia for 48 h. Non-transduced cells were
cultured in a normoxic environment (21% O2) or in a
hypoxic environment (5% O2) as the controls.
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
cell proliferation assay
At 24 h after viral infection, the PASMCs were
seeded into 96-well plates at a density of 3,000 cells/well, and
the cells were exposed to hypoxia or normoxia for 48 h. Thereafter,
the cells were incubated with 0.2 mg/ml MTT (Sigma-Aldrich) for 4 h
at 37°C in a normoxic environment. Subsequently, the medium was
carefully aspirated and 200 µl DMSO was added in to each
well to dissolve the MTT-formazan crystals completely. Optical
density (OD) values at 490 nm were measured using an ELX-800
microplate reader (BioTek, Winooski, VT, USA). This experiment was
performed in 5 replicates.
Flow cytometry for cell cycle
analysis
Following viral transduction and 48 h of exposure to
hypoxia or normoxia, the cell cycle of the PASMCs was analyzed by
flow cytometry. The cells were fixed in 70% ethanol for 2 h at 4°C,
and incubated in propidium iodide (PI) solution containing 50
µg/ml RNase A (Beyotime) for 30 min, followed by analysis
using a FACSCalibur flow cytometer (BD Biosciences, Franklin Lakes,
NJ, USA).
Statistical analysis
Raw data were analyzed using GraphPad PRISM software
(version 5.0; GraphPad Software, San Diego, CA, USA). Values are
expressed as the means ± standard deviation (SD) of individual rats
within the group or of 3 independent experiments for the in
vitro assays. Comparison between multiple groups was performed
using one-way analysis of variance (ANOVA), followed by Bonferroni
post-hoc test for comparisons between 2 groups. The difference was
considered statistically significant when P<0.05.
Results
Reduction of hypoxia-induced HIF-1α
expression in pulmonary arteries by RNA interference (RNAi)
The inhibition efficiency of HIF-1α shRNA #1 and
HIF-1α shRNA #2 on HIF-1α expression was tested in an in
vitro pilot experiment (data not shown). The
EGFP-encoding lentiviruses harboring HIF-1α shRNA #1 or
HIF-1α shRNA #2 were administered into the lungs of the SD rats by
intratracheal instillation, and the signals of GFP were
predominantly detected in the cells aligning the pulmonary arteries
and arterioles 3 weeks after lentiviral administration (Fig. 1A). This probably occurred as the
lentivirus underwent replication in the proliferating PAECs and
PASMCs in response to chronic hypoxia, and thus only the
proliferating cells could produce adequate GFP for microscopic
detection. Following exposure to hypoxia for 3 weeks, the
expression of HIF-1α in the pulmonary arteries was increased by
3.59-fold at the mRNA level (P<0.001) and by 3.43-fold at the
protein level (P<0.001), as compared to the normoxia group
(Fig. 1B and C). By contrast,
both HIF-1α shRNA #1 and HIF-1α shRNA #2 effectively reduced the
hypoxia-induced elevation of HIF-1α at the mRNA and protein level
(P<0.001) in the pulmonary arteries, whereas the scramble shRNA
had no effect on HIF-1α expression. These results indicated a
successful lentivirus-mediated gene delivery to the pulmonary
arteries and a marked inhibition efficiency of the HIF-1α shRNAs
against the hypoxia-induced upregulation of HIF-1α.
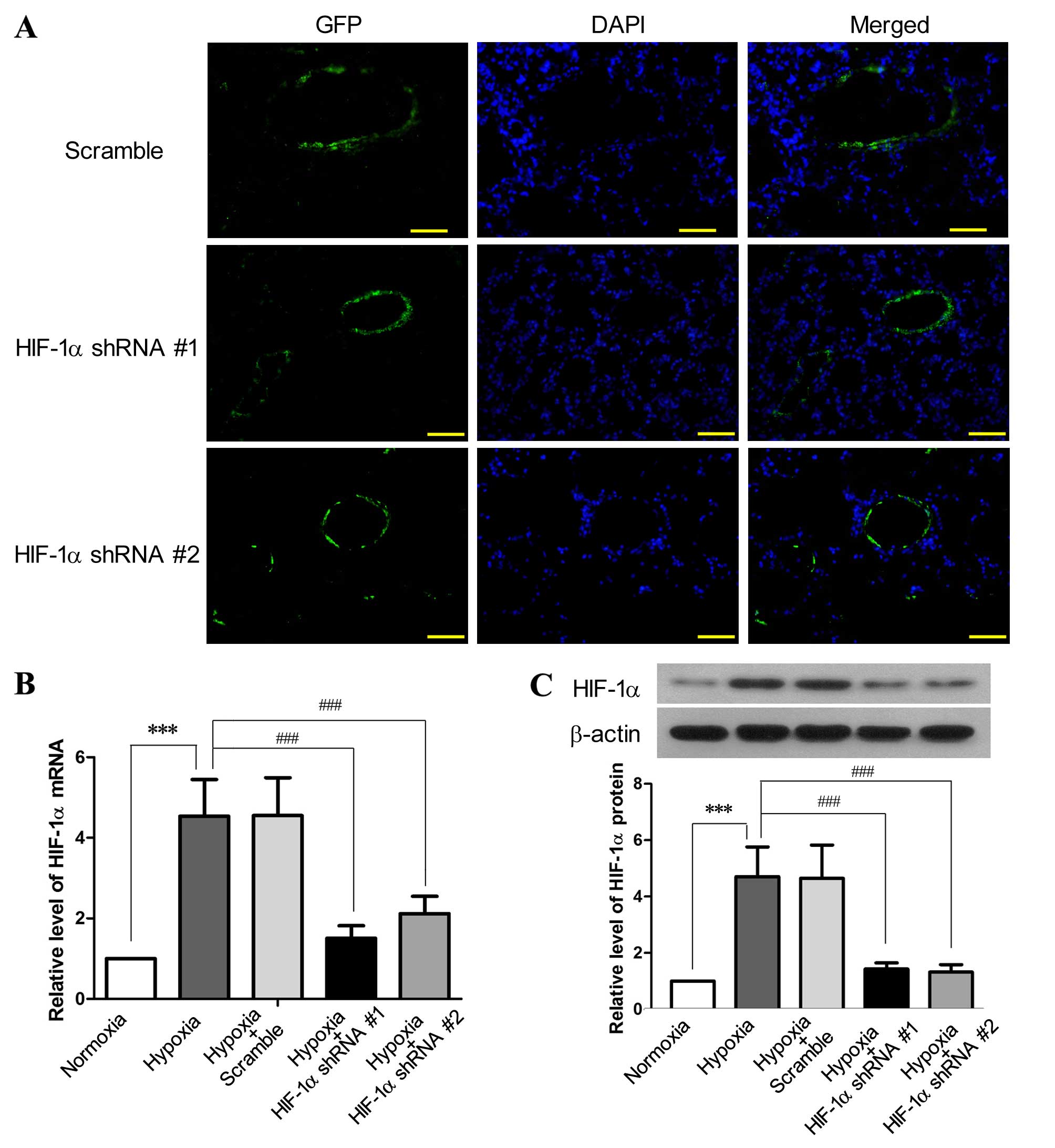 | Figure 1Lentivirus-mediated delivery of
hypoxia-inducible factor-1α (HIF-1α) shRNA to pulmonary arteries
inhibits the hypoxia-induced upregulation of HIF-1α. SD rats
received EGFP-encoding lentiviruses carrying HIF-1α shRNA
#1, HIF-1α shRNA #2 or scramble shRNA by intratracheal instillation
for 6 days, followed by exposure to hypoxia (10% O2) for
3 weeks. SD rats without lentiviral treatment were exposed to
hypoxia or normoxia for 3 weeks as the control (n=6 for each
group). (A) Following exposure to hypoxia, the cryosections of the
lungs from the rats receiving lentiviral treatment were prepared
and examined under a fluorescence microscope (×200 magnification;
scale bar, 100 µm) to detect the expression of green
fluorescence protein (GFP), which was primarily aligned with the
pulmonary vessels. Following lentiviral treatment and exposure to
hypoxia, the rats were sacrificed and the pulmonary arteries were
isolated. The levels of (B) HIF-1α mRNA and (C) HIF-1α protein in
the pulmonary arteries were assessed by RT-qPCR and western blot
analysis, respectively. This figure shows the representative images
from each group, and the values are expressed as the means ± SD.
***P<0.001, hypoxia vs. normoxia;
###P<0.001, hypoxia + HIF-1α shRNAs vs. hypoxia. |
Suppression of the expression of HIF-1α
in pulmonary arteries attenuates hypoxia-induced PH and pulmonary
arterial remodeling
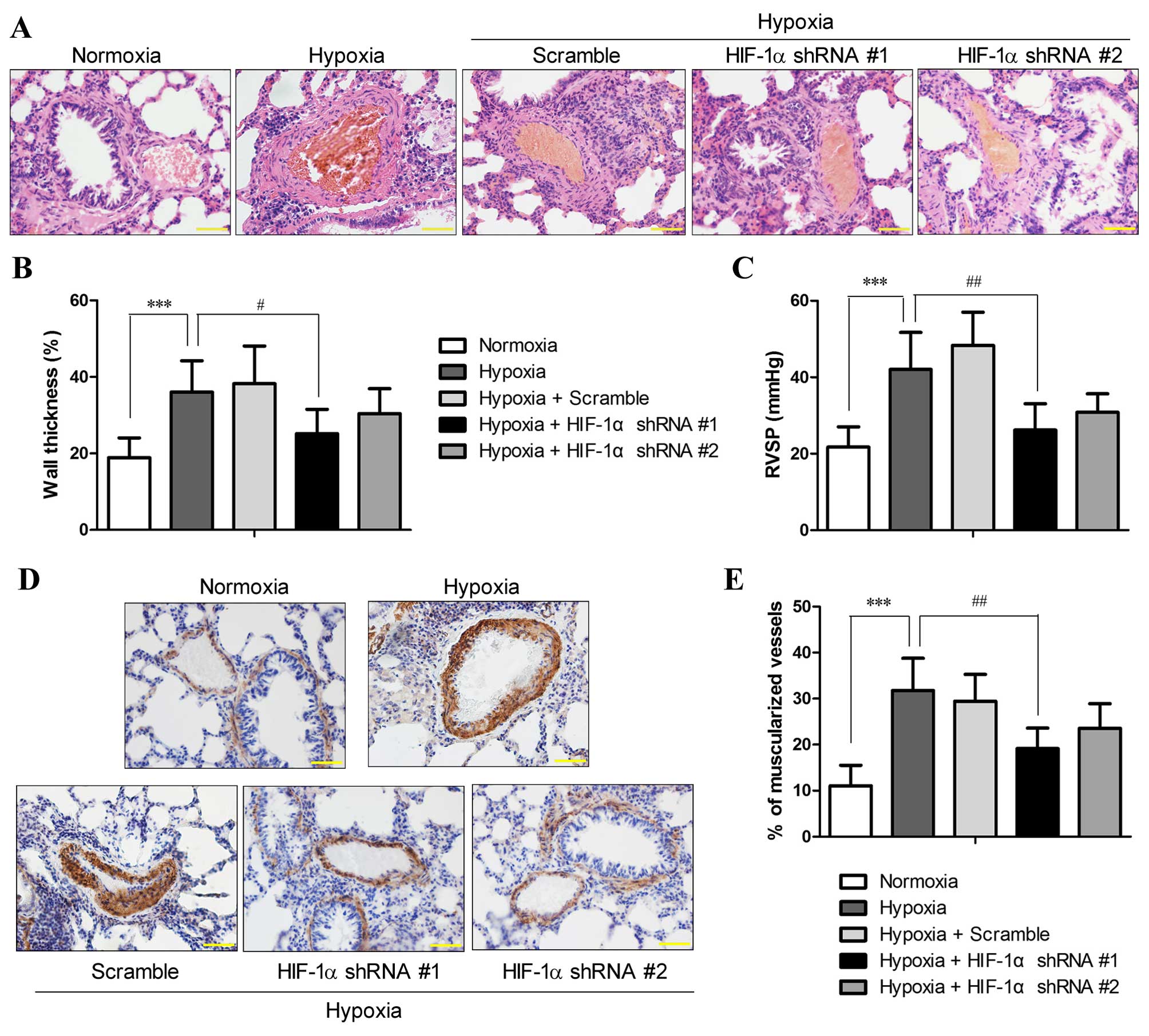
The morphology of the lung vessels following
exposure to chronic hypoxia with and without manipulation of HIF-1α
expression was examined by H&E staining. The results revealed
an increased thickness of the pulmonary arterioles due to smooth
muscle cell (SMC) hypertrophy (increase in cell volume) and
hyperplasia (over-proliferation) (Fig. 2A), which are hallmarks of PH
(20). Morphometric analysis
revealed that chronic hypoxia increased the medial thickness of the
pulmonary arterioles compared with the normoxic controls (36±8.2
vs. 18.9±5.2%; P<0.001), and HIF-1α shRNA #1 significantly
attenuated the hypoxia-induced vascular remodeling and the
thickening of the pulmonary arterioles (P<0.05; Fig. 2B). However, the reduction in
hypoxia-induced vessel wall thickening by HIF-1α shRNA #2 lacked
statistical significance, which was probably due to the small
sample size.
RVSP, an indicator of pulmonary arterial pressure,
was measured in the rats from all groups (n=6 each group). RVSP in
rats under normoxics conditions was 21.73±5.3 mmHg, and the rats
that were exposed to 10% O2 for 3 weeks developed PH
with an RVSP of 42.04±9.7 mmHg, which was significantly higher than
that of the normoxic controls (P<0.01; Fig. 2C). By contrast, the RVSP of the
rats that were pre-treated with HIF-1α shRNA was 26.19±6.9 and
30.83±4.82 mmHg for HIF-1α shRNA #1 and HIF-1α shRNA #2,
respectively. The RVSP values of the rats that were pre-treated
with HIF-1α shRNA were markedly reduced compared to those of the
hypoxia group (P<0.05), and no statistically significant
differences were observed compared to the normoxia group,
suggesting that the rats treated with HIF-1α shRNA did not develop
PH after 3 weeks of exposure to hypoxia.
Immunohistochemical staining for α-SMC, a typical
marker of SMCs, indicated that the hypoxia-induced thickening of
the pulmonary arterioles was attributed to the increased numbers of
SMCs, which was markedly reduced by HIF-1α shRNA #1 and #2
(Fig. 2D). In normal lungs, only
11.1±4.4% of the pulmonary arterioles were fully muscularized,
whereas exposure to chronic hypoxia significantly induced
muscularization of the pulmonary arterioles (31.8±7%; P<0.001).
Compared with the hypoxia group, pre-treatment with HIF-1α shRNA #1
significantly reduced the percentage of fully muscularized
pulmonary arterioles (19.1±4.5%; P<0.01), whereas HIF-1α shRNA#2
reduced the muscularization of pulmonary arterioles to a lesser
extent (23.6±5.3%) (Fig. 2E).
These results demonstrated that the suppression of HIF-1α
expression by RNAi attenuated the symptoms of hypoxia-induced PH
and pulmonary vascular remodeling.
Reduced expression of HIF-1α in PASMCs
inhibits the hypoxia-induced acceleration of cell proliferation and
cell cycle progression
The deregulated proliferation of PAECs is regarded
to play a central role in hypoxia-induced PH and vascular
remodeling as vascular endothelial growth factor (VEGF) has been
demonstrated to be a HIF-1 target that drives vascular remodeling
(21), and PAEC-secreted factors
can also promote PASMC proliferation (6,7).
In this study, we focused on the endogenous role of HIF-1α in
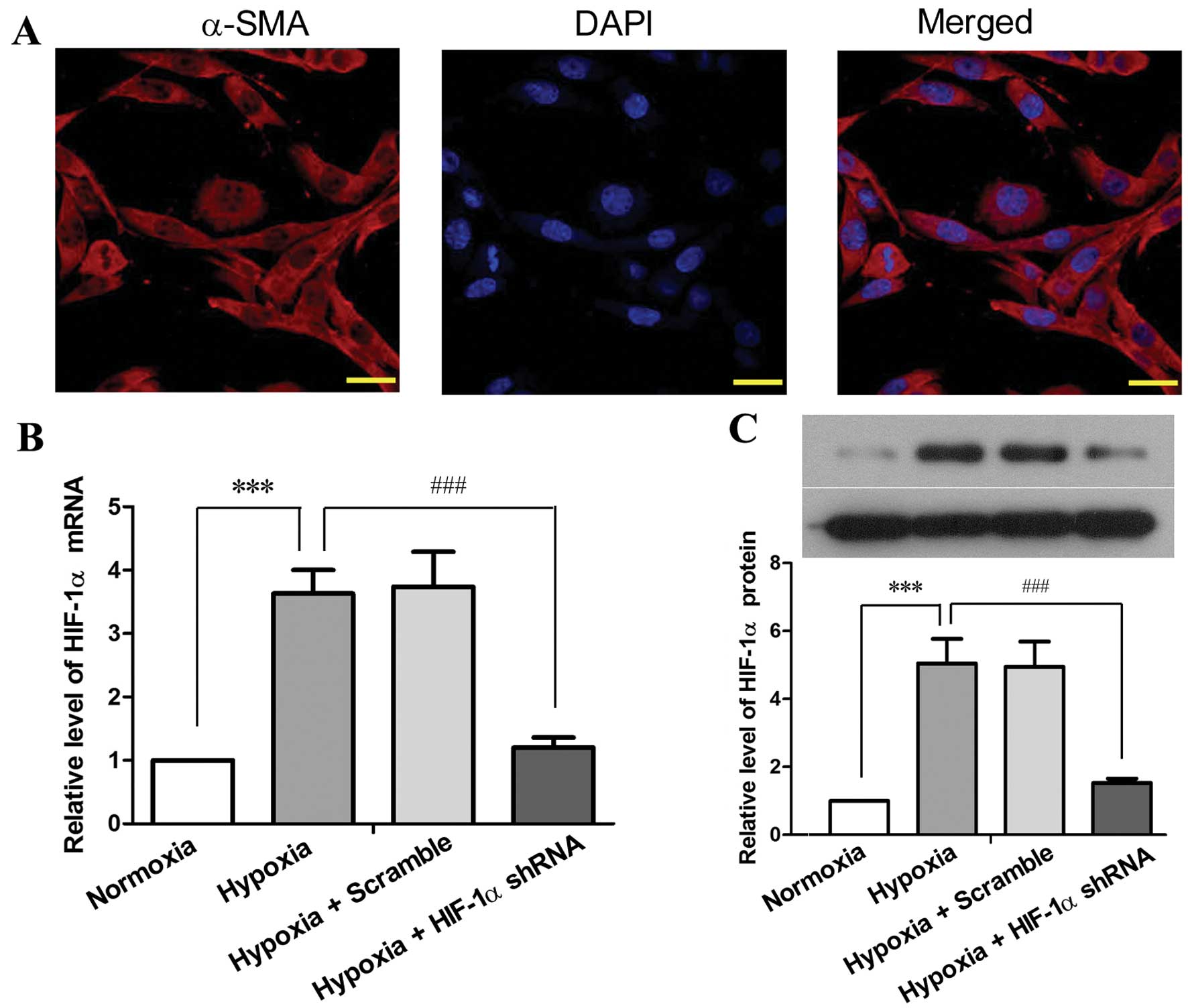
PASMCs. PASMCs were prepared from 6–8-week-old SD rats, and the
purity of the isolated cells was determined by immunofluorescence
staining using a specific antibody against α-SMC (Fig. 3A). Primary PASMCs were cultured
in vitro, and infected with lentiviruses carrying HIF-1α
shRNA #1 or scramble shRNA. Exposure to hypoxia (5% O2)
for 48 h resulted in a 2.63-fold elevation of HIF-1α mRNA
expression (P<0.001) and in a 4.05-fold elevation of HIF-1α
protein expression (P<0.001) in the PASMCs compared with the
cells under normoxic conditions (Fig.
3B and C). By contrast, HIF-1α shRNA reduced the
hypoxia-induced elevation of HIF-1α by 85–90% at the mRNA and
protein level (P<0.001). These results indicated a high
efficiency of the HIF-1α shRNA-mediated inhibition of HIF-1α
expression in PASMCs under hypoxic conditions.
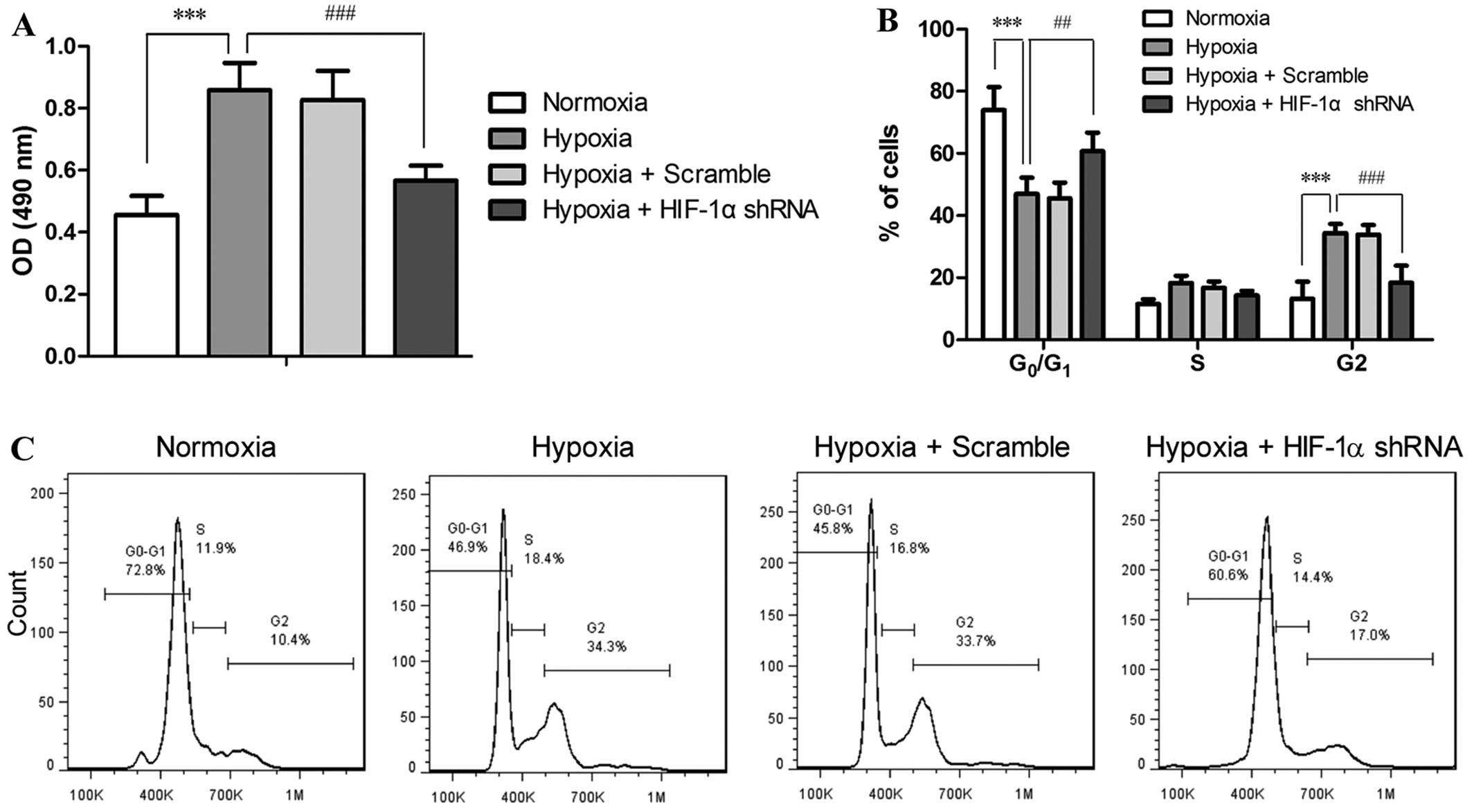
The proliferation and cell cycle of the PASMCs
treated with or without RNAi and exposed to hypoxia were assessed.
As shown in Fig. 4A, 48 h of
exposure to hypoxia doubled the number of PASMCs compared with
those under normoxic conditions (P<0.001), while HIF-1α shRNA
markedly suppressed the hypoxia-accelerated proliferation of the
PASMCs (P<0.001). Cell cycle analysis revealed that hypoxia
disrupted the progression of the cell cycle of PASMCs, as indicated
by the reduced number of cells in the G0/G1
phase (P<0.001) and the accumulation of the cells in the G2
phase (P<0.001) in the hypoxia-exposed cells as compared to the
cells under normoxic conditions (Fig.
2B and C). This result implied hypoxia induced accelerated cell
cycle and enhanced proliferation of PASMCs. On the contrary,
treatment with HIF-1α shRNA significantly attenuated the
hypoxia-induced disruption of the cell cycle distribution at both
the G0/G1 phase (P<0.01) and G2 phase
(P<0.001) in the PASMCs. These data suggest that the
proliferation and cell cycle of PASMCs are directly regulated by
HIF-1α in response to hypoxia.
Discussion
HIF-1α has been demonstrated to play an essential
role in the pathophysiology of chronic hypoxia-induced PH in a mice
with the heterozygous deletion of HIF-1α (11) and in mice with the smooth
muscle-specific disruption of HIF-1α (12). Recently, HIF-1α has been
implicated in several molecular mechanisms accounting for the
pathogenesis of PH (22–24). In addition, the increased
expression of HIF-1α has been found in the pulmonary arteries of
many patients with idiopathic PH (13,25). Although HIF-1α is recognized to be
a key player in PH, surprisingly, the potential of HIF-1α as a
therapeutic target in the treatment of PH has seldom been evaluated
in previous studies. In this study, we employed a
lentivirus-mediated gene delivery approach to introduce HIF-1α
shRNA to the pulmonary vessels in SD rats, and found that the
suppression of the expression of HIF-1α in the pulmonary arteries
effectively attenuated chronic hypoxia-induced PH and pulmonary
vascular remodeling. In addition, the reduction of HIF-1α
expression in primary PASMCs by RNAi significantly inhibited the
hypoxia-induced acceleration of the proliferation and cell cycle of
PASMCs, suggesting that the HIF-1α-mediated over-proliferation of
PASMCs under hypoxic conditions may play an important part in the
development of hypoxic PH.
The protein level HIF-1α is highly regulated by
intracellular O2 tension. Under normoxic conditions,
HIF-1α is continuously synthesized and expressed in the cytoplasm,
but is rapidly degraded by the ubiquitin-mediated pathway. However,
under hypoxic conditions, the low availability of O2
inhibits the hydroxylation of HIF-1α and the subsequent
ubiquitination and proteasomal degradation, resulting in the
accumulation of HIF-1α protein in the nucleus and the initiation of
the transcription of hypoxic responsive genes (26). In addition, the level of HIF-1α is
also regulated at the transcriptional level in response to a
variety of growth factors via the activation of various signaling
pathways, such as the phosphatidylinositol 3-kinase (PI3K) pathway
or mitogen-activated protein kinase (MAPK) pathway (27,28). In this study, exposure to chronic
hypoxia resulted in the elevation of HIF-1α mRNA and protein levels
in the pulmonary arteries of SD rats and in the in vitro
cultured PASMCs, suggesting that, in addition to the instant
protein stabilization in the absence of O2, HIF-1α
expression is enhanced at the transcriptional level as an adaptive
response to chronic hypoxia. Thus, the inhibition of HIF-1α
expression by RNAi is apparently an efficient approach to delay or
alleviate hypoxia-induced pathological alterations.
As a critical feature of pulmonary vascular
remodeling, the hypertrophy and hyperplasia of PASMCs contributed
greatly to the sustained increase of pulmonary vascular resistance
and pulmonary artery pressure in PH (20). In this study, we observed that the
lentivirus-mediated delivery of HIF-1α shRNA effectively inhibited
HIF-1α expression in the pulmonary arteries of SD rats that were
exposed to chronic hypoxia, and such a reduction in HIF-1α
expression markedly attenuated the hypoxia-induced hyperplasia of
PASMCs and the elevation of RVSP, an indicator of pulmonary
arterial pressure. These results suggest a promising therapeutic
potential of HIF-1α shRNA for hypoxic PH.
Previous studies have demonstrated that PAECs can
regulate PASMC proliferation by releasing growth stimulators, such
as serotonin and fibroblast growth factor-2 (FGF-2) (6,7),
or by reducing the production of factors that normally suppress
PASMC proliferation, such as apelin (29). In this study, by culturing PASMCs
in vitro, we demonstrated that hypoxia induced the
acceleration of PASMC proliferation independent of PAECs, but via
HIF-1α-mediated hypoxic responses, suggesting that HIF-1α plays a
critical role in PASMC hyperplasia in hypoxia-induced PH. These
in vitro findings also support the observation in the mice
with PH with smooth muscle-specific deletion of HIF-1α that HIF-1α
in the smooth muscle cells contributes to the development of PH in
chronic hypoxia (12). In
addition to hypoxia, HIF-1α has also been implicated in the growth
factor-induced proliferation of PASMCs, presumably through the
HIF-1-dependent expression of cyclin A (30,31), which may be also involved in the
accelerated cell cycle of PASMCs under chronic hypoxic conditions
in this study.
Our in vivo study demonstrated that the
intratracheal instillation of lentivirus-carried HIF-1α shRNA
effectively attenuated the hypoxia-induced elevation of RVSP,
pulmonary vascular remodeling and the muscularization of
arterioles, providing evidence for the therapeutic potential of
HIF-1α shRNA for hypoxic PH. Genetic medicine has long been
proposed to be of tremendous therapeutic potential and has been
assessed in several diseases (32,33). One major issue associated with the
virus-based gene transduction system is the host immune response,
thus non-viral delivery methods with satisfactory efficiency are
being developed for potential clinical applications (34). Hence, the efficiency of
non-virus-based delivery of HIF-1α shRNA to pulmonary arteries and
its efficacy for the treatment of PH is to be evaluated in future
studies for the potential use of HIF-1α shRNA in the treatment of
PH.
In conclusion, this study demonstrated that the
inhibition of HIF-1α expression by the lentivirus-mediated delivery
of HIF-1α shRNA into rat pulmonary arteries effectively attenuated
the symptoms associated with hypoxia-induced PH, which was, at
least partially, attributed to the suppression of the
hypoxia-induced hyperplasia of PASMCs when HIF-1α was silenced.
Moreover, this study provides the preliminary evidence for the
therapeutic potential of HIF-1α shRNA in the treatment of hypoxic
PH.
References
|
1
|
Simonneau G, Gatzoulis MA, Adatia I,
Celermajer D, Denton C, Ghofrani A, Gomez Sanchez MA, Krishna Kumar
R, Landzberg M, Machado RF, et al: Updated clinical classification
of pulmonary hypertension. J Am Coll Cardiol. 62(Suppl 25):
D34–D41. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
D'Alonzo GE, Barst RJ, Ayres SM, Bergofsky
EH, Brundage BH, Detre KM, Fishman AP, Goldring RM, Groves BM,
Kernis JT, et al: Survival in patients with primary pulmonary
hypertension. Results from a national prospective registry. Ann
Intern Med. 115:343–349. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Humbert M, Sitbon O, Chaouat A, Bertocchi
M, Habib G, Gressin V, Yaïci A, Weitzenblum E, Cordier JF, Chabot
F, et al: Survival in patients with idiopathic, familial, and
anorexigen-associated pulmonary arterial hypertension in the modern
management era. Circulation. 122:156–163. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Shimoda LA and Semenza GL: HIF and the
lung: Role of hypoxia-inducible factors in pulmonary development
and disease. Am J Respir Crit Care Med. 183:152–156. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Stenmark KR, Fagan KA and Frid MG:
Hypoxia-induced pulmonary vascular remodeling: Cellular and
molecular mechanisms. Circ Res. 99:675–691. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Thompson K and Rabinovitch M: Exogenous
leukocyte and endogenous elastases can mediate mitogenic activity
in pulmonary artery smooth muscle cells by release of
extracellular-matrix bound basic fibroblast growth factor. J Cell
Physiol. 166:495–505. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Dewachter L, Adnot S, Fadel E, Humbert M,
Maitre B, Barlier-Mur AM, Simonneau G, Hamon M, Naeije R and
Eddahibi S: Angiopoietin/Tie2 pathway influences smooth muscle
hyperplasia in idiopathic pulmonary hypertension. Am J Respir Crit
Care Med. 174:1025–1033. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Jones PL, Cowan KN and Rabinovitch M:
Tenascin-C, proliferation and subendothelial fibronectin in
progressive pulmonary vascular disease. Am J Pathol. 150:1349–1360.
1997.PubMed/NCBI
|
|
9
|
Wang GL, Jiang BH, Rue EA and Semenza GL:
Hypoxia-inducible factor 1 is a basic-helix-loop-helix-PAS
heterodimer regulated by cellular O2 tension. Proc Natl
Acad Sci USA. 92:5510–5514. 1995. View Article : Google Scholar
|
|
10
|
Semenza GL: Targeting HIF-1 for cancer
therapy. Nat Rev Cancer. 3:721–732. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Yu AY, Shimoda LA, Iyer NV, Huso DL, Sun
X, McWilliams R, Beaty T, Sham JS, Wiener CM, Sylvester JT and
Semenza GL: Impaired physiological responses to chronic hypoxia in
mice partially deficient for hypoxia-inducible factor 1alpha. J
Clin Invest. 103:691–696. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
12
|
Ball MK, Waypa GB, Mungai PT, Nielsen JM,
Czech L, Dudley VJ, Beussink L, Dettman RW, Berkelhamer SK,
Steinhorn RH, et al: Regulation of hypoxia-induced pulmonary
hypertension by vascular smooth muscle hypoxia-inducible factor-1α.
Am J Respir Crit Care Med. 189:314–324. 2014. View Article : Google Scholar :
|
|
13
|
Tuder RM, Chacon M, Alger L, Wang J,
Taraseviciene-Stewart L, Kasahara Y, Cool CD, Bishop AE, Geraci M,
Semenza GL, et al: Expression of angiogenesis-related molecules in
plexiform lesions in severe pulmonary hypertension: Evidence for a
process of disordered angiogenesis. J Pathol. 195:367–374. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Brusselmans K, Compernolle V, Tjwa M,
Wiesener MS, Maxwell PH, Collen D and Carmeliet P: Heterozygous
deficiency of hypoxia-inducible factor-2alpha protects mice against
pulmonary hypertension and right ventricular dysfunction during
prolonged hypoxia. J Clin Invest. 111:1519–1527. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhang R, Shi L, Zhou L, Zhang G, Wu X,
Shao F, Ma G and Ying K: Transgelin as a therapeutic target to
prevent hypoxic pulmonary hypertension. Am J Physiol Lung Cell Mol
Physiol. 306:L574–L583. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang L, Zhou Y, Li M and Zhu Y: Expression
of hypoxia-inducible factor-1α, endothelin-1 and adrenomedullin in
newborn rats with hypoxia-induced pulmonary hypertension. Exp Ther
Med. 8:335–339. 2014.PubMed/NCBI
|
|
17
|
Ko EA, Song MY, Donthamsetty R, Makino A
and Yuan JX: Tension measurement in isolated rat and mouse
pulmonary artery. Drug Discov Today Dis Models. 7:123–130. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Christou H, Morita T, Hsieh CM, Koike H,
Arkonac B, Perrella MA and Kourembanas S: Prevention of
hypoxia-induced pulmonary hypertension by enhancement of endogenous
heme oxygenase-1 in the rat. Circ Res. 86:1224–1229. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Sreejayan N and Yang X: Isolation and
functional studies of rat aortic smooth muscle cells. Methods Mol
Med. 139:283–292. 2007. View Article : Google Scholar
|
|
20
|
Jones R, Zapol WM and Reid L: Pulmonary
artery remodeling and pulmonary hypertension after exposure to
hyperoxia for 7 days. A morphometric and hemodynamic study. Am J
Pathol. 117:273–285. 1984.PubMed/NCBI
|
|
21
|
Hänze J, Weissmann N, Grimminger F, Seeger
W and Rose F: Cellular and molecular mechanisms of
hypoxia-inducible factor driven vascular remodeling. Thromb
Haemost. 97:774–787. 2007.PubMed/NCBI
|
|
22
|
Fukai K, Nakamura A, Hoshino A, Nakanishi
N, Okawa Y, Ariyoshi M, Kaimoto S, Uchihashi M, Ono K, Tateishi S,
et al: Pyk2 aggravates hypoxia-induced pulmonary hypertension by
activating HIF-1α. Am J Physiol Heart Circ Physiol. 308:H951–H959.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shan F, Li J and Huang QY: HIF-1
alpha-induced upregulation of miR-9 contributes to phenotypic
modulation in pulmonary artery smooth muscle cells during hypoxia.
J Cell Physiol. 229:1511–1520. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Chettimada S, Gupte R, Rawat D, Gebb SA,
McMurtry IF and Gupte SA: Hypoxia-induced glucose-6-phosphate
dehydrogenase overexpression and -activation in pulmonary artery
smooth muscle cells: Implication in pulmonary hypertension. Am J
Physiol Lung Cell Mol Physiol. 308:L287–L300. 2015. View Article : Google Scholar :
|
|
25
|
Fijalkowska I, Xu W, Comhair SA, Janocha
AJ, Mavrakis LA, Krishnamachary B, Zhen L, Mao T, Richter A,
Erzurum SC and Tuder RM: Hypoxia inducible-factor1alpha regulates
the metabolic shift of pulmonary hypertensive endothelial cells. Am
J Pathol. 176:1130–1138. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Maxwell PH, Wiesener MS, Chang GW,
Clifford SC, Vaux EC, Cockman ME, Wykoff CC, Pugh CW, Maher ER and
Ratcliffe PJ: The tumour suppressor protein VHL targets
hypoxia-inducible factors for oxygen-dependent proteolysis. Nature.
399:271–275. 1999. View
Article : Google Scholar : PubMed/NCBI
|
|
27
|
Fukuda R, Hirota K, Fan F, Jung YD, Ellis
LM and Semenza GL: Insulin-like growth factor 1 induces
hypoxia-inducible factor 1-mediated vascular endothelial growth
factor expression, which is dependent on MAP kinase and
phosphatidylinositol 3-kinase signaling in colon cancer cells. J
Biol Chem. 277:38205–38211. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhong H, Chiles K, Feldser D, Laughner E,
Hanrahan C, Georgescu MM, Simons JW and Semenza GL: Modulation of
hypoxia-inducible factor 1alpha expression by the epidermal growth
factor/phosphatidylinositol 3-kinase/PTEN/AKT/FRAP pathway in human
prostate cancer cells: Implications for tumor angiogenesis and
therapeutics. Cancer Res. 60:1541–1545. 2000.PubMed/NCBI
|
|
29
|
Alastalo TP, Li M, Perez VJ, Pham D,
Sawada H, Wang JK, Koskenvuo M, Wang L, Freeman BA, Chang HY and
Rabinovitch M: Disruption of PPARγ/β-catenin-mediated regulation of
apelin impairs BMP-induced mouse and human pulmonary arterial EC
survival. J Clin Invest. 121:3735–3746. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Schultz K, Fanburg BL and Beasley D:
Hypoxia and hypoxia-inducible factor-1alpha promote growth
factor-induced proliferation of human vascular smooth muscle cells.
Am J Physiol Heart Circ Physiol. 290:H2528–H2534. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Imanishi M, Tomita S, Ishizawa K, Kihira
Y, Ueno M, Izawa-Ishizawa Y, Ikeda Y, Yamano N, Tsuchiya K and
Tamaki T: Smooth muscle cell-specific Hif-1α deficiency suppresses
angiotensin II-induced vascular remodelling in mice. Cardiovasc
Res. 102:460–468. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Clayman GL, el-Naggar AK, Lippman SM,
Henderson YC, Frederick M, Merritt JA, Zumstein LA, Timmons TM, Liu
TJ, Ginsberg L, et al: Adenovirus-mediated p53 gene transfer in
patients with advanced recurrent head and neck squamous cell
carcinoma. J Clin Oncol. 16:2221–2232. 1998.PubMed/NCBI
|
|
33
|
Manno CS, Pierce GF, Arruda VR, Glader B,
Ragni M, Rasko JJ, Ozelo MC, Hoots K, Blatt P, Konkle B, et al:
Successful transduction of liver in hemophilia by AAV-Factor IX and
limitations imposed by the host immune response. Nat Med.
12:342–347. 2006. View
Article : Google Scholar : PubMed/NCBI
|
|
34
|
Kim SS, Garg H, Joshi A and Manjunath N:
Strategies for targeted nonviral delivery of siRNAs in vivo. Trends
Mol Med. 15:491–500. 2009. View Article : Google Scholar : PubMed/NCBI
|