Introduction
With the progress made in the study of nervous
system diseases, considerable evidence has been provided to support
the hypothesis that oxidative stress, inflammation and apoptosis
are closely associated with the development of neurodegenerative
diseases (1–3). The use of the rat pheochromocytoma
cell line, PC12, as an in vitro model system is quite
acceptable for neurological and neurochemical studies (4,5).
Moreover, apoptosis can be induced by a variety of methods,
including the use of lipopolysaccharide (LPS), a significant
component of the Gram-negative bacteria cell wall, and it has been
widely used in the study of neuronal apoptosis (6). The Toll-like receptor (TLR)4 can
specifically bind with LPS and can thus trigger the release of
inflammatory factors, free radicals and cysteinyl aspartate
specific proteinases (known as caspases) that subsequently cause
apoptosis (7–9). Hence, the development of a novel
drug to reverse neurodegeneration through the inhibition of
apoptosis is feasible. The exploration of new chemicals with high
efficiency and low toxicity for the treatment of neurodegenerative
diseases associated with oxidative stress, inflammation and
apoptosis is of utmost importance.
Bioflavonoids, a group of polyphenolic substances,
are found in most plants and are a sustainable supplement for human
consumption (10). Due to their
widespread availability, coupled with their low toxicity, they can
be developed for use as therapeutic materials (11–14). Naringin (Nar; 4′,
5,7-trihydroxyflavanone-7-rhamnoglucoside) is a proverbial
flavanone glycoside, which is found in abundance in citrus fruit,
grapefruit and juices (15). Nar
has been shown to have multiple biological and pharmacological
properties, including anti-inflammatory, anti-carcinogenic,
lipid-lowering and antioxidant activities (16–18).
In the study of pharmaceuticals for the treatment of
central nervous system diseases, the critical threshold depends on
whether or not these agents can cross the blood-brain barrier
(19). Naringenin
(4′,5,7-trihydroxyflavanone), a metabolic product of Nar, can
easily cross the blood brain barrier (20), and due to this fact, the study of
Nar instantly acquires more importance.
All in all, the mechanisms responsible for the
protective effects of Nar against LPS-stimulated PC12 cell damage
are not well understood. In the present study, we demonstrate that
Nar protects PC12 cells from LPS-induced apoptosis by exerting
antioxidant, anti-inflammatory and anti-apoptotic effects. Firstly,
Nar reduces the level of intracellular reactive oxygen species
(ROS) through the downregulation of cytochrome P450 2E1 (CYP2E1)
expression directly, rather than through the upregulation of
antioxidant-related protein expression, progressively maintaining
the balance of the pro-oxidant and antioxidant enzyme system. Nar
also attenuates the inflammatory response through the
downregulation of the TLR4 pathway. Finally, we also explore the
underlying anti-apoptotic mechanisms in PC12 cells.
Materials and methods
Materials
PC12 rat pheochromocytoma cells were obtained from
Shanghai Biochemistry Co., Ltd (Shanghai, China).
3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT),
RPMI-1640 medium, fetal bovine serum (FBS), penicillin and
streptomycin, were obtained from Sigma Chemical Co. (St. Louis, MO,
USA). 4-(2-Hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES),
acridine orange (AO) and ethidium bromide (EB) fluorescent dyes,
4′,6-damidino-2-phenylindole (DAPI) and TRIzol reagent were from
Nanjing KeyGen Biotech Co. Ltd. (Nanjing, China). The reactive
oxygen assay kit and DCFH-DA were provided by Dojindo Molecular
Technologies, Inc. (Kumamoto, Japan). The Annexin V/propidium
iodide (PI) apoptosis detection kit was obtained from Invitrogen
Life Technologies, Inc. (Carlsbad, CA, USA).
Cell culture and treatment
The PC12 cells have been diffusely used as an analog
neuron model in research (21).
In this study, the cell culture medium contained RPMI-1640 with 5%
FBS, and appropriate penicillin and streptomycin. In the process of
cell culture, the culture medium was changed 3 times a week.
MTT assay and cell viability
In order to determine the efficacy and dose, as well
as optimal treaqtment time, the PC12 cells were treated with
various concentrations (0–2,000 ng/ml) of Nar for 0.5, 1 and 2 h
before being exposed to 400 µg/ml LPS for 18 h at 37°C with
5% CO2. MTT solution was added to terminate the drug
reaction process, and DMSO was then utilized to dissolve the
crystals. The results were gauged by a POLARstar OPTIMA
multi-detection microplate reader (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA) at 490 nm.
Morphological inspection
AO can penetrate viable cells, and is embedded in
nuclear DNA to emit a bright green fluorescence. EB can only
penetrate damaged cells, and is embedded in nuclear DNA to emit
orange fluorescence. DAPI, as a fluorescent dye, can penetrate the
cell membrane and nucleus of the double-stranded DNA. Thus, we used
AO, EB, DAPI as dyes. The half lethal concentrations of LPS and the
optimal protective concentrations of Nar were determined by MTT
assays, and these were then used in subsequent experiments. The
cells were treated with Nar (200, 600 and 1,000 ng/ml) for 1 h
before being exposed to 400 µg/ml LPS for 18 h at 37°C with
5% CO2. The treated PC12 cells were inoculated into
6-well plates and stained with AO, EB and DAPI (the final
concentration of AO and EB was 100 mg/ml, and that of DAPI was 100
ng/ml; these were dissolved in PBS). Staining was carried out at
room temperature for 2–5 min, and the cells were then rinsed well
with PBS. Finally, images of the cells were acquired utilizing an
inverted fluorescence microscope (Olympus BX63; Olympus, Tokyo,
Japan).
Transmission electron microscopy
(TEM)
The PC12 cells were subjected to various treatments
as described above. The cells were collected in 1.5 ml Eppendorf
tubes, and 4% glutaraldehyde was then slowly added along the tube
wall. The tubes were then placed in a refrigerator at 4°C for
preservation. The the process of dehydration, infiltration,
embedding, slicing and heavy metal staining was carried out in
accordance with conventional TEM techniques. A transmission
electron microscope was used to obtain microscopic images.
Determination of apoptosis
The PC12 cells were treated as described above. In
this assay, we used the Annexin V/PI apoptosis detection kit
(Invitrogen Life Technologies, Inc.) and a flow cytometer
(FACSCalibur, Becton-Dickson, San Diego CA, USA). The cells
(1×105/sample) were washed twice with PBS, followed by
centrifugation at 300 × g at 4°C for 5 min. The cells were then
resuspended in the 1X binding buffer (100 µl), followed by
the addition of Annexin V (5 µl) and PI staining solution (5
µl). Finally, the cells were exposed to room temperature for
10 min, avoiding cell-cell contact, and 400 µl 1X binding
buffer was then added. Apoptosis was detected by flow cytometry.
The third quadrant in the flow cytometric plots rerepsents the
distribution of viable cells. The second and fourth quadrant are
considered to indicate the distribution of apoptotic cells.
Intracellular ROS assay
The PC12 cells were treated as described above. The
cells (1×105/sample) were washed twice with PBS,
followed by centrifugation at 300 × g at 4°C for 5 min. DCFH-DA
solution was then added followed by incubation for 30 min in room
temperature. The density distribution of ROS was analyzed by flow
cytometry.
Detection of cytochrome c (Cyto c)
release
The PC12 cells were subjected to various treatments
as described above. The cells were washed with PBS, fixed with 4%
paraformaldehyde for 15 min at 4°C, washed with PBS again, and then
permeabi-lized with 0.2% Trixon-100 for 8 min. Non-specific binding
was blocked by incubating the cells in 3% BSA for 45 min at 37°C,
and the cells were then incubated with the primary antibody
overnight at 4°C. The cells were then washed twice with PBS. The
cells were incubated with the secondary antibody for 1 h at 37°C,
washed with PBS, and then stained with DAPI (1 µg/ml) for 5
min. A laser scanning confocal microscope (TCS SP5, Leica
Microsystems CMS GmbH., Mannheim, Germany) was used to acquire
images. Cyto c was combined with the antibody to emit green
light, and DAPI was used to stain the nuclei blue. In addition, we
respectively extracted the cytoplasmic and mitochondrial proteins
for use in western blot analysis.
Reverse transcrtiption-quantitative PCR
(RT-qPCR)
Total RNA was extracted from the cells using TRIzol
reagent. One microgram total RNA from each sample was then
converted into complementary DNA (cDNA) by reverse transcription
[using the PrimeScript™ RT reagent kit (Takara, Dalian, China)].
Quantitative PCR (qPCR) was performed to gauge the mRNA levels
using the 7500 Real-Time PCR system (Applied Biosystems, Foster
City, CA, USA). The PCR cycling conditions were as follows: 94°C
pre-degeneration for 4 min, 94°C degeneration for 30 sec, 60°C
annealing for 30 sec, 72°C extension for 2 min, for 35 cycles. The
data were analyzed using System SDS software (Applied Biosystems).
The expression level of the GAPDH gene was considered as a
standard. The relative gene expression levels of interleukin
(IL)-1β, IL-6, tumor necrosis factor-α (TNF-α), fatty acid synthase
(Fas), Fas ligand (FasL) and Bcl-2-associated X protein (Bax) were
expressed as a percentage of the expression of GAPDH. Respective
primers (Table I) were applied by
calculating target gene expression.
 | Table IThe primer sequences used for
RT-qPCR. |
Table I
The primer sequences used for
RT-qPCR.
| Genes | GenBank
accession no. | Forward primers
(5′→3′) | Reverse primers
(5′→3′) |
|---|
| GAPDH | NM_017008.3 |
GGCACAGTCAAGGCTGAGAATG |
ATGGTGGTGAAGACGCCAGTA |
| IL-1β | NM_031512.2 |
CCCTGAACTCAACTGTGAAATAGCA |
CCCAAGTCAAGGGCTTGGAA |
| IL-6 | NM_012589.1 |
ATTGTATGAACAGCGATGATGCAC |
CCAGGTAGAAACGGAACTCCAGA |
| TNF-α | NM_012675.3 |
TCAGTTCCATGGCCCAGAC |
GTTGTCTTTGAGATCCATGCCATT |
| Fas | NM_139194.2 |
CACAGCATTCAGTCCTATCCACAGA |
CACAGCCAACCAGATGCTTCA |
| FasL | NM_012908.1 |
CACCAACCACAGCCTTAGAGTATCA |
CACTCCAGAGATCAAAGCAGTTCC |
| Bax | NM_017059.2 |
CGAATTGGCGATGAACTGGA |
CAAACATGTCAGCTGCCACAC |
Western blot analysis
We used lysis buffer to extract the total proteins,
and the extracted proteins were then quantified using the Bradford
assay kit (Blossom Bio Inc., Hangzhou, China). The cells were lysed
using lysis buffer containing phenylmethanesulfonyl fluoride (PMSF)
on ice for 30 min, The lysates were then centrifuged at 20,000 × g
at 4°C for 5 min. The supernatant was placed in −20°C. A total of
50 µg protein was placed on a 12% SDS-PAGE gel and separated
electrophoretically. The target proteins were then transferred onto
PVDF membranes (Millipore, Billerica, MA, USA). After blocking the
PVDF membranes in 5% dried skim milk (Boster Biological Technology,
Wuhan, China) for 3 h at room temperature, the membranes were
incubated overnight at 4°C with primary antibodies (Table II). Subsequently, the membranes
were incubated at room temperature for 2 h with horseradish
peroxidase-conjugated goat anti-rabbit IgG or goat anti-mouse IgG
antibodies at a 1: 2,000 dilution (Beyotime Institute of
Biotechnology, Beijing, China). Protein detection was performed by
enhanced chemiluminescence (ECL) and imaging was carried out using
a BioSpectrum Gel Imaging System (HR410, UVP, USA). In order to
eliminate the variations, the expression level of the GAPDH gene
was considered as a standard. It should be noted that all protein
extractions were carried out once, and thus the internal reference
was same in this experiment. Moreover, in order to detect the
intra- and extranuclear content of Cyto c, we used the
Nuclear and Cytoplasmic Protein Extraction kit (Blossom Bio Inc.)
to extract the protein, and the remaining process was carried out
as described above..
| Table IIInformation for the antibodies used
in the present study. |
Table II
Information for the antibodies used
in the present study.
| Antibodies | Sources | Dilutions | Companies |
|---|
| GAPDH | Rat | 1:5,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| Nrf2 | Rabbit | 1:1,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| HO-1 | Rabbit | 1:1,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| SOD2 | Rabbit | 1:1,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| GSS | Rabbit | 1:1,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| CYP2E1 | Rabbit | 1:1,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| HMGB1 | Rabbit | 1:1,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| COX-2 | Rabbit | 1:1,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| TLR4 | Rabbit | 1:1,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| MyD88 | Rabbit | 1:1,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| TRAF6 | Rabbit | 1:1,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| NF-κB | Rabbit | 1:1,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| AP-1 | Rabbit | 1:1,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| Bak | Rabbit | 1:1,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| Bcl-2 | Rabbit | 1:1,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| Bcl-xL | Rabbit | 1:1,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| Caspase-9 | Rabbit | 1:1,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| Cyto c | Rabbit | 1:1,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| Caspase-8 | Rabbit | 1:1,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| Caspase-3 | Rabbit | 1:1,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| p53 | Rabbit | 1:1,000 | Proteintech Group,
Inc., Chicago, IL, USA |
| p-p38 | Rabbit | 1:1,000 | Bioworld
Technology, Inc., St. Louis Park, MN, USA |
| p-38 | Rabbit | 1:1,000 | Bioworld
Technology, Inc., St. Louis Park, MN, USA |
| p-ERK | Rabbit | 1:1,000 | Bioworld
Technology, Inc., St. Louis Park, MN, USA |
| ERK | Rabbit | 1:1,000 | Bioworld
Technology, Inc., St. Louis Park, MN, USA |
| p-JNK | Rabbit | 1:1,000 | Bioworld
Technology, Inc., St. Louis Park, MN, USA |
| JNK | Rabbit | 1:1,000 | Bioworld
Technology, Inc., St. Louis Park, MN, USA |
Statistical analysis
All data are presented as the means ± standard
deviation (SD). The data between groups were compared by one-way
analysis of variance (ANOVA) and inter-group differences were
compared using a t-test. A value of p<0.05 was considered to
indicate a statistically significant difference.
Results
Effects of Nar on LPS-induced cell
death
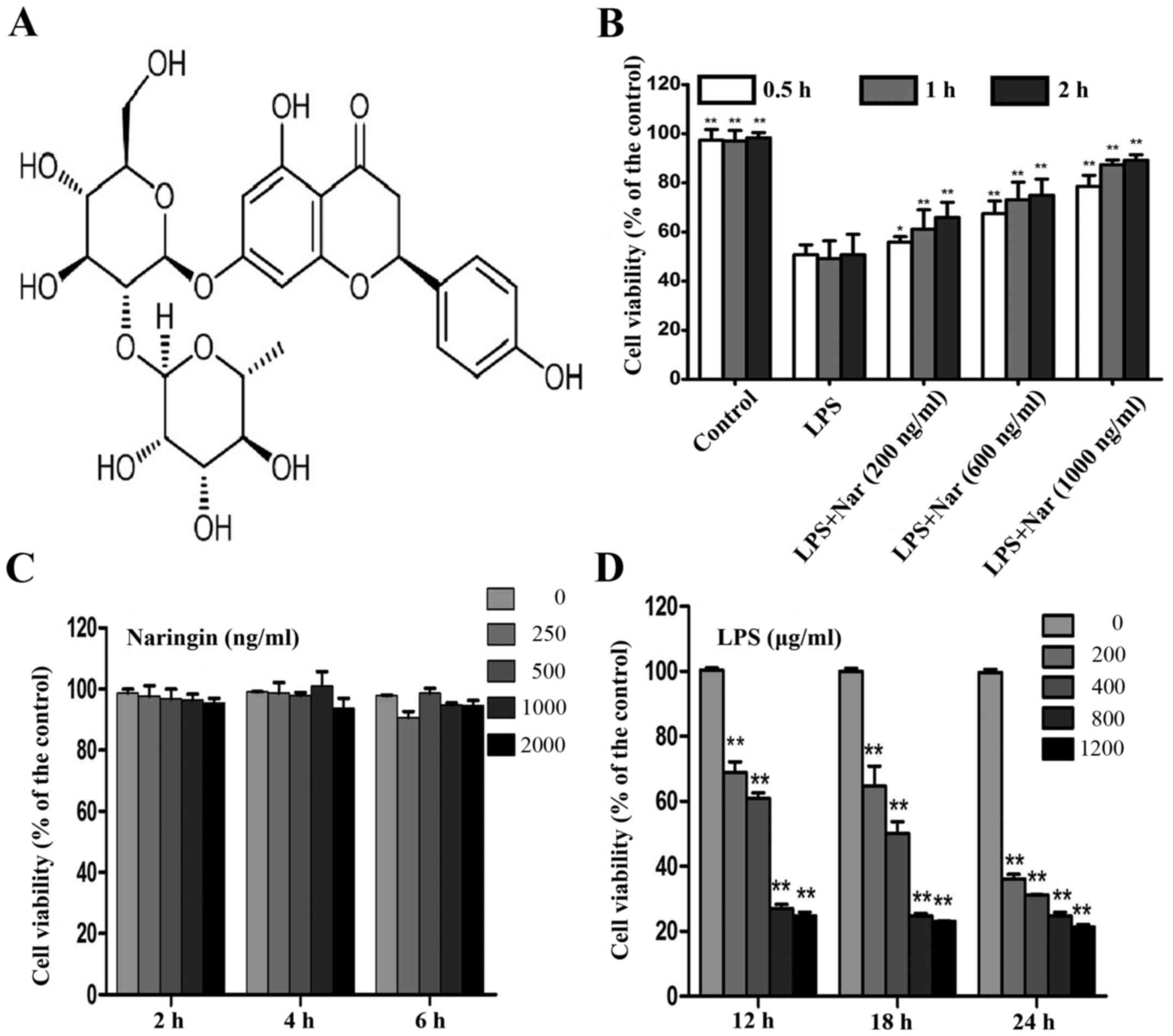
The chemical structures of Nar is shown in Fig. 1A. MTT assay is widely used in
pharmacodynamics experiments (22,23). Treatment with various
concentrations (200, 600 and 1,000 ng/ml) of Nar for 1 h prior to
treatment with LPS (400 µg/ml, 18 h) induced a time and
dose-dependent increase in the cell survival rate (cell viability
increased to 61.2±1.3, 73.1±1.4 and 87.36±1.8%, respectively)
(Fig. 1B).
Treatment with Nar at the range of 0–2,000 ng/ml had
no obvious effect on the survival rate of the PC12 cells (Fig. 1C). Therefore, the Nar
concentrations of 200, 600 and 1,000 ng/ml used in the subsequent
experiments were not considered to be toxic to the cells. In
addition, the viability of the PC12 cells exposed to LPS (400
µg/ml, 18 h) was decreased to 50.15±1.1% (Fig. 1D); thus, LPS at the concentration
of 400 µg/ml was selected for use in the subsequent
experiments to induce cell damage. Our results demonstrated the
potential protective effects of Nar on LPS-induced PC12 cell
viabiltiy in a time- and dose-dependent manner.
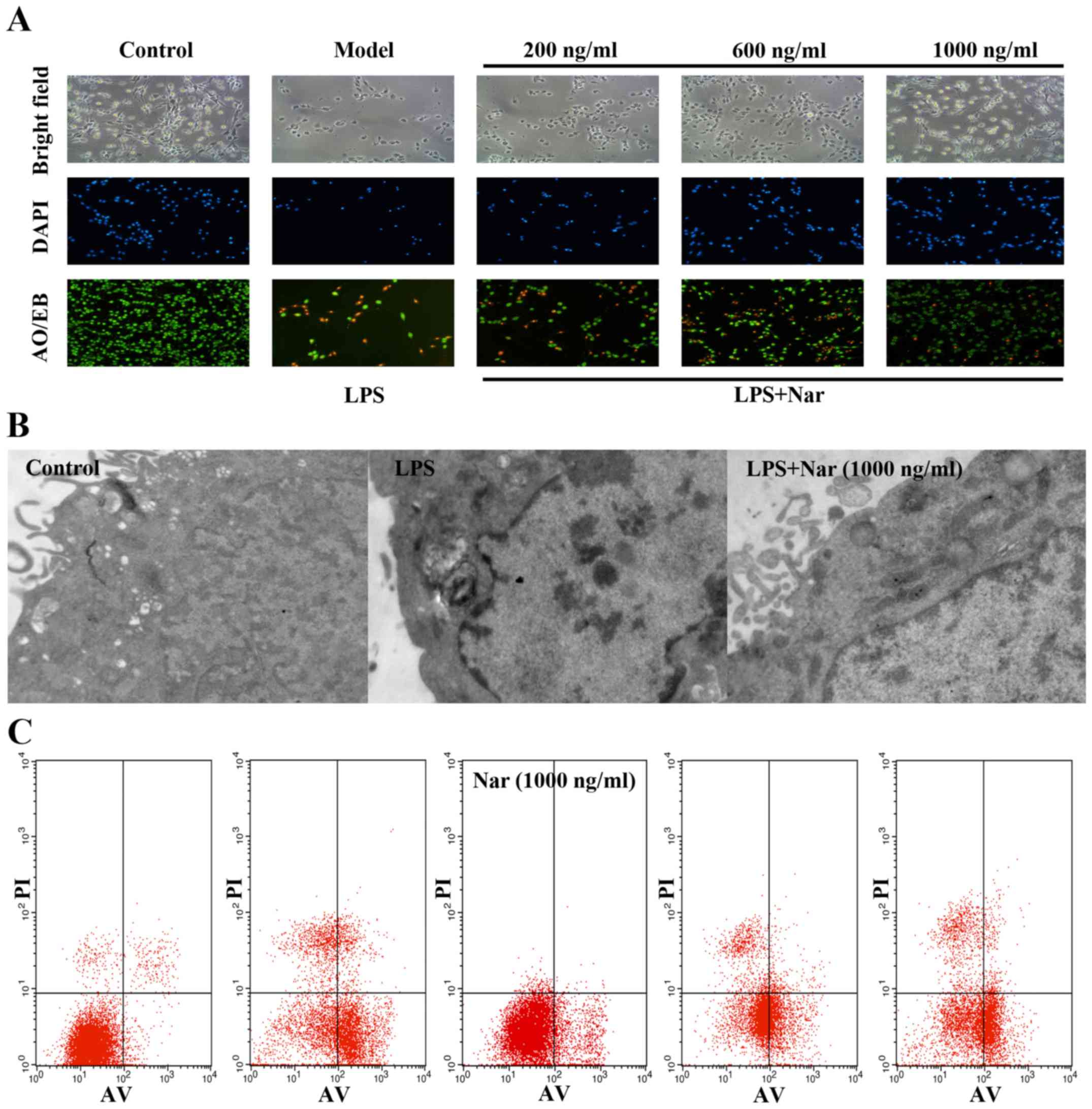
Cell morphology
Black and white microscopic images and images of
AO/EB and DAPI staining provided morphological evidence to confirm
our results. As shown in Fig. 2A,
the control PC12 cells exhibited green fluorescence, whereas the
model group cells exposed to LPS exhibited yellow-stained cells,
indicating apoptosis. The apoptotic cells were significantly
decreased by pre-treatment with Nar (1,000 ng/ml, 1 h), as
evidenced by the decreased number of yellow-stained cells.
In order to further confirm our results, the changes
in the ultrastructure of the PC12 cells were observed under a
transmission electron microscope. As shown in Fig. 2B, the control group cells
exhibited intact cell membranes, intact nuclei, and a large number
of villi structures; however, the cells exposed to LPS exhibited
distinct changes, including cytoplasmic vacuoles, as well as the
disappearance of microvilli structures. Compared with the model
group exposed to LPS, pre-treatment with Nar markedly reversed the
above-mentioned changes.
Finally, flow cytometry was used to measure the
amount of apoptotic cells (Fig.
2C). In relation to the condition, LPS provoked a 14-fold
increase in the number of apoptotic cells (from 3.9% of cells in
the control condition to 55% of cells following exposure to LPS).
However, pre-treatment with Nar decreased the apoptotic rate
significantly (Fig. 2C).
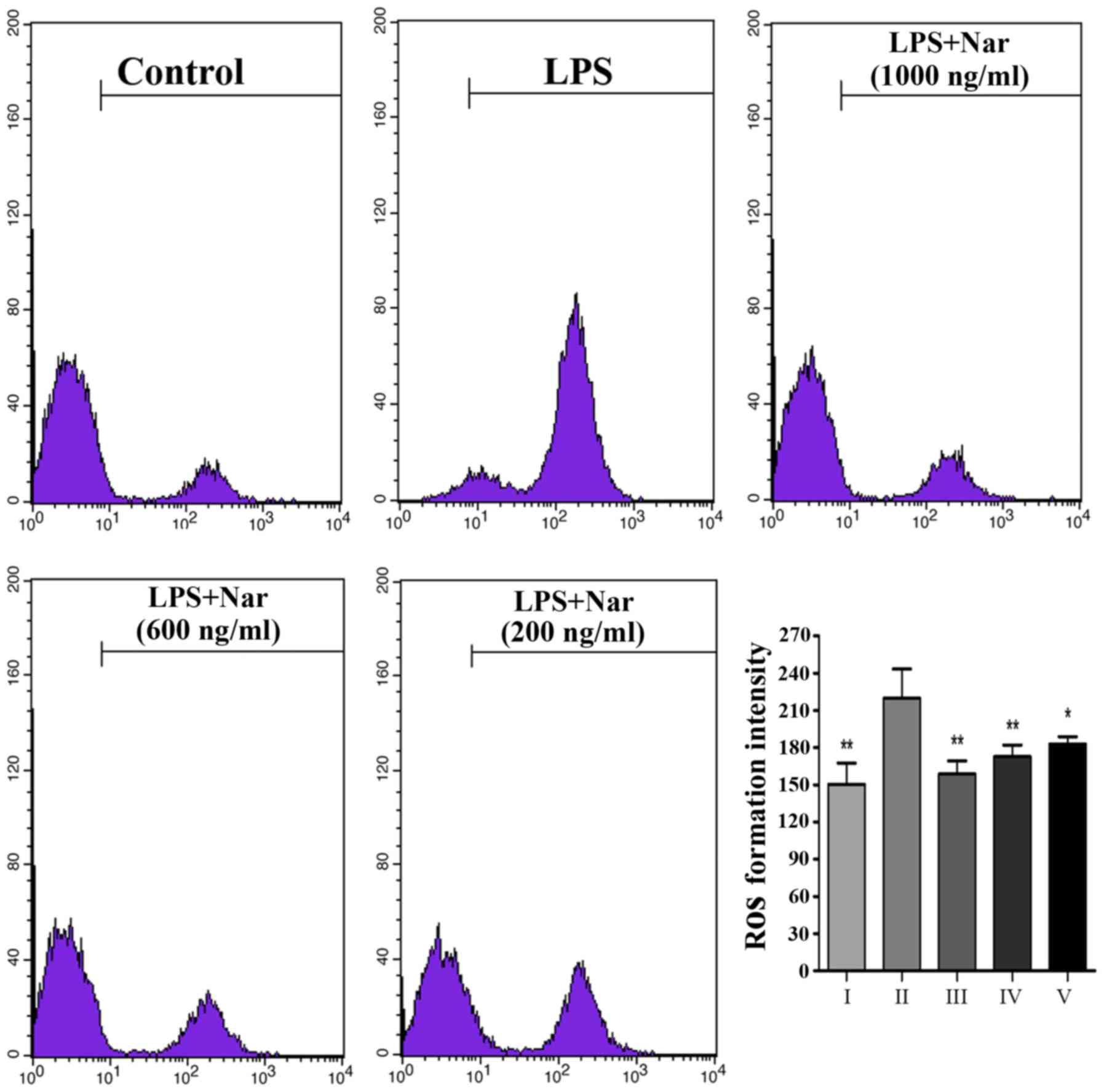
Effects of Nar on intracellular ROS
levels
Exposure to LPS prompted the release of a large
number of ROS from the cells (Fig.
3); however, ROS production was markedly inhibited by
pre-treatment with Nar, particularly pre-treatment Nar at 1,000
ng/ml. Our findings demonstrated that the LPS-induced PC12 cell
apoptosis may be associated with oxidative stress, as also
previously demonstrated (4,24).
However, Nar protected the PC12 cells from apoptosis and attenuated
the production of intracellular ROS.
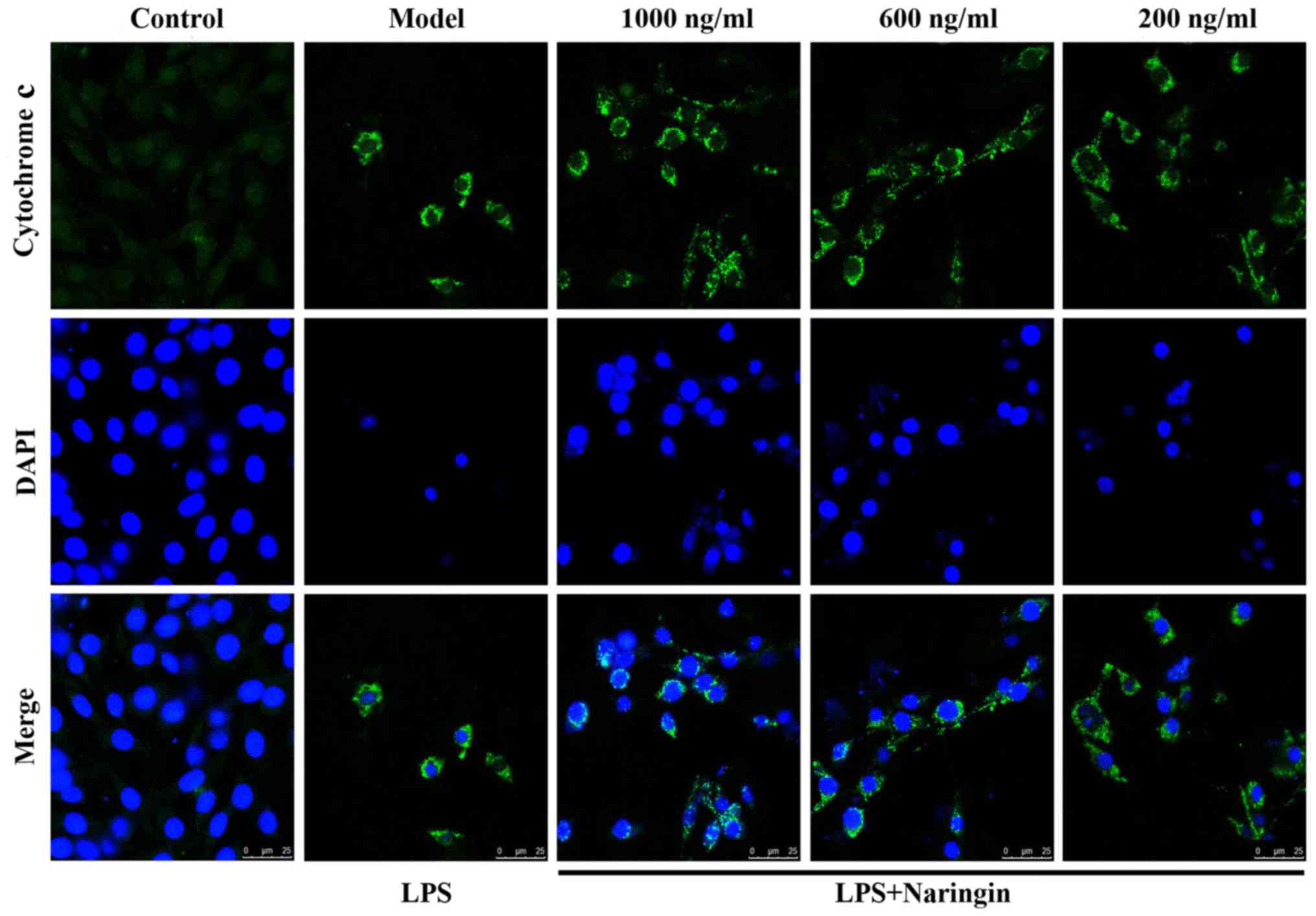
Suppression of Cyto c release
As is known, Cyto c plays a vital role in the
process of apoptosis (25,26).
In Fig. 4, green fluorescence
reflects staining with Cyto c fluorescent antibody, and blue
fluorescence reflects staining with DAPI, which directly reflects
nuclear staining. As shown in the merge image of Fig. 4, little fluorescence was observed
in the control group, whereas in the LPS group, even though the
number of viable cells was significantly decreased, the cell size
was reduced and the nuclear staining also decreased, the content of
extranuclear Cyto c was still evident. Thus, there was a
mass of diffuse cytoplasmic fluorescence. As also shown in Fig. 8G and H, Nar effectively abated the
transfer of Cyto c from the mitochondria to the cytoplasm.
Therefore, it exerted an anti-apoptotic effect.
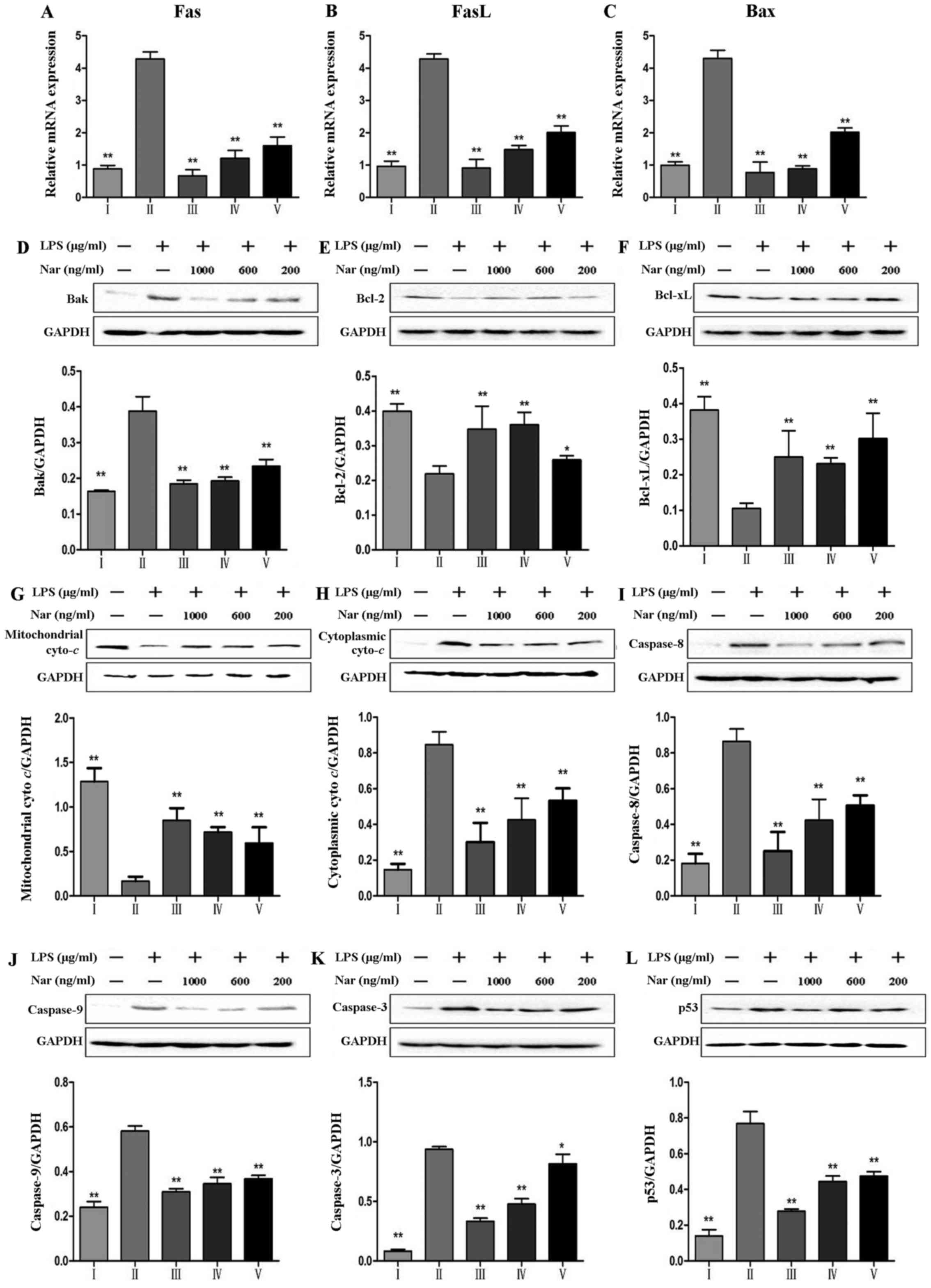 | Figure 8Effects of naringin (Nar) on the
expression of apoptosis-related genes and proteins. mRNA expression
of (A) fatty acid synthase (Fas), (B) Fas ligand (FasL), (C)
Bcl-2-associated X protein (Bax), and (D) protein expression of
Bak, (E) Bcl-2, B-cell lymphoma 2 (Bcl-2), (F) Bcl-xL, (G)
mitochondrial cytochrome c (Mito Cyto c) (H)
cytoplasmic cytochrome c (Cyto Cyto c), (I)
caspase-8, (J) capase-9, (K) caspase-3 and (L) p53.
*p<0.05 and **p<0.05 compared with the
LPS group. For the bar charts: I, control; II, LPS only; III, Nar
at 1,000 ng/ml; IV, Nar at 600 ng/ml; and V, Nar at 200 ng/ml. |
Effects of the Nar on the expression
levels of oxidative stress-related markers
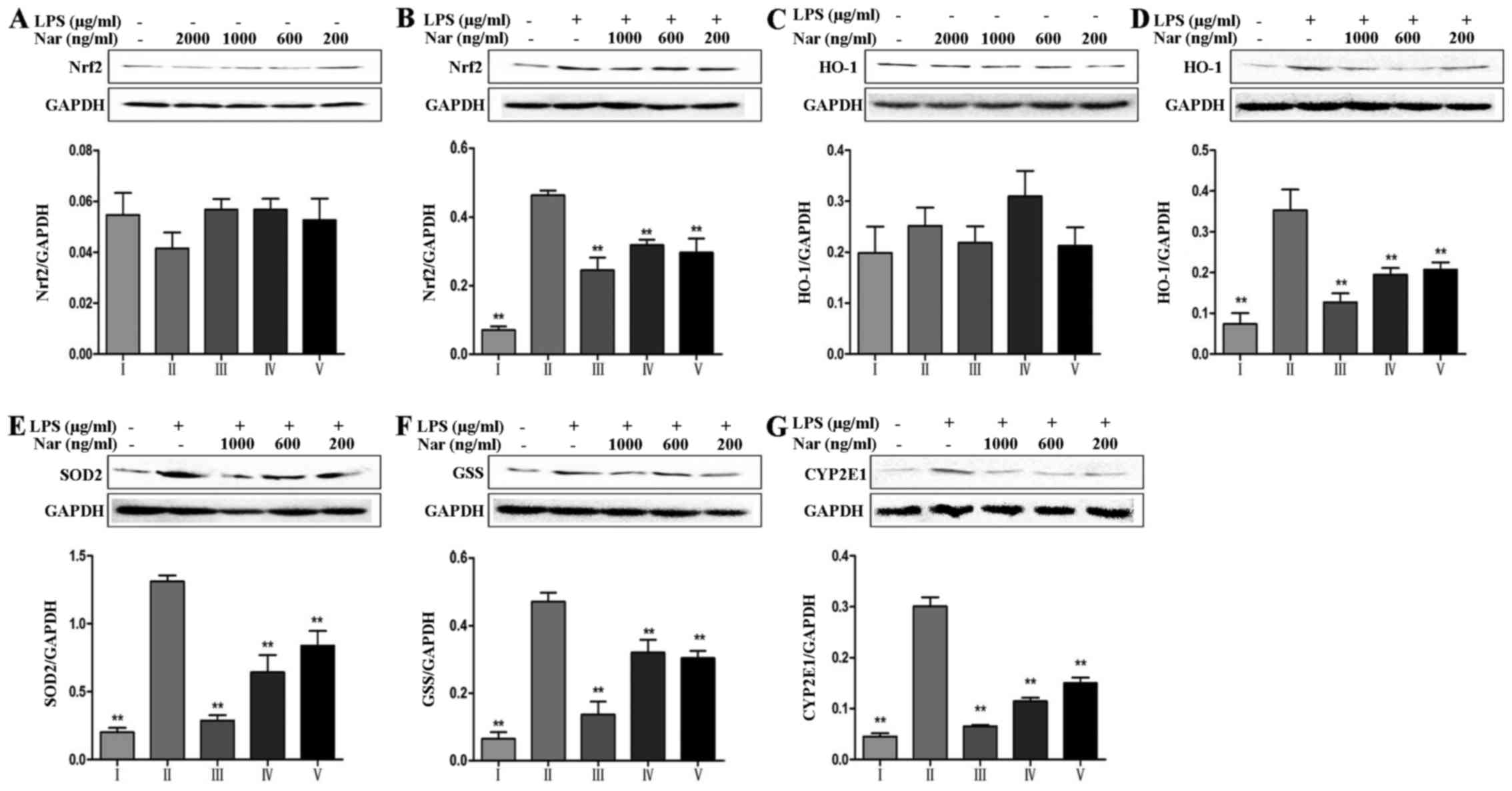
It is worth noting that nuclear factor E2-related
factor 2 (Nrf2) can mediate antioxidant gene expression (27). As shown in Fig. 5A and B, in the absence of LPS,
treatment of the PC12 cells with Nar at any concentration did not
alter the total Nrf2 protein level compared with the untreated
control group. However, in the cells exposed to LPS, the total Nrf2
protein level was significantly increased, indicating the
resistance to the oxidative stress reaction through compensatory
mechanisms. Pre-treatment of the cells with Nar improved the cell
survival rate and gradually corrected the balance of the
intracellular total Nrf2 levels. In addition, other
oxidative-related proteins, including heme oxygenase-1 (HO-1),
superoxide dismutase 2 (SOD2) and glutathione synthetase (GSS) also
exhibited the same trend in expression (Fig. 5C–F).
It is worth emphasizing that CYP2E1 plays an
essential role in the production of ROS (28). As shown in Fig. 5G, relative to the specific
condition, LPS provoked a 6.7-fold increase in the CYP2E1 levels
(from 4.5% of cells in the control condition to 30.1% of cells
exposed to LPS). However, pre-treatment with Nar significantly
decreased the levels of CYP2E1.
Effects of the Nar on the expression
levels of inflammatory cytokines and TLR4-related proteins
As shown in Fig.
6, we detected the expression levels of inflammation-related
genes and proteins, including IL-1β, IL-6, TNF-α, high mobility
group box 1 protein (HMGB1) and cyclooxygenase-2 (COX-2), as well
as TLR4-related proteins, including TLR4, myeloid differentiation
factor 88 (MyD88), TNF receptor-associated factor 6 (TRAF6),
nuclear factor κ-light-chain-enhancer of activated B cells (NF-κB)
and activator protein transcription factor-1 (AP-1), and the
results revealed that LPS promoted the overexpression of
inflammation-related proteins, whereas pre-treatment with Nar
decreased the levels of inflammation-related proteins in a
dose-dependent manner.
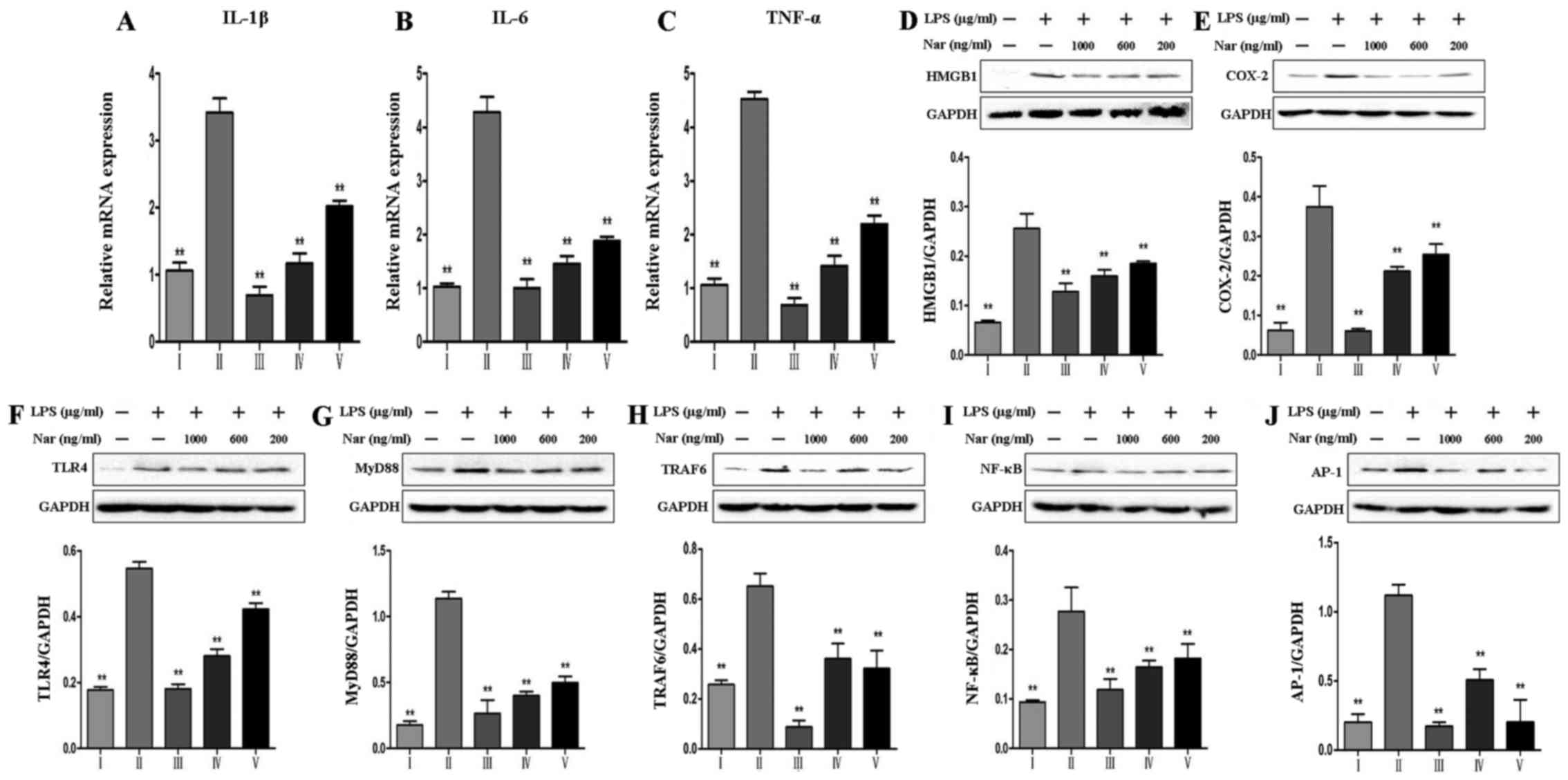 | Figure 6Effects of naringin (Nar) on the
expression of inflammation-related markers. mRNA levels of (A)
interleukin (IL)-1β, (B) IL-6, (C) tumor necrosis factor-α (TNF-α),
and protein expressions of (D) high mobility group box 1 protein
(HMGB1), (E) cyclooxygenase-2 (COX-2), (F) Toll-like receptor 4
(TLR4), (G) myeloid differentiation factor 88 (MyD88), (H) TNF
receptor-associated factor 6 (TRAF6), (I) nuclear factor
κ-light-chain-enhancer of activated B cells (NF-κB) and (J)
activator protein transcription factor-1 (AP-1). Values are
expressed as the means ± SD (n=5 treatment groups).
**p<0.01 compared with the LPS group. For the bar
charts: I, control; II, LPS only; III, Nar at 1,000 ng/ml; IV, Nar
at 600 ng/ml; and V, Nar at 200 ng/ml. |
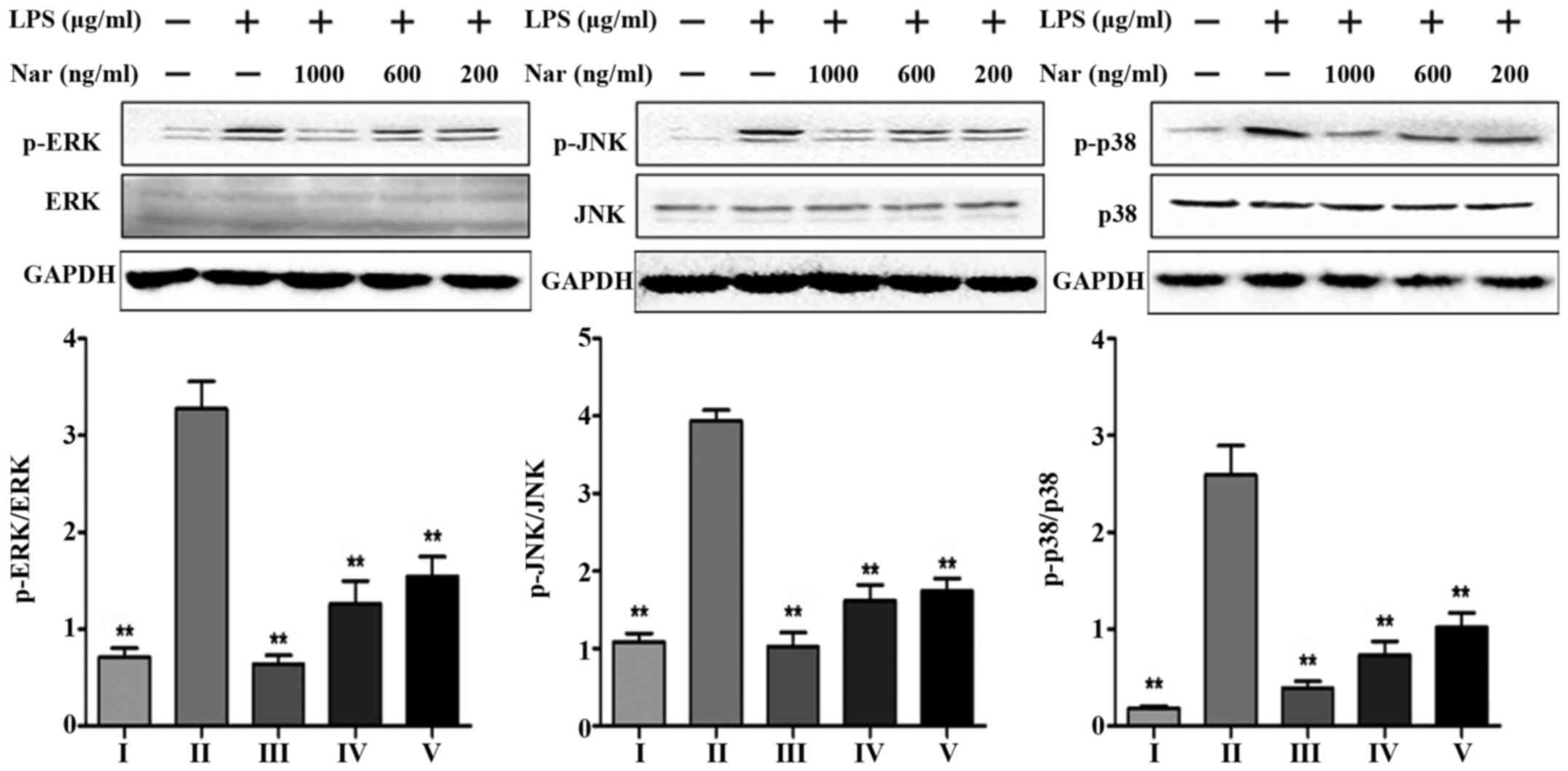
Effects of Nar on the expression levels
of mitogen-activated protein kinase (MAPK) phosphorylation
Consistent with the in vitro experimental
results of the TLR4 pathway (Fig.
6F–J), we detected the levels of MAPK phosphorylation at the
same time. LPS markedly increased the protein expression levels
(p-ERK, p-JNK, p-p38) by 4.6-, 3.6- and 14.3-fold. However, Nar
markedly decreased MAPK phosphorylation (Fig. 7). On the whole, pre-treatment with
Nar significantly inhibited the LPS-induced activation of
extracellular signal-related protein kinase (ERK), c-Jun N-terminal
kinase (JNK) and the p38 signaling pathways. Furthermore, our
results demonstrated that the anti-inflammatory effects of Nar are
associated with the inhibition of the TLR4 pathway.
Effects of Nar on the expression levels
of apoptosis-related genes and proteins
As shown in Fig.
8, LPS increased the expression levels of Fas, FasL and Bax
(Fig. 8A–C). Furthermore, LPS
increased the protein levels of Bak, caspase-9, caspase-8,
caspase-3 and p53, and decreased the levels of the anti-apoptotic
proteins, Bcl-xL, B-cell lymphoma 2 (Bcl-2). However, pre-treatment
with Nar markedly decreased the levels of Fas, FasL, Bax, Bak,
caspase-9, caspase-3 and p53. The levels of Bcl-2 and Bcl-xL were
evidently upregulated by 1.9-fold (from 21.9 to 42.5%) and 3.0-fold
(from 10.5 to 31.7%) (p<0.01) by pre-treatment with Nar (1,000
ng/ml).
Discussion
Nar, a flavanone glycoside of plants and fruits, has
been shown to possess potent antioxidant (29), anti-inflammatory proper ties
(30–32). As it has been previously reported,
the long-term consumption of Nar improves memory in animal models
of neurodegenerative diseases (33–35). To date, to the best of our
knowedge, in vitro research on the effects of Nar on neurons
is limited. Thus, in this study, in order to explore and confirm
the potential neuroprotective effects of Nar, we examined the
effects of Nar on the LPS-induced apoptosis of PC12 cells in
vitro. In addition, we investigated the potential underlying
mechanisms.
On the one hand, chronic toxicological assessment
analysis of Nar has indicated that Nar is a substance with low
toxicity (36), and our results
also demonstrated this fact. On the other hand, naringenin, a
metabolic product of Nar, can easily cross the blood brain barrier
to protect brain cells (20). All
these data make the research of Nar feasible.
Oxidative stress is closely associated with
neurodegenerative diseases (37).
ROS play an essential role in oxidative stress. In the present
study, LPS markedly increased the release of intracellular ROS from
PC12 cells; however, pre-treatment with various concentrations of
Nar decreased the intracellular ROS levels. It should be noted that
CYP2E1 is also associated with LPS-induced oxidative stress and ROS
production (38). Our results
from western blot analysis demonstrated this view, which also
revealed that the protective effects of Nar against LPS-induced
PC12 cell apoptosis may be associated with its specific inhibition
of CYP2E1.
In addition, the enhanced tolerance capacity against
oxidetive stress gained our attention (Fig. 5). The protein levels of Nrf2,
HO-1, SOD2 and GSS increased following exposure to LPS. Of note,
the levels of these proteins were not altered when Nar treatment
was used alone. Therefore, we speculate that Nar can not only
enhance the antioxidant capacity through the downregulation of
CYP2E1 protein expression, but can also maintain the balance of
intracellular antioxidant protein expression.
LPS is a major activator of inflammation and a
ligand for TLR4 (39). Upon LPS
stimulation, inflammation is generated by TLR4 through the
activation of downstream proteins, such as MyD88, TRAF6, NF-κB,
MAPKs and AP-1, resulting in the production of a wide range of
pro-inflammatory cytokines and chemokines (40,41). Moreover, NF-κB is an inducible
transcription factor, which can be sensitized by TNF-α, IL-1β, LPS
and ROS (42–44). The present study demonstrated that
LPS has the ability to cause the robust transcriptional activity of
NF-κB and to markedly increase the expression of pro-inflammatory
cytokines, leading to PC12 cell apoptosis. However, all of changes
were reversed by pre-treatment with Nar. In addition, our results
of western blot analysis demonstrated that the phosphorylation of
ERK, p38, JNK was inhibited in the presence of Nar. Thus, it is
possible that Nar mediates the TLR4-related inflammatory pathway,
and thereby alleviate inflammation in PC12 cells.
Apoptosis is accompanied by increased mRNA levels of
pro-inflammatory cytokines, such as IL-1β, IL-6, TNF-α (45). COX-2 is one of inflammatory
early-response proteins (46),
and its overexpression is associated with the stimulation of HMGB1.
HMGB1 is a member of the damage-associated molecular pattern (DAMP)
family of proteins that is secreted by necrotic brain cells
(47). HMGB1 can bind to its
receptor (TLR4) for mediating the inflammatory reaction (48); however, the potential mechanisms
of action of the HMGB1-TLR4 pathway remain unclear. Nevertheless,
in this study, Nar downregulated the levels of HMGB1 and
TLR4-related proteins, and these results indicated that Nar
effectively mitigated the secretion of inflammatory cytokines.
Finally, the death of PC12 cells induced by LPS is
considered the common effect of oxidative stress, and inflammatory
and apoptosis reactions. Bcl-2 is a critical unit for pro-survival
function by maintaining the mitochondrial membrane. p53 is a
critical suppressor molecule for Bcl-2. Upon oxidative or
inflammatory stimulation, p53 is activated and associates with
Bcl-2 by direct binding. The association of p53 with Bcl-2 can
promote Bcl-2 dissociation from the mitochondria and accelerate the
permeabilization of the mitochondrial membrane. This causes Cyto
c release from the mitochondria to the cytoplasm and
activates downstream of caspase apoptotic cascades (49,50). Caspase-3 is considered to be a
unique factor of apoptosis in this cascade reaction. On the one
hand, Fas/FasL and caspase-8 are the upstream regulators of
caspase-3 (51), and LPS can
upregulate caspase-3 through Fas/FasL-caspase-3 pathway; on the
other hand, upon LPS stimulation, LPS can downregulate Bcl-2,
Bcl-xL, and upregulate Bax, Bak, cytoplasmic Cyto c and
capase-9 (52). Finally, the
caspase-3-related pathway is activated which eventually leads to
apoptosis. The present study demonstrated that Nar markedly
downregulate the protein expression levels of p53, Bak, cytoplasmic
Cyto c, caspase-9, caspase-8 and caspase-3, and the mRNA
expression levels of Fas, FasL and Bax; Nar increased the
expression of Bcl-2 and Bcl-xL. Our results demonstrated that the
Nar suppressed LPS-induced PC12 cell apoptosis by mediating the
expression of a series of caspase-3-related proteins.
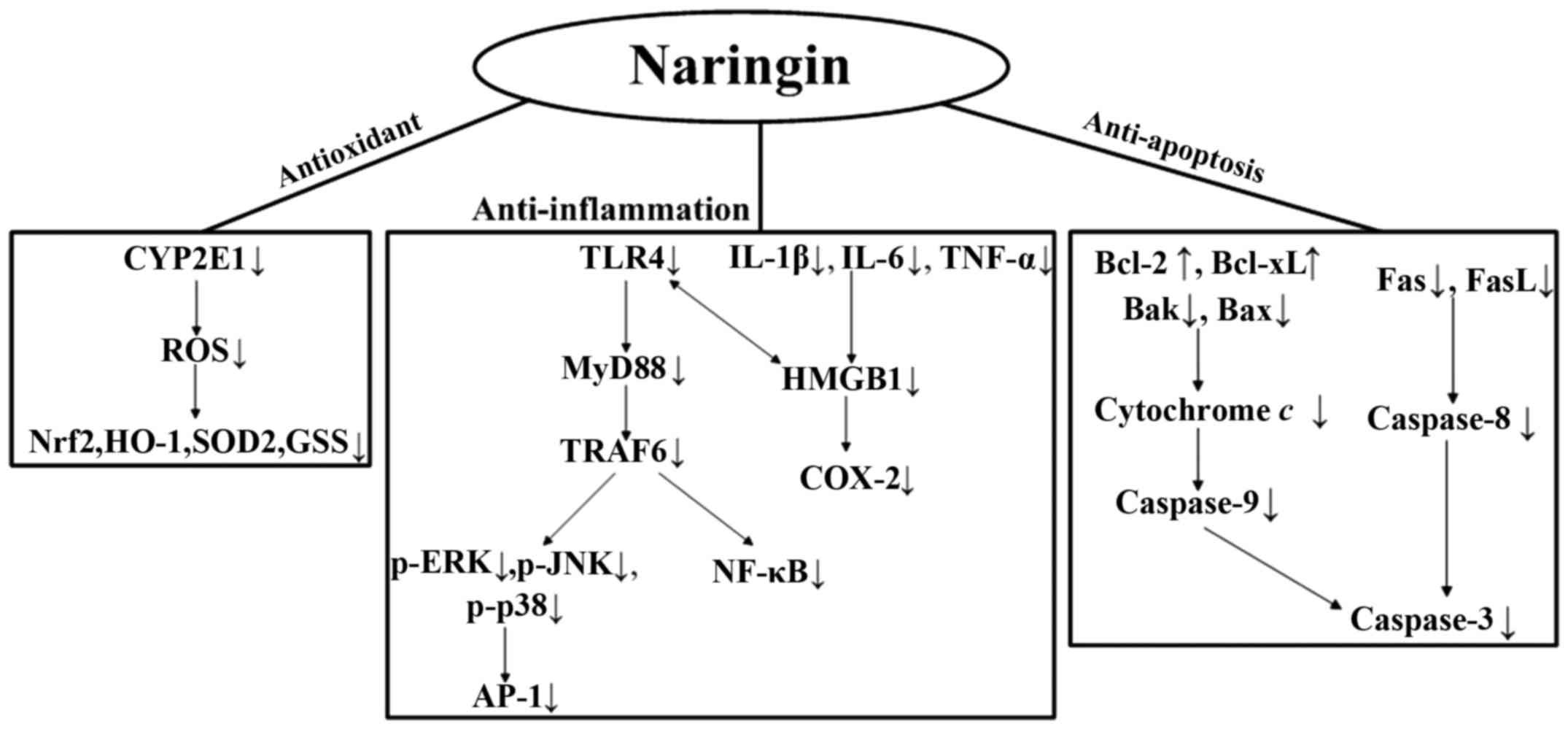
In conclusion, our results demonstrate for the first
time, at least to the best of our knowledge, that Nar reverses the
oxidative stress, inflammation and the apoptosis of PC12 cells
resulting from exposure to LPS. In Fig. 9, we summarize the underlying
mechanisms responsible for the protective effects of Nar. As
neuroinflammation is involved in the development of
neurodegenerative diseases, Nar seems to act as a possible
neuroprotective substance. Further studies are warranted to fully
determine the protective effects of Nar and to elucidate all the
underlying molecular mechanisms.
Abbreviations:
|
LPS
|
lipopolysaccharide
|
|
CYP
|
cytochrome P450 superfamily
|
|
CYP2E1
|
cytochrome P450 2E1
|
|
ROS
|
reactive oxygen species
|
|
Nrf2
|
nuclear factor erythroid 2-related
factor 2
|
|
HO-1
|
heme oxygenase-1
|
|
SOD
|
superoxide dismutase
|
|
GSS
|
glutathione synthetase
|
|
IL-1
|
interleukin-1
|
|
TNF-α
|
tumor necrosis factor-α
|
|
HMGB1
|
high mobility group box 1 protein
|
|
COX-2
|
cyclooxygenase-2
|
|
TLR4
|
Toll-like receptor 4
|
|
MyD88
|
myeloid differentiation factor 88
|
|
TRAF6
|
TNF receptor-associated factor 6
|
|
NF-κB
|
nuclear factor κ- light-chain-enhancer
of activated B cells
|
|
ERK
|
extracellular signal-related protein
kinase
|
|
JNK
|
c-Jun N-terminal kinase
|
|
MAPKs
|
mitogen activated protein kinases
|
|
AP-1
|
activator protein transcription
factor-1
|
|
Fas
|
fatty acid synthase
|
|
FasL
|
Fas ligand
|
|
Bcl-2
|
B-cell lymphoma 2
|
|
Bcl-xL
|
B-cell lymphoma-extra large
|
|
Bax
|
Bcl-2- associated X protein
|
|
Mito Cyto c
|
mitochondrial cytochrome c
|
|
Cyto Cyto c
|
cytoplasmic cytochrome c
|
|
LD 50
|
median lethal dose
|
Acknowledgments
We would like to thank all colleagues within the
Department of Pharmaceutical Analysis and correlative laboratory
technicians from the Science Research Center of Dalian Medical
University for their kind help and support.
References
|
1
|
Zhou WW, Lu S, Su YJ, Xue D, Yu XL, Wang
SW, Zhang H, Xu PX, Xie XX and Liu RT: Decreasing oxidative stress
and neuroinflammation with a multifunctional peptide rescues memory
deficits in mice with Alzheimer disease. Free Radic Biol Med.
74:50–63. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Frankola KA, Greig NH, Luo W and Tweedie
D: Targeting TNF-α to elucidate and ameliorate neuroinflammation in
neurodegenerative diseases. CNS Neurol Disord Drug Targets.
10:391–403. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Khalil WK, Assaf N, ElShebiney SA and
Salem NA: Neuroprotective effects of bee venom acupuncture therapy
against rotenone-induced oxidative stress and apoptosis. Neurochem
Int. 80:79–86. 2015. View Article : Google Scholar
|
|
4
|
Ansari N, Khodagholi F, Amini M and
Shaerzadeh F: Attenuation of LPS-induced apoptosis in
NGF-differentiated PC12 cells via NF-κB pathway and regulation of
cellular redox status by an oxazine derivative. Biochimie.
93:899–908. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
He AY, Qiu LJ, Gao Y, Zhu Y, Xu ZW, Xu JM
and Zhang ZH: The role of oxidative stress in neuromelanin
synthesis in PC12 cells. Neuroscience. 189:43–50. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Shaerzadeh F, Ansari N, Amini M and
Khodagholi F: Neuroprotective effects of oxazine derivative against
LPS-induced oxidative stress via attenuation of NF-κB pathway and
regulation of cellular redox status in PC12 cells. Alzheimers
Dement. 7:S616–S617. 2011. View Article : Google Scholar
|
|
7
|
Lee JW, Lee YK, Yuk DY, Choi DY, Ban SB,
Oh KW and Hong JT: Neuro-inflammation induced by lipopolysaccharide
causes cognitive impairment through enhancement of beta-amyloid
generation. J Neuroinflammation. 5:372008. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Song X, Guo M, Wang T, Wang W, Cao Y and
Zhang N: Geniposide inhibited lipopolysaccharide-induced apoptosis
by modulating TLR4 and apoptosis-related factors in mouse mammary
glands. Life Sci. 119:9–17. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Gadad BS, Britton GB and Rao KS: Targeting
oligomers in neurodegenerative disorders: lessons from α-synuclein,
tau, and amyloid-β peptide. J Alzheimers Dis. 24(Suppl 2): 223–232.
2011.
|
|
10
|
Nijveldt RJ, van Nood E, van Hoorn DE,
Boelens PG, van Norren K and van Leeuwen PA: Flavonoids: a review
of probable mechanisms of action and potential applications. Am J
Clin Nutr. 74:418–425. 2001.PubMed/NCBI
|
|
11
|
Maher P, Salgado KF, Zivin JA and Lapchak
PA: A novel approach to screening for new neuroprotective compounds
for the treatment of stroke. Brain Res. 1173:117–125. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Lv L, Zheng L, Dong D, Xu L, Yin L, Xu Y,
Qi Y, Han X and Peng J: Dioscin, a natural steroid saponin, induces
apoptosis and DNA damage through reactive oxygen species: a
potential new drug for treatment of glioblastoma multiforme. Food
Chem Toxicol. 59:657–669. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Zhang S and Peng J: Response to: 'hormetic
effect of Rosa laevigata Michx in CCl4-induced
hepatotoxicity and the presumptive role of PPARs'. Food Chem
Toxicol. 57:3892013. View Article : Google Scholar
|
|
14
|
Wei Y, Xu Y, Han X, Qi Y, Xu L, Xu Y, Yin
L, Sun H, Liu K and Peng J: Anti-cancer effects of dioscin on three
kinds of human lung cancer cell lines through inducing DNA damage
and activating mitochondrial signal pathway. Food Chem Toxicol.
59:118–128. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Yu J, Dandekar DV, Toledo RT, Singh RK and
Patil BS: Supercritical fluid extraction of limonoids and naringin
from grapefruit (Citrus paradisi Macf.) seeds. Food Chem.
105:1026–1031. 2007. View Article : Google Scholar
|
|
16
|
Choi MS, Do KM, Park YS, Jeon SM, Jeong
TS, Lee YK, Lee MK and Bok SH: Effect of naringin supplementation
on cholesterol metabolism and antioxidant status in rats fed high
cholesterol with different levels of vitamin E. Ann Nutr Metab.
45:193–201. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Raza SS, Khan MM, Ahmad A, Ashafaq M and
Islam F, Wagner AP, Safhi MM and Islam F: Neuroprotective effect of
naringenin is mediated through suppression of NF-κB signaling
pathway in experimental stroke. Neuroscience. 230:157–171. 2013.
View Article : Google Scholar
|
|
18
|
Sachdeva AK, Kuhad A and Chopra K:
Naringin ameliorates memory deficits in experimental paradigm of
Alzheimer's disease by attenuating mitochondrial dysfunction.
Pharmacol Biochem Behav. 127:101–110. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Ding G, Zhang Z, Chopp M, Li L, Zhang L,
Li Q, Wei M and Jiang Q: MRI evaluation of BBB disruption after
adjuvant AcSDKP treatment of stroke with tPA in rat. Neuroscience.
271:1–8. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zbarsky V, Datla KP, Parkar S, Rai DK,
Aruoma OI and Dexter DT: Neuroprotective properties of the natural
phenolic antioxidants curcumin and naringenin but not quercetin and
fisetin in a 6-OHDA model of Parkinson's disease. Free Radic Res.
39:1119–1125. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Rogers SW, Mandelzys A, Deneris ES, Cooper
E and Heinemann S: The expression of nicotinic acetylcholine
receptors by PC12 cells treated with NGF. J Neurosci. 12:4611–4623.
1992.PubMed/NCBI
|
|
22
|
Zhao X, Cong X, Zheng L, Xu L, Yin L and
Peng J: Dioscin, a natural steroid saponin, shows remarkable
protective effect against acetaminophen-induced liver damage in
vitro and in vivo. Toxicol Lett. 214:69–80. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Jia Y, Ji L, Zhang S, Xu L, Yin L, Li L,
Zhao Y and Peng J: Total flavonoids from Rosa Laevigata Michx fruit
attenuates hydrogen peroxide induced injury in human umbilical vein
endothelial cells. Food Chem Toxicol. 50:3133–3141. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Omata Y, Saito Y, Fujita K, Ogawa Y,
Nishio K, Yoshida Y and Niki E: Induction of adaptive response and
enhancement of PC12 cell tolerance by lipopolysaccharide primarily
through the upregulation of glutathione S-transferase A3 via Nrf2
activation. Free Radic Biol Med. 45:1437–1445. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Balachandran C, Sangeetha B, Duraipandiyan
V, Raj MK, Ignacimuthu S, Al-Dhabi NA, Balakrishna K, Parthasarathy
K, Arulmozhi NM and Arasu MV: A flavonoid isolated from
Streptomyces sp. (ERINLG-4) induces apoptosis in human lung cancer
A549 cells through P53 and cytochrome c release caspase dependant
pathway. Chem Biol Interact. 224:24–35. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Blatt NB, Boitano AE, Lyssiotis CA,
Opipari AW Jr and Glick GD: Bz-423 superoxide signals apoptosis via
selective activation of JNK, Bak, and Bax. Free Radic Biol Med.
45:1232–1242. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen YT, Shi D, Yang D and Yan B:
Antioxidant sulforaphane and sensitizer trinitrobenzene sulfonate
induce carboxylesterase-1 through a novel element transactivated by
nuclear factor-E2 related factor-2. Biochem Pharmacol. 84:864–871.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Valencia-Olvera AC, Morán J,
Camacho-Carranza R, Prospéro-García O and Espinosa-Aguirre JJ:
CYP2E1 induction leads to oxidative stress and cytotoxicity in
glutathione-depleted cerebellar granule neurons. Toxicol In Vitro.
28:1206–1214. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kumar A, Prakash A and Dogra S: Naringin
alleviates cognitive impairment, mitochondrial dysfunction and
oxidative stress induced by D-galactose in mice. Food Chem Toxicol.
48:626–632. 2010. View Article : Google Scholar
|
|
30
|
Golechha M, Chaudhry U, Bhatia J, Saluja D
and Arya DS: Naringin protects against kainic acid-induced status
epilepticus in rats: evidence for an antioxidant, anti-inflammatory
and neuroprotective intervention. Biol Pharm Bull. 34:360–365.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kandhare AD, Ghosh P and Bodhankar SL:
Naringin, a flavanone glycoside, promotes angiogenesis and inhibits
endothelial apoptosis through modulation of inflammatory and growth
factor expression in diabetic foot ulcer in rats. Chem Biol
Interact. 219:101–112. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dong D, Xu L, Yin L, Qi Y and Peng J:
Naringin prevents carbon tetrachloride-induced acute liver injury
in mice. J Funct Foods. 12:179–191. 2015. View Article : Google Scholar
|
|
33
|
Wang D, Gao K, Li X, Shen X, Zhang X, Ma
C, Qin C and Zhang L: Long-term naringin consumption reverses a
glucose uptake defect and improves cognitive deficits in a mouse
model of Alzheimer's disease. Pharmacol Biochem Behav. 102:13–20.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Golechha M, Sarangal V, Bhatia J, Chaudhry
U, Saluja D and Arya DS: Naringin ameliorates
pentylenetetrazol-induced seizures and associated oxidative stress,
inflammation, and cognitive impairment in rats: possible mechanisms
of neuroprotection. Epilepsy Behav. 41:98–102. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mani VM and Sadiq AMM: Naringin modulates
the impairment of memory, anxiety, locomotor, and emotionality
behaviors in rats exposed to deltamethrin; a possible mechanism
association with oxidative stress, acetylcholinesterase and ATPase.
Biomed Prev Nutr. 4:527–533. 2014. View Article : Google Scholar
|
|
36
|
Li P, Wang S, Guan X, Cen X, Hu C, Peng W,
Wang Y and Su W: Six months chronic toxicological evaluation of
naringin in Sprague-Dawley rats. Food Chem Toxicol. 66:65–75. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Liu S, Han Y, Zhang T and Yang Z:
Protective effect of trifluoperazine on hydrogen peroxide-induced
apoptosis in PC12 cells. Brain Res Bull. 84:183–188. 2011.
View Article : Google Scholar
|
|
38
|
Pérez MJ and Cederbaum AI: Antioxidant and
pro-oxidant effects of a manganese porphyrin complex against
CYP2E1-dependent toxicity. Free Radic Biol Med. 33:111–127. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Li X, Lian LH, Bai T, Wu YL, Wan Y, Xie
WX, Jin X and Nan JX: Cryptotanshinone inhibits LPS-induced
proinflammatory mediators via TLR4 and TAK1 signaling pathway. Int
Immunopharmacol. 11:1871–1876. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Cui L, Feng L, Zhang ZH and Jia XB: The
anti-inflammation effect of baicalin on experimental colitis
through inhibiting TLR4/NF-κB pathway activation. Int
Immunopharmacol. 23:294–303. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Lu SM, Yu CJ, Liu YH, Dong HQ, Zhang X,
Zhang SS, Hu LQ, Zhang F, Qian YN and Gui B: S100A8 contributes to
postoperative cognitive dysfunction in mice undergoing tibial
fracture surgery by activating the TLR4/MyD 88 pathway. Brain Behav
Immun. 44:221–234. 2015. View Article : Google Scholar
|
|
42
|
Nadeau S and Rivest S: Role of
microglial-derived tumor necrosis factor in mediating CD14
transcription and nuclear factor kappa B activity in the brain
during endotoxemia. J Neurosci. 20:3456–3468. 2000.PubMed/NCBI
|
|
43
|
Castello L, Froio T, Maina M, Cavallini G,
Biasi F, Leonarduzzi G, Donati A, Bergamini E, Poli G and
Chiarpotto E: Alternate-day fasting protects the rat heart against
age-induced inflammation and fibrosis by inhibiting oxidative
damage and NF-κB activation. Free Radic Biol Med. 48:47–54. 2010.
View Article : Google Scholar
|
|
44
|
Zheng Y, Guo Z, He W, Yang Y, Li Y, Zheng
A, Li P, Zhang Y, Ma J, Wen M, et al: Ephedrine hydrochloride
protects mice from LPS challenge by promoting IL-10 secretion and
inhibiting proinflammatory cytokines. Int Immunopharmacol.
13:46–53. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Mukhopadhyay P, Rajesh M, Horváth B,
Bátkai S, Park O, Tanchian G, Gao RY, Patel V, Wink DA, Liaudet L,
et al: Cannabidiol protects against hepatic ischemia/reperfusion
injury by attenuating inflammatory signaling and response,
oxidative/nitrative stress, and cell death. Free Radic Biol Med.
50:1368–1381. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Abdelazeem AH, Abdelatef SA, El-Saadi MT,
Omar HA, Khan SI, McCurdy CR and El-Moghazy SM: Novel
pyrazolopyrimidine derivatives targeting COXs and iNOS enzymes;
design, synthesis and biological evaluation as potential
anti-inflammatory agents. Eur J Pharm Sci. 62:197–211. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Zhang J, Takahashi HK, Liu K, Wake H, Liu
R, Maruo T, Date I, Yoshino T, Ohtsuka A, Mori S and Nishibori M:
Anti-high mobility group box-1 monoclonal antibody protects the
blood-brain barrier from ischemia-induced disruption in rats.
Stroke. 42:1420–1428. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Park JS, Gamboni-Robertson F, He Q,
Svetkauskaite D, Kim JY, Strassheim D, Sohn JW, Yamada S, Maruyama
I, Banerjee A, et al: High mobility group box 1 protein interacts
with multiple toll-like receptors. Am J Physiol Cell Physiol.
290:C917–C924. 2006. View Article : Google Scholar
|
|
49
|
Chipuk JE and Green DR: How do BCL-2
proteins induce mitochondrial outer membrane permeabilization?
Trends Cell Biol. 18:157–164. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Zeng KW, Liao LX, Zhao MB, Song FJ, Yu Q,
Jiang Y and Tu PF: Protosappanin B protects PC12 cells against
oxygen-glucose deprivation-induced neuronal death by maintaining
mitochondrial homeostasis via induction of ubiquitin-dependent P53
protein degradation. Eur J Pharmacol. 751:13–23. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Miller DK: The role of the Caspase family
of cysteine proteases in apoptosis. Semin Immunol. 9:35–49. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Islam Z, Amuzie CJ, Harkema JR and Pestka
JJ: Neurotoxicity and inflammation in the nasal airways of mice
exposed to the macrocyclic trichothecene mycotoxin roridin a:
kinetics and potentiation by bacterial lipopolysaccharide
coexposure. Toxicol Sci. 98:526–541. 2007. View Article : Google Scholar : PubMed/NCBI
|