Introduction
Amikacin (AMK) is one of the aminoglycoside
antibiotics most often used for the treatment of severe,
hospital-acquired infections due to multidrug resistant
Gram-negative bacteria (1). The
common side-effect of AMK is ototoxicity, which results in hearing
loss or deafness (2,3). Aminoglycosides induce the generation
of reactive oxygen species (ROS) and impair redox homeostasis in
the inner ear (4). It has
previously been demonstrated that AMK-induced ototoxicity is most
likely caused by the production and accumulation of ROS, which
damage the hair cells of the basilar membrane in cochlea (5).
Punica granatum L., often referred to as the
pomegranate, is an edible fruit that is widely cultivated in a
number of countries (6). The
antioxidant effects of pomegranate peel extract (PPE) have been
studied on the removal of ROS, and have been shown to inhibit lipid
peroxidation in vivo (7).
The antioxidant status of the cochlea has emerged as an important
factor in the death or survival of hair cells with respect to 3
primary contributors that induce hearing loss: noise trauma, drugs
(aminoglycoside antibiotics and cisplatin) and the aging process
(8–10). A previous study indicated that
oral PPE decreases the levels of ROS, as well as acute inflammation
in the tympanic membrane following myringotomy (11). Moreover, the oral administration
of PPE in rats has been shown to reduce cisplatin-induced toxicity
in the cochlea; therefore, an oral experimental dose of PPE may
have a powerful effect in protecting cisplatin-induced ototoxicity
in rats (12). However, the
molecular mechanisms underlying PPE-induced protection from
ototoxic drugs are not yet fully understood.
Forkhead box proteins O (FoxO) are a subclass of the
forkhead family of transcription factors, and have been linked to
the regulation of oxidative stress and cellular apoptosis (13). Mammalian FoxOs consist of 4
homologous genes: FoxO1, FoxO3a, FoxO4 and FoxO6, all of which
regulate various cellular functions, such as proliferation,
differentiation and longevity in diverse cell types (14). ROS induce FoxO3a phosphorylation
via the mitogen-activated protein kinase (MAPK) pathway, and
suppress the expression of antioxidant genes, including p21, p27
and Bim, ultimately inducing cell death (15). Activated JNK is responsible for
the nuclear translocation and transcriptional activity of FoxO3a
(16). More importantly, FoxO3-KO
mice display mild high-frequency hearing loss compared to wild-type
mice (17).
In this study, we evaluated the protective effects
of PPE on AMK-induced ototoxicity and the potential underlying
mechanisms in vivo. We hypothesized that PPE may attenuate
hearing loss and hair cell damage in response to AMK, and that
these protective effects may mediated via the regulation of
MAPK/FoxO3a signaling.
Materials and methods
Drugs and reagents
PPE was purchased from Xi'an Acetar Bio-Tech Inc.
(Shaanxi, China). PPE, a brown powder that contains ≥98% ellagic
acid (phenolic compound with antioxidant and anti-inflammatory
effects), was dissolved in saline for gavage. AMK sulfate
injections (0.1 g/ml) were purchased from Qilu Pharmaceutical Co.,
Ltd. (Shandong, China). Rabbit polyclonal anti-FoxO3a (ab109629),
anti-phospho-FoxO3a (ab47285) and anti-4-hydroxynonenal (4-HNE;
ab46545) primary antibodies were from Abcam (Cambridge, UK); rabbit
polyclonal anti-p38 (8690), anti-phospho-p38 (9215), anti-ERK 1/2
(4695), anti-phospho-ERK (4370), anti-JNK (9252), anti-p-JNK
(9251), anti-Bcl-2 (2876), anti-Bax (5023) and anti-cleaved
caspase-3 (9579) were from Cell Signaling Technology, Inc.
(Danvers, MA, USA).
Animals
All animal manipulations were conducted in
accordance with the regulations for the Management of Laboratory
Animals published by the Ministry of Science and Technology of the
People's Republic of China, and was approved by the Institutional
Animal Care and Use Committee of Jinzhou Medical University. BALB/c
mice (18–22 g), which were 6–8 weeks old and had normal
auropalpebral reflexes and otomicroscopic examination results, were
purchased from the Animal Experimental Center of Dalian Medical
University [Liaoning, China; license no. SCXK (Liao) 2008-0002].
All mice were fed a standard commercial diet, and housed at an
ambient temperature of 22°C with a relative humidity of 50±5% under
12 h/12 h light-dark cycle in a specific pathogen-free
facility.
Animal experiments
The experimental mice were divided into 4 groups
(n=20 ears in each group): i) the control group received
physiological saline (100 µl/day) via gavage; ii) the AMK
group intraperitoneally received AMK intramuscular injection at 500
mg/kg/day for 15 consecutive days, as previously described
(18); iii) the PPE plus AMK
group not only received hypodermic injection for AMK at 500
mg/kg/day for 15 consecutive days, but also PPE (34 mg/kg, 100
µl/day) via gavage for 5 days prior to AMK injection and for
15 days concomitantly with AMK injections; and iv) the PPE group
received PPE via gavage for 20 days.
Measurement of hearing function
For the analysis of the auditory threshold, the
auditory brainstem response (ABR) was recorded 1 day before and 15
days after AMK treatment with tone bursts of 8, 12, 24 and 32 kHz
(1-msec rise/fall time, 2-msec plateau) using the Smart EP and OAE
auditory evoked potential recording system (Intelligent Hearing
Systems Co., Miami, FL, USA). The mice were anesthetized using
pentobarbital sodium (40 mg/kg) and kept warm with a heating pad
during ABR recording. A subdermal (active) needle electrode was
inserted at the vertex, while ground and reference electrodes were
inserted subdermally in the loose skin beneath the pinnae of
opposite ears. The technique used to record ABRs has been
previously described in detail (19). The ABR waveforms were averaged
over a 10-msec time window using the Smart EP and OAE auditory
evoked potential recording system software. The sound intensity was
varied at 5 dB intervals near the hearing threshold. The
differences in ABR thresholds shift for each frequency between the
starting and the terminal points of the experimental time course
were noted. The threshold was determined off-line by two
independent, experimentally blinded observers on the basis of the
ABR records. Briefly, thresholds were recorded using the Smart EP
and OAE auditory evoked potential recording system. In addition,
the mice were euthanized and double cochleas were removed for
further analysis.
Sensory hair cell counting
After the ABR test, the temporal bones were
harvested. Each bulla was opened using rongeurs to expose the
cochlea. The oval and round windows were then opened. Following the
creation of a hole in the cochlea apex, 4% paraformaldehyde was
perfused through the cochlea for at least 24 h. The cochlea was
decalcified in 4% EDTA for 7 days at 4°C. Subsequently, the basilar
membrane was dissected under a dissecting microscope, and the stria
vascularis and tectorial membrane were removed. To identify F-actin
in the organ of Corti, tetramethyl rhodamine isothiocyanate (TRITC)
(Sigma-Aldrich) was applied for 20 min at room temperature and
protected from light. The specimens were then rinsed 3 times with
0.01 M phosphate-buffered saline (PBS) (pH 7.4). Fluorescence
signals from the hair cells were counted under a BX41 microscope
with epifluorescence (Olympus, Tokyo, Japan), and the images were
obtained with TCS-SP5II laser-scanning confocal microscope (Leica
Biosystems, Wetzlar, Germany). Three rows of the outer hair cells
(OHCs) were counted from the apex through the basilar turn of the
cochlea under ×200 magnification in 20 consecutive fields.
Immunohistofluorescence and
immunohistochemical analysis
Tissue sections were incubated with rabbit
anti-4-HNE antibody (1:200 dilution; Abcam), followed by
appropriate fluorescent-secondary antibody for 1 h at room
temperature. After the samples were counterstained with DAPI,
immunohistofluorescence images were obtained via confocal
microscopy (TCS-SP5II; Leica Biosystems). Immunofluorescence
analysis was performed as previously described (19). After preparation and blocking, the
sections were incubated with rabbit anti-phospho-FoxO3a antibody
(1:200 dilution; Abcam) overnight at 4°C, and rinsed with TBS plus
Tween-20 (TBST). The sections were incubated with biotinylated goat
anti-rabbit Boost Detection reagent (Cell Signaling Technology,
Inc.) for 30 min at room temperature, and rinsed with TBST. The
chromogen reaction was performed with DAB (Cell Signaling
Technology, Inc.) at room temperature, and rinsed with distilled
water to terminate the reaction. The sections were observed by
using Nikon microscope digital camera software (Nikon, Tokyo,
Japan).
Measurement of malondialdehyde (MDA),
catalase (CAT) and superoxide dismutase (SOD) activities
Cochlea tissue was homogenized in 0.01 M cold
phosphate buffer (pH 7.4) using a homogenizer. The homogenate was
centrifuged at 1,500 × g for 10 min. Tissue supernatant was
collected and measured for oxidative stress using the MDA assay kit
[2-thio-barbituric acid (TBA) method]. SOD and CAT activities were
assessed using the SOD and CAT assay kit (Nanjing Jiancheng
Bioengineering Institute, Nanjing, China), according to the
manufacturer's instructions.
Western blot analysis
Total cochlea tissue was homogenized in RIPA buffer
(Aidlab Biotechnologies Co., Ltd, Beijing, China) containing
protease inhibitor cocktail tablets and phosphostop cocktails
(cOmplete; Roche Diagnostics GmbH, Mannheim, Germany). The tissue
homogenate was sonicated for 30 sec and centrifuged at 12,000 rpm
at 4°C for 30 min to extract the supernatant. Protein
concentrations were determined using the BCA kit (Thermo Fisher
Scientific, Inc., San Francisco, CA, USA). The protein samples (30
µg each lane) were separated using SDS-PAGE. Following
electrophoresis, the proteins were transferred onto nitrocellulose
membranes (Millipore Corp., Boston, MA, USA). The membranes were
then blocked in 2% BSA in TBS for 1 h, and probed with primary
antibodies rabbit anti-FoxO3a, anti-phospho-FoxO3a (1:300 dilution;
Abcam), anti-p38, phospho-p38, ERK 1/2, phospho-ERK, JNK,
phospho-JNK, Bcl-2, Bax and cleaved caspase-3 (1:500 dilution; Cell
Signaling Technology, Inc.) overnight at 4°C, and then washed 3
times with TBST. Subsequently, the membranes were incubated with
their respective secondary antibodies (1:15,000, IRDye antibodies;
LI-COR Biotechnology, Lincoln, NE, USA) for 1 hour at RT. Membranes
were scanned using the Odyssey infrared imaging system (LI-COR
Biotechnology). The value of the relative density of each target
protein band was normalized to the density of the corresponding
β-actin band.
Statistical analysis
Data were analyzed using Prism 5 software (GraphPad
Software, Inc., La Jolla, CA, USA), and presented as mean ± SD.
Comparisons of parameters between different groups were performed
by one-way analysis of variance (ANOVA), followed by a Newman-Keuls
test. A value of P<0.05 was considered to indicate a
statistically significant difference.
Results
PPE prevents AMK-induced hearing loss and
hair cell loss in mice
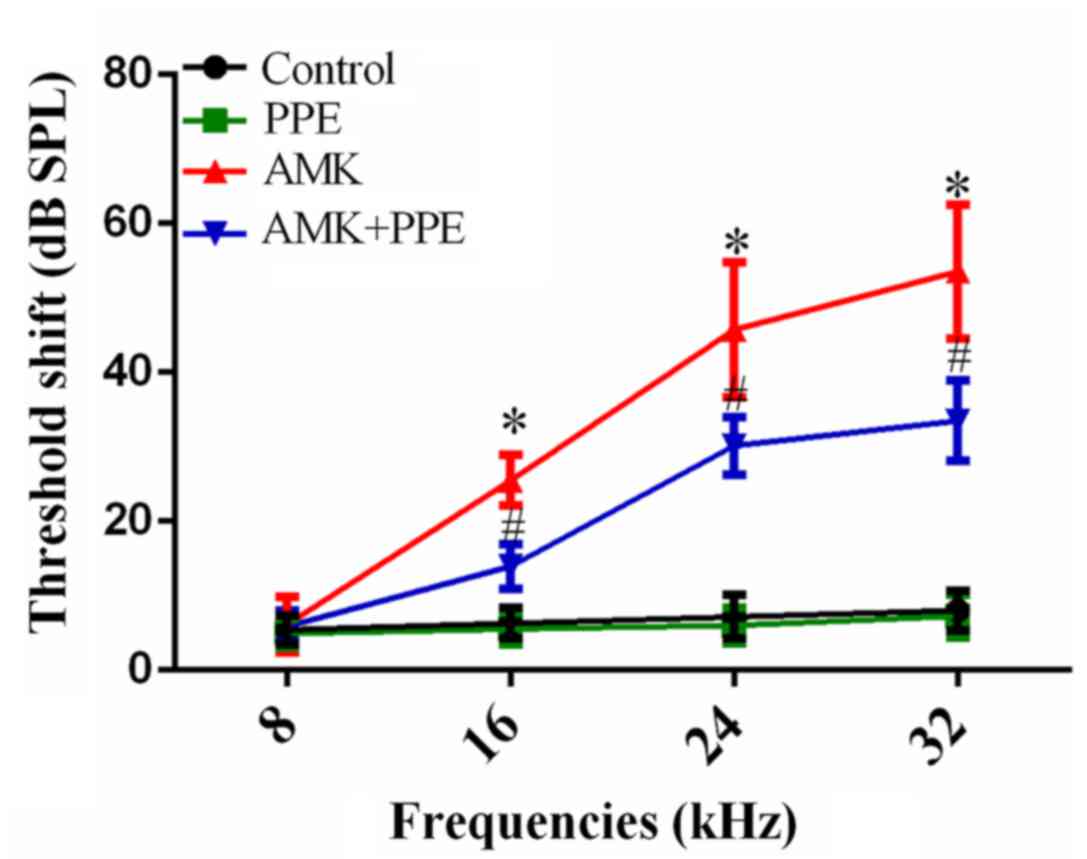
To verify whether PPE prevents hearing loss induced
by AMK, the ABR of the experimental groups (n=20 ears for each
group, two from each animal) was recorded. The ABR test showed that
mice in the control group maintained stable thresholds throughout
the experiment. By contrast, ABR threshold shifts in the AMK group
were significantly elevated at 16, 24 and 32 kHz (25.50±3.40,
45.75±9.07 and 53.55±8.97 dB, respectively) compared with the
control group (6.30±2.08, 7.15±2.94 and 8.00±2.73 dB, respectively)
after continuous injections for 15 days, and the hearing functional
deficit was greater at the higher frequency. However, following
concurrent treatment with PPE and AMK for 15 days, the ABR
threshold shifts were markedly reduced compared to the AMK group at
16, 24 and 32 kHz (13.95±2.98, 30.15±3.85 and 33.50±5.40 dB). PPE
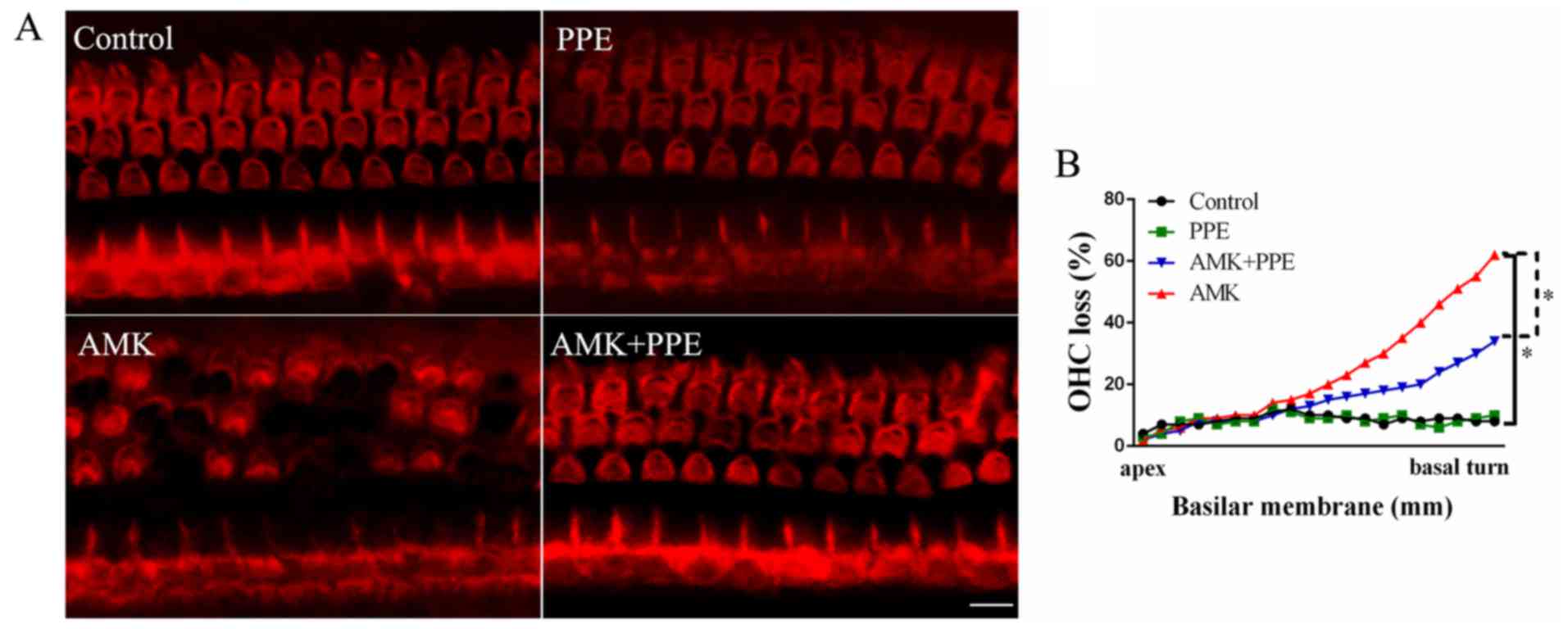
alone had no effect on the ABR threshold shifts (Fig. 1; P<0.05). To further determine
the in vivo protective effects of PPE against AMK, we
performed TRITC staining of stereociliary bundles of hair cells and
sensory hair cell counting in the 4 experimental groups. The
results revealed that in the control group or PPE group alone, the
morphology of the OHCs appeared normal in the lower basal turn of
the cochlea. In the AMK group, OHCs had more apparent morphological
changes or disorganization of stereocilia than the control group,
whereas many of the OHCs in the AMK plus PPE group survived AMK
insult (Fig. 2A). In addition,
quantitative hair cell counting revealed that the amount of OHC
loss was approximately 28% compared to that of approximately 58% in
the AMK group without PPE (Fig.
2B; P<0.05).
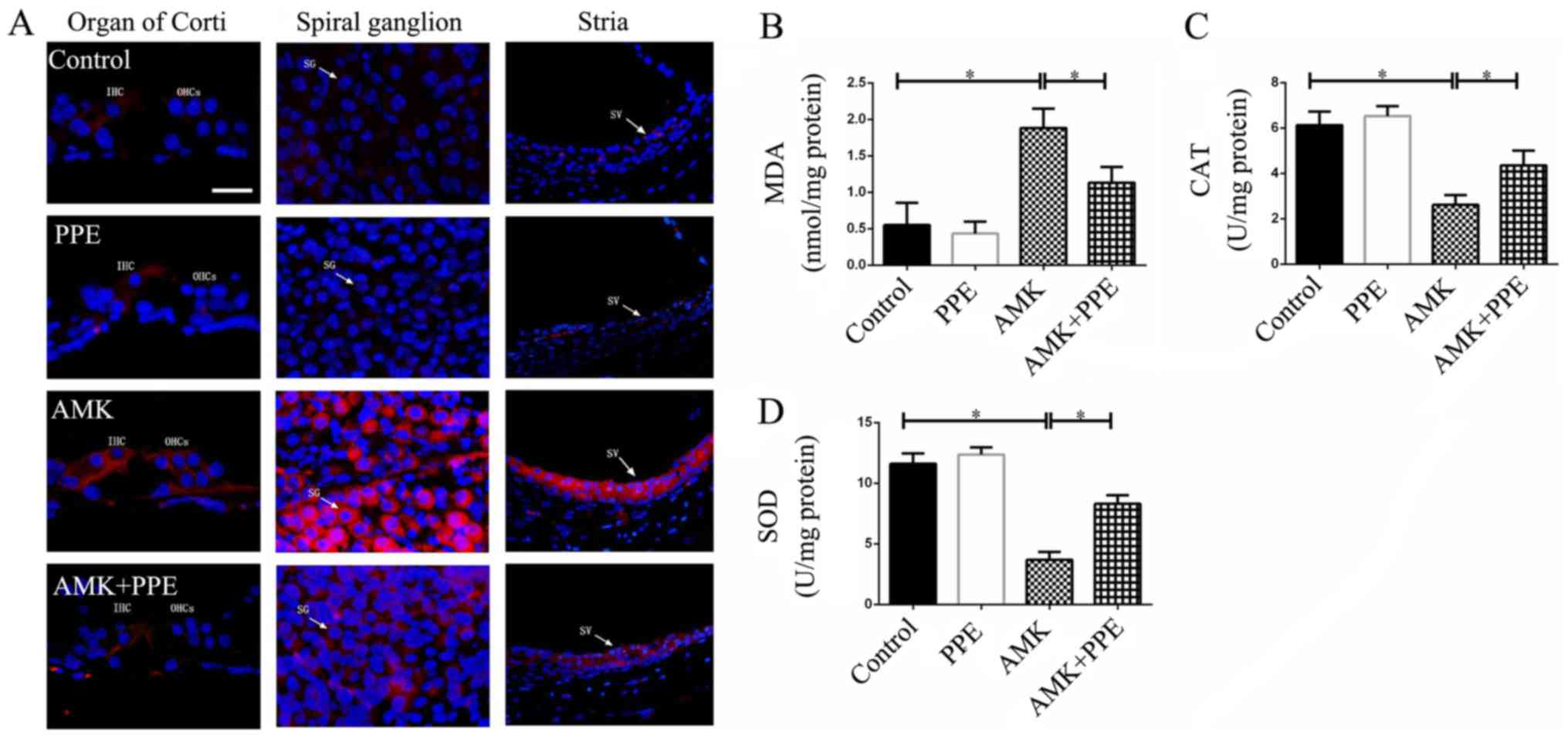
PPE alleviates oxidative stress in mice
with AMK-induced ototoxicity
AMK signifincantly increased the staining for 4-HNE
in the organ of Corti, spiral ganglion and stria vascularis of the
mouse cochleae. However, the production of 4-HNE was inhibited in
the mice treated with AMK plus PPE. However, PPE alone had no
effect on the expression of 4-HNE in the mouse cochleae, as
indicated by immunohistochemical analysis (Fig. 3A). We also observed that the
AMK-treated mice had lower protein levels of CAT and SOD (the
potent superoxide scavenging enzymes), and higher protein levels of
MDA (a marker for oxidative stress) compared with the control mice.
However, PPE administration prevented the increase in oxidative
stress-related markers in the cochleae of mice treated with AMK
(Fig. 3B–D; P<0.05).
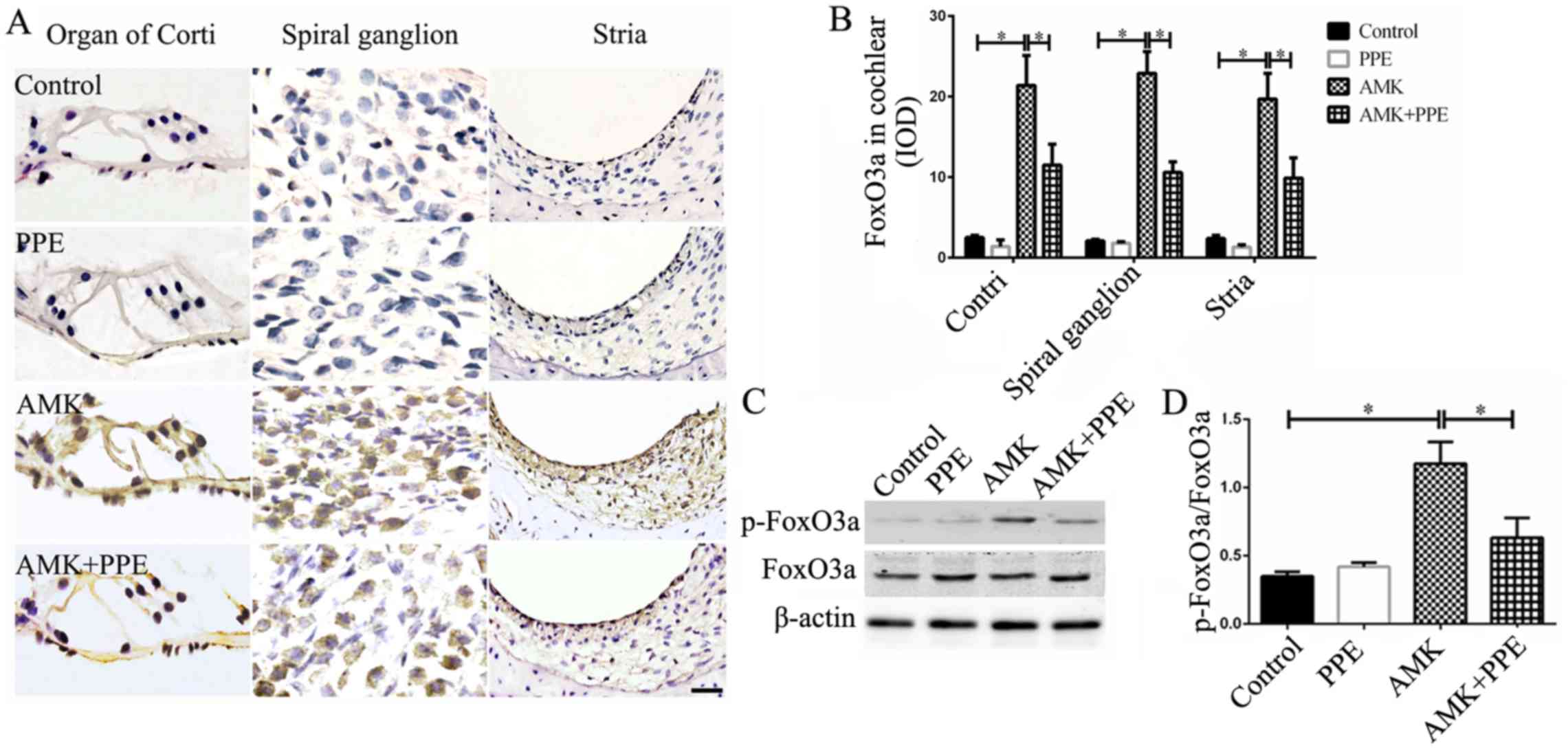
PPE inhibits the AMK-induced
phosphorylation of FoxO3a in mouse cochleae
Immunofluorescence staining revealed a significantly
increased phosphorylation of FoxO3a in the organ of Corti, spiral
ganglion and stria vascularis of the cochleae of mice treated with
AMK, which was prevented to a significant degree in mice treated
with AMK plus PPE (Fig. 4A and
B). Similarly, the results of western blot analysis also
revealed that the phosphorylation of FoxO3a in the cochleae of mice
treated with AMK was increased; however PPE treatment inhibited the
AMK induced the phosphorylation of FoxO3 proteins (Fig. 4C and D; P<0.05, n=3).
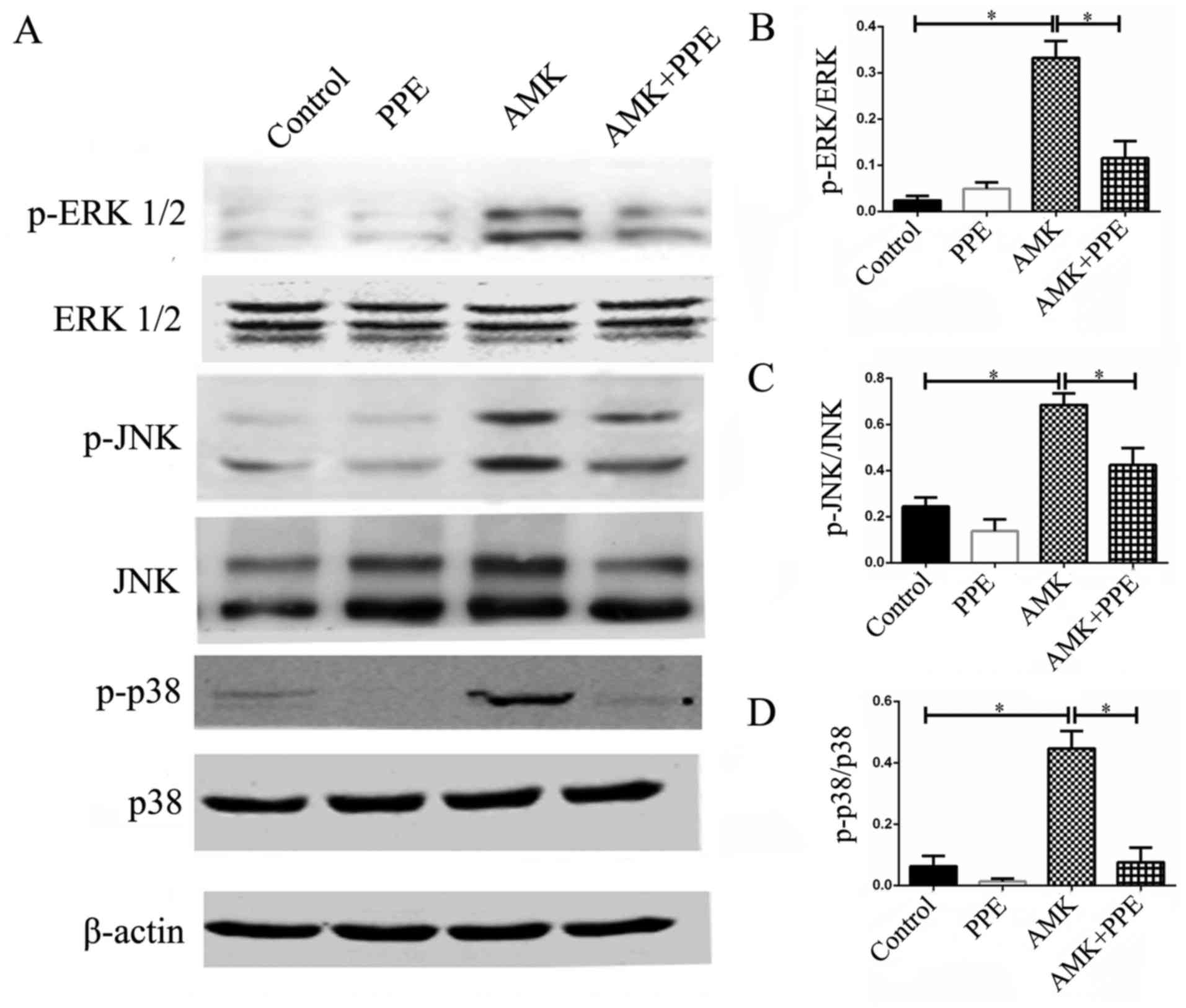
PPE inhibits the activation of MAPK in
cochleae of mice treated with AMK
The results of western blot analysis revealed the
activated phosphorylation of ERK 1/2 (Thr202/Tyr204), phospho-JNK
(Thr183/Tyr185) and phospho-p38 MAPK (Thr180/Tyr182), and the
upregulated protein expression of JNK in the cochleae of mice
treated with AMK. Of note, all these effects were inhibited in the
mice treated with AMK plus PPE (Fig.
5; P<0.05, n=3).
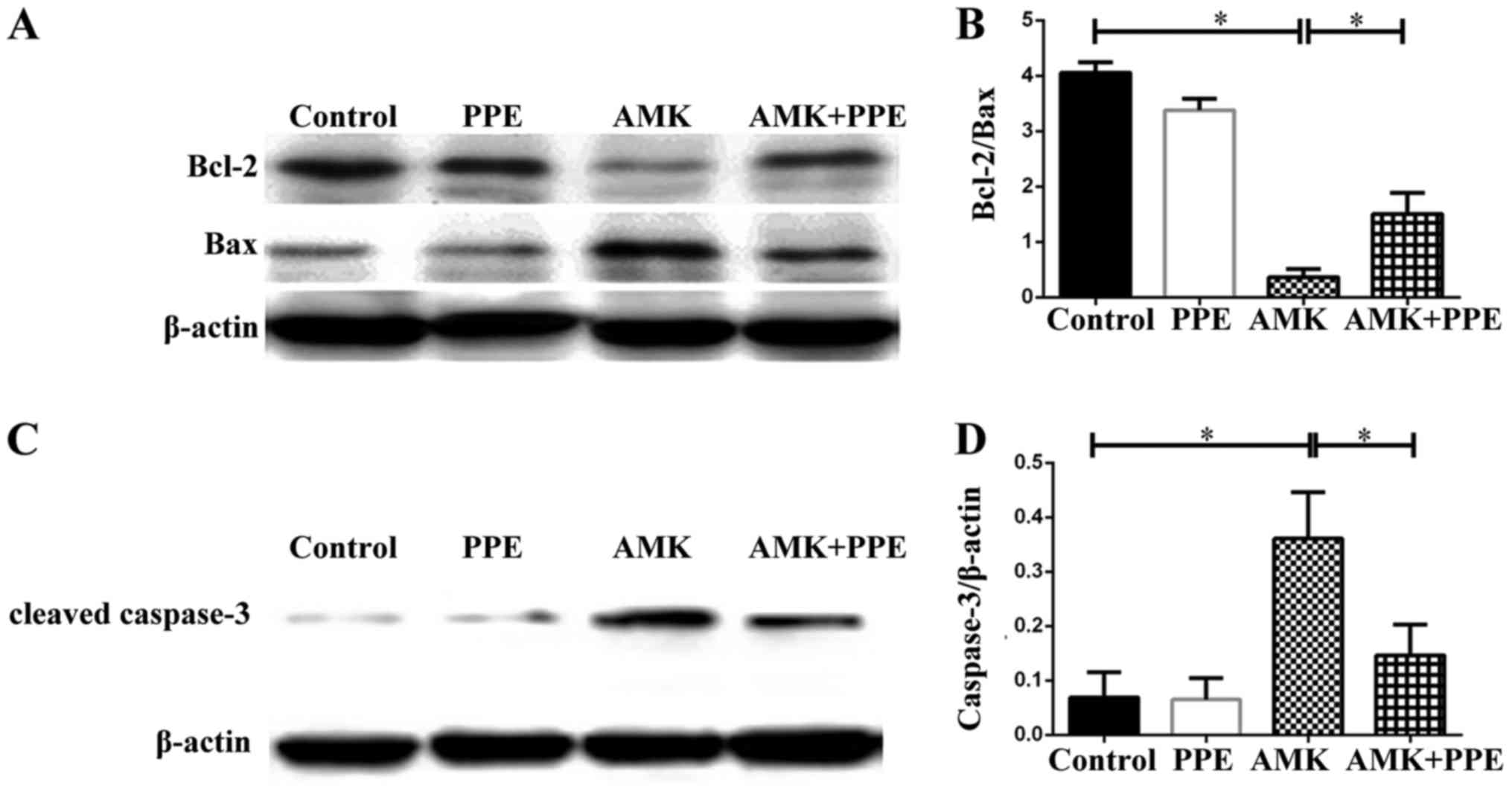
PPE attenuates the expression of
apoptotic family proteins in the cochleae of mice treated with
AMK
The results of western blot analysis revealed that
the protein expression of Bcl-2 was degraded, while the expression
of Bax and cleaved caspase-3 was upregulated in the cochleae of
mice in the AMK group. However, PPE inhibited the degradation of
Bcl-2 and prevented the activation of caspase-3 in the cochleae of
mice treated with AMK plus PPE (Fig.
6; P<0.05, n=3).
Discussion
Hair cell loss and the activation of oxidative
stress may play crucial roles in the development and progression of
aminoglycoside-induced ototoxicity (20). In this study, we confirmed that
PPE, a crude food antioxidant, exerted protective effects against
AMK-induced oxidative stress and hearing loss. In addition, the
expression of phosphorylated FoxO3a was distinctly visible in the
organ of Corti, spiral ganglion and stria vascularis of the
cochleae of mice with AMK-induced ototoxicity. Finally, we
demonstrated that PPE inhibited AMK-induced hair cell apoptosis in
the cochlea via the regulation of MAPK/FoxO3a signaling.
AMK has been shown to significantly enhance the ABR
threshold shifts in guinea pigs (21). Similarly, in this study, we found
that AMK-induced hearing injury in mice followed by the common
characteristics of aminoglycoside ototoxicity, and more severe
damage was caused at a higher frequency. A clinical study reported
that the hearing thresholds of 53 hospitalized patients
administered aminoglycosides were monitored at serial conventional
(0.25–8 kHz) and high-frequency (9–20 kHz) sound stimuli. The
results revealed that hearing loss occurred in 47% of the ears
studied, with hearing loss first appearing in the high-frequency
range in 71% of ears showing change (22). Another clinical study also showed
that uremic patients with continuous ambulatory peritoneal dialysis
(CAPD) peritonitis who received AMK treatment, had hearing loss,
particularly at higher frequencies (2). In addition, in our study, we found
hair cell deficiency from AMK ototoxicity in the lower basal turn
of cochlear cells. These results also confirmed that high frequency
hearing loss was mainly affected by hair cell loss at the base of
the cochlea. We also found that following concurrent administration
with AMK plus PPE, the ABR threshold shifts significantly decreased
at 16, 24 and 32 kHz compared with AMK, demonstrating that PPE
effectively protected the mice from AMK-induced hearing injury. In
addition, PPE alone had no effect on ABR threshold shifts in mice,
indicating that PPE is safe and non-toxic; thus, PPE may
effectively protect from aminoglycoside-induced ototoxicity.
Oxidative stress plays an important role in the
pathogenesis of drug-induced ototoxicity. 4-HNE, an aldehydic
product of lipid peroxidation, has been implicated in the etiology
of pathological changes under oxidative stress as a key mediator of
oxidative stress induced cell death (23). We observed that AMK augmented
4-HNE immunostaining in the cochlea. Furthermore, we assessed MDA,
a product of lipid peroxidation, as well as two antioxidant
enzymes, SOD and CAT, which are considered as biomarkers of
oxidative damage (24). Our
results indicated that AMK-induced ototoxicity was closely
associated with elevated MDA levels, and decreased SOD and CAT
levels in cochlea. However, PPE treatment reversed the expression
of AMK-induced these enzymes in the cochlea. PPE reduced ROS
production to block the oxidative stress, and acted as a potential
guardian against aminoglycoside ototoxicity. These results
suggested that the antioxidant PPE attenuated oxidative stress,
which may have beneficial therapeutic effects in the course of
AMK-induced ototoxicity.
FoxO3a has recently been widely investigated as a
transcription factor that is involved in regulation of the stress
response, apoptosis and autophagy (25). The efficiency of FoxO3a is
primarily regulated by phosphorylation. Several kinases, such as
protein kinase B (Akt) and MAPK, directly phosphorylate FoxO3a.
Phosphorylated FoxO3a combines with the 14-3-3 nuclear export
protein and remains in the cytoplasm in cells (26). FoxO3a also plays a key role in ROS
production (27) and regulation
of FoxO3a by MAPK signaling in response to oxidative stress
(28). In addition, FoxO3a
mediates activation of antioxidant genes, such as SOD and CAT
(29). A recent study showed that
saponins activated FoxO3a, and nuclear factor-erythroid 2-related
factor 2 (Nrf2) increased the expression and function of multiple
antioxidants, at least partly contributing to the protection of
D-galactose-induced aging in rats (30). In this study, we observed that AMK
ototoxicity enhanced the phosphorylation of FoxO3a, and inhibited
the nuclear translocation of FoxO3a in the cochlea. Our results
also demonstrated that PPE inhibited the high expression of
phosphorylated FoxO3a induced by AMK, which may exert beneficial
effects in AMK-induced auditory insults. It has been previously
reported that MAPK pathway is activated by aminoglycosides induced
in rodent models, and his pathway is at least partially responsible
for hair cell apoptosis (31).
This may be one of the reasons that antioxidant and free radical
scavengers prevent hair cell damage via MAPK dependent pathway on
cochlea (32). Our group
previously showed that α-lipoic acid effectively attenuated
kanamycin-induced hair cell toxicity, and was involved in MAPK
activation and apoptosis in mouse hair cells (19). The present study suggested that
PPE attenuates oxidative stress and ototoxicity by regulating
MAPK/FoxO3a signaling, and the contribution of FoxO3a to the
molecular mechanisms associated with AMK-induced hearing loss is a
key area for future studies.
Recent studies have shown that the overexpression of
Bcl-xL prevented gentamicin-induced hair cell apoptosis on the
cochlea in mice (33,34). In this study, we demonstrated that
PPE elevated the ratio of Bcl-2/Bax iin mice cochleae, and
inhibition of cleaved caspase-3 activation led to hair cell
apoptosis when treated with AMK. These results suggest that PPE may
contribute to alleviated hair cell apoptosis induced by AMK.
In conclusion, our in vivo data demonstrated
that PPE prevented AMK-induced ototoxicity by regulating the
MAPK/FoxO3a signaling pathway in the cochlea. Targeting FoxO3a in
the cochlea may thus prove be an effective therapeutic option in
drug-induced ototoxicity.
Acknowledgments
This study was funded by the Jinzhou Medical
University Youth Science and Technology Staring Foundation Program
(Y2012Z018), Liaoning Science and Technology Program (2014022029),
Liaoning Education Program (L2015316), Undergraduate of Liaoning
Province Innovation and Entrepreneurship Training Program
(201410160023).
References
|
1
|
Michalopoulos A and Papadakis E: Inhaled
anti-infective agents: emphasis on colistin. Infection. 38:81–88.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Tokgoz B, Ucar C, Kocyigit I, Somdas M,
Unal A, Vural A, Sipahioglu M, Oymak O and Utas C: Protective
effect of N-acetylcysteine from drug-induced ototoxicity in uraemic
patients with CAPD peritonitis. Nephrol Dial Transplant.
26:4073–4078. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Aksoy F, Dogan R, Ozturan O, Eren SB,
Veyseller B, Pektas A and Hüseyinbas Ö: Protective effect of
trimetazidine on amikacin-induced ototoxicity in rats. Int J
Pediatr Otorhinolaryngol. 78:663–669. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Forge A and Schacht J: Aminoglycoside
antibiotics. Audiol Neurootol. 5:3–22. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Szczepanik W, Kaczmarek P and
Jezowska-Bojczuk M: Oxidative activity of copper(II) complexes with
aminoglycoside antibiotics as implication to the toxicity of these
drugs. Bioinorg Chem Appl. 2:55–68. 2004. View Article : Google Scholar
|
|
6
|
Faria A and Calhau C: The bioactivity of
pomegranate: impact on health and disease. Crit Rev Food Sci Nutr.
51:626–634. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Al-Gubory KH, Blachier F, Faure P and
Garrel C: Pomegranate peel extract decreases small intestine lipid
peroxidation by enhancing activities of major antioxidant enzymes.
J Sci Food Agric. 96:3462–3468. 2016. View Article : Google Scholar
|
|
8
|
Yamashita D, Jiang HY, Schacht J and
Miller JM: Delayed production of free radicals following noise
exposure. Brain Res. 1019:201–209. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Jiang H, Sha SH and Schacht J: NF-kappaB
pathway protects cochlear hair cells from aminoglycoside-induced
ototoxicity. J Neurosci Res. 79:644–651. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Jiang H, Talaska AE, Schacht J and Sha SH:
Oxidative imbalance in the aging inner ear. Neurobiol Aging.
28:1605–1612. 2007. View Article : Google Scholar
|
|
11
|
Kahya V, Meric A, Yazici M, Yuksel M, Midi
A and Gedikli O: Antioxidant effect of pomegranate extract in
reducing acute inflammation due to myringotomy. J Laryngol Otol.
125:370–375. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Yazici ZM, Meric A, Midi A, Arınc YV,
Kahya V and Hafız G: Reduction of cisplatin ototoxicity in rats by
oral administration of pomegranate extract. Eur Arch
Otorhinolaryngol. 269:45–52. 2012. View Article : Google Scholar
|
|
13
|
Webb AE and Brunet A: FOXO transcription
factors: key regulators of cellular quality control. Trends Biochem
Sci. 39:159–169. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Almeida M: Unraveling the role of FoxOs in
bone - insights from mouse models. Bone. 49:319–327. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Parody JP, Ceballos MP, Quiroga AD,
Frances DE, Carnovale CE, Pisani GB, Alvarez ML and Carrillo MC:
FoxO3a modulation and promotion of apoptosis by interferon-α2b in
rat preneoplastic liver. Liver Int. 34:1566–1577. 2014. View Article : Google Scholar
|
|
16
|
Shukla S, Rizvi F, Raisuddin S and Kakkar
P: FoxO proteins' nuclear retention and BH3-only protein Bim
induction evoke mitochondrial dysfunction-mediated apoptosis in
berberine-treated HepG2 cells. Free Radic Biol Med. 76:185–199.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Gilels F, Paquette ST, Zhang J, Rahman I
and White PM: Mutation of Foxo3 causes adult onset auditory
neuropathy and alters cochlear synapse architecture in mice. J
Neurosci. 33:18409–18424. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Murillo-Cuesta S, Contreras J, Cediel R
and Varela-Nieto I: Comparison of different aminoglycoside
antibiotic treatments to refine ototoxicity studies in adult mice.
Lab Anim. 44:124–131. 2010. View Article : Google Scholar
|
|
19
|
Wang A, Hou N, Bao D, Liu S and Xu T:
Mechanism of alpha-lipoic acid in attenuating kanamycin-induced
ototoxicity. Neural Regen Res. 7:2793–2800. 2012.PubMed/NCBI
|
|
20
|
Shulman E, Belakhov V, Wei G, Kendall A,
Meyron-Holtz EG, Ben-Shachar D, Schacht J and Baasov T: Designer
amino-glycosides that selectively inhibit cytoplasmic rather than
mitochondrial ribosomes show decreased ototoxicity: a strategy for
the treatment of genetic diseases. J Biol Chem. 289:2318–2330.
2014. View Article : Google Scholar
|
|
21
|
Amora LdeA, Murashima AdeA, Rossato M,
Moreira MB, Hyppolito MÂ and Fagundes DJ: The effects of hyperbaric
oxygen therapy upon ototoxic injuries produced by amikacin in
guinea pigs. Braz J Otorhinolaryngol. 79:342–348. 2013. View Article : Google Scholar
|
|
22
|
Fausti SA, Henry JA, Schaffer HI, Olson
DJ, Frey RH and McDonald WJ: High-frequency audiometric monitoring
for early detection of aminoglycoside ototoxicity. J Infect Dis.
165:1026–1032. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Choi CH, Chen K, Du X, Floyd RA and Kopke
RD: Effects of delayed and extended antioxidant treatment on acute
acoustic trauma. Free Radic Res. 45:1162–1172. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Khalil SR, Abd-Elhakim YM, Selim ME and
Al-Ayadhi LY: Apitoxin protects rat pups brain from propionic
acid-induced oxidative stress: the expression pattern of Bcl-2 and
caspase-3 apoptotic genes. Neurotoxicology. 49:121–131. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Storz P: Forkhead homeobox type O
transcription factors in the responses to oxidative stress.
Antioxid Redox Signal. 14:593–605. 2011. View Article : Google Scholar :
|
|
26
|
Vogt PK, Jiang H and Aoki M: Triple layer
control: phosphorylation, acetylation and ubiquitination of FOXO
proteins. Cell Cycle. 4:908–913. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Zhang L, Yang X, Li X, Li C, Zhao L, Zhou
Y and Hou H: Butein sensitizes HeLa cells to cisplatin through the
AKT and ERK/p38 MAPK pathways by targeting FoxO3a. Int J Mol Med.
36:957–966. 2015.PubMed/NCBI
|
|
28
|
Sun L, Zang WJ, Wang H, Zhao M, Yu XJ, He
X, Miao Y and Zhou J: Acetylcholine promotes ROS detoxification
against hypoxia/reoxygenation-induced oxidative stress through
FoxO3a/PGC-1α dependent superoxide dismutase. Cell Physiol Biochem.
34:1614–1625. 2014. View Article : Google Scholar
|
|
29
|
Rangarajan P, Karthikeyan A, Lu J, Ling EA
and Dheen ST: Sirtuin 3 regulates Foxo3a-mediated antioxidant
pathway in microglia. Neuroscience. 311:398–414. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Li YN, Guo Y, Xi MM, Yang P, Zhou XY, Yin
S, Hai CX, Li JG and Qin XJ: Saponins from Aralia taibaiensis
attenuate D-galactose-induced aging in rats by activating FOXO3a
and Nrf2 pathways. Oxid Med Cell Longev. 2014:3205132014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yang J, Wu Z, Renier N, Simon DJ, Uryu K,
Park DS, Greer PA, Tournier C, Davis RJ and Tessier-Lavigne M:
Pathological axonal death through a MAPK cascade that triggers a
local energy deficit. Cell. 160:161–176. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Dinh CT, Goncalves S, Bas E, Van De Water
TR and Zine A: Molecular regulation of auditory hair cell death and
approaches to protect sensory receptor cells and/or stimulate
repair following acoustic trauma. Front Cell Neurosci. 9:962015.
View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Dong Y, Liu D, Hu Y and Ma X: NaHS
protects cochlear hair cells from gentamicin-induced ototoxicity by
inhibiting the mitochondrial apoptosis pathway. PLoS One.
10:e01360512015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chi NU, Tabuchi K, Nakamagoe M, Nakayama
M, Nishimura B and Hara A: Ceramide/sphingomyelin cycle involvement
in gentamicin-induced cochlear hair cell death. Arch Toxicol.
89:415–421. 2015. View Article : Google Scholar
|