Introduction
Melanin determines the color of human skin, hair and
eyes as a final product of melanogenesis (1,2),
and plays a protective role against sun exposure-associated
ultraviolet radiation (3). This
pigment is synthesized by a complex pathway in highly specialized
cells of the epidermis known as melanocytes (4). Melanocytes are normally positioned
at the basal layer of the epidermal part of the skin (5). Specifically, melanin is produced in
cellular sub-compartments denominated as melanosomes and is
transported to other cells of the epidermis, including
keratinocytes and Langerhans cells (5). Importantly, the overproduction or
abnormal distribution of melanin can lead to skin disorders and
cosmetic complications, such as melisma and freckles (5). Melanin is synthesized by the three
enzymes, tyrosinase, tyrosinase-related protein (TRP)-1 and
DOPAchrome tautomerase (DCT) also known as TRP-2 (6). UV radiation-sensitized human
epidermal keratinocytes produce factors which stimulate
melanogenesis, including α-melanocyte-stimulating hormone (α-MSH)
(7). Secreted α-MSH/ACTH binds to
the melanocortin 1 receptor (MC1R) on melanocytes to activate the
cAMP/PKA/CREB pathway, which subsequently increases the expression
levels of tyrosinase, TRP-1 and TRP-2 (7,8).
Tyrosinase converts L-tyrosine into L-3,4-dihydroxyphenylalanine
(L-DOPA); L-DOPA can then be converted into DOPAquinone, which
spontaneously undergoes cyclization and further rearrangement,
yielding l-DOPAchrome (9).
DCT/TRP-2 catalyzes the conversion of DOPAchrome to
5,6-dihydroxyindole-2-carboxylic acid (DHICA). TRP-1 is important
for tyrosinase stabilization and trafficking, thus ultimately
facilitating tyrosinase activity (10). Two types of melanin are produced
in mammalian cells, the reddish-yellow-colored pheomelanin and the
dark-brown-colored eumelanin. Tyrosinase is required for the
synthesis of pheomelanin and eumelanin, whereas TRP-1 and TRP-2 are
required for the synthesis of only eumelanin (11,12). Therefore, the downregulation of
tyrosinase and related protein expression and/or activity have been
proposed to be involved in the reduction of melanin production
(13).
Soyasaponins are bioactive secondary plant
metabolites in soybeans (Glycine max) and other legumes,
such as lentils (Lens culinaris) and green peas (Pisum
sativum L. cv. Azad) (14–16). Soyasaponins are oleanene-type
triterpenoid saponins with polar sugar chains and a non-polar
pentacyclic ring structure (17).
These metabolites are generally divided into four main groups on
the basis of their aglycone (soyasapogenol) structure, referred to
as groups A, B, E and DDMP
(2,3-dihydro-2,5-dihydroxy-6-methyl-4-pyrone) (14). Group A soyasaponins contain
bidesmosides with two sugar chains at the C-3 and C-22 position of
soyasapogenol A. Previously, eight isomers of group A saponins,
named Aa, Ab, Ac, Ad, Ae, Af, Ag and Ah, according to their elution
order in reversed-phase high-performance liquid chromatography
(HPLC), were isolated from soybeans and characterized in the study
by Shiraiwa et al (15).
Soyasaponins have been shown to have a number of
biological functions, including anti-obesity, anti-inflammatory and
anti-carcinogenic effects, as well as hepatoprotective effects
(14,18,19). However, to the best of our
knowledge, no study to date has investigated the putative
anti-melanogenic effects of soyasaponins. Therefore, in the present
study, we investigated the anti-melanogenic effects of soyasaponin
Ag on α-MSH-induced melanin synthesis in B16F10 melanoma cells.
Materials and methods
Chemicals and antibodies
α-MSH, 3-(4,5,-dimethylthiazol-2-yl)-2,5-diphenyl
tetrazolium bromide (MTT), L-DOPA and arbutin were purchased from
Sigma-Aldrich (St. Louis, MO, USA). Antibodies to tyrosinase
(sc-73244), TRP-1 (sc-58438), TRP-2 (sc-74439) and
microphthalmia-associated transcription factor (MITF; sc-52938)
were obtained from Santa Cruz Biotechnology, Inc. (Santa Cruz, CA,
USA).
Isolation of soyasaponin
Soybean saponins Ag were purified by the method
previously described in the study by Shiraiwa et al
(15) with some modifications.
The seed hypocotyls were extracted in a 10-fold volume (v/v) of 80%
(v/v) aqueous methanol for 24 h at room temperature. The extracts
were concentrated in vacuo to evaporate the solvents. The
dried extracts were suspended in a mixture of 2-butanol, water and
acetic acid (final concentration, 0.1%; equal to the 10-fold volume
used for extraction) and followed by overnight incubation at room
temperature. Following incubation, the upper (butanol) phase was
collected, evaporated and dried under a vacuum. To prepare crude
saponin solutions, the dried powders were dissolved in a small
amount of preparative HPLC solvent [34% (v/v) acetonitrile, 6%
(v/v) 2-propanol, 0.1% (v/v) acetic acid and 59.9% (v/v) water].
The crude saponin solutions were then filtered through 0.45-mm
membrane filters and were analyzed repeatedly on a preparative
reverse-phase HPLC system. The purity of saponin Ag measured by
liquid chromatography-photodiode array-tandem mass spectrometry
(LC-PDA-MS/MS) analysis was >95%, based on the total peak area
monitored at 205 nm.
LC-PDA-MS/MS analysis
Analysis of the saponin components was carried out
by LC-PDA-MS/MS with an analytical reverse-phase column (Develosil
C30-UG-5, 4.6×150 mm; Nomura Chemical Co., Ltd., Aichi, Japan) at
40°C. Solvent A (94% acetonitrile, 6% 2-propanol and 0.1% formic
acid) and solvent B (94% water, 6% 2-propanol, and 0.1% formic
acid) were used. Gradient elution from 10.64% (v/v) solvent A (10%
acetonitrile) to 53.19% (v/v) solvent A (50% acetonitrile) was
performed for 40 min (gradient increased by 1% acetonitrile per
minute) at a flow rate of 0.15 ml/min (Prominence UFLP system;
Shimadzu Corp., Kyoto, Japan). Then, the column was washed by
increasing the concentration of solvent A to 100% (v/v) for 5 min;
after which, the eluates composition returned to the initial state
of 10.64% (v/v) solvent A for 15 min. The eluate from the column
was monitored at 205 and 292 nm using a PDA detector, and MS and
MS/MS data of the eluates were acquired with a tandem mass
spectrometer (LTQ Orbitrap XL; Thermo Fisher Scientific, Inc.,
Waltham MA, USA) in positive-ion mode using electrospray ionization
[ESI(+)]. Saponin contents were calculated from a standard curve on
the basis of the peak area of purified saponin Ag monitored at 205
nm.
Cell culture
B16F10 cells, established to produce melanin and
respond to α-MSH (20) were
purchased from the American Type Culture Collection (ATCC;
CRL-6475). Typically, the B16F10 cells were cultured in α-modified
Eagle's minimal essential medium containing 10% fetal bovine serum
(HyClone, Logan, UT, USA) and 1% penicillin-streptomycin (10,000
unit/10,000 µg/ml) (Gibco, Grand Island, NY, USA) in a
humidified atmosphere containing 5% CO2 in air at
37°C.
Cell viability assay
Cell viability was determined using
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
(Sigma-Aldrich). The B16F10 cells were cultured in 96-well plates.
After a 24-h incubation, the cells were treated with soyasaponin Ag
(25–100 µM) and then incubated for 48 h in an atmosphere of
5% CO2 at 37°C. After 48 h, MTT solution (5 mg/ml) was
added to each well and the cells were incubated for 4 h at 37°C.
The supernatants were removed, and dimethylsulphoxide (DMSO) was
added to dissolve the formazan crystals. The absorbance of each
well was estimated at 570 nm using an ELISA plate reader (Tecan,
Mannedorf, Switzerland). The percentage of living cells was
determined relative to the control group.
Melanin content assay
The B16F10 cells (3.5×104) were cultured
in 6-well plates at 37°C in an atmosphere of 5% CO2 for
24 h. After 24 h, the cells were treated with α-MSH (20 nM) and
medium containing arbutin (Sigma-Aldrich; arbutin was diluted in
0.1 M phosphate buffer and used at 1 and 2 mM concentrations as a
positive control) or soyasaponin Ag (25, 50 and 100 µM) for
48 h. After 48 h, the cells were washed with phosphate-buffered
saline (PBS), and the cells were detached by incubation with
trypsin-EDTA and centrifuged at 3,000 rpm for 5 min. The
supernatant was then discarded and the cell pellet was solubilized
in 1 N NaOH at 60°C for 2 h. To determine the melanin content, the
absorbance was measured at 405 nm using an ELISA plate reader and
then normalized to the total protein concentration.
Cellular tyrosinase activity assay
B16F10 cell seeding and treatment methods were the
same as those used to measure melanin contents. Briefly, the cells
were cultured in 6-well plates, and after 24 h, the cells were
treated with α-MSH (20 nM) and soyasaponin Ag-containing medium
(25–100 μM) for 48 h. After treatment, the cells were washed twice
with cold PBS and lysed with 100 µl of RIPA buffer
containing 0.1 mM phenylmethylsulfonyl fluoride (PMSF) for 30 min
on ice. The lysates were centrifuged (13,000 rpm for 15 min), 20
µl of the supernatant was mixed with 100 µl of 4%
N,N-dimethylformamide, 50 µl of 5 mM L-DOPA and 50 µl
of 20.7 mM 3-methyl-2-benzothiazolinone hydrazine (MBTH) in a
96-well plate and then incubated at 37°C for 30 min. To determine
the tyrosinase activity, the absorbance was measured at an
absorbance wavelength of 505 nm, and then normalized to the total
protein concentration.
Western blot analysis
The B16F10 cells were treated with soyasaponin Ag
(25–100 µM) for 24 h and then further treated with α-MSH (20
nM) and the soyasaponin sample for a further 24 h. After these
treatments, the cells were collected and lysed with RIPA buffer
containing 1X solution (50 mM Tris HCl, 150 mM NaCl, 1% Nonidet
P-40, 0.5% deoxycholate, 0.1% SDS) for 30 min on ice. The cell
lysates were centrifuged (10,000 rpm for 10 min), and the protein
concentrations were determined using the BCA protein assay.
Proteins were separated by 10% SDS-PAGE and transferred onto
nitrocellulose membranes (GE Healthcare, Buckinghamshire, UK). The
membranes were blocked with blocking buffer (Thermo Fisher
Scientific, Inc.) and then incubated with the appropriate
antibodies. The membranes were washed with wash buffer and
incubated with anti-mouse (sc-2371) or anti-rabbit (sc-2379)
horseradish peroxidase-conjugated secondary antibodies (Santa Cruz
Biotechnology, Inc.) at 1:1,000 dilution for 1 h. The
immunoreactive bands were enhanced using chemiluminescence
reagents. The loading control was assessed using an anti-β-actin
antibody.
Statistical analysis
All data were obtained in triplicate and are
presented as the means ± standard deviation. Values were compared
using the Student's t-test. P<0.05 was considered to indicate a
statistically significant difference.
Results
Effect of soyasaponin Ag on cell
viability and melanin synthesis in α-MSH-stimulated B16F10
cells
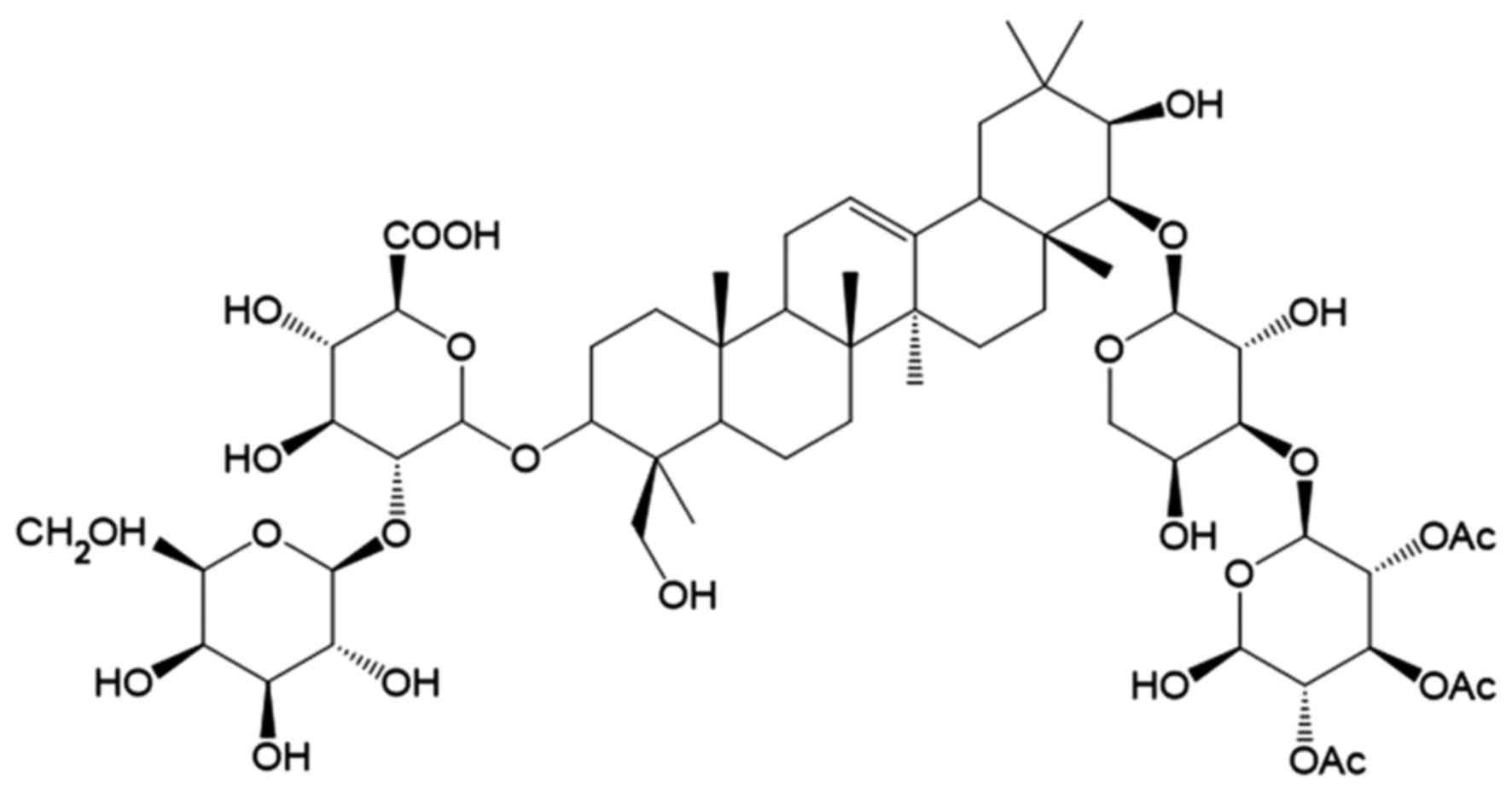
The chemical structure of saponin Ag is shown in
Fig. 1. The first step in this
study was to determine any potential cytotoxic effects of
soyasaponin Ag in view of its potential utilization as a functional
food or cosmetic agent. We performed cell viability assays with
soyasaponin Ag to determine its in vitro cytotoxicity in
murine B16F10 melanoma cells. More specifically, we utilized the
MTT colorimetric assay for cell viability. In short, MTT is reduced
to purple formazan crystals by the enzymatic activity of the
mitochondria. Since for most living cells total mitochondria
activity correlates with the number of viable cells, the assay is a
simple and reliable method for determining potential cytotoxic
effects (21). To determine the
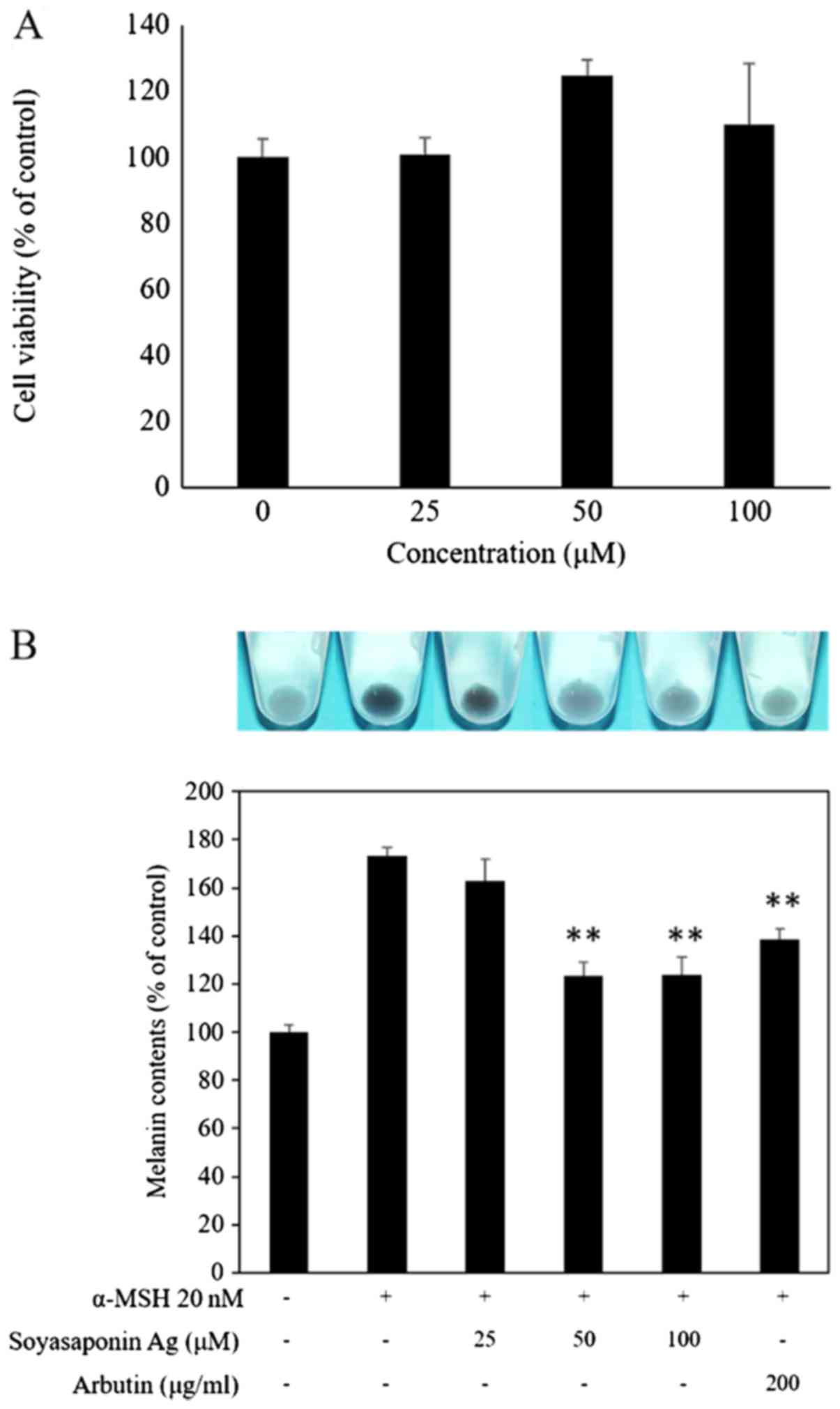
effect of soyasaponin Ag on B16F10 cell viability, the cells were
treated with 25–100 µM of soyasaponin Ag for 48 h (Fig. 2A). No significant effect on cell
viability was detected when using this concentration range of
soyasaponin Ag. To determine the effect of soyasaponin Ag (25–100
µM) treatment on α-MSH-induced melanin synthesis, the
melanin content of the B16F10 cells was evaluated photometrically.
As shown in Fig. 2B, α-MSH
markedly upregulated the B16F19 cell melanin content (P<0.05).
Of ntoe, treatment with soyasaponin Ag at 50 and 100 µM
significantly decreased the cellular melanin content of the
α-MSH-stimulated B16F10 cells (P<0.01). In the control
experiments, arbutin (positive control), a known melanogenic
inhibitor (22) was shown to
attenuate the melanin content of stimulated B16F10 cells (Fig. 2B).
Effect of soyasaponin Ag on tyrosinase
activity in α-MSH-stimulated B16F10 cells
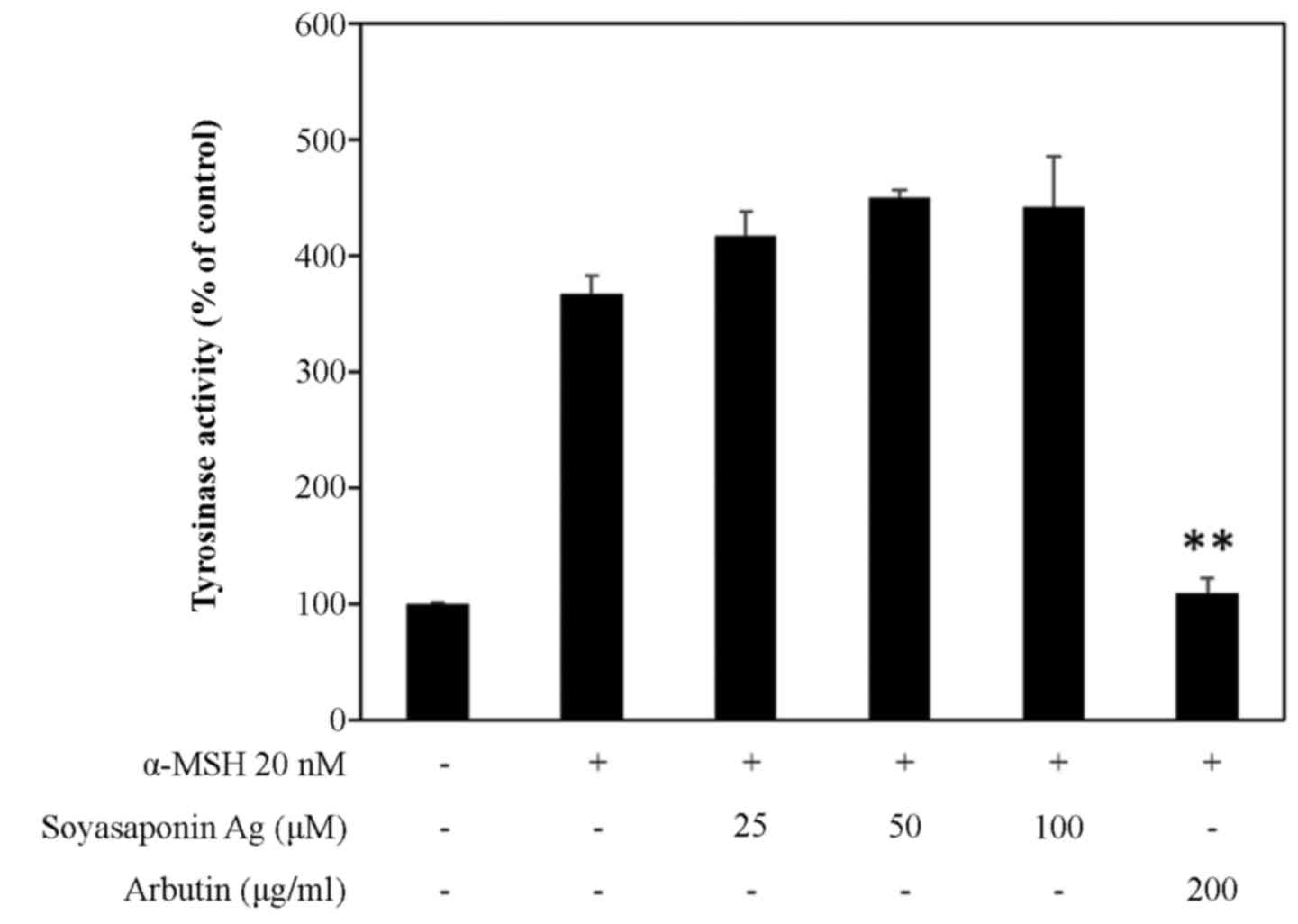
Tyrosinase is known to play a key role in melanin
synthesis. Thus, in this study, to investigate the mechanisms
underlying the inhibition of melanogenesis, we measured tyrosinase
activity in α-MSH-stimulated B16F10 cells (Fig. 3). B16F10 melanoma cells treated
with up to 100 μM soyasaponin Ag did not exhibit any inhibition of
tyrosinase activity. On the other hand, in the control experiments,
arbutin (22), a direct inhibitor
of tyrosinase activity, at 200 µg/ml completely inhibited
α-MSH stimulated tyrosinase activity to basal levels (Fig. 3).
Effects of soyasaponin Ag on the
expression of melanin synthesis factors in α-MSH-stimulated B16F10
cells
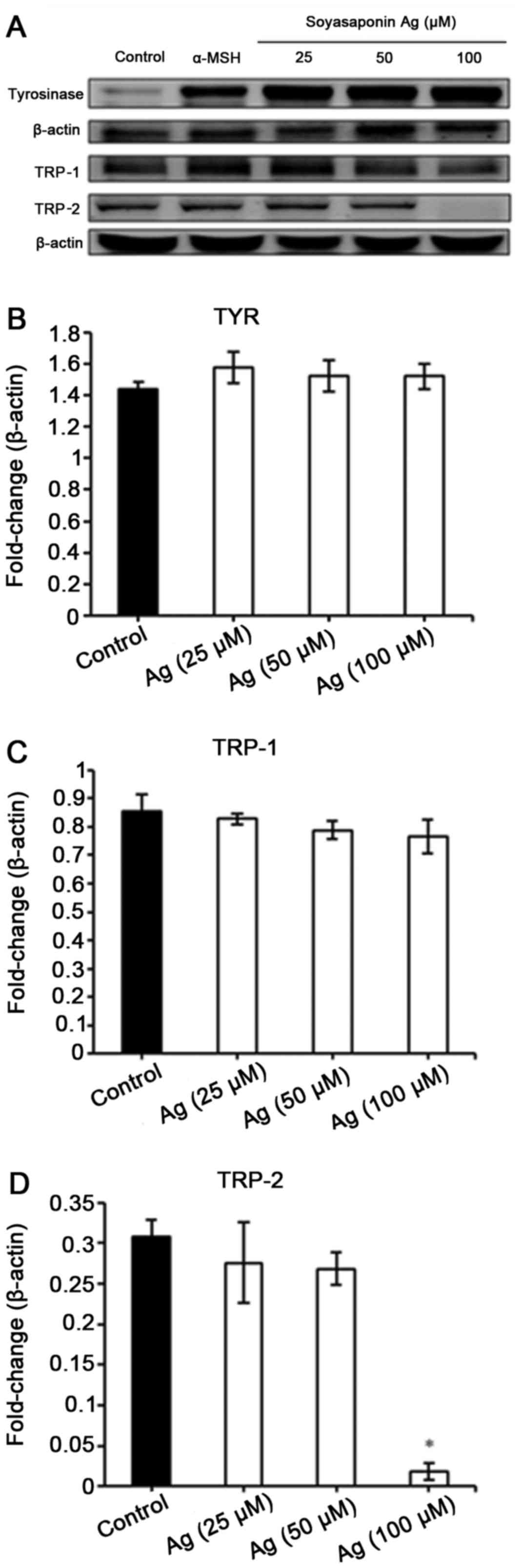
We then examined the effect of soyasaponin Ag
treatment on the the expression of tyrosinase, TRP-1 and TRP-2 in
B16F10 cells stimulated with α-MSH. As demonstrated by western blot
analysis, soyasaponin did not affect α-MSH-induced tyrosinase and
TRP-1 expression (Fig. 4). Of
note, treatment of the B16F10 cells with 100 µM of
soyasaponin Ag abolished TRP-2 expression (Fig. 4).
Discussion
Saponins are beneficial bioactive molecules that are
widely distributed in plants and exhibit vast structural and
functional diversity; thus, they may be important substances which
are worthy of investigation for potential pharmaceutical
applications.
The process of melanogenesis is under complex
regulatory control by at least 125 genetic loci (23). The enzyme tyrosinase plays a
critical role in melanin biosynthesis, and in concert with TRP-1
and TRP-2 is responsible for modifying melanin into different forms
(5,24). Thus, the expression level and
catalytic activity of these enzymes correlate with melanin
production and have been widely investigated with respect to the
whitening properties of natural compounds (25). The putative effects of soyasaponin
Ag on tyrosinase activity, gene expression, or maturation in
melanin biosynthesis during skin pigmentation have not been studied
to date, at least to the best of our knowledge. In this study, we
demonstrate that soyasaponin Ag inhibits melanogenesis in B16F10
murine melanoma cells via the downregulation of tyrosinase TRP-2
expression. In our initial experiments, we determined that
soyasaponin Ag had no cytotoxic effect on B16F10 melanoma cell
viability, even at high concentrations. This first indication of
the safe use of soyasaponin Agas an anti-melanogenic agent needs
however, to be verified in normal melanocytes and in in vivo
models.
There are different types of inhibitory mechanisms
for melanin synthesis in melanocytes. Indeed, hypopigmenting agents
have been classified according to their mechanism of action into
three categories: a) regulation of enzyme, which is subdivided into
three categories: i) regulation of transcription and maturation of
tyrosinase, ii) inhibition of tyrosinase activity, and iii)
post-transcriptional control of tyrosinase; b) inhibition of
melanosome transfer; and c) additional mechanisms, such as
regulation of the melanocyte environment and antioxidant agents
(26). Thus, many
anti-melanogenic agents, such as kojic acid, hydroquinone and
arbutin, are direct inhibitors of tyrosinase enzyme activity
(25). Additionally, some
compounds exert their anti-melanogenic activities via the
suppression of tyrosinase gene expression, although they do not
have any direct inhibitory effect on tyrosinase activity in general
(27). This type of mechanism was
previously identified for acetylsalicylic acid (ASA) in B16
melanoma cells, where ASA exerted a strong dose-dependent
inhibitory effect on tyrosinase expression (28). Moreover, ASA did not exert any
effect on mushroom tyrosinase activity (28). Other compounds, such as
oleoylethanolamide were found to have a more general effect on
melanogenesis, inhibiting the expression levels of MITF, TRP-1 and
tyrosinase (29). Indeed, MITF
plays a crucial role in melanogenesis as the master regulator of
tyrosinase, TRP-1 and TRP-2 expression (30). The hydrolyzed ginseng extract, was
found to downregulate MITF and TRP-1 expression B16F10 cells, but
only indirectly to attenuate cellular tyrosine activity (31). Moreover, the coumarin derivative,
osthol, was found to decrease tyrosinase, TRP-1 and TRP-2
expression, but not to modulate tyrosinase activity (32).
The results of the present study indicate that
soyasapogenol A treatment decreased the expression of TRP-2 protein
and attenuated melanin production. The most likely explanation for
these findings is that soyasapogenol A affects MITF
activity/expression, which in other studies has been associated
with hypopigmentation (33–38). Importantly, a number of studies
have reported that not all skin-whitening agents, such as nicotinic
acid hydroxamate, glutathione-monoisopropyl ester and plumbagin,
simultaneously inhibit TRP-1 and TRP-2 expression (39–41). In addition, the melanin content
can be affected by mutation and degradation of the tyrosinase
protein (42).
In conclusion, to the best of our knowledge, the
present study is the first to demonstrate that soyasapogenol A
inhibits melanogenesis by downregulating the protein levels of
TRP-1 and TRP-2, resulting in reduced melanin production.
Therefore, given the biological properties of soyasapogenol A, we
suggest that saponin Ag is a putative skin-whitening agent and may
be effective in the treatment of hyperpigmentation disorders, and
may also be used in skin-whitening cosmetics. Further
investigations are required in order to fully determine the
depigmenting mechanisms of action of soyasapogenol A, as well as
its anti-melanogenic activity in vivo.
Acknowledgments
This study was carried out with the support of the
Basic Science Research Program through the National Research
Foundation of Korea (NRF) funded by the Ministry of Education
(NRF-2015R1D1A1A09060925).
References
|
1
|
Videira IF dos S, Moura DF and Magina S:
Mechanisms regulating melanogenesis. An Bras Dermatol. 88:76–83.
2013. View Article : Google Scholar
|
|
2
|
Regad T: Molecular and cellular
pathogenesis of melanoma initiation and progression. Cell Mol Life
Sci. 70:4055–4065. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Brenner M and Hearing VJ: The protective
role of melanin against UV damage in human skin. Photochem
Photobiol. 84:539–549. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Levine N and Maibach HI: Pigmentation and
Pigmentary Disorders A Volume in the Dermatology: Clinical and
Basic Science Series. 1st Edition. CRC Press; Boca Raton, FL:
1993
|
|
5
|
Costin GE and Hearing VJ: Human skin
pigmentation: Melanocytes modulate skin color in response to
stress. FASEB J. 21:976–994. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
D'Mello SAN, Finlay GJ, Baguley BC and
Askarian-Amiri ME: Signaling pathways in melanogenesis. Int J Mol
Sci. 17:11442016. View Article : Google Scholar :
|
|
7
|
Millington GWM: Proopiomelanocortin
(POMC): The cutaneous roles of its melanocortin products and
receptors. Clin Exp Dermatol. 31:407–412. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Abdel-Malek Z, Swope VB, Suzuki I, Akcali
C, Harriger MD, Boyce ST, Urabe K and Hearing VJ: Mitogenic and
melanogenic stimulation of normal human melanocytes by melanotropic
peptides. Proc Natl Acad Sci USA. 92:1789–1793. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Prota G: Melanin-producing cells. Melanins
melanogenesis. Academic Press; San Diego, CA: pp. 14–33. 1992,
View Article : Google Scholar
|
|
10
|
Kobayashi T, Imokawa G, Bennett DC and
Hearing VJ: Tyrosinase stabilization by Tyrp1 (the brown locus
protein). J Biol Chem. 273:31801–31805. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Choe YS: The effect of basic cosmetic
ingredients on the melanogenesis in the B16F10 mouse melanocyte.
Konkuk University; Seoul: 2006
|
|
12
|
del Marmol V and Beermann F: Tyrosinase
and related proteins in mammalian pigmentation. FEBS Lett.
381:165–168. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kim SS, Kim MJ, Choi YH, Kim BK, Kim KS,
Park KJ, Park SM, Lee NH and Hyun CG: Down-regulation of
tyrosinase, TRP-1, TRP-2 and MITF expressions by citrus press-cakes
in murine B16 F10 melanoma. Asian Pac J Trop Biomed. 3:617–622;
discussion 621–622. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Guang C, Chen J, Sang S and Cheng S:
Biological functionality of soyasaponins and soyasapogenols. J
Agric Food Chem. 62:8247–8255. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Shiraiwa M, Kudo S, Shimoyamada M, Harada
K and Okubo K: Composition and structure of 'group A saponin' in
soybean seed. Agric Biol Chem. 55:315–322. 1991.
|
|
16
|
Shiraiwa M, Harada K and Okubo K:
Composition and structure of 'group B saponin' in soybean seed.
Agric Biol Chem. 55:911–917. 1991.PubMed/NCBI
|
|
17
|
Zhang W and Popovich DG: Chemical and
biological characterization of oleanane triterpenoids from soy.
Molecules. 14:2959–2975. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Yang SH, Ahn EK, Lee JA, Shin TS,
Tsukamoto C, Suh JW, Mei I and Chung G: Soyasaponins Aa and Ab
exert an anti-obesity effect in 3T3-L1 adipocytes through
downregulation of PPARγ. Phytother Res. 29:281–287. 2015.
View Article : Google Scholar
|
|
19
|
Xiao JX, Huang GQ and Zhang SH:
Soyasaponins inhibit the proliferation of Hela cells by inducing
apoptosis. Exp Toxicol Pathol. 59:35–42. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Park SH, Kim DS, Park SH, Shin JW, Youn SW
and Park KC: Inhibitory effect of p-coumaric acid by Rhodiola
sachalinensis on melanin synthesis in B16F10 cells. Pharmazie.
63:290–295. 2008.PubMed/NCBI
|
|
21
|
van Meerloo J, Kaspers GJL and Cloos J:
Cell sensitivity assays: the MTT assay. Methods Mol Biol.
731:237–245. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Maeda K and Fukuda M: Arbutin: Mechanism
of its depigmenting action in human melanocyte culture. J Pharmacol
Exp Ther. 276:765–769. 1996.PubMed/NCBI
|
|
23
|
Yamaguchi Y, Brenner M and Hearing VJ: The
regulation of skin pigmentation. J Biol Chem. 282:27557–27561.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Hearing VJ and Tsukamoto K: Enzymatic
control of pigmentation in mammals. FASEB J. 5:2902–2909.
1991.PubMed/NCBI
|
|
25
|
Chang TS: An updated review of tyrosinase
inhibitors. Int J Mol Sci. 10:2440–2475. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Kim H, Choi HR, Kim DS and Park KC:
Topical hypopigmenting agents for pigmentary disorders and their
mechanisms of action. Ann Dermatol. 24:1–6. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chung SY, Seo YK, Park JM, Seo MJ, Park
JK, Kim JW and Park CS: Fermented rice bran downregulates MITF
expression and leads to inhibition of α-MSH-induced melanogenesis
in B16F1 melanoma. Biosci Biotechnol Biochem. 73:1704–1710. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Sato K, Takahashi H, Iraha R and Toriyama
M: Down-regulation of tyrosinase expression by acetylsalicylic acid
in murine B16 melanoma. Biol Pharm Bull. 31:33–37. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhou J, Ren T, Li Y, Cheng A, Xie W, Xu L,
Peng L, Lin J, Lian L, Diao Y, et al: Oleoylethanolamide inhibits
α-melanocyte stimulating hormone-stimulated melanogenesis via ERK,
Akt and CREB signaling pathways in B16 melanoma cells. Oncotarget.
May 23–2017.Epub ahead of print.
|
|
30
|
Yasumoto K, Amae S, Udono T, Fuse N,
Takeda K and Shibahara S: A big gene linked to small eyes encodes
multiple Mitf isoforms: Many promoters make light work. Pigment
Cell Res. 11:329–336. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Han JS, Sung JH and Lee SK:
Antimelanogenesis activity of hydrolyzed ginseng extract (GINST)
via inhibition of JNK mitogen-activated protein kinase in B16F10
cells. J Food Sci. 81:H2085–H2092. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Beom Kim S, Kim C, Liu Q, Hee Jo Y, Joo
Choi H, Hwang BY, Kyum Kim S and Kyeong Lee M: Optimization of
extraction conditions for osthol, a melanogenesis inhibitor from
Cnidium monnieri fruits. Pharm Biol. 54:1373–1379. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kim DS, Hwang ES, Lee JE, Kim SY, Kwon SB
and Park KC: Sphingosine-1-phosphate decreases melanin synthesis
via sustained ERK activation and subsequent MITF degradation. J
Cell Sci. 116:1699–1706. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Koo JH, Rhee KS, Koh HW, Jang HY, Park BH
and Park JW: Guggulsterone inhibits melanogenesis in B16 murine
melanoma cells by downregulating tyrosinase expression. Int J Mol
Med. 30:974–978. 2012.PubMed/NCBI
|
|
35
|
Kim DS, Kim SY, Chung JH, Kim KH, Eun HC
and Park KC: Delayed ERK activation by ceramide reduces melanin
synthesis in human melanocytes. Cell Signal. 14:779–785. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Tachibana M: MITF: A stream flowing for
pigment cells. Pigment Cell Res. 13:230–240. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Li X, Guo L, Sun Y, Zhou J, Gu Y and Li Y:
Baicalein inhibits melanogenesis through activation of the ERK
signaling pathway. Int J Mol Med. 25:923–927. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Lee JH, Jang JY, Park C, Kim BW, Choi YH
and Choi BT: Curcumin suppresses α-melanocyte stimulating
hormone-stimulated melanogenesis in B16F10 cells. Int J Mol Med.
26:101–106. 2010.PubMed/NCBI
|
|
39
|
Lin YS, Chuang MT, Chen CH, Chien MY and
Hou WC: Nicotinic acid hydroxamate downregulated the melanin
synthesis and tyrosinase activity through activating the MEK/ERK
and AKT/GSK3β signaling pathways. J Agric Food Chem. 60:4859–4864.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chung BY, Choi SR, Moon IJ, Park CW, Kim
YH and Chang SE: The glutathione derivative, GSH monoethyl ester,
may effectively whiten skin but GSH does not. Int J Mol Sci.
17:6292016. View Article : Google Scholar :
|
|
41
|
Oh TI, Yun JM, Park EJ, Kim YS, Lee YM and
Lim JH: Plumbagin suppresses α-MSH-induced melanogenesis in B16F10
mouse melanoma cells by inhibiting tyrosinase activity. Int J Mol
Sci. 18:3202017. View Article : Google Scholar
|
|
42
|
Ando H, Kondoh H, Ichihashi M and Hearing
VJ: Approaches to identify inhibitors of melanin biosynthesis via
the quality control of tyrosinase. J Invest Dermatol. 127:751–761.
2007. View Article : Google Scholar : PubMed/NCBI
|