Introduction
It is well known that platelet-derived growth factor
(PDGF) is a major mitogen for connective tissue cells and certain
other cell types (1). The PDGF
family consists of four different polypeptide chains encoded by
different genes, which have been identified as PDGF-A, PDGF-B and
the recently discovered PDGF-C and PDGF-D, and 4 homodimers
(PDGF-AA, PDGF-BB, PDGF-CC and PDGF-DD) and a heterodimer (PDGF-AB)
(2). To maintain structural bone
integrity and mineral homeostasis, bone metabolism is tightly
regulated by bone remodeling, which consists mainly of osteoblast
bone formation and osteoclastic bone resorption (3). In bone metabolism, PDGF reportedly
increases proliferation of osteoblasts and inhibits differentiation
(4). As for the fracture healing
process, PDGF has a crucial role as a systemic factor, and
regulates bone remodeling (5).
Additionally, it has been reported that administration of
recombinant platelet-derived growth factor-BB (PDGF-BB) accelerates
fracture healing in the geriatric osteoporotic rat (6). We previously demonstrated that
PDGF-BB stimulates interleukin-6 (IL-6) synthesis via p44/p42
mitogen-activated protein kinase (MAPK), p38 MAPK and
stress-activated protein kinase/c-Jun N-terminal kinase (SAPK/JNK)
in osteoblast-like MC3T3-E1 cells (7), and that phosphoinositide 3-kinase
(PI3K)/Akt and p70 S6 kinase negatively regulate the synthesis
(7,8). It has recently been reported that
PDGF-BB induces the migration of human osteoblasts (9). However, the details behind the
effect of PDGF effect in osteoblast migration has not been fully
clarified.
It is generally recognized that heat shock proteins
(HSPs) play pivotal roles as molecular chaperones, which facilitate
the refolding of improperly folded proteins induced by stress, or
assist in their elimination via chaperone-mediated autophagy or the
ubiquitin proteasome system (10). Among them, HSP27 (HSPB1) belongs
to the HSPB family, with monomeric molecular masses ranging from 12
to 43 kDa, so called small HSPs. HSP27 is ubiquitously expressed in
human cells and tissues, and its functions have been clarified in
the studies of tissues which have higher expression levels such as
smooth, skeletal and cardiac muscles (11). Furthermore, it is generally known
that post-translational modifications such as phosphorylation
regulate the functions of HSP27 (12). HSP27 ordinarily exists as
unphosphorylated multioligomers in cells and possesses three serine
residues (Ser-15, Ser-78 and Ser-82) and one threonine residue
(Thr-143) that can be phosphorylated by related enzymes such as p38
MAPK (12,13). HSP27 oligomerization is mainly
regulated by Ser-78 and/or Ser-82 phosphorylation (13). Once HSP27 is phosphorylated, a
conformational change occurs from the aggregated form to the dimer
(11,12). With regard to HSP27 in
osteoblasts, HSP27 is reportedly involved in their differentiation
and apoptosis (14). The
expression levels of HSP27 in osteoblasts under unstimulated
conditions (15) including
osteoblast-like MC3T3-E1 cells (16) are quite low. In our previous
studies, we demonstrated that various bone remodeling mediators,
such as prostaglandin D2 (PGD2),
endothelin-1, PGE2, PGF2α and transforming
growth factor-β (TGF-β), enable HSP27 to be induced in
osteoblast-like MC3T3-E1 cells (17–21). In addition, we reported that
phosphorylated HSP27 changes its localization from the cytosol to
the perinuclear region, and acts as a negative regulator in
triiodothyronine-induced osteocalcin synthesis in MC3T3-E1 cells,
but has a stimulatory effect on mineralization (22). However, the exact roles of HSP27
especially in the function of osteoblasts including migration
remain to be elucidated.
In the present study, we investigated the role of
HSP27 in the PDGF-BB-stimulated migration of osteoblast-like
MC3T3-E1 cells. We herein showed that HSP27 acts as a negative
regulator in PDGF-BB-stimulated cell migration, and the inhibitory
effect of phosphorylated HSP27 on the migration is stronger than
that of unphosphorylated HSP27.
Materials and methods
Materials
PDGF-BB and TGF-β were both purchased form R&D
Systems, Inc. (Minneapolis, MN, USA). HSP27 antibodies used to
detect the expression of HSP27, glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) antibodies and deguelin, were obtained from
Santa Cruz Biotechnology, Inc. (Santa Cruz, CA, USA). HSP27
antibodies used to confirm the transfection with the WT HSP27
vector, and phospho-specific (p)-HSP27 (Ser-82) antibodies were
purchased from Enzo Life Sciences, Inc. (Famingdale, NY, USA).
PD98059, SB203580, SP600125, Akt inhibitor and rapamycin were all
obtained from Calbiochem-Novabiochem (La Jolla, CA, USA). p-p44/p42
MAPK, p-p38 MAPK and p-SAPK/JNK antibodies were all purchased from
Cell Signaling Technology, Inc. (Beverly, MA, USA). WT HSP27 and
mutant human HSP27 subcloned into the pcDNA3.1(+) mammalian
expression vector were kindly provided by Dr. C. Schafer (Klinikum
Grosshadern, Ludwig Maximilians University, Munich, Germany). Rega
rding mutant HSP27 vectors, three serine residues (Ser-15, 78 and
82) of the HSP27 cDNAs had been mutated to alanine (3A) to prevent
the phosphorylation of HSP27, or mutated to aspartic acid (3D) to
imitate the phosphorylated HSP27 form, as previously described
(23). The eukaryotic expression
vector, pcDNA3.1(+) was obtained from Life Technologies (Carlsbad,
CA, USA). A BCA protein assay kit was purchased from Thermo Fisher
Scientific Inc. (Waltham, MA, USA). An ECL western blotting
detection system was obtained from GE Healthcare UK Ltd.
(Buckinghamshire, UK). Other materials and chemicals were obtained
from commercial sources. PD98059, SB203580, SP600125, Akt
inhibitor, deguelin and rapamycin were dissolved in dimethyl
sulfoxide (DMSO). The maximum concentration of DMSO was 0.3%, which
did not affect the cell migration assay.
Cell culture
Cloned osteoblast-like MC3T3-E1 cells, which were
originally derived from newborn mouse calvaria (24) were maintained as previously
described (25). In brief, the
cells were maintained in α-minimum essential medium (α-MEM)
containing 10% fetal bovine serum (FBS) in a humidified 95% air, 5%
CO2 atmosphere at 37°C. The cells were seeded into 90-mm
diameter dishes (1–2×105 cells/dish) in α-MEM containing
10% FBS. After 4 days, the medium was exchanged for α-MEM
containing 0.3% FBS. Thereafter, the cells were cultured for 6 h
and then used for the cell migration assay.
Transient transfections
For transient transfections, the MC3T3-E1 cells were
seeded into 90-mm diameter dishes (5–15×104 cells/dish)
in α-MEM containing 10% FBS. After 4 days, the medium was removed,
and the cells were washed with 10 ml of α-MEM medium without FBS.
The cultured cells were then transfected with 6.6 µg of the
WT, control (empty), mutant 3A or 3D HSP27 plasmids pcDNA3.1(+)
vector using the UniFector transfection reagent (B-Bridge
International, Inc., Mountain View, CA, USA) in 5.6 ml of α-MEM
medium without FBS. Five hours after transfection, 6.6 ml of medium
with α-MEM containing 0.6% FBS was added. At 3 days after
transfection, the medium was exchanged to α-MEM containing 0.3% FBS
for the experiments.
Cell migration assay
Cell migration assay was performed as previously
described in the method of Kara giosis et al (26) using Boyden chamber [polycarbonate
membrane with 8-µm pores (Transwell®; Corning
Costar, Cambridge, MA, USA)]. Briefly, the cultured cells were
trypsinized, and seeded (5–10×104 cells/well) onto the
upper chamber in α-MEM containing 0.3% FBS. PDGF-BB (3 ng/ml) was
added to the lower chamber, and the cells were incubated for 16 h
at 37°C. After mechanical removal of the cells on the upper surface
of the membrane, the migrated cells adherent to the underside of
the membrane were fixed with 4% paraformaldehyde and stained with
4′,6-diamidino-2-phenylindole (DAPI) solution. The number of
stained cells was counted using fluorescence microscopy from three
randomly chosen high power fields per well at x20 magnification.
When indicated, the cells were used after pretreatment with
PD98059, SB203580, SP600125, Akt inhibitor, deguelin or rapamycin
in the lower chamber for 60 min.
Western blot analysis
The cultured cells or transfected cells were
stimulated by 30 ng/ml of PDGF-BB or 5 ng/ml of TGF-β in α-MEM
containing 0.3% FBS for the indicated periods. The cells were
washed twice in phosphate-buffered saline (PBS) and then lysed,
homogenized and sonicated in a lysis buffer: 62.5 mM Tris-HCl, pH
6.8, 2% sodium dodecyl sulfate (SDS), 50 mM dithiothreitol and 10%
glycerol. The protein concentration of each sample was measured by
a BCA protein assay kit. SDS-polyacrylamide gel electrophoresis
(PAGE) was performed by the method described by Laemmli (27) on 10% polyacrylamide gels. The
samples from the transfected cell cultures to be quantitatively
compared by the western blot analysis were run on the same gel. The
protein was fractionated and transferred onto an Immun-Blot
polyvinylidene difluoride membrane (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA). The membranes were blocked with 5% fat-free dry
milk in Tris-buffered saline-Tween (TBS-T; 20 mM Tris-HCl, pH 7.6,
137 mM NaCl, 0.1% Tween-20) for 1 h before incubation with the
primary antibodies. Western blot analysis was performed as
previously described (28) using
the indicated primary antibodies with the appropriate secondary
antibodies. Immunoreactive bands were visualized on X-ray film by
means of the ECL western blotting detection system.
Determinations
The densitometric analysis was carried out using
image analysis software (ImageJ version 1.49; National Institutes
of Health, Bethesda, MD, USA). The background-subtracted signal
intensity for each protein and phosphorylation was normalized to
the respective intensity of GAPDH, and plotted as the fold increase
in comparison to the control cells.
Statistical analysis
All data are presented as the mean ± standard error
of the mean (SEM) of triplicate determinations from three
independent cell preparations. Statistical analysis were performed
using the Bonferroni method for multiple comparisons between pairs,
and a P-value <0.05 was considered to be significant.
Results
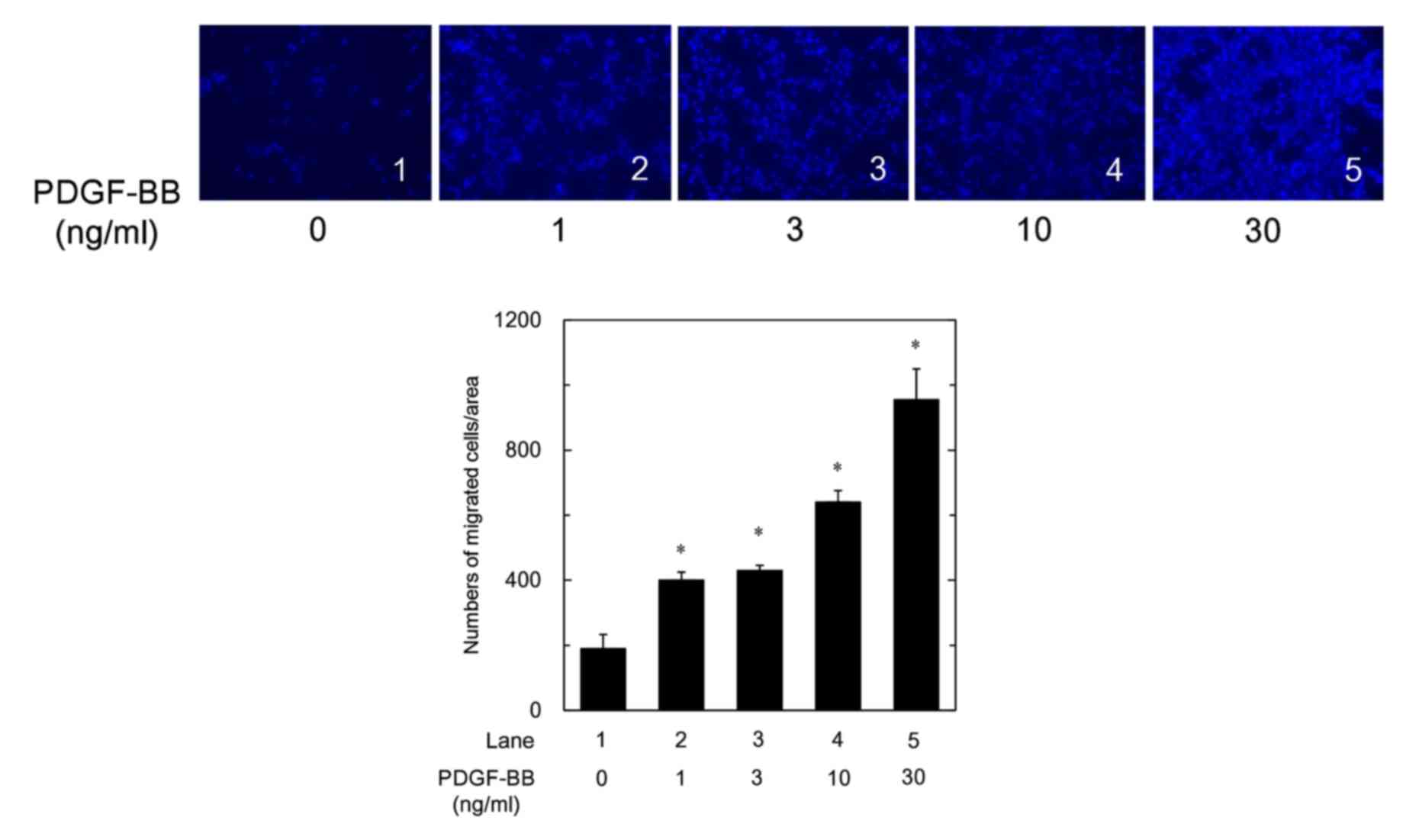
Effects of PDGF-BB on the migration,
HSP27 induction and HSP27 phosphorylation in MC3T3-E1 cells
It has been reported that PDGF-BB induces the
migration of human osteoblasts (9). We found that PDGF-BB truly
stimulated the migration of osteoblast-like MC3T3-E1 cells in a
dose-dependent manner in the range between 1 and 30 ng/ml (Fig. 1). In our previous studies
(16,29,30), we demonstrated that
osteoblast-like MC3T3-E1 cells normally have low levels of HSP27,
and HSP27 expression is induced by sphingosine 1-phosphate, basic
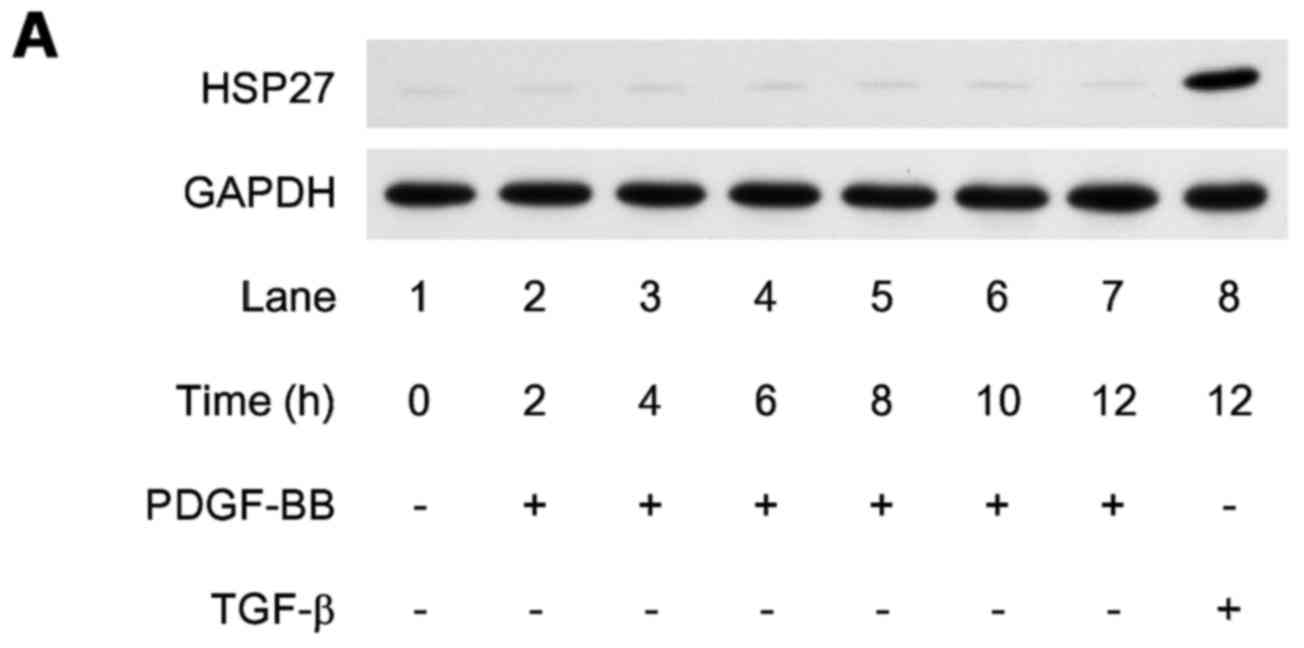
fibroblast growth factor or TGF-β. We confirmed that HSP27 levels
without stimulation were low in these cells. PDGF-BB barely induced
the expression of HSP27 up to 12 h in comparison to TGF-β (Fig. 2A).
We previously established WT HSP27-over expressing
MC3T3-E1 cells transfected with the WT HSP27 vector (31). We confirmed that the protein
expression levels of HSP27 were clearly detectable in the WT HSP27
vector-transfected MC3T3-E1 cells, in comparison with the control
empty vector-transfected cells (Fig.
2B). We next examined the effect of PDGF-BB on the
phosphorylation of HSP27 in the WT HSP27 vector-transfected
MC3T3-E1 cells. We confirmed that the phosphorylation of HSP27 was
barely detected before the PDGF-BB stimulation in the
HSP27-overexpressing MC3T3-E1 cells, but found that PDGF-BB
markedly stimulated the phosphorylation of HSP27 (Fig. 2C). The effect of PDGF-BB on the
phosphorylation of HSP27 was detected from 10 to 30 min after the
stimulation. We confirmed that PDGF-BB markedly induced the
phosphorylation of HSP27 in the HSP27-overexpressing cells at 10
min after the stimulation in comparison with that without PDGF-BB
stimulation (Fig. 2D).
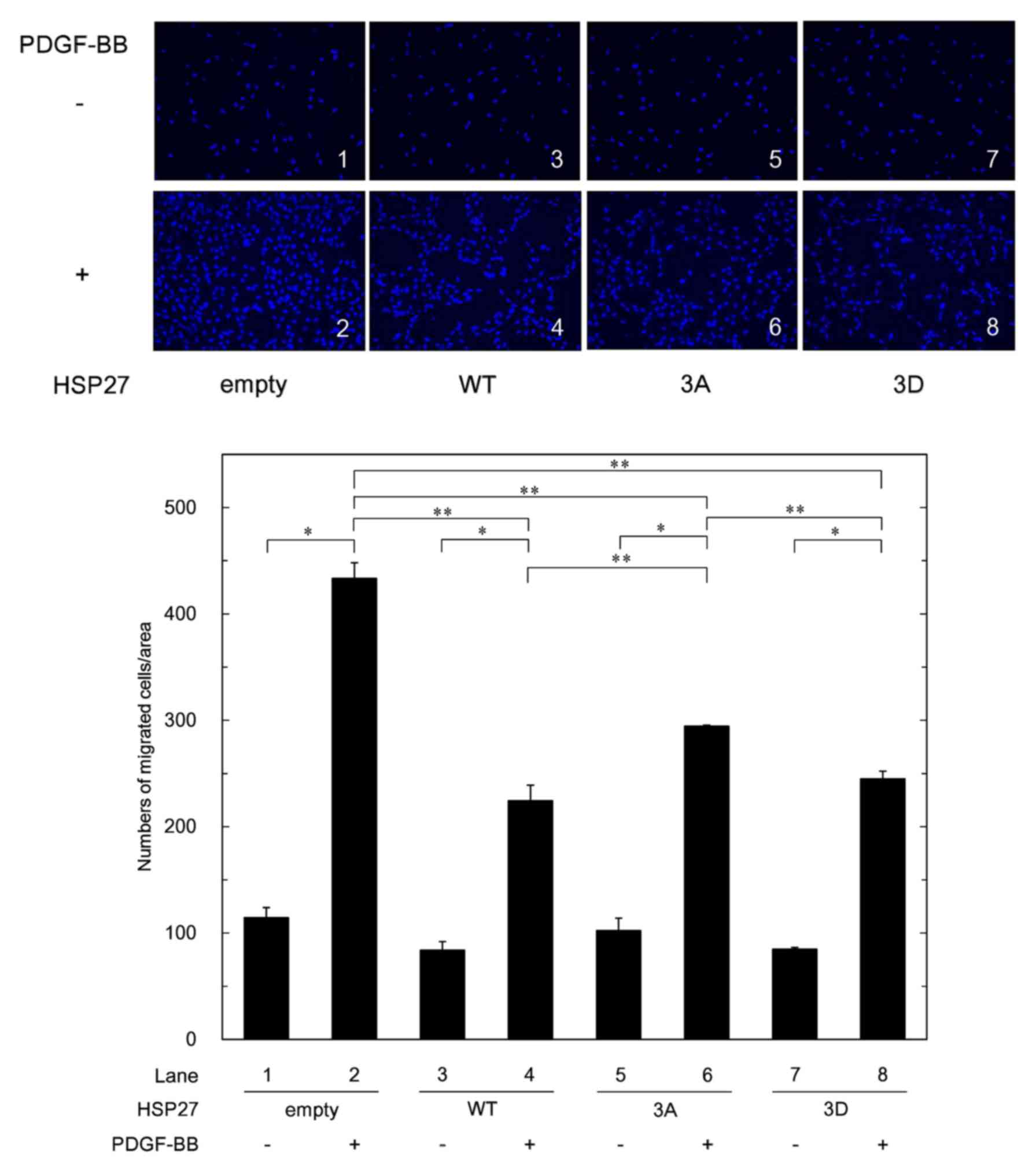
Effects of HSP27 overexpression and the
status of HSP27 phosphorylation on the PDGF-BB-induced migration of
MC3T3-E1 cells
It is currently recognized that the functions of
HSP27 are modulated by phosphorylation, resulting from its
conformational changes through dimerization of multioligomers
(10). We previously established
two mutant HSP27-transfected MC3T3-E1 cell lines, in which serine
residues (Ser-15, Ser-78 and Ser-82) were mutated into alanine (3A)
to prevent the phosphorylation or were mutated into aspartic acid
(3D) to imitate the phosphorylated form, and reported that the
phosphorylation levels of HSP27 in the 3D-transfected cells were
clearly greater than those in the 3A-transfected cells (22). We investigated the effects of
PDGF-BB on the migration of control empty vector-transfected cells
which represent normal MC3T3-E1 cells, WT HSP27-overexpressing
cells, 3A cells and 3D cells on the same day. There were no
differences in the migrated cell numbers among the different cell
groups without PDGF-BB stimulation. In comparison with the control
empty vector-transfected cells, the PDGF-BB-induced migration in
the WT HSP27-overexpressing cells was significantly reduced.
Although the difference in the PDGF-BB-induced migrated cell
numbers between the 3D cells and the 3A cells appeared subtle, the
migrated numbers of the 3D cells were significantly reduced
compared with those of the 3A cells. In addition, the
PDGF-BB-induced migrated cell numbers of the WT
HSP27-overexpressing cells were less than those of the 3A cells
(Fig. 3).
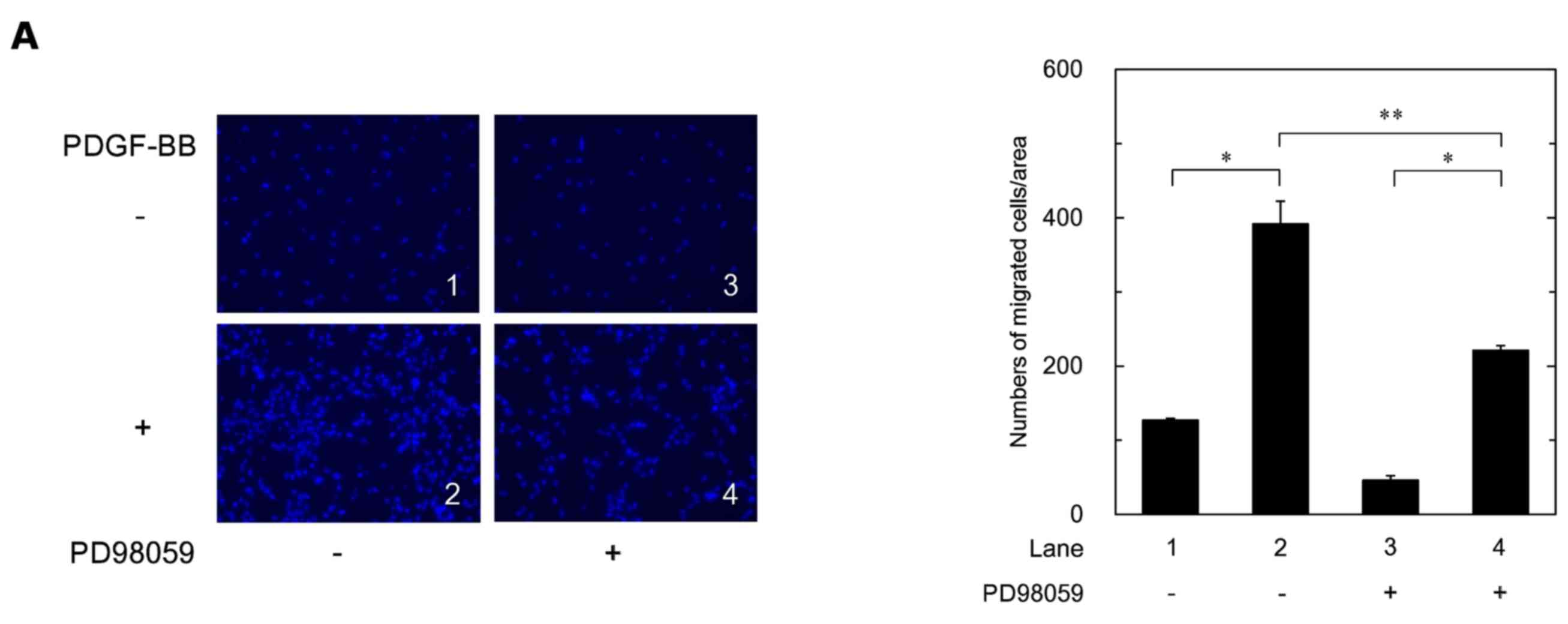
Effects of PD98059, SB203580 or SP600125
on the PDGF-BB-stimulated migration of MC3T3-E1 cells
Regarding the intracellular signaling of PDGF-BB in
osteoblasts, we previously demonstrated that p44/p42 MAPK, p38 MAPK
and SAPK/JNK function as positive regulators in the
PDGF-BB-stimulated IL-6 synthesis in osteoblast-like MC3T3-E1
cells, whereas the PI3K/Akt and p70 S6 kinase pathways negatively
regulate the IL-6 synthesis (7,8).
In order to investigate the roles of three MAPKs in the
PDGF-BB-stimulated cell migration, we examined the effects of
PD98059, an inhibitor of the upstream kinase activating p44/p42
MAPK (MEK1/2) (32), SB203580, an
inhibitor of p38 MAPK (33), or
SP600125, an inhibitor of SAPK/JNK (34), on the cell migration. PD98059,
SB203580 and SP600125 significantly reduced the PDGF-BB-stimulated
migration of the MC3T3-E1 cells (Fig.
4A–C).
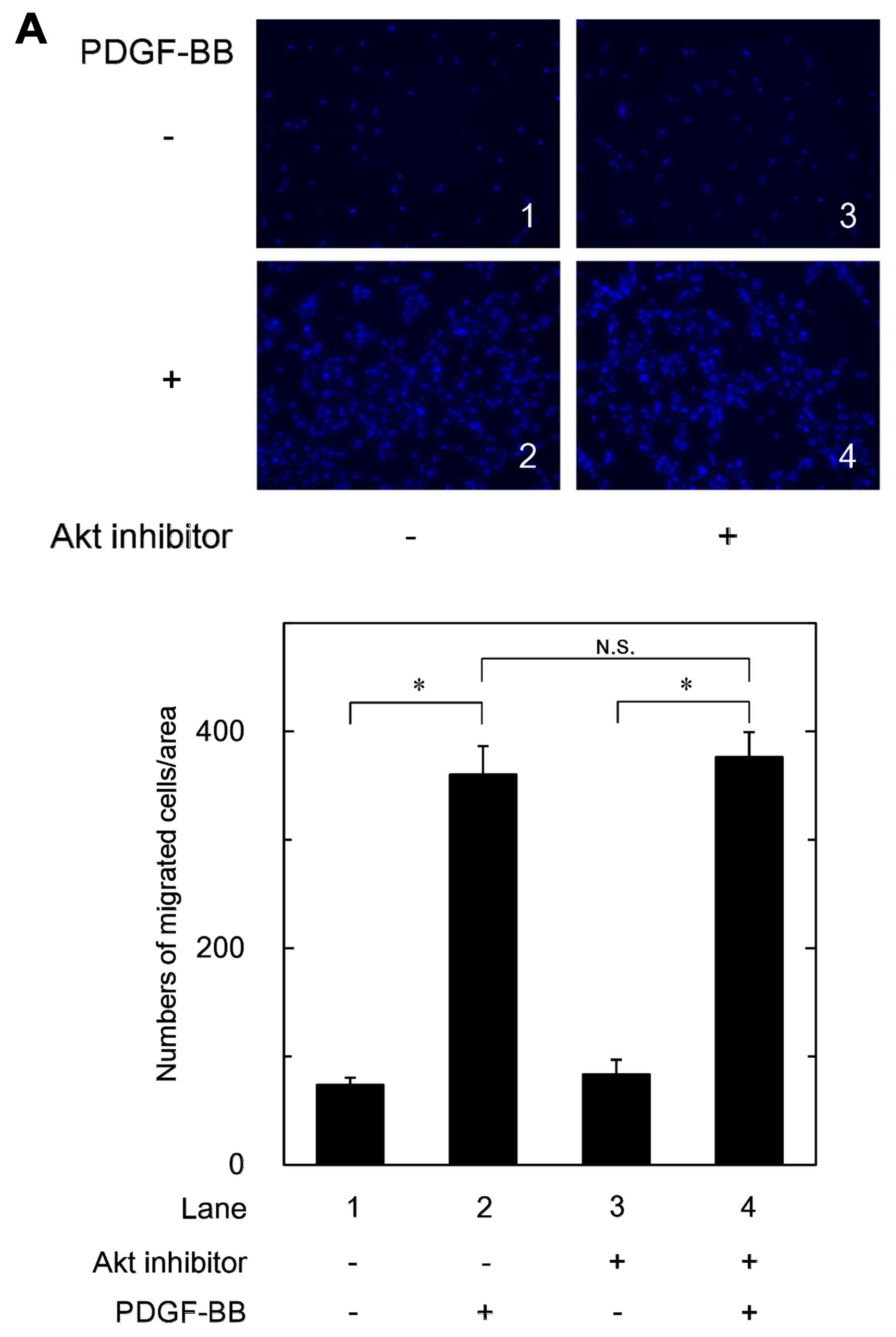
Effects of Akt inhibitor or rapamycin on
the PDGF-BB-stimulated migration of MC3T3-E1 cells
In order to clarify the role of PI3K/Akt or p70 S6
kinase in the PDGF-BB-stimulated migration of MC3T3-E1 cells, we
next examined the effects of Akt inhibitor,
1L-6-hydroxymethyl-chiro-inositol
2-(R)-2-O-methyl-3-O-octadecylcarbonate
(35) and rapamycin, an inhibitor
of upstream kinase of p70 S6 kinase (mTOR) (36), on the cell migration. The Akt
inhibitor failed to affect the PDGF-BB-stimulated migration of
MC3T3-E1 cells (Fig. 5A). In
addition, we found that deguelin, another Akt inhibitor (37), did not suppress the
PDGF-BB-induced cell migration (data not shown). On the other hand,
rapamycin barely affected the PDGF-BB-stimulated migration of
MC3T3-E1 cells (Fig. 5B).
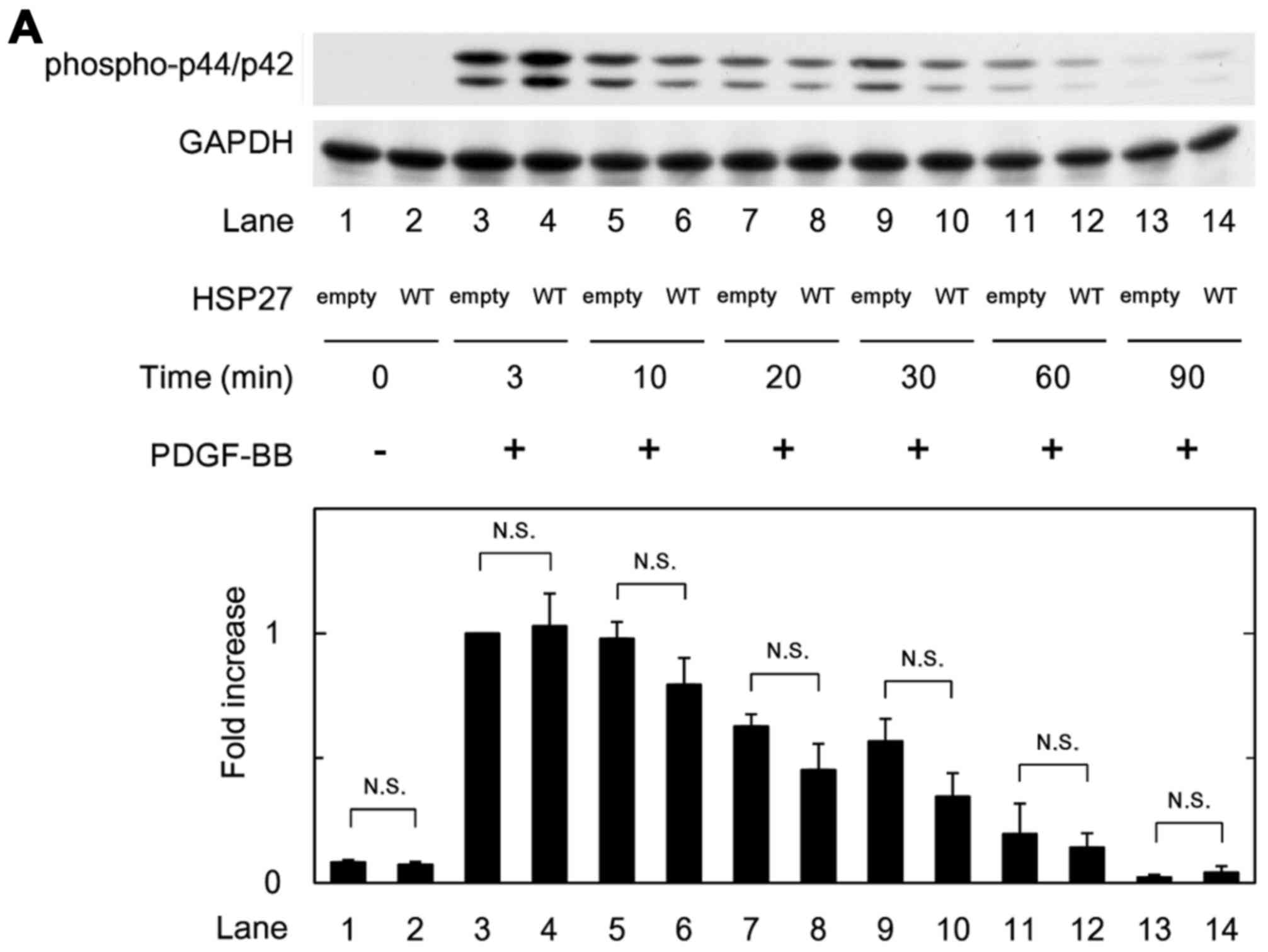
Effects of HSP27 overexpression on the
PDGF-BB-induced phosphorylation of p44/p42 MAPK, p38 MAPK or
SAPK/JNK in MC3T3-E1 cells
In order to clarify whether HSP27 affects
PDGF-BB-stimulated MC3T3-E1 cell migration through the activation
of p44/p42 MAPK, p38 MAPK or SAPK/JNK, we examined the effect of
PDGF-BB on the phosphorylation of p44/p42 MAPK, p38 MAPK or
SAPK/JNK in the WT HSP27 vector-transfected MC3T3-E1 cells and the
control empty vector-transfected cells. However, there were no
significant differences in the phosphorylated levels of p44/p42
MAPK, p38 MAPK or SAPK/JNK between the WT HSP27-overexpressing
cells and the control cells (Fig.
6A–C).
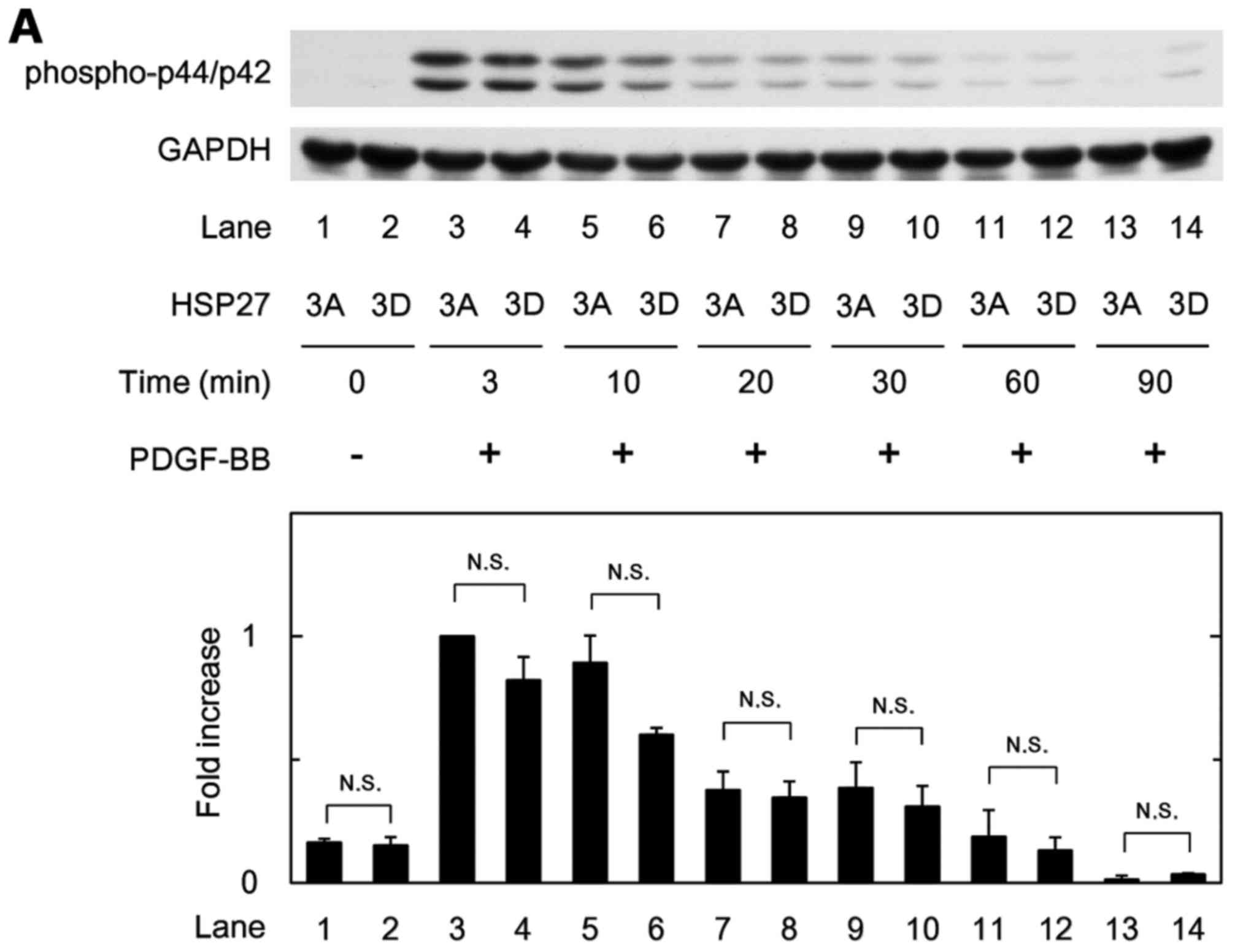
We further examined the effect of PDGF-BB on the
phosphorylation of p44/p42 MAPK, p38 MAPK or SAPK/JNK in the 3D and
3A cells. There were no significant differences in their
phosphorylated levels between the 3A and 3D cells (Fig. 7A–C).
Discussion
We conducted the present study to clarify whether
HSP27 (HSPB1), one of the low-molecular-weight HSPs, is implicated
in the migration of osteoblast-like MC3T3-E1 cells. Since PDGF-BB,
a potent mitogen of connective tissue cells (1), reportedly induces migration of human
osteoblasts, we confirmed that PDGF-BB truly stimulated the
migration of osteoblast-like MC3T3-E1 cells. Although HSP27 is
ubiquitously expressed in human cells and tissues such as skeletal,
smooth and cardiac muscles, we herein confirmed that the expression
levels of HSP27 without stimulation are quite low in MC3T3-E1 cells
as we previously reported (16).
We also found that PDGF-BB barely induced the protein expression
levels of HSP27 in contrast to TGF-β. We previously established WT
HSP27-overexpressing MC3T3-E1 cells (22,31). In the present study, we
investigated the role of HSP27 in the PDGF-BB-induced osteoblast
cell migration using the mutant cells. The PDGF-BB-stimulated
migrated cell numbers in the HSP27-overexpressing cells were
markedly less than those in the control cells. We previously
demonstrated that HSP27 in the WT HSP27-transfected MC3T3-E1 cells
was not phosphorylated (22).
However, we found here that PDGF-BB induced the phosphorylation of
HSP27 in the WT HSP27-overexpressing MC3T3-E1 cells. Based on our
findings, it is probable that HSP27 plays a suppressive role in the
PDGF-BB-induced migration of osteoblast-like MC3T3-E1 cells, and
that HSP27 in the WT HSP27-expressing cells could be phosphorylated
at least in part stimulated by PDGF-BB.
It is currently established that the functions of
HSP27 are regulated by post-translational modifications including
phosphorylation (10). Regarding
the phosphorylation of HSP27, unphosphorylated HSP27 forms
aggregated multimers while the phosphorylation of HSP27 results in
conformational changes, such as dimers (12,13). During the processes of cell
migration, phosphorylated HSP27 reportedly regulates actin filament
dynamics in cytoskeleton organization (38,39). It is likely that the
phosphorylation of HSP27 stimulated by PDGF-BB itself could affect
the PDGF-BB-induced migration of HSP27-expressing osteoblasts.
Thus, we established two mutant HSP27-transfected osteoblast-like
MC3T3-E1 cell lines, 3A and 3D cells, which transiently express the
unphosphorylated state and the phospho-mimic state of HSP27,
respectively (31), and
investigated the effect of HSP27 phosphorylation on the
PDGF-BB-induced migration of MC3T3-E1 cells. We demonstrated that
the PDGF-BB-stimulated migrated cell numbers of the 3D cells were
markedly decreased compared with those of the 3A cells. We also
showed that the PDGF-BB-induced migrated cell numbers of the WT
HSP27-overexpressing cells were less than those of the 3A cells. As
HSP27 in the WT HSP27-overexpressing cells could be phosphorylated
at least in part stimulated by PDGF-BB, our findings suggest that
the suppressive effect by HSP27 in its phosphorylated form of the
PDGF-BB-stimulated cell migration is greater than in its
unphosphorylated form. HSP27 oligomerization is mainly regulated by
Ser-78 and/or Ser-82 phosphorylation (13). Therefore, it is probable that we
could cover major relevant phosphorylation sites while making 3A
and 3D mutants. Based on these findings, it is possible that the
conformational change of HSP27 by its phosphorylation affects
PDGF-BB-stimulated migration of osteoblast-like MC3T3-E1 cells.
We previously reported that PDGF-BB stimulates IL-6
synthesis at least in part via p44/p42 MAPK, p38 MAPK and SAPK/JNK
in osteoblast-like MC3T3-E1 cells (7). Thus, in order to clarify the
involvement of these three MAPKs in the PDGF-BB-induced MC3T3-E1
cell migration, we examined the effects of PD98059 (32), SB203580 (33) or SP600125 (34) on the migration. PD98059, SB203580
and SP600125 significantly reduced the PDGF-BB-stimulated
migration. Although it is possible that some pharmacological
effects other than the specificity may be involved in the
suppression, our findings suggest that PDGF-BB induced the
migration of osteoblast-like MC3T3-E1 cells through the activation
of p44/p42 MAPK, p38 MAPK and SAPK/JNK. On the other hand, in our
previous studies (7,8) we demonstrated that PI3K/Akt and p70
S6 kinase limit the PDGF-BB-stimulated IL-6 synthesis in
osteoblast-like MC3T3-E1 cells. In order to investigate whether
PI3K/Akt or p70 S6 kinase is involved in the PDGF-BB-induced
MC3T3-E1 cell migration, we examined the effects of Akt inhibitor,
1L-6-hydroxymethyl-chiro-inositol
2-(R)-2-O-methyl-3-O-octadecylcarbonate
(35) or rapamycin (36) on the migration. The Akt inhibitor
or rapamycin failed to affect the PDGF-BB-stimulated migration of
these cells. Thus, it seems unlikely that either PI3K/Akt or p70 S6
kinase regulates the migration stimulated by PDGF-BB in MC3T3-E1
cells. Taking our findings into account, it is most likely that
p44/p42 MAPK, p38 MAPK and SAPK/JNK, but not PI3K/Akt or p70 S6
kinase, act as positive regulators in the PDGF-BB-induced migration
of osteoblast-like MC3T3-E1 cells.
In order to further investigate the relationship
between HSP27 and PDGF signaling in osteoblast-like MC3T3-E1 cells,
we examined the effects of PDGF-BB on the phosphorylation of
p44/p42 MAPK, p38 MAPK or SAPK/JNK in the WT HSP27
vector-transfected cells compared with those in the control empty
vector-transfected cells. However, we did not observe any
significant differences in their phosphorylation levels between the
WT HSP27-overexpressing cells and the control cells. In addition,
we found that there were no significant differences between the 3A
and 3D cells in the PDGF-BB-induced phosphorylation levels of
p44/p42 MAPK, p38 MAPK or SAPK/JNK. Taking our findings into
account, it seems unlikely that HSP27, regardless of its
phosphorylation, regulates PDGF-BB-induced osteoblast-like MC3T3-E1
cell migration via activation of p44/p42 MAPK, p38 MAPK or
SAPK/JNK. It is possible that HSP27 may act at a point downstream
of these molecules or another target, resulting in the
downregulation of osteoblast-like MC3T3-E1 cell migration. Our
present findings, showing that HSP27 could function as a negative
regulator in the PDGF-BB-stimulated migration of osteoblasts, and
phosphorylated HSP27 enhances the suppression, provide a new
insight concerning small molecular HSP as a bone remodeling
modulator. It has been reported that other migratory mediators
downstream of PI3K such as neuronal Wiskott-Aldrich syndrome
protein, Rac and RhoA are involved in the migration induced by PDGF
in NIH-3T3 fibroblasts (40).
Therefore, it is possible that the mediators described above are
involved in the suppression by HSP27 of the PDGF-BB-induced
migration in osteoblast-like MC3T3-E1 cells. The Boyden chamber
assay adopted here does not distinguish between cells that lose
their chemoattraction and those that are no longer changing to a
migratory phenotype or dysregulation in adherence. Further
investigation including cell adhesion, lamellipodia or filopodia
formation would be required to clarify the detailed mechanism
underlying the suppressive effect of HSP27 on the migration of
osteoblasts.
In conclusion, our results strongly suggest that
HSP27 functions as a negative regulator in the PDGF-BB-stimulated
migration of osteoblasts and the suppressive effect is amplified by
the phosphorylation state of HSP27.
Acknowledgments
We thank Dr. C. Schafer (Klinikum Grosshadern,
Ludwig Maximilians University, Munich, Germany) for providing the
mutant HSP27 cDNA and Mrs. Yumiko Kurokawa for her skillful
technical assistance. This investigation was supported in part by a
Grant-in-Aid for Scientific Research (19591042) from the Ministry
of Education, Culture, Sports, Science and Technology of Japan, a
Grant-in-Aid for Scientific Research (H25-Aging-General-004) from
the Ministry of Health, Labor and Welfare of Japan, and the
Research Funding for Longevity Sciences (25-4, 26-12) from the
National Center for Geriatrics and Gerontology (NCGG), Japan.
References
|
1
|
Heldin CH and Westermark B: Mechanism of
action and in vivo role of platelet-derived growth factor. Physiol
Rev. 79:1283–1316. 1999.PubMed/NCBI
|
|
2
|
Wang Z, Ahmad A, Li Y, Kong D, Azmi AS,
Banerjee S and Sarkar FH: Emerging roles of PDGF-D signaling
pathway in tumor development and progression. Biochim Biophys Acta.
1806:122–130. 2010.PubMed/NCBI
|
|
3
|
Kular J, Tickner J, Chim SM and Xu J: An
overview of the regulation of bone remodelling at the cellular
level. Clin Biochem. 45:863–873. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Canalis E: Growth factor control of bone
mass. J Cell Biochem. 108:769–777. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Caplan AI and Correa D: PDGF in bone
formation and regeneration: new insights into a novel mechanism
involving MSCs. J Orthop Res. 29:1795–1803. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Hollinger JO, Hart CE, Hirsch SN, Lynch S
and Friedlaender GE: Recombinant human platelet-derived growth
factor: biology and clinical applications. J Bone Joint Surg Am.
90(Suppl 1): 48–54. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Takai S, Tokuda H, Hanai Y and Kozawa O:
Limitation by p70 S6 kinase of platelet-derived growth
factor-BB-induced interleukin 6 synthesis in osteoblast-like
MC3T3-E1 cells. Metabolism. 56:476–483. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Hanai Y, Tokuda H, Ohta T,
Matsushima-Nishiwaki R, Takai S and Kozawa O: Phosphatidylinositol
3-kinase/Akt auto-regulates PDGF-BB-stimulated interleukin-6
synthesis in osteoblasts. J Cell Biochem. 99:1564–1571. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Hengartner NE, Fiedler J, Ignatius A and
Brenner RE: IL-1β inhibits human osteoblast migration. Mol Med.
19:36–42. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Mymrikov EV, Seit-Nebi AS and Gusev NB:
Large potentials of small heat shock proteins. Physiol Rev.
91:1123–1159. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bakthisaran R, Tangirala R and Rao ChM:
Small heat shock proteins: role in cellular functions and
pathology. Biochim Biophys Acta. 1854:291–319. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Kostenko S and Moens U: Heat shock protein
27 phosphorylation: kinases, phosphatases, functions and pathology.
Cell Mol Life Sci. 66:3289–3307. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Katsogiannou M, Andrieu C and Rocchi P:
Heat shock protein 27 phosphorylation state is associated with
cancer progression. Front Genet. 5:3462014. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Leonardi R, Barbato E, Paganelli C and Lo
Muzio L: Immunolocalization of heat shock protein 27 in developing
jaw bones and tooth germs of human fetuses. Calcif Tissue Int.
75:509–516. 2004. View Article : Google Scholar
|
|
15
|
Shakoori AR, Oberdorf AM, Owen TA, Weber
LA, Hickey E, Stein JL, Lian JB and Stein GS: Expression of heat
shock genes during differentiation of mammalian osteoblasts and
promyelocytic leukemia cells. J Cell Biochem. 48:277–287. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Kozawa O, Niwa M, Matsuno H, Ishisaki A,
Kato K and Uematsu T: Stimulatory effect of basic fibroblast growth
factor on induction of heat shock protein 27 in osteoblasts: role
of protein kinase C. Arch Biochem Biophys. 388:237–242. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kozawa O, Otsuka T, Hatakeyama D, Niwa M,
Matsuno H, Ito H, Kato K, Matsui N and Uematsu T: Mechanism of
prostaglandin D2-stimulated heat shock protein 27
induction in osteoblasts. Cell Signal. 13:535–541. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Hatakeyama D, Kozawa O, Niwa M, Matsuno H,
Kato K, Tatematsu N, Shibata T and Uematsu T: Inhibition by
adenylyl cyclase-cAMP system of ET-1-induced HSP27 in osteoblasts.
Am J Physiol Endocrinol Metab. 281:E1260–E1266. 2001.PubMed/NCBI
|
|
19
|
Tokuda H, Kozawa O, Niwa M, Matsuno H,
Kato K and Uematsu T: Mechanism of prostaglandin
E2-stimulated heat shock protein 27 induction in
osteoblast-like MC3T3- E1 cells. J Endocrinol. 172:271–281. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Tokuda H, Niwa M, Ishisaki A, Nakajima K,
Ito H, Kato K and Kozawa O: Involvement of stress-activated protein
kinase (SAPK)/c-Jun N-terminal kinase (JNK) in prostaglandin
F2α-induced heat shock protein 27 in osteoblasts.
Prostaglandins Leukot Essent Fatty Acids. 70:441–447. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Hayashi K, Takai S, Matsushima-Nishiwaki
R, Hanai Y, Kato K, Tokuda H and Kozawa O: (−)-Epigallocatechin
gallate reduces transforming growth factor β-stimulated HSP27
induction through the suppression of stress-activated protein
kinase/c-Jun N-terminal kinase in osteoblasts. Life Sci.
82:1012–1017. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Kato K, Adachi S, Matsushima-Nishiwaki R,
Minamitani C, Natsume H, Katagiri Y, Hirose Y, Mizutani J, Tokuda
H, Kozawa O, et al: Regulation by heat shock protein 27 of
osteocalcin synthesis in osteoblasts. Endocrinology. 152:1872–1882.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kubisch C, Dimagno MJ, Tietz AB, Welsh MJ,
Ernst SA, Brandt-Nedelev B, Diebold J, Wagner AC, Göke B, Williams
JA, et al: Overexpression of heat shock protein Hsp27 protects
against cerulein-induced pancreatitis. Gastroenterology.
127:275–286. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Sudo H, Kodama HA, Amagai Y, Yamamoto S
and Kasai S: In vitro differentiation and calcification in a new
clonal osteogenic cell line derived from newborn mouse calvaria. J
Cell Biol. 96:191–198. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kozawa O, Tokuda H, Miwa M, Kotoyori J and
Oiso Y: Cross-talk regulation between cyclic AMP production and
phosphoinositide hydrolysis induced by prostaglandin E2
in osteoblast-like cells. Exp Cell Res. 198:130–134. 1992.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Karagiosis SA, Chrisler WB, Bollinger N
and Karin NJ: Lysophosphatidic acid-induced ERK activation and
chemotaxis in MC3T3-E1 preosteoblasts are independent of EGF
receptor transactivation. J Cell Physiol. 219:716–723. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Laemmli UK: Cleavage of structural
proteins during the assembly of the head of bacteriophage T4.
Nature. 227:680–685. 1970. View
Article : Google Scholar : PubMed/NCBI
|
|
28
|
Kato K, Ito H, Hasegawa K, Inaguma Y,
Kozawa O and Asano T: Modulation of the stress-induced synthesis of
hsp27 and αB-crystallin by cyclic AMP in C6 rat glioma cells. J
Neurochem. 66:946–950. 1996. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Kozawa O, Niwa M, Matsuno H, Tokuda H,
Miwa M, Ito H, Kato K and Uematsu T: Sphingosine 1-phosphate
induces heat shock protein 27 via p38 mitogen-activated protein
kinase activation in osteoblasts. J Bone Miner Res. 14:1761–1767.
1999. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hatakeyama D, Kozawa O, Niwa M, Matsuno H,
Ito H, Kato K, Tatematsu N, Shibata T and Uematsu T: Upregulation
by retinoic acid of transforming growth factor-β-stimulated heat
shock protein 27 induction in osteoblasts: involvement of
mitogen-activated protein kinases. Biochim Biophys Acta.
1589:15–30. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Kuroyanagi G, Tokuda H, Yamamoto N,
Matsushima-Nishiwaki R, Kozawa O and Otsuka T: Unphosphorylated
HSP27 (HSPB1) regulates the translation initiation process via a
direct association with eIF4E in osteoblasts. Int J Mol Med.
36:881–889. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Alessi DR, Cuenda A, Cohen P, Dudley DT
and Saltiel AR: PD 098059 is a specific inhibitor of the activation
of mitogen-activated protein kinase kinase in vitro and in vivo. J
Biol Chem. 270:27489–27494. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Cuenda A, Rouse J, Doza YN, Meier R, Cohen
P, Gallagher TF, Young PR and Lee JC: SB 203580 is a specific
inhibitor of a MAP kinase homologue which is stimulated by cellular
stresses and interleukin-1. FEBS Lett. 364:229–233. 1995.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Bennett BL, Sasaki DT, Murray BW, O'Leary
EC, Sakata ST, Xu W, Leisten JC, Motiwala A, Pierce S, Satoh Y, et
al: SP600125, an anthrapyrazolone inhibitor of Jun N-terminal
kinase. Proc Natl Acad Sci USA. 98:13681–13686. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hu Y, Qiao L, Wang S, Rong SB, Meuillet
EJ, Berggren M, Gallegos A, Powis G and Kozikowski AP:
3-(Hydroxymethyl)-bearing phosphatidylinositol ether lipid
analogues and carbonate surrogates block PI3-K, Akt, and cancer
cell growth. J Med Chem. 43:3045–3051. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Price DJ, Grove JR, Calvo V, Avruch J and
Bierer BE: Rapamycin-induced inhibition of the 70-kilodalton S6
protein kinase. Science. 257:973–977. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Chun KH, Kosmeder JW II, Sun S, Pezzuto
JM, Lotan R, Hong WK and Lee HY: Effects of deguelin on the
phosphatidylinositol 3-kinase/Akt pathway and apoptosis in
premalignant human bronchial epithelial cells. J Natl Cancer Inst.
95:291–302. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Clarke JP and Mearow KM: Cell stress
promotes the association of phosphorylated HspB1 with F-actin. PLoS
One. 8:e689782013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Lavoie JN, Hickey E, Weber LA and Landry
J: Modulation of actin microfilament dynamics and fluid phase
pinocytosis by phosphorylation of heat shock protein 27. J Biol
Chem. 268:24210–24214. 1993.PubMed/NCBI
|
|
40
|
Jiménez C, Portela RA, Mellado M,
Rodríguez-Frade JM, Collard J, Serrano A, Martínez-A C, Avila J and
Carrera AC: Role of the PI3K regulatory subunit in the control of
actin organization and cell migration. J Cell Biol. 151:249–262.
2000. View Article : Google Scholar : PubMed/NCBI
|