Introduction
Innate immune signaling receptors play a pivotal
role in defense against invasion by pathogenic microorganisms or
tissue damage (1). Toll-like
receptors (TLRs), as one of the important innate immune signaling
receptors, initiate complicated signaling pathways leading to acute
but transient inflammatory responses aimed at the clearance of
pathogens and cellular debris (2). TLR3 and TLR4 recruit
TIR-domain-containing adapter-inducing interferon-β (TRIF) and̸or
MyD88, leading to the expression of proinflammatory cytokines and
the induction of interferon (IFN)-β (2,3).
Although proinflammatory cytokines and IFN-β are essential for
resistance against invading pathogens, uncontrolled TLR3/4
activation by poly(I:C)/lipopolysaccharide (LPS) and production of
proinflammatory cytokines, such as tumor necrosis factor (TNF)-α,
interleukin (IL)-6 and IFN-β are the main cause of septic shock
(4). Therefore, elucidating the
mechanisms underlying the production of these cytokines is crucial
for developing treatments for such conditions.
The cytoskeleton plays an important role in
modulating cell morphology, migration and division through
contractile ring formation and certain intracellular signaling
pathways (5). Recent studies have
provided evidence supporting the importance of the cytoskeleton in
immunocytes, including macrophage-, B-cell- and dendritic
cell-mediated inflammation (6–8).
poly(I:C) and LPS are two TLR3/4 agonists commonly used to induce
macrophage activation and investigate the effect of other signaling
molecules in cellular responses (3). However, the role of the cytoskeleton
in the TLR3/4 signaling pathway inducing macrophage activation has
not been clearly determined.
Vincristine (VCR) is a vinca alkaloid extracted from
the plant Catharanthus roseus (9), which has long been used as a
chemotherapeutic agent for the treatment of childhood and adult
acute lymphocytic leukemia, Hodgkin's and non-Hodgkin's lymphoma,
and various solid tumors, including germ cell tumors, small-cell
lung cancer, Ewing's sarcoma, neuroblastoma, breast cancer,
melanoma and multiple myeloma (10,11). VCR may lead to microtubule
depolymerization via binding to the tubulin protein in a
dose-dependent manner, similar to colchicine (12). However, the effect of VCR on
macrophage activation by poly(I:C) and LPS has not been fully
elucidated.
The aim of the present study was to demonstrate that
macrophage activation, leading to cytokine production and changes
in cell morphology, differs significantly between LPS and
poly(I:C). In addition, tubulin expression was decreased following
LPS stimulation. By contrast, tubulin expression was somewhat
increased following poly(I:C) treatment. There was no significant
effect on the expression of β-actin and glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) in LPS- or poly(I:C)-induced macrophages.
Furthermore, it was observed that VCR pretreatment represses the
cytoskeleton rearrangement in macrophages and reduces the
production of proinflammatory cytokines and cell migration caused
by LPS. In addition, it was demonstrated that the mitogen-activated
protein kinase (MAPK)/p38 signaling pathway plays a pivotal role in
cytoskeleton rearrangement leading to cytokine production and
macrophage invasion induced by LPS.
Materials and methods
Reagents and preparation
LPS (Escherichia coli, 055:B5) was obtained
from Sigma-Aldrich (Merck KGaA, St. Louis, MO, USA; cat. no.
L2880). poly(I:C) was obtained from InvivoGen (San Diego, CA, USA;
cat. no. tlrl-picw). VCR was purchased from Dalian Meilun Biotech
Co., Ltd. (Dalian, Liaoning, China; cat. no. 2068-78-2). Mouse
monoclonal antibodies to GAPDH (cat. no. 60004-1-lg), β-actin (cat.
no. 60008-1-lg), α-tubulin (cat. no. 66031-1-lg) and β-tubulin
(cat. no. 66240-1-lg) were purchased from Proteintech Group, Inc.
(Rosemont, IL, USA). LPS and poly(I:C) were dissolved and diluted
with corresponding liquid according to the manufacturer's
instructions. The VCR was stored as powder at room temperature. For
cytology experiments, VCR was dissolved in a stock solution of
sterile deionized H2O to a concentration of 1 mg/ml. For
further experiments, stock solution was diluted in sterile
deionized with H2O or Dulbecco's modified Eagle's medium
(DMEM; Gibco-BRL; Thermo Fisher Scientific, Grand Island, NY, USA)
containing 10% fetal bovine serum (FBS; Gibco® Sera;
Thermo Fisher Scientific, Newcastle, Australia) to a final
concentration working solution of 10, 20, 50, 100, 300, 500 and
1,000 ng/µl.
Mice and cells
Male C57BL/6j mice, 6–8 weeks old, were obtained
from the Animal Research Committee of the Institute of Biology and
Cell Biology (Shanghai, China) and housed in a specific
pathogen-free environment. The animal room was kept at 20–22°C
under a 12-h light/dark cycle. All animal experiments were
conducted in accordance with the National Institutes of Health
Guide for the Care and use of Laboratory Animals, with the approval
of the Scientific Investigation Board of the Medical School of
Shandong University (Jinan, China). Mouse primary peritoneal
macrophages were prepared as previously described (3). The mouse macrophage cell line
RAW264.7 was obtained from American Type Culture Collection
(Manassas, VA, USA) and cultured in DMEM containing 10% (v/v) FBS,
100 U/ml penicillin and 100 µg/ml streptomycin (Gibco-BRL;
Thermo Fisher Scientific, Grand Island, NY, USA). The cell lines
were maintained at 37°C in a humidified incubator with 5%
CO2. RAW264.7 cells were stimulated with 100 ng/ml LPS
or 20 µg/ml poly(I:C) for different times, with or without
pretreatment with VCR or MAPK pathway inhibitors.
RNA extraction and quantification
Total RNA was extracted using TRIzol reagent
(Invitrogen; Thermo Fisher Scientific, Carlsbad, CA, USA) according
to the manufacturer's instructions, and reverse transcription was
performed using a Takara reverse transcription kit (Takara, Shiga,
Japan). The expression of IL-6, TNF-α and IFN-β was quantified
using SYBR Premix Ex Tap™, with GAPDH as an internal normalized
reference. The specific sequences of the primers used were as
previously described (3,13). Quantitative polymerase chain
reaction (qPCR) was performed under the following conditions: 95°C
for 30 sec, followed by 45 cycles at 95°C for 5 sec, 60°C for 5
sec, 72°C for 5 sec and 65°C for 20 sec, using the LightCycler
Real-time PCR system (Roche Diagnostics, Indianapolis, IN, USA) as
previously described (14).
Enzyme-linked immunosorbent assay
(ELISA)
The cell culture supernatants were collected, and
the concentration of IL-6 (cat. no. KMC0061) and TNF-α (cat. no.
KMC3011) were measured using a commercially available ELISA kit
(all from Invitrogen; Thermo Fisher Scientific), in accordance with
the manufacturer's instructions.
Boyden chamber assays
The Boyden chamber was obtained from Corning, Inc.
(Corning, NY, USA; cat. no. 3422). DMEM (600 µl) with 10%
FBS was added to the lower chamber. Next, RAW264.7 cells were
collected and washed with DMEM. The cells were resuspended in 200
µl of DMEM with/without VCR and/or LPS, and then seeded to
the upper chamber at a concentration of 2×105
cells/well. Following incubation for 20 h at 37°C in a humidified
incubator with 5% CO2, the Boyden chamber was fixed with
4% paraformaldehyde. Non-migrated cells on the upper surface of the
filter were removed with a cotton swab and the cells that traversed
and spread on the lower surface of the membrane were stained with
hematoxylin. The filter was dehydrated with gradient alcohol and
dried at room temperature. These membranes were sealed using
neutral gum and cell invasiveness was observed under a light
microscope as previously described (14).
Cell counting kit-8 (CCK-8) assay
Cell proliferation was evaluated using CCK-8
(Dojindo Molecular Technologies, Inc., Kumamoto, Japan). Cells were
seeded at a density of 3×104/well in 96-well plates and
were then treated with VCR at different concentrations. After 0, 2,
4, 8, 12, 24, 36 and 48 h, 20 µl of CCK-8 solution was added
to each well and incubated at 37°C for 1 h. At the end of the
incubation, the optical density was read at 450 nm using a
microplate reader (Thermo Fisher Scientific, Waltham, MA, USA). The
mean values were determined from different wells as previously
described (14).
Confocal microscopy analysis
RAW264.7 cells were grown on coverslips and
stimulated with 100 ng/ml LPS or 20 µg/ml poly(I:C) for 24 h
at 37°C, with or without VCR pretreatment. Non-adherent cells were
then washed away with phosphate-buffered saline (PBS). The cells
were fixed in 4% paraformaldehyde for 15 min and permeabilized with
0.1% Triton X-100. After blocking with 10% bovine serum albumin
(BSA) in PBS for 2 h at room temperature, they were incubated with
primary antibodies against α-tubulin, β-tubulin and phalloidin-FITC
(50 µg/ml; cat. no. P5282; Sigma Aldrich; Merck KGaA) for 1
h at room temperature, followed by washing 3 times with PBS for 5
min/time. Cells were incubated with secondary antibodies (cat. no.
CA11005s; Invitrogen; Thermo Fisher Scientific) for 1 h at room
temperature in the dark, except for the phalloidin-FITC staining
groups, followed by washing 3 times with PBS for 5 min/time. Cell
nuclei were stained with DIPA for 8 min and washed 3 times with PBS
for 5 min/time. After sealing the coverslips with neutral gum,
images were captured using a high sensitivity laser scanning
confocal microscope (LSM780; Zeiss, Jena, Germany) with the
appropriate filters and laser (488, 561 and 633 nm) and a ×63
objective lens.
Simple western assays
Cells were lysed with the CelLytic™ Cell Lysis
Reagent (Sigma Aldrich; Merck KGaA) supplemented with a protease
inhibitor cocktail; the protein concentrations were measured on a
Multiskan GO microplate reader (Thermo Fisher Scientific) using the
bicinchoninic acid assay and BSA standards (Pierce, Rockford, IL,
USA), and the volumes were then made equal using the extraction
reagent. Automated capillary western blot analyses were performed
according to the ProteinSimple user manual provided by the
manufacturer. In brief, cell lysate samples were diluted with 1X
sample buffer to 0.2 mg/ml; the diluted samples were then mixed at
1:4 ratio with 5X master mix (ProteinSimple) containing 1X
fluorescent molecular weight markers, sodium dodecyl sulfate and
dithiothreitol, and were then incubated at 98°C for 10 min for
protein denaturation. The biotinylated ladder, treated samples,
blocking buffer, primary antibodies, horseradish
peroxidase-conjugated secondary antibodies, luminol-peroxidase 1:1
mix and wash buffer were dispensed to designated assay plate with
13-Capillary Cartridge (PS-CC02). Chemiluminescence was detected at
5, 15, 30, 60, 120, 240 and 480 sec. All the primary antibodies
were used at 1:50 dilution for simple western blot analysis.
Statistical analysis
All data are presented as the result of three or
four independent experiments. All data are expressed as mean ±
standard deviation, and analyzed via one-way analysis of variance
and two-tailed Student's t-test using SPSS 16.0 statistical
software (SPSS, Inc., Chicago, IL, USA). In all cases, P-values
<0.05 were considered to indicate statistically significant
differences.
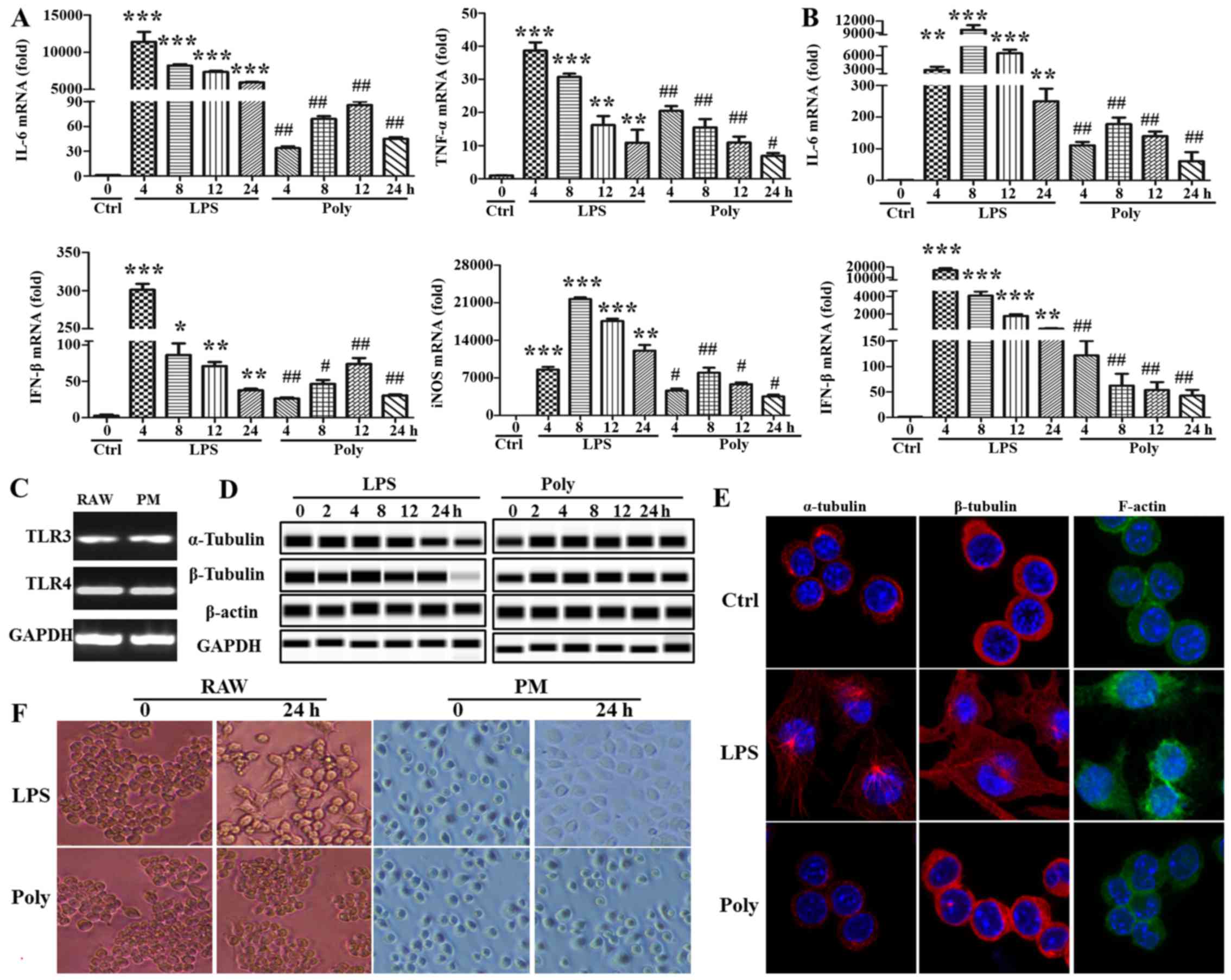
Results
LPS stimulation, but not poly(I:C)
stimulation, leads to changes in macrophage morphology
In order to investigate the possibly different
mechanism of TLR3/4-induced macrophage activation, the mouse
macrophage cell line RAW264.7 and mouse primary peritoneal
macrophages were stimulated with LPS or poly(I:C) at different
timepoints. First, the activating effect of LPS or poly(I:C) was
examined by PCR using IL-6 and IFN-β, well-known downstream
molecules in the signaling pathway of TLR3/4. As shown in Fig. 1A and B, the synthesis of IL-6 and
IFN-β was markedly increased with LPS or poly(I:C) stimulation. The
expression of other cytokines, such as TNF-α and inducible nitric
oxide synthase, was also upregulated (Fig. 1A). However, surprisingly, the
expression of the abovementioned cytokines induced by poly(I:C)
stimulation was always lower compared with that of the LPS group at
any given timepoint. Moreover, TLR3 and TLR4 expression was
detected with the PCR assay, and TLR3 and TLR4 were found to be
expressed in RAW264.7 cells and mouse primary peritoneal
macrophages, although the level of TLR3 was lower compared with
that of TLR4 (Fig. 1C). In
addition, there was a major distinction between the two groups
regarding cell morphology. As shown in Fig. 1D, the cells stimulated by LPS were
more stretched and developing multiple pseudopodia or flattened. In
sharp contrast, poly(I:C)-induced cells maintained their round
morphology, without marked changes compared with the unstimulated
group. Subsequently, cytoskeleton rearrangement following LPS or
poly(I:C) treatment was observed using a confocal microscope. As
shown in Fig. 1E, the
cytoskeleton was found to be markedly rearranged via staining of
the α- and β-tubulin and F-actin. Finally, the expression of
cytoskeletal proteins (α- and β-tubulin and β-actin) and
non-cytoskeletal proteins (GAPDH) was evaluated following LPS or
poly(I:C) stimulation using simple western assays. Surprisingly,
LPS stimulation significantly reduced α- and β-tubulin expression,
but not that of β-actin and GAPDH. By contrast, α- and β-tubulin
expression were mildly increased, whereas there was no significant
effect on the expression of β-actin and GAPDH following poly(I:C)
stimulation (Fig. 1F). Taken
together, these data indicate that cytokine and cytoskeletal
protein expression and cell morphology differed significantly
following TLR3/4-induced macrophage activation.
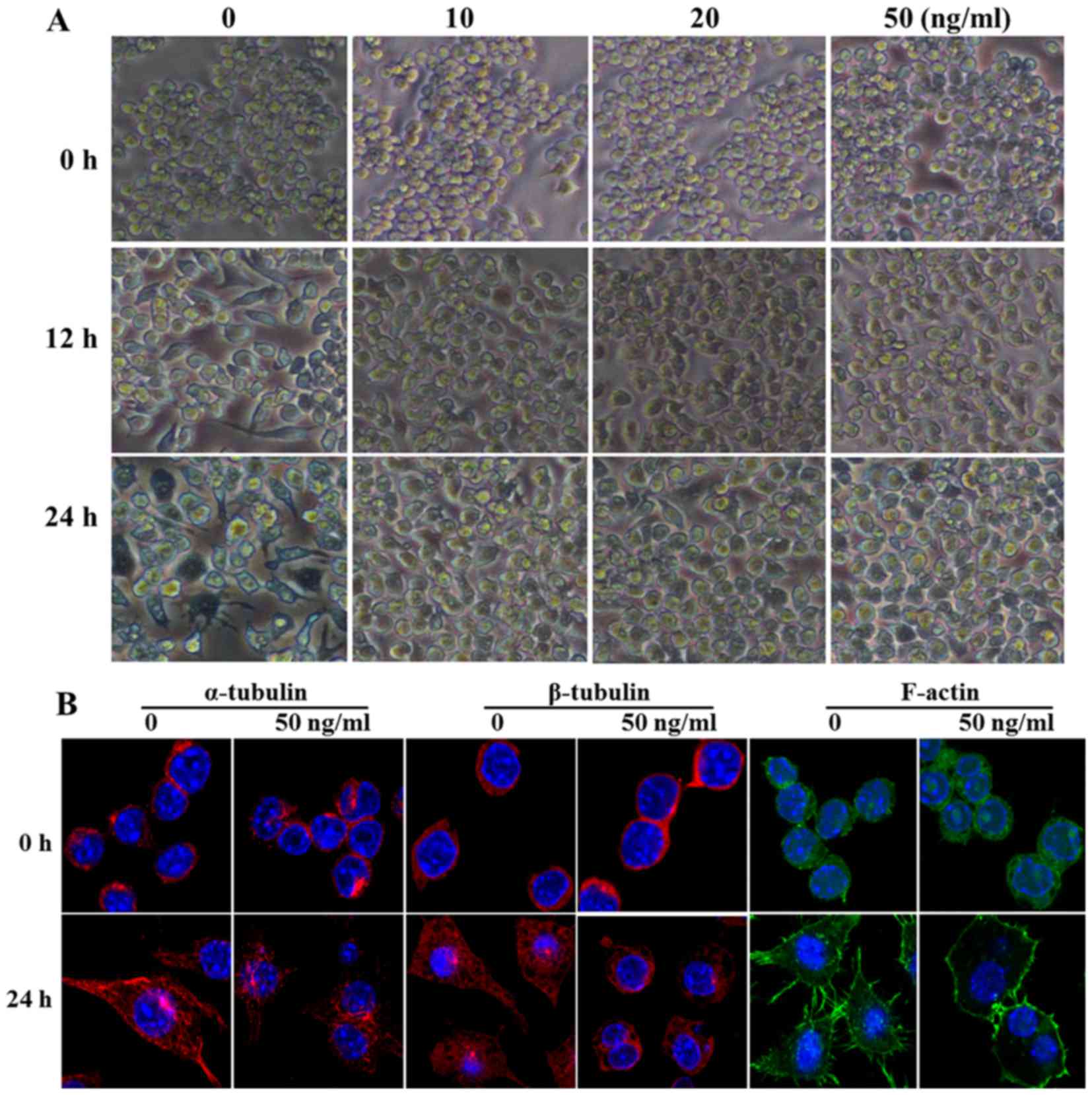
 | Figure 1Lipopolysaccharide (LPS) stimulation,
but not poly(I:C) stimulation, led to changes in macrophage
morphology. (A) RAW264.7 cells and (B) mouse primary peritoneal
macrophages (PM) were stimulated with LPS (100 ng/ml) or poly(I:C)
(20 µg/ml) for 0, 4, 8, 12 and 24 h, and interleukin (IL)-6,
interferon (IFN)-β, tumor necrosis factor (TNF)-α and inducible
nitric oxide synthase (iNOS) expression was detected with
quantitative polymerase chain reaction (PCR).
*P<0.05, **P<0.01,
***P<0.001, #P<0.05 and
##P<0.01. (C) Toll-like receptor (TLR)3 and TLR4
expression was detected with the PCR assay in RAW264.7 cells and
mouse PM. (D) The expression of α- and β-tubulin and β-actin and
glyceraldehyde 3-phosphate dehydrogenase (GAPDH) were measured at
indicated timepoints with simple western assays. Similar
observations were obtained from three independent experiments. (E)
After RAW264.7 cells were stimulated with LPS (100 ng/ml) or
poly(I:C) (20 µg/ml) for 24 h, α- and β-tubulin and F-actin
were stained and detected using a confocal microscope. (F) Cell
morphology was observed under a light microscope after LPS (100
ng/ml) or poly(I:C) (20 µg/ml) treatment for 24 h (original
magnification, ×100). |
VCR pretreatment represses the
cytoskeleton rearrangement induced by LPS
VCR may combine with tubulin, actin and 10-nm
filament proteins to inhibit cytoskeletal rearrangement, and plays
an important role in the cytoskeleton and signaling pathways, such
as colchicine. Therefore, in order to further confirm the effect of
cell morphological changes on macrophage activation induced by LPS,
the mouse macrophage cell line RAW264.7 was pretreated with
gradually increasing concentrations of VCR for 30 min, and then
stimulated with 100 ng/ml LPS for 12 or 24 h. As shown in Fig. 2A, a concentration as low as 10
ng/ml was able to significantly inhibit LPS-induced cell
morphological changes and the cells maintained a round shape,
rather than developing cytoplasmic projections. Furthermore,
cytoskeleton rearrangement following VCR pretreatment was observed
using a confocal microscope. As shown in Fig. 2B, the arrangement of α- and
β-tubulin and F-actin was found to be disordered and the
cyto-skeleton rearrangement was significantly inhibited following
VCR pretreatment and LPS stimulation for 24 h. Taken together,
these data indicate that VCR pretreatment repressed the
cytoskeleton rearrangement induced by LPS.
VCR pretreatment reduces the synthesis
and secretion of cytokines and attenuates the migration of
macrophages
To investigate the status of macrophage activation
during the inhibition of cell morphological changes, the expression
and secretion of TNF-α and IL-6 was detected with PCR and ELISA. As
shown in Fig. 3A and B, in
RAW264.7 cells, VCR priming markedly decreased the expression and
secretion of IL-6 in a dose-dependent manner. However, VCR priming
diminished the secretion of TNF-α, but not its expression. As shown
in Fig. 3C and D, the expression
and secretion of IL-6 and TNF-α were all decreased in mouse primary
peritoneal macrophages during VCR pretreatment. Macrophage invasion
is a key factor in the inflammatory response; hence, their invasion
ability was detected with Boyden chamber assays. As shown in
Fig. 3E, macrophage invasion was
significantly accelerated following LPS stimulation. However, only
few cells invaded through the filter membrane in the poly(I:C)
group. VCR pretreatment remarkably abrogated macrophage invasion
caused by LPS, but not poly(I:C). Taken together, these data
indicate that VCR pretreatment inhibited the LPS-induced cytokine
expression and secretion and attenuated invasion in
macrophages.
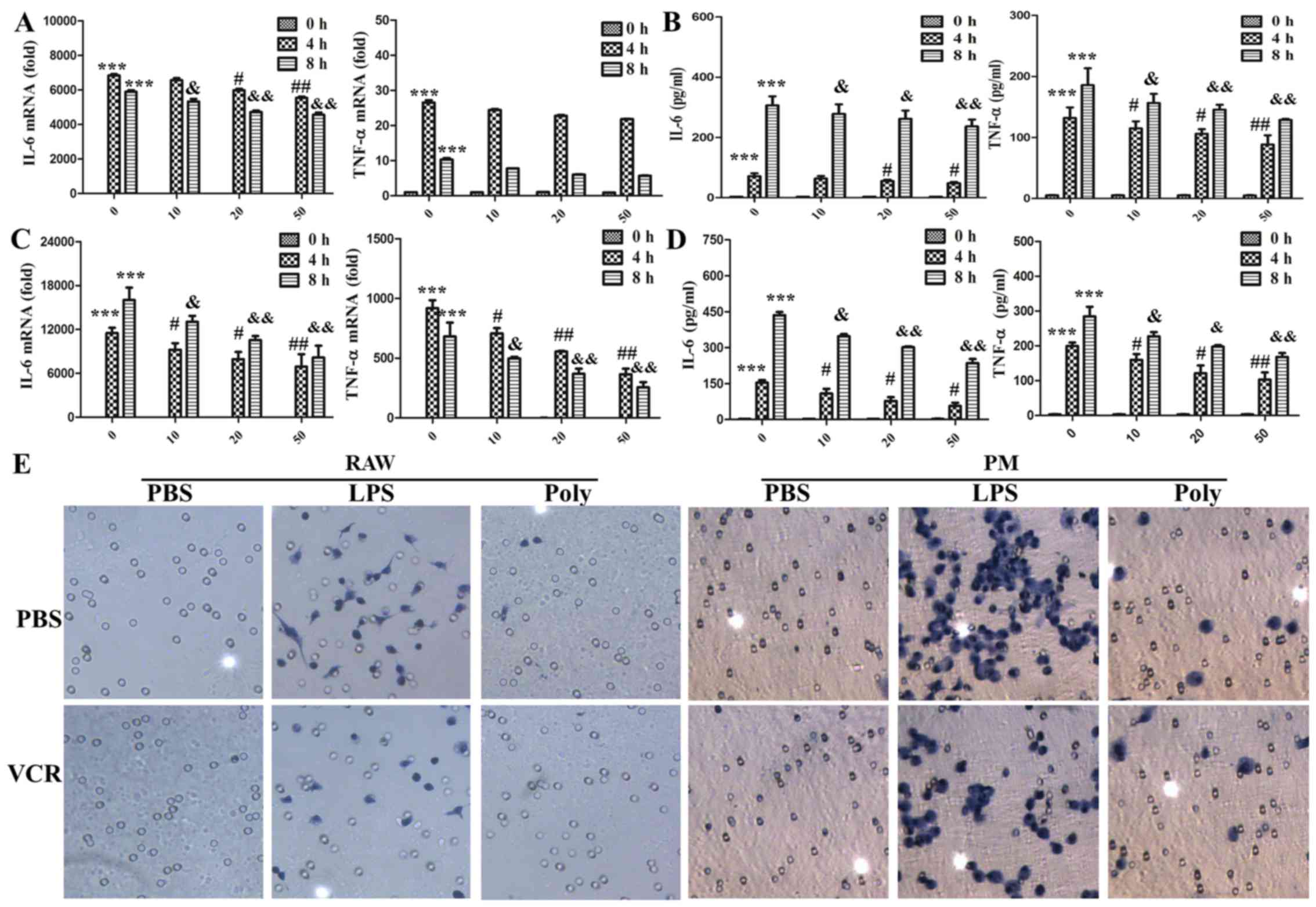 | Figure 3Vincristine (VCR) pretreatment
reduced the synthesis and secretion of cytokines and cell migration
caused by lipopolysaccharide (LPS). (A and B) RAW264.7 cells were
primed for 30 min with VCR at different concentrations and then
treated for 0, 4 and 8 h with 100 ng/ml LPS. (A) Interleukin (IL)-6
and tumor necrosis factor (TNF)-α expression was measured by
quantitative polymerase chain reaction (PCR).
***P<0.001, #P<0.05,
##P<0.01, &P<0.05 and
&&P<0.01. (B) Secretion of IL-6 and TNF-α in
the cell culture supernatants was determined by ELISA
(***P<0.001, #P<0.05,
##P<0.01, &P<0.05 and
&&P<0.01). (C and D) Peritoneal macrophages
were primed for 30 min with VCR at different concentrations and
then treated for 0, 4 and 8 h with 100 ng/ml LPS. (C) IL-6 and
TNF-α expression was measured by quantitative PCR
(***P<0.001, #P<0.05,
##P<0.01, &P<0.05 and
&&P<0.01). (D) Secretion of IL-6 and TNF-α in
the cell culture supernatants was determined by ELISA
(***P<0.001, #P<0.05,
##P<0.01, &P<0.05 and
&&P<0.01). The data are shown as mean ±
standard deviation of triplicate samples. (E) After RAW264.7 cells
and peritoneal macrophages (PM) were primed for 30 min with 50
ng/ml VCR and then treated for 20 h (RAW264.7) or 36 h (PM) with
100 ng/ml LPS and poly(I:C) (20 µg/ml), migrated cells on
the lower surface of the membrane were stained with hematoxylin
(original magnification, ×200). Similar observations were obtained
from three independent experiments. PBS, phosphate-buffered
saline. |
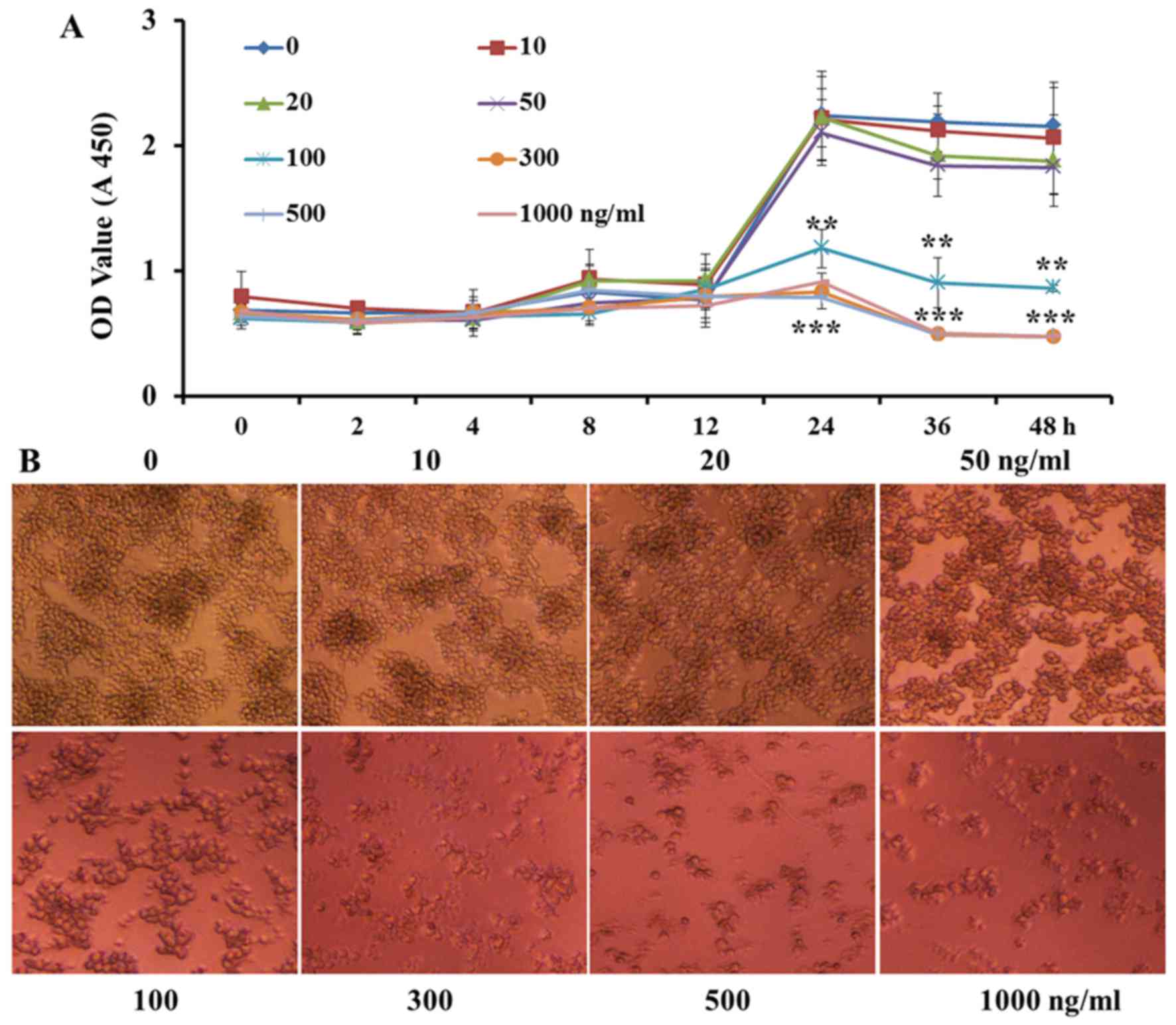
VCR treatment at low concentrations and
for a short time does not inhibit cell proliferation
In order to verify that the inhibition of
proinflammatory cytokine expression and secretion and cell invasion
in RAW264.7 macrophages by VCR pretreatment was not due to VCR
decreasing cell proliferation, the macrophages were pretreated with
increasing concentrations of VCR (0, 10, 20, 50, 100, 300, 500 and
1,000 ng/ml). As shown in Fig.
4A, high or low concentrations of VCR did not significantly
affect cell proliferation within 12 h. However, over time, VCR at
≥100 ng/ml achieved a higher cell mortality rate. Following VCR
primed for 36 h, low concentrations of VCR were also cytotoxic to a
certain extent. As shown in Fig.
4B, VCR at <50 ng/ml did not significantly affect the cell
count for 24 h. However, the cytotoxic effect was markedly
increased at VCR concentrations of ≥100 ng/ml. Taken together,
these data indicate that VCR pretreatment at low doses and for a
short time did not significantly affect cell proliferation and
activity.
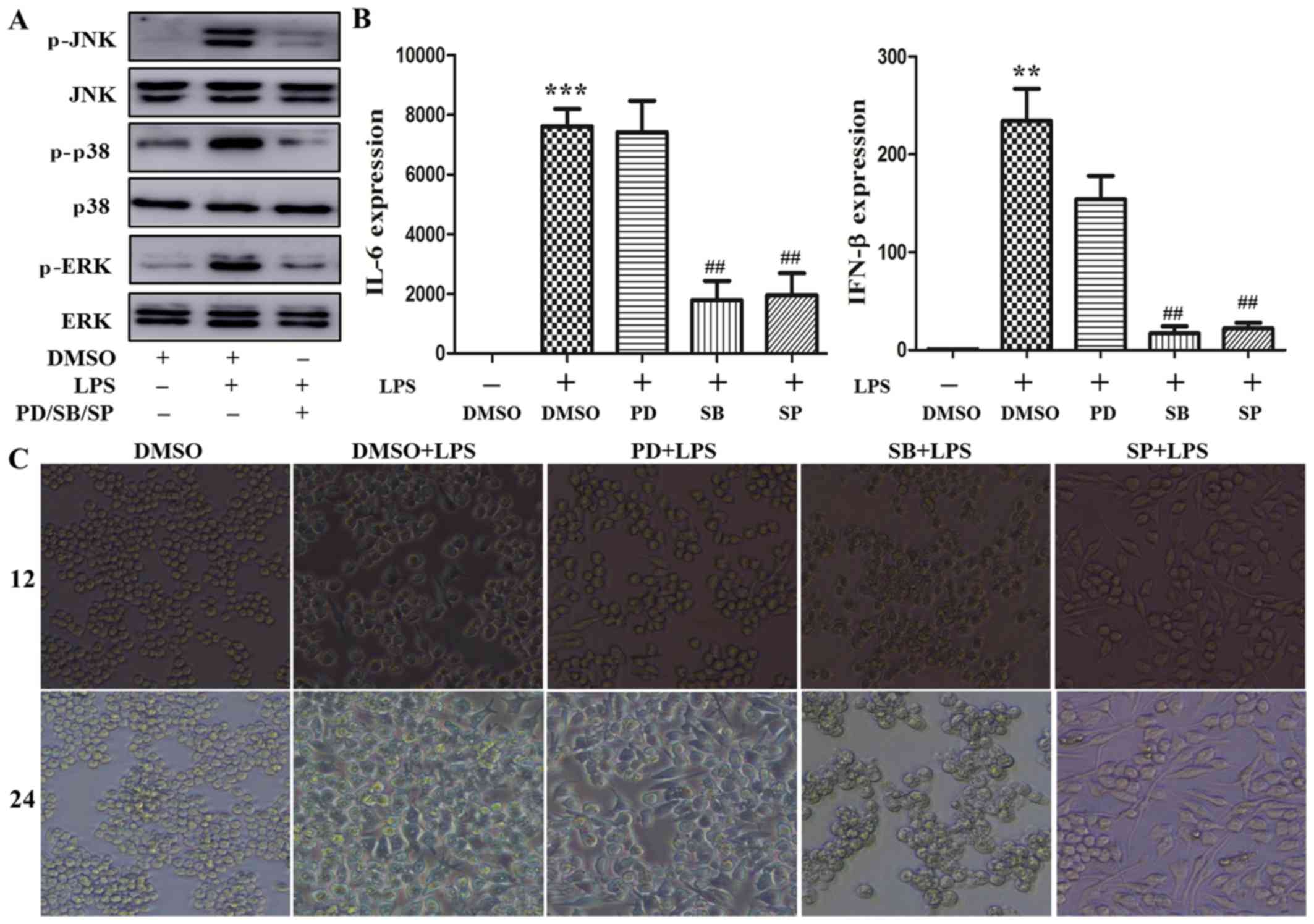
The MAPK/p38 signaling pathway plays a
predominant role in cytoskeleton rearrangement by augmenting
cytokine production induced by LPS
TLR4 initiates the MyD88-dependent and -independent
pathway that involves TRAF6 to activate the MAPK signaling pathway,
resulting in phosphorylation of the extracellular signal-regulated
kinase (ERK), p38 and c-Jun N-terminal kinase (JNK), with
subsequent nuclear translocation to regulate proinflammatory
cytokine transcription (15).
PD98059, SB203580 and SP600125 are effective inhibitors
downregulating the phosphorylation level of MAPK signaling pathway
molecules (16). To determine the
role of the MAPK signaling pathway in cytoskeleton rearrangement
caused by LPS, the macrophages were respectively pretreated with
these inhibitors for 30 min, and then stimulated with 100 ng/ml
LPS. As shown in Fig. 5A, ERK,
p38 and JNK phosphorylation were markedly increased following LPS
stimulation, which was consistent with previously reported data on
LPS inducing MAPK activation (16). Treatment with the abovementioned
inhibitors significantly attenuated LPS-induced ERK, p38 and JNK
phosphorylation. Next, the effect of these inhibitors on
LPS-induced cytokine expression was further investigated. As shown
in Fig. 5B, LPS increased the
levels of IL-6 and IFN-β mRNA following stimulation for 4 h. This
increase was reversed by treatment with SB203580 and SP600125, but
not PD98059. Furthermore, the effect of these inhibitors on
macrophage morphology was assessed. As shown in Fig. 5C, LPS induced cell stretching
after stimulation for 12 and 24 h. This effect was abrogated by
treatment with SB203580, but not PD98059 and SP600125. Moreover,
the phosphorylation level of p38 and p65 was measured after VCR
pretreatment followed by LPS stimulation. As shown in Fig. 5D, p38 phosphorylation was lower
following VCR pretreatment; however, there was little change in the
phosphorylation of p65. Taken together, these data indicate that
the MAPK/p38 signaling pathway may play a prominent role in
cytoskeleton rearrangement leading to cytokine production induced
by LPS.
Discussion
The present study demonstrated a striking difference
in cytokine production and cell morphology between LPS- and
poly(I:C)-induced macrophage activation. Stimulation by LPS, but
not poly(I:C), significantly decreased tubulin protein expression,
leading to cytoskeleton rearrangement, while the expression of
actin and GAPDH were not significantly affected. VCR, a drug
accelerating microtubule depolymerization via tubulin binding, was
found to inhibit cytoskeleton rearrangement in LPS-induced
macrophages, decrease the production of proinflammatory cytokines
and diminish macrophage migration. Furthermore, the MAPK/p38
signaling pathway may play a prominent role in cytoskeleton
rearrangement leading to cytokine production induced by LPS. Our
data suggest that the cytoskeleton regulates LPS-induced macrophage
activation via the MAPK/p38 signaling pathway.
Macrophages represent a heterogeneous population of
immune cells with various functions in body homeostasis and disease
initiation, maintenance and resolution (17,18). Macrophages express a myriad of
pattern recognition receptors (PRRs), including TLRs, that enable
them to rapidly respond to pathogen infections and to coordinate
innate and adaptive immunity (2,13).
Disturbances in macrophage function may lead to abnormal repair,
such as uncontrolled production of inflammatory and growth
mediators, deficient production of anti-inflammatory macrophages,
or failed communication between macrophages and other non-immune
cells, all of which are conducive to a status of persistent injury
(18). Therefore, it is
particularly important to elucidate the mechanism underlying
macrophage activation. TLRs are membrane-associated PRRs that
consist of an ectodomain with leucine-rich repeats (LRR) that
mediate interactions with activator and coreceptors, a
transmembrane region, and an intracellular TIR signaling domain
(19). TLR3/4 are the main PRRs
for recognition of Gram-negative bacterial LPS (20,21) and viral dsRNA (22). Their binding to corresponding
ligands leads to accumulation of intracellular TIR domains to
enable recruitment of adapter proteins by TLR dimerization. The
MyD88-dependent signaling pathway is activated by TLR4 from the
cell surface (23), and then TLR4
translocates into the endosome in a GTPase- and CD14-dependent
manner to trigger the TRIF-dependent signaling pathway that
activates the TBK1-IRF3 to induce production of type≈I IFNs. TLR3
only utilizes the TRIF-dependent signaling pathway (19). The TLR signaling pathway is
regulated at multiple levels, such as the expression of TLR
(24), TLR signal complex
assembly, ubiquitination and phosphorylation of associated proteins
(25,26), induction of negative and positive
regulators (13,26,27), and epigenetic and
post-transcriptional modification regulation (28,29). It was observed that the expression
and secretion of IL-6, TNF-α and IL-12 p40 and p70 were always
lower during poly(I:C) compared with LPS stimulation in previous
studies (16,30,31). Consistently, in the present study,
it was demonstrated that IL-6 and IFN-β expression following
poly(I:C) stimulation were always lower compared with LPS
stimulation at any given timepoint, although the reason for this
difference remains unclear.
The cytoskeleton plays a central role in cell
morphology maintenance, cell migration and division, and organelle
movement and localization; in addition, it is involved in intra-
and extracellular signal transduction. The three elementary
structural components of the cytoskeleton are microtubules,
microfilaments and intermediate filaments via the polymerization
and assembling of different monomers. These polymers undergo
continual turnover and rearrangement and specifically bind
different proteins to yield their respective functions. In the
present study, a significant difference in cell morphology was
observed between LPS and poly(I:C)-induced macrophage activation.
LPS stimulation, but not poly(I:C) stimulation, significantly
decreased tubulin protein expression, leading to cytoskeleton
rearrangement. VCR, a drug accelerating microtubule
depolymerization by binding to tubulin, inhibits cytoskeleton
rearrangement in LPS-induced macrophages, decreasing the production
of proinflammatory cytokines and diminishing cell migration. Thus,
it was validated that cytoskeleton regulated LPS- but not
poly(I:C)-induced macrophage activation. In addition, actin and
GAPDH were found to be more stable as internal reference compared
with tubulin in detecting the change in the expression of other
proteins during LPS-induced macrophage activation.
The MAPK intracellular signaling pathway is a key
mediator of TLR3/4-induced signal transduction. MAPKs, including
ERK, p38 and JNK, regulate the synthesis of inflammatory mediators
at the transcriptional and translational levels through NF-κB
activation (32,33). In the present study, it was
demonstrated that pretreatment with PD98059, an EKR inhibitor, did
not affect cytokine expression mediated by the MAPK/ERK signaling
pathway. Similarly, it was observed that SB203580, a p38 signaling
pathway inhibitor, repressed cytoskeleton rearrangement in
LPS-induced macrophages and led to inhibition of the production of
proinflammatory cytokines. PD98059 and SP600125, MAPK/ERK and JNK
signaling pathway inhibitors, respectively, exerted no effect on
cytoskeleton rearrangement leading to cytokine production. In
previous studies, the MAPK signaling pathway was found to
participate in tubulin and/or actin polymerization, regulating
migration of vascular smooth muscle cells (34), podocyte response to ox-LDL
(35) and hepatocellular
cholestasis induced by oxidative stress (36). In addition, it was demonstrated
that p38 phosphorylation, but not p65 phosphorylation, was ablated
following VCR priming and LPS stimulation compared with LPS alone.
In conclusion, our results demonstrated that the cytoskeleton
played a different role in LPS- and poly(I:C)-induced macrophage
activation. The MAPK/p38 signaling pathway, but not ERK and JNK,
promoted cytoskeleton rearrangement in LPS-induced macrophage to
promote the production of proinflammatory cytokines and cell
migration. Given the pathological role of the macrophage
inflammatory response in certain autoimmune diseases, VCR at low
doses may be of therapeutic value in the treatment of autoimmune
diseases with uncontrolled inflammatory response. In addition, due
to the differences in the stability of cytoskeletal proteins,
particularly tubulin, in LPS- and poly(I:C)-induced macrophage
activation, GAPDH was used as an internal control to determine the
expression level of the other proteins.
Acknowledgments
The present study was supported in part by grants
from the National Natural Science Foundation of China (nos.
81472685 and 81600469), the Major Special Plan of Science and
Technology of Shandong Province (no. 2015ZDXX0802A01), the
Promotive Research Fund for Excellent Young and Middle-aged
Scientisits of Shandong Province (no. BS2014yy037), and the Science
and Technology Development Projects of Shandong Province (no.
2016GSF201126).
References
|
1
|
Pelka K, Shibata T, Miyake K and Latz E:
Nucleic acid-sensing TLRs and autoimmunity: Novel insights from
structural and cell biology. Immunol Rev. 269:60–75. 2016.
View Article : Google Scholar
|
|
2
|
Kawai T and Akira S: The role of
pattern-recognition receptors in innate immunity: Update on
Toll-like receptors. Nat Immunol. 11:373–384. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Zhao W, Qi J, Wang L, Zhang M, Wang P and
Gao C: LY294002 inhibits TLR3/4-mediated IFN-β production via
inhibition of IRF3 activation with a PI3K-independent mechanism.
FEBS Lett. 586:705–710. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Bosshart H and Heinzelmann M: Targeting
bacterial endotoxin: Two sides of a coin. Ann NY Acad Sci.
1096:1–17. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Kustermans G, Piette J and Legrand-Poels
S: Actin-targeting natural compounds as tools to study the role of
actin cytoskeleton in signal transduction. Biochem Pharmacol.
76:1310–1322. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Diesel B, Hoppstädter J, Hachenthal N,
Zarbock R, Cavelius C, Wahl B, Thewes N, Jacobs K, Kraegeloh A and
Kiemer AK: Activation of Rac1 GTPase by nanoparticulate structures
in human macrophages. Eur J Pharm Biopharm. 84:315–324. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Song W, Liu C and Upadhyaya A: The pivotal
position of the actin cytoskeleton in the initiation and regulation
of B cell receptor activation. Biochim Biophys Acta. 1838:569–578.
2014. View Article : Google Scholar
|
|
8
|
Chen YR, Feng F, Wang L, Qu SY, Zhang ZQ,
Liu L, Qin HY, Liang YM and Han H: Deletion of RBP-j in dendritic
cells compromises TLR-mediated DC activation accompanied by
abnormal cytoskeleton reorganization. Mol Biol Rep. 40:1531–1539.
2013. View Article : Google Scholar
|
|
9
|
Moudi M, Go R, Yien CY and Nazre M: Vinca
alkaloids. Int J Prev Med. 4:1231–1235. 2013.
|
|
10
|
Pathak P, Hess R and Weiss MA: Liposomal
vincristine for relapsed or refractory Ph-negative acute
lymphoblastic leukemia: A review of literature. Ther Adv Hematol.
5:18–24. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Thakur V, Kush P, Pandey RS, Jain UK,
Chandra R and Madan J: Vincristine sulfate loaded dextran
microspheres amalgamated with thermosensitive gel offered sustained
release and enhanced cytotoxicity in THP-1, human leukemia cells:
In vitro and in vivo study. Mater Sci Eng C. 61:113–122. 2016.
View Article : Google Scholar
|
|
12
|
Ruiz-Gómez MJ, Souviron A,
Martínez-Morillo M and Gil L: P-glycoprotein, glutathione and
glutathione S-transferase increase in a colon carcinoma cell line
by colchicine. J Physiol Biochem. 56:307–312. 2000. View Article : Google Scholar
|
|
13
|
Qi J, Qiao Y, Wang P, Li S, Zhao W and Gao
C: microRNA-210 negatively regulates LPS-induced production of
proinflammatory cytokines by targeting NF-κB1 in murine
macrophages. FEBS Lett. 586:1201–1207. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Qi J, Li T, Bian H, Li F, Ju Y, Gao S, Su
J, Ren W and Qin C: SNAI1 promotes the development of HCC through
the enhancement of proliferation and inhibition of apoptosis. FEBS
Open Bio. 6:326–337. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Wu J, Zhang H, Hu B, Yang L, Wang P, Wang
F and Meng X: Coptisine from Coptis chinensis inhibits production
of inflammatory mediators in lipopolysaccharide-stimulated RAW264.7
murine macrophage cells. Eur J Pharmacol. 780:106–114. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Bode KA, Schmitz F, Vargas L, Heeg K and
Dalpke AH: Kinetic of RelA activation controls magnitude of
TLR-mediated IL-12p40 induction. J Immunol. 182:2176–2184. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Devisscher L, Verhelst X, Colle I, Van
Vlierberghe H and Geerts A: The role of macrophages in
obesity-driven chronic liver disease. J Leukoc Biol. 99:693–698.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Wynn TA and Vannella KM: Macrophages in
tissue repair, regeneration, and fibrosis. Immunity. 44:450–462.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Beutler B: Microbe sensing, positive
feedback loops, and the pathogenesis of inflammatory diseases.
Immunol Rev. 227:248–263. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Alexopoulou L, Holt AC, Medzhitov R and
Flavell RA: Recognition of double-stranded RNA and activation of
NF-kappaB by Toll-like receptor 3. Nature. 413:732–738. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Ruiz J, Kanagavelu S, Flores C, Romero L,
Riveron R, Shih DQ and Fukata M: Systemic activation of
TLR3-dependent TRIF signaling confers host defense against
gram-negative bacteria in the intestine. Front Cell Infect
Microbiol. 5:1052016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Poltorak A, Smirnova I, Clisch R and
Beutler B: Limits of a deletion spanning Tlr4 in C57BL/10ScCr mice.
J Endotoxin Res. 6:51–56. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Kagan JC and Medzhitov R:
Phosphoinositide-mediated adaptor recruitment controls Toll-like
receptor signaling. Cell. 125:943–955. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Romieu-Mourez R, François M, Boivin MN,
Bouchentouf M, Spaner DE and Galipeau J: Cytokine modulation of TLR
expression and activation in mesenchymal stromal cells leads to a
proinflammatory phenotype. J Immunol. 182:7963–7973. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Xiong Y and Medvedev AE: Induction of
endotoxin tolerance in vivo inhibits activation of IRAK4 and
increases negative regulators IRAK-M, SHIP-1, and A20. J Leukoc
Biol. 90:1141–1148. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Xiong Y, Qiu F, Piao W, Song C, Wahl LM
and Medvedev AE: Endotoxin tolerance impairs IL-1
receptor-associated kinase (IRAK) 4 and TGF-beta-activated kinase 1
activation, K63-linked polyubiquitination and assembly of IRAK1,
TNF receptor-associated factor 6, and IkappaB kinase gamma and
increases A20 expression. J Biol Chem. 286:7905–7916. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Murphy MB, Xiong Y, Pattabiraman G,
Manavalan TT, Qiu F and Medvedev AE: Pellino-3 promotes endotoxin
tolerance and acts as a negative regulator of TLR2 and TLR4
signaling. J Leukoc Biol. 98:963–974. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Foster SL, Hargreaves DC and Medzhitov R:
Gene-specific control of inflammation by TLR-induced chromatin
modifications. Nature. 447:972–978. 2007.PubMed/NCBI
|
|
29
|
O'Neill LA, Sheedy FJ and McCoy CE:
MicroRNAs: The fine-tuners of Toll-like receptor signalling. Nat
Rev Immunol. 11:163–175. 2011. View
Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhao W, Wang L, Zhang M, Wang P, Qi J,
Zhang L and Gao C: Nuclear to cytoplasmic translocation of
heterogeneous nuclear ribonucleoprotein U enhances TLR-induced
proinflammatory cytokine production by stabilizing mRNAs in
macrophages. J Immunol. 188:3179–3187. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Zhang M, Wang L, Zhao X, Zhao K, Meng H,
Zhao W and Gao C: TRAF-interacting protein (TRIP) negatively
regulates IFN-β production and antiviral response by promoting
proteasomal degradation of TANK-binding kinase 1. J Exp Med.
209:1703–1711. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Chan ED and Riches DW: IFN-gamma + LPS
induction of iNOS is modulated by ERK, JNK/SAPK, and p38(mapk) in a
mouse macrophage cell line. Am J Physiol Cell Physiol.
280:C441–C450. 2001.PubMed/NCBI
|
|
33
|
Kaminska B: MAPK signalling pathways as
molecular targets for anti-inflammatory therapy–from molecular
mechanisms to therapeutic benefits. Biochim Biophys Acta.
1754:253–262. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lee GL, Wu JY, Tsai CS, Lin CY, Tsai YT,
Lin CS, Wang YF, Yet SF, Hsu YJ and Kuo CC: TLR4-activated
MAPK-IL-6 axis regulates vascular smooth muscle cell function. Int
J Mol Sci. 17:E13942016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Hu M, Fan M, Zhen J, Lin J, Wang Q, Lv Z
and Wang R: FAK contributes to proteinuria in hypercholesterolaemic
rats and modulates podocyte F-actin re-organization via activating
p38 in response to ox-LDL. J Cell Mol Med. 21:552–567. 2017.
View Article : Google Scholar
|
|
36
|
Toledo FD, Basiglio CL, Barosso IR,
Boaglio AC, Zucchetti AE, Sánchez Pozzi EJ and Roma MG:
Mitogen-activated protein kinases are involved in hepatocanalicular
dysfunction and cholestasis induced by oxidative stress. Arch
Toxicol. 91:2391–2403. 2017. View Article : Google Scholar
|