Introduction
Chronic neuroinflammation serves an important role
in progressive neurodegenerative disorders, including Alzheimer's
disease (AD) and Parkinson's disease (PD) (1). Neuroinflammation may occur as a
result of endotoxins, injury or infection. Microglial cells are
macrophages specific to the central nervous system (CNS), which
serve a crucial role in host defenses against toxins in the CNS
(2). Specific stimulators may
activate the microglial cells by binding with pattern-recognition
receptors (PRR) on their surface (3). Microglial cells may be activated
into two distinct types, which are designated as classically
activated M1 and alternatively activated M2 cells. The activation
of M1 microglial cells initiates inflammatory responses via the
production of inflammatory mediators, including nitric oxide (NO),
reactive oxygen species (ROS), tumor necrosis factor-α (TNF-α) and
prostaglandin E2 (PGE2), which may promote
neuronal injury (4). Activated M2
microglial cells produce anti-inflammatory molecules that reduce
inflammation and repair injured tissue (5). The maintenance of microglial cell
activation is important for brain development and the repair of
injured sites within the brain (6). However, under pathological
conditions, extensive classically activated microglia-mediated
inflammation contributes to neuronal injury through the release of
cytotoxic agents, including NO, ROS and other inflammatory
mediators (7).
During chronic neuroinflammation, intracellular
signaling cascades, including nuclear factor-κB (NF-κB) and
mitogen-activated protein kinases (MAPKs), become activated
(8). This subsequently increases
inflammatory mediators, including TNF-α, interleukin-1β (IL-1β),
IL-6 and cyclooxygenase-2 (COX-2), as well as increases the
expression of genes that regulate cell survival and growth
(9). Additionally, the p38 MAPK
signaling pathway in activated microglia has been associated with
several inflammatory diseases, including rheumatoid arthritis, AD
and inflammatory bowel disease (10). Inflammatory signaling pathways
following microglial activation are considered a prime target for
inhibition of neuroinflammation-mediated neurodegeneration
(2). A recent study suggested
that the activation of the nuclear factor (erythroid-derived
2)-like 2 (Nrf2)/antioxidant response element (ARE) signaling
pathway was an important target in blocking neuroinflammation and
neurodegenerative diseases (11).
Another previous study indicated that Nrf2 inhibited microglial
hyper-activation by suppressing the p38 MAPK and NF-κB signaling
pathways (12).
Under normal conditions, Nrf2 activation is
suppressed by the Kelch-like ECH-associated protein 1, protein
dimer, which acts as an adaptor molecule of the cullin3 ubiquitin
protein ligase complex. However, in response to stimuli, Nrf2
translocates to the nucleus and binds with the ARE sequence located
in the promoter region of the oxidative stress response genes,
NAD(P)H dehydrogenase [quinone] (NQO)-1 and heme oxygenase-1 (HO-1)
(13). The HO-1 gene has been
previously studied for its potential neuroprotective and
anti-inflammatory effects (14).
The hippocampi of Nrf2 and HO-1 knockout mice were demonstrated to
be hypersensitive to neuroinflammation induced by
lipopolysaccharide (LPS), as indicated by an increase in the
secretion of inflammatory markers, including inducible nitric oxide
synthase (iNOS), IL-6 and TNF-α (15,16). The results of these previous
studies suggest that targets to activate Nrf2 or HO-1 in the
microglia would be a beneficial therapeutic strategy to reduce
neuroinflammation.
Curcumin is one of the main components of the
rhizomes of Curcuma longa. It has been previously used in
oriental medicine as a sedative, analgesic and antipyretic, as well
as a treatment for pancreatic cancer, AD and epilepsy (16,17). Curcumin is a highly lipophilic
natural compound that is able to pass through the blood-brain
barrier (18). Within the brain
curcumin appears primarily in the hippocampus (18). A previous study has reported that
curcumin may inhibit amyloid β oligomers and rescue neuronal injury
in models of AD (19).
Furthermore, curcumin has also been reported to promote the
development of the M2 microglial phenotype in an HO-1-dependent
manner and reduce iNOS induction, thus protecting microglial cells
against oxidative stress (16).
The anti-inflammatory effects of curcumin occur through several
different mechanisms (16,17).
Curcumin is able to regulate the activation of transcription
factors, such as activator protein-1 and NF-κB and it may block
COX-2 and iNOS (20). It has also
been reported that curcumin has an inhibitory effect on the
LPS-induced production of inflammatory mediators, including IL-1β,
IL-6, TNF-α and IFN-α (21).
However, the effect of curcumin on neuroinflammation is not
thoroughly understood. Therefore, the present study investigated
the anti-inflammatory effects of curcumin and the underlying
molecular mechanisms by which it affects microglial cells and may
confer neuroprotection.
Materials and methods
Materials
Curcumin was purchased from Sigma-Aldrich (Merck
KGaA, Darmstadt, Germany). Protoporphyrin IX (SnPP) and antibodies
directed against HO-1 (sc-390991), Nrf2 (sc-722), TATA-binding
protein (TBP; sc-74595), α-tubulin (sc-134237) and β-actin
(sc-130065) were purchased from Santa Cruz Biotechnology, Inc.
(Dallas, TX, USA). Antibodies directed against COX-2 (4842S), iNOS
(13120), phosphorylated (p)-MAPK (9910s), MAPK (9926), protein
kinase B (Akt; 4685), p-Akt (13038), and NF-κB pathway kit (9936)
were purchased from Cell Signaling Technology, Inc. (Danvers, MA,
USA). Pam3CSK4, Dulbecco's modified Eagle's medium (DMEM;
11995-065) and fetal bovine serum (FBS; 10099-1) were purchased
from Gibco (Thermo Fisher Scientific, Inc., Waltham, MA, USA).
c-Jun NH2-terminal protein kinase (JNK) inhibitor (JNK inhibitor
II; 420119); Akt inhibitor (wortmannin; 12-338), extracellular
signal-regulated kinase (ERK) inhibitor (PD98059, 513000) and p38
inhibitor (SB230580, 559395) were purchased from EMD Millipore
(Billerica, MA, USA).
Cell culture
Mouse BV-2 microglial cells were purchased from the
American Type Culture Collection (Manassas, VA, USA). Cells were
cultured in DMEM supplemented with 10% heat-inactivated FBS and 1%
penicillin-streptomycin at 37°C in a 5% CO2 humidified
atmosphere.
Cell viability assay
The cytotoxicity of curcumin was assessed using a
MTT-based colorimetric assay. BV-2 cells were seeded in 24-well
plates at a density of 5×105 cells/well. The cells were
treated with different concentrations of curcumin (5, 10 and 20
µl) or co-treated with Pam3CSK4 (0.1 µg/ml) for 24 h
at 37°C. Subsequently, MTT solution (5 µl of 5 mg/ml) was
added to each well (final concentration, 62.5 µg/ml).
Following incubation for 3 h at 37°C in 5% CO2, the
supernatant was removed and the formazan crystals produced in
viable cells were solubilized with 150 µl dimethyl
sulfoxide. The absorbance of each well was read at 570 nm using a
microplate reader.
Measurement of TNF-α and PGE2
concentration
BV-2 cells were firstly incubated with various
concentrations (0-20 µM) of curcumin for 1 h (37°C) and
subsequently treated with Pam3CSK4 (0.1 µg/ml) for 16 h at
37°C. The release of TNF-α and PGE2 were determined.
Cells were pretreated with SnPP (HO-1 inhibitor, 20 µM) for
30 min at 37°C, and then treated with curcumin in the presence or
absence of Pam3CSK4 for 16 h. The release of TNF-α was determined.
Following 16 h incubation at 37°C, TNF-α (MTA00B) and/or
PGE2 (KGE004B) secretion were quantified in the culture
media using enzyme-linked immunosorbent assay (ELISA) kits (R&D
Systems, Inc., Minneapolis, MN, USA) according to the
manufacturer's protocol.
Preparation of nuclear extract
Cells were treated with 20 µM curcumin for
the indicated times (0, 1, 2 and 4 h) or various doses of curcumin
(0, 1, 5, 10 or 20 µM) for 2 h at 37°C. Nuclear extracts
were prepared and examined for Nrf2 expression using western blot
analysis. BV-2 cells were treated with curcumin followed by
treatment with Pam3CSK4 for 0.5 h at 37°C. The nucleus and cytosol
extracts of BV-2 cells were harvested and analyzed by western blot
analysis to detect the degree of p65 translocation and IκBα
degradation. BV-2 microglial cells were washed three times with
cold phosphate-buffered saline (PBS) and collected in 300 µl
PBS using centrifugation at 800 x g for 5 min (4°C). The cell
pellets were suspended in buffer A [10 mM HEPES-KOH (pH 7.9); 1.5
mM MgCl2; 10 mM KCl; 0.5 mM dithiothreitol (DTT); 0.2 mM
protease inhibitor (PI)] and incubated for 5 min on ice. Buffer B
[10 mM HEPES-KOH (pH 7.9); 1.5 mM MgCl2; 420 mM NaCl;
0.2 mM EDTA; glycerol 25% v/v; 0.1 mM DTT; 0.2 mM PI] was added to
the cell extract and it was incubated on ice for 5 min prior to
centrifugation at 11,000 x g for 1 min at 4°C. Nuclear proteins
were extracted by the addition of complete lysis buffer B [10 mM
HEPES-KOH (pH 7.9); 1.5 mM MgCl2; 10 mM KCL; 0.5 mM DTT;
0.2 mM PI; 25% (w/v) glycerin; 420 mM NaCl; 0.2 mM EDTA] for 30 min
at 4°C with occasional vortexing. Following centrifugation at
11,000 x g for 5 min at 4°C, the supernatants were collected and
stored at −70°C.
Western blot analysis
BV-2 cells were treated with various concentrations
of curcumin (1, 5, 10 or 20 µM) for 1 h at 37°C, and
subsequently treated with Pam3CSK4 (0.1 µg/ml) for 8 h at
37°C, and the COX-2 expression level was determined. Cells were
treated with 20 µM curcumin for the indicated times (0–24 h)
or various doses of curcumin (0–20 µM) for 8 h at 37°C.
Total cellular extracts were harvested and examined for HO-1
expression using western blot analysis. BV-2 cells were treated
with curcumin for 1 h followed by stimulation with Pam3CSK4 for 0.5
h at 37°C. Cell extracts were collected and subjected to western
blot analysis to determine p-Akt and p-MAPK expression levels. BV-2
cells were harvested in ice-cold lysis buffer (1% Triton X-100; 1%
deoxycholate; 0.1% sodium dodecyl sulfate). The protein content of
the cell lysates was subsequently determined using Bradford reagent
(Bio-Rad Laboratories, Inc., Hercules, CA, USA). Total proteins in
each sample (50 µg) were separated by 7.5% sodium dodecyl
sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) and
transferred to polyvinylidene difluoride membranes. Following
blocking of the non-specific binding sites with 5% non-fat milk at
room temperature for 30 min, the membranes were incubated with
primary antibodies directed against COX-2 (1:500), iNOS (1:500),
p-Akt (1:1,000), p-MAPK (1:1,000), MAPK (1:1,000), p-p65, p65
(1:500) p-IκBα, IκBα (1:1,000), HO-1 (1:1,000), Nrf2 (1:1,000), TBP
(1:3,000), α-tubulin (1:3,000) and β-actin (1:3,000) for 16 h at
4°C. This was followed by incubation with horseradish
peroxidase-conjugated anti-rabbit (sc-2768; 1:5,000) or anti-mouse
(sc-2371; 1:5,000) secondary antibodies (Santa Cruz Biotechnology,
Inc.) at room temperature for 1 h. Tubulin was used as the loading
control for each lane. The proteins were visualized using an
enhanced chemiluminescence detection kit (GE Healthcare, Chicago,
IL, USA). Following washing with PBS with Tween-20, the protein
bands were visualized using the Gel Doc™ EZ Imaging system (Bio-Rad
Laboratories, Inc.) and analyzed using an ImageQuant 350 analyzer
(GE Healthcare).
Measurement of nitrite concentration
BV-2 cells were treated with various concentrations
of curcumin (5, 10 or 20 µM) for 1 h and subsequently
treated with Pam3CSK4 (0.1 µg/ml) for 16 h at 37°C. NO
synthesis in cell cultures was measured by a microplate assay.
Cells were pretreated with SnPP (HO-1 inhibitor, 20 µM) for
30 min at 37°C, and then treated with curcumin in the presence or
absence of Pam3CSK4 for 16 h at 37°C. The release of NO was
determined. BV-2 cells were treated with the JNK inhibitor (JNK II,
10 µM), Akt inhibitor (Wor, 5 µM), ERK inhibitor
(PD98059, 10 µM) or p38 inhibitor (SB230580, 10 µM)
for 1 h at 37°C, following treatment with Pam3CSK4 (0.1
µg/ml) for 16 h at 37°C. The release of NO was determined.
To measure nitrite, 100-µl aliquots were removed from the
supernatant and incubated with an equal volume of the Griess
reagent [1% sulfanilamide; 0.1%
N-(1-naphthyl)-ethylenediaminedihydrochloride; 2.5%
H3PO4] at room temperature for 10 min.
Nitrite concentration was determined by measuring the absorbance at
540 nm with a Vmax 96-well microplate spectrophotometer. Sodium
nitrite was used as a standard.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
BV-2 cells were treated with various concentrations
of curcumin (0, 5, 10 or 20 µM) for 1 h at 37°C, and
subsequently treated with Pam3CSK4 (0.1 µg/ml) for 4 h at
37°C, the mRNA expression levels of iNOS and COX-2 were determined.
BV-2 cells were incubated with 20 µM curcumin for the
indicated times (2, 4, 6, 8 and 10 h) or cells were incubated with
increasing doses of curcumin (1, 5, 10 and 20 µM) for 4 h at
37°C and the relative HO-1 mRNA expression level was measured. BV-2
cells were then treated with the JNK inhibitor (10 µM), Akt
inhibitor (5 µM), ERK inhibitor (10 µM) or p38
inhibitor (10 µM) for 1 h at 37°C, followed by treatment
with Pam3CSK4 (0.1 µg/ml) for 4 h at 37°C. The expression
levels of iNOS were determined. Total RNA was isolated from cells
using an Axyprep multisource total RNA miniprep kit (AP-MN-MS-RNA;
Corning, Inc., Corning, NY, USA) according to the manufacturer's
protocol. cDNA was synthesized from 1 µg total RNA using a
Maxime RT-PCR PreMix kit (Takara Bio, Inc., Otsu, Japan) and
anchored oligo(dT)15-primers. qPCR was performed using a
Chromo 4™ system (Bio-Rad Laboratories, Inc.) and
SYBR®-Green PCR Master Mix (Thermo Fisher Scientific,
Inc.). The following thermocycling conditions were used: 40 cycles
of 95°C for 5 min, 95°C for 15 sec and 62°C for 30 sec. Relative
amounts of target mRNA were determined using the 2−ΔΔCq
method (22) by normalizing
target mRNA comparative threshold values to those of β-actin. The
PCR primers were designed using Primier-E version 6 software
(Premier Group of Companies, Vancouver, BC, Canada) and are listed
in Table I. The relative amount
of mRNA was presented as the fold-change value compared to the
control value.
 | Table IName and sequence of primers used for
reverse transcription-quantitative polymerase chain reaction. |
Table I
Name and sequence of primers used for
reverse transcription-quantitative polymerase chain reaction.
| Gene name | Primer sequence
(5′–3′) | Gene ID | Product size
(bp) |
|---|
| Inducible nitric
oxide synthase | F:
GGCACCGAGATTGGAGTTC
R: GGTCACATTCTGCTTCT | NM001313921 | 174 |
|
Cyclooxygenase-2 | F:
TCAGGTCATTGGTGGAGAGG
R: ATGGTGGCATACATCATCAGAC | NM011198.4 | 150 |
| Heme
oxygenase-1 | F:
AGGTCCTGAAGAAGATTGC
R: TCTCCAGAGTGTTCATTCG | NM010442.2 | 175 |
| β-actin | F:
GCACCACACCTTCTACAA
R TACGACCAGAGGCATACA | NM007393.5 | 156 |
Statistical analysis
Data were expressed as the mean ± standard
deviation. Each experiment was repeated a minimum of three times.
Statistical analysis was performed using SPSS version 16.0 (SPSS,
Inc., Chicago, IL, USA) to determine significant differences.
Either a Student's t-test or one-way analysis of variance, followed
by Dunn's post hoc test was used for analyses. P<0.05 was
considered to indicate a statistically significant difference.
Results
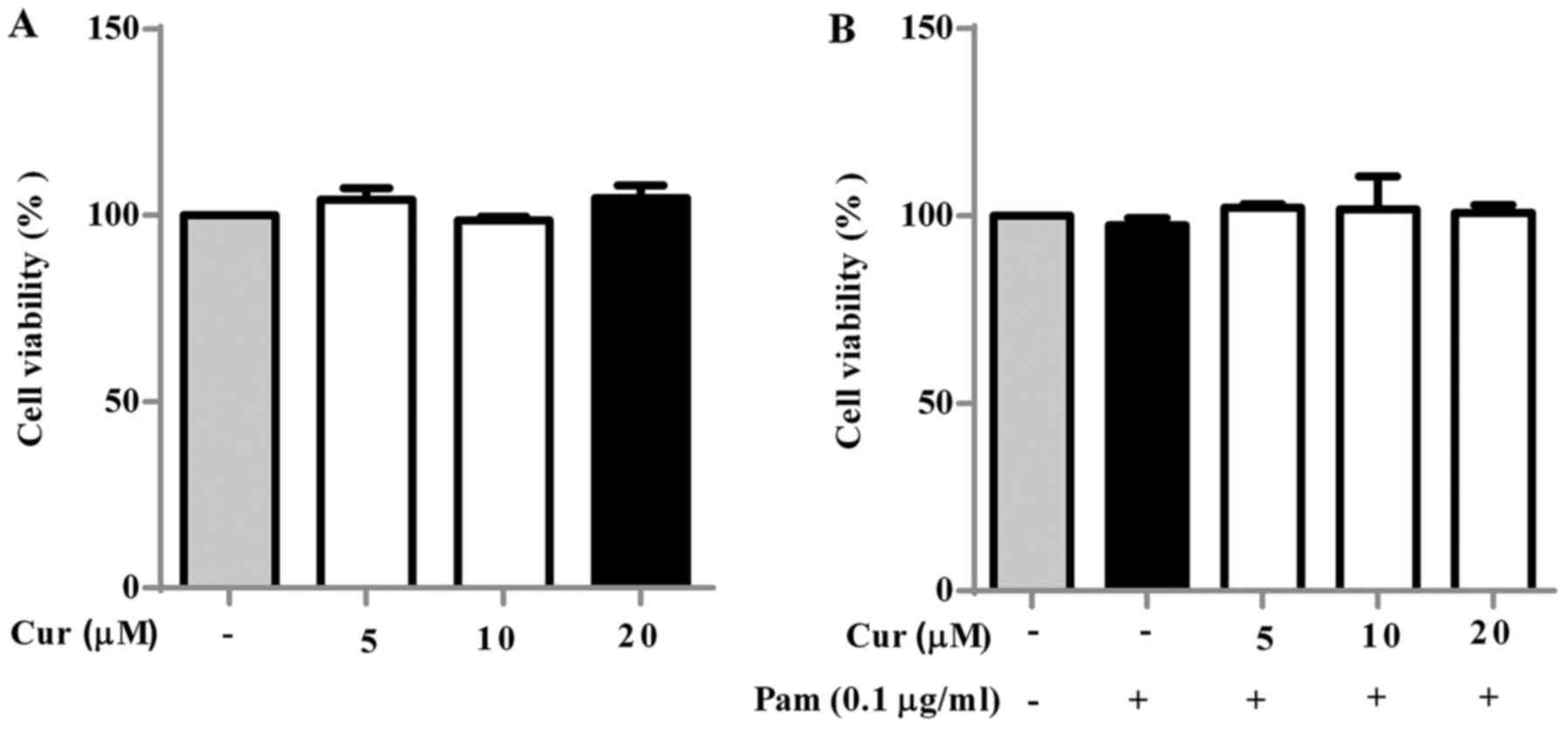
Toxicity of curcumin and PamCSK4 on BV-2
cells
To assess the cytotoxic effect of curcumin on BV-2
cells, they were treated with increasing concentrations (0–20
µM) of curcumin for 24 h and their viability was measured
(Fig. 1A). BV-2 cells without any
treatment were used as the negative control. The results revealed
that curcumin at concentrations of 5–20 µM had no notable
cytotoxic effect on BV-2 microglial cells. Thus, 5–20 µM
curcumin was selected for subsequent experiments. The cytotoxic
effect of treatment of BV-2 cells with curcumin and Pam3CSK4 (0.1
µg/ml) in combination was also investigated (Fig. 1B). No notable cytotoxic effects
were observed at all concentrations.
Inhibitory effect of curcumin on
Pam3CSK4-induced inflammatory mediators in BV-2 microglial
cells
NO, TNF-α, PGE2, iNOS and COX-2 are
inflammatory mediators present in activated microglial cells
(7). Treatment of BV-2 cells with
Pam3CSK4 alone notably increased the production of NO (Fig. 2A), PGE2 (Fig. 2B) and TNF-α (Fig. 2C) and the expression of iNOS
(Fig. 2D) and COX-2 (Fig. 2E) compared with the negative
control. Curcumin had significant inhibitory effects on the
secretion of NO and PGE2 in Pam3CSK4-stimulated BV-2
microglial cells at higher concentrations (10 and 20 µM;)
(Fig. 2A and B) compared with the
Pam3CSK4-treated control cells. The secretion of TNF-α was also
significantly suppressed by curcumin (Fig. 2C) compared with the level in the
Pam3CSK4-treated control cells, however only at the highest
concentration of 20 µM. The mRNA expression levels of iNOS
and COX-2 were significantly increased following exposure to
Pam3CSK4 compared with the negative control group, but
significantly decreased compared with Pam3CSK4-treated control
cells following treatment with 10 and 20 µM curcumin
(Fig. 2D and E). Curcumin also
notably decreased the protein expression levels of COX-2 in
Pam3CSK4-induced BV-2 cells (Fig.
2F).
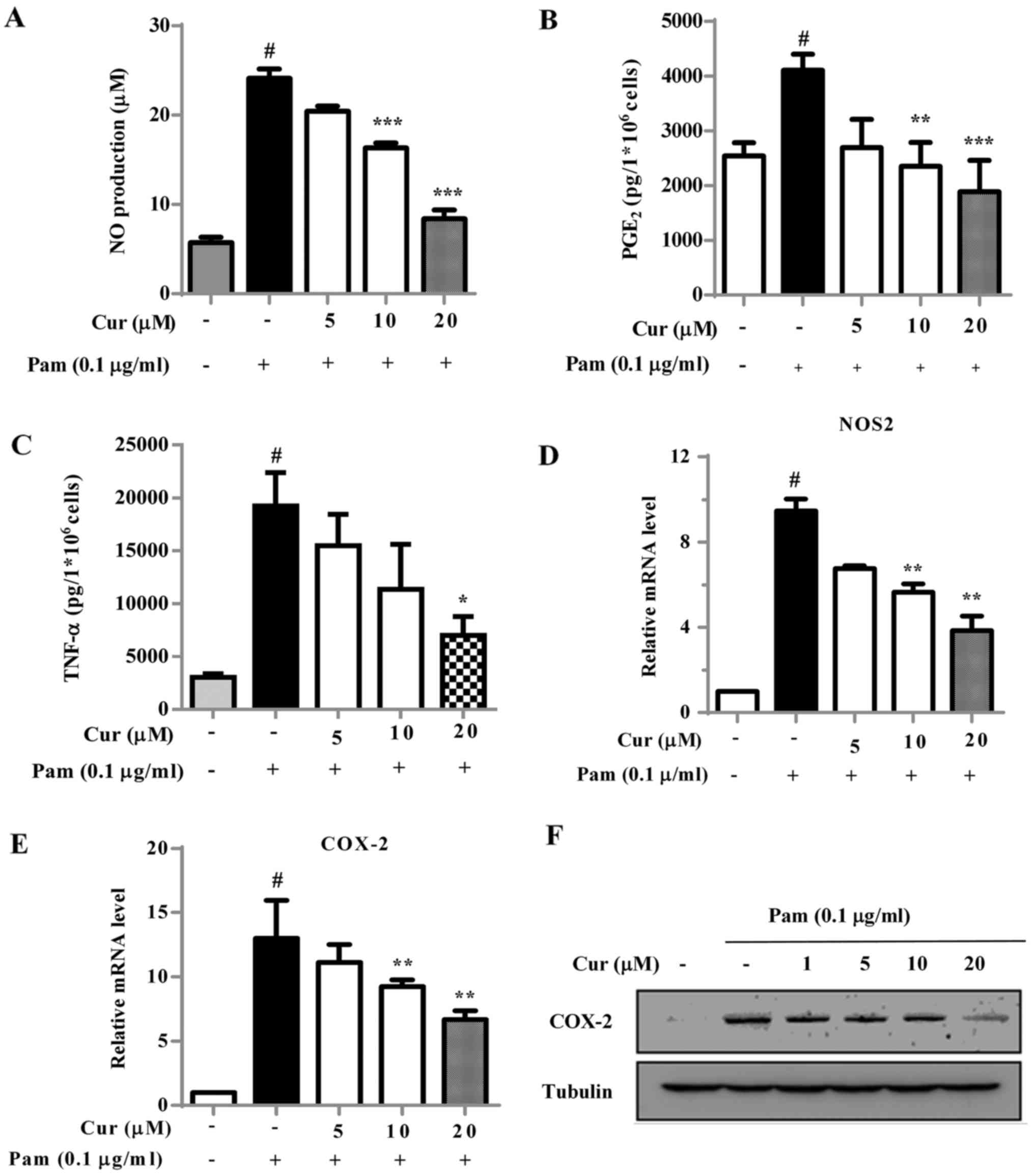 | Figure 2Effect of Cur on Pam-induced NO,
PGE2, TNF-α secretion, iNOS and COX-2 expression in BV-2
cells. Cells were treated with Cur at increasing doses (0–20
µM) and stimulated with Pam (0.1 µg/ml). (A) Nitrite
levels were measured in the collected media supernatant using the
Griess method. The secretion of (B) PGE2 and (C) TNF-α
were determined by ELISA. The mRNA expression levels of (D) iNOS,
encoded by NOS2, and (E) COX-2 were detected using reverse
transcription-quantitative polymerase chain reaction analysis. (F)
The expression of COX-2 was detected using western blot analysis.
Tubulin was used as a control. Data were presented as the mean ±
standard deviation from three independent experiments.
#P<0.01 vs. negative control; *P<0.05,
**P<0.01 and ***P<0.001 vs. the Pam3CSK4-treated
control. Pam, Pam3CSK4; Cur, curcumin; TNF, tumor necrosis factor;
PGE2, prostaglandin E2; COX-2,
cyclooxygenase-2; iNOS, inducible nitric oxide synthase; NO, nitric
oxide; NOS2, nitric oxide synthase 2; ELISA, enzyme-linked
immunosorbent assay. |
Treatment with curcumin causes an
increase of HO-1 in microglial cells
To assess the association of HO-1 with the
anti-neuroinflammatory properties of curcumin, the effect of
curcumin on HO-1 induction at the transcriptional and translational
levels was investigated. Treatment of BV-2 cells with curcumin
significantly increased HO-1 mRNA levels compared with the
untreated cells, with a maximal effect being observed at 4 h
(Fig. 3A). Curcumin increased the
HO-1 mRNA in a dose-dependent manner with significant increases
observed at 10 and 20 µM (Fig.
3B) compared with the untreated cells. The protein expression
level of HO-1 was measured by western blot analysis and similar
results were observed (Fig. 3C and
D). The protein expression of HO-1 was notably increased at 8 h
following treatment and at the higher concentrations of
curcumin.
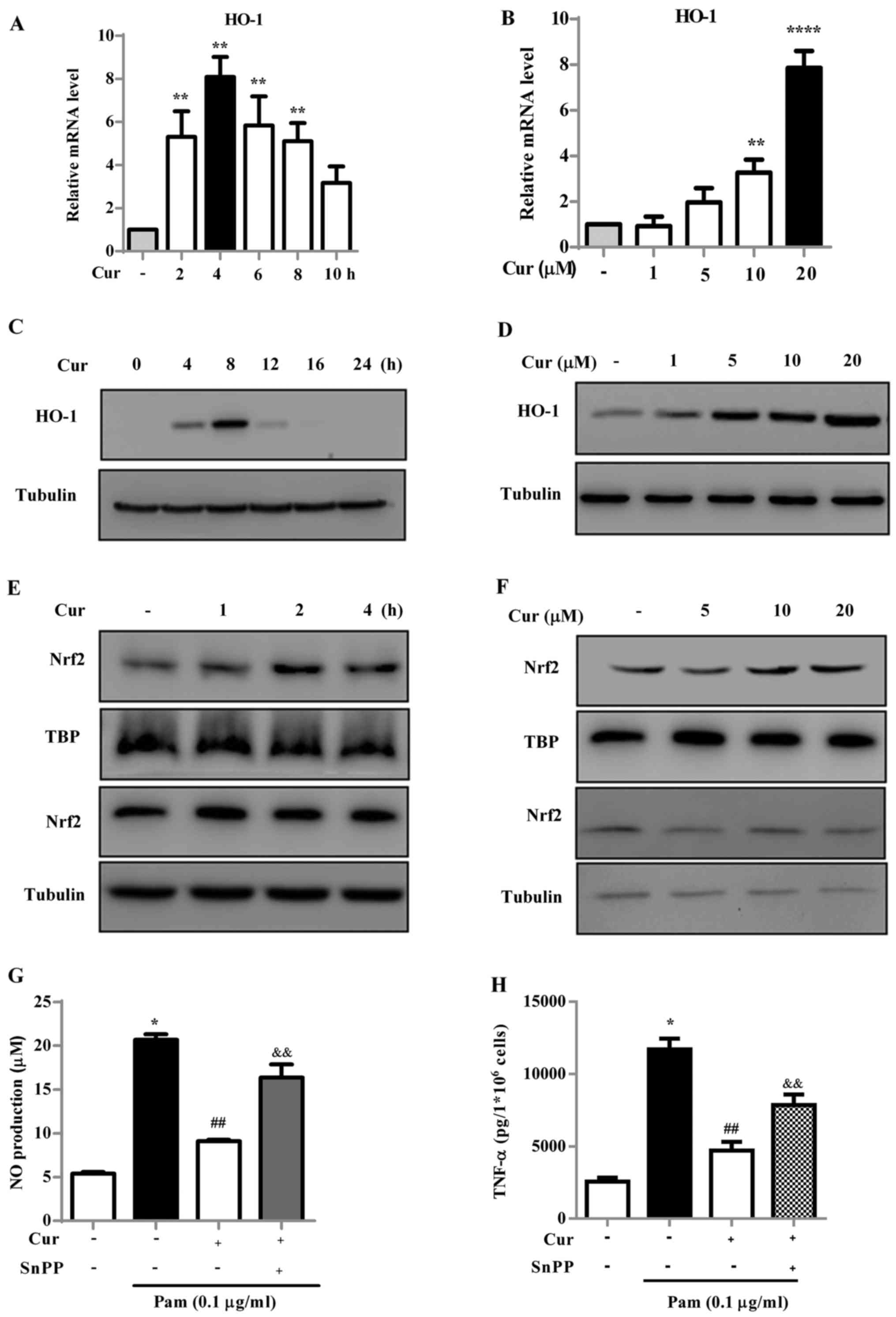 | Figure 3Cur induces HO-1 production through
the Nrf2/antioxidant response element signaling pathway in BV-2
microglial cells. (A) BV-2 cells were incubated with 20 µM
Cur for the indicated times (2, 4, 6, 8 and 10 h) or (B) cells were
incubated with increasing doses of Cur (5, 10 and 20 µM) for
4 h and the relative HO-1 mRNA expression level was measured using
reverse transcription-quantitative polymerase chain reaction. (C)
Cells were treated with 20 µM Cur for the indicated times
(0–24 h) or (D) various doses of Cur (0–20 µM) for 8 h.
Total cellular extracts were harvested and examined for HO-1
expression using western blot analysis. Tubulin was used as the
loading control for each lane. (E) Cells were treated with 20
µM Cur for the indicated times (0–24 h) or (F) various doses
of Cur (0–20 µM) for 2 h. Nuclear extracts were prepared and
examined for Nrf2 expression using western blot analysis. TBP and
tubulin was detected as a loading control. Cells were pretreated
with SnPP (HO-1 inhibitor, 20 µM) for 30 min, and then
treated with Cur in the presence or absence of Pam for 16 h. The
release of (G) NO and (H) TNF-α were determined. Three independent
experiments were performed and the data are presented as the mean ±
standard deviation. *P<0.05, **P<0.01
and ****P<0.0001 vs. negative control;
##P<0.01 vs. Pam-treated control;
&&P<0.01 vs. Cur ± Pam-treated group. Cur,
curcumin; HO, heme oxygenase; TBP, TATA-binding protein; NO, nitric
oxide; SnPP, protoporphyrin IX; Nrf2, nuclear factor
(erythroid-derived 2)-like 2; TNF, tumor necrosis factor; Pam,
Pam3CSK4. |
To investigate the effect of curcumin on Nrf2
nuclear translocation, which is associated with HO-1 induction, the
accumulation of Nrf2 in the nucleus was measured following
treatment with curcumin (Fig. 3E and
F). The quantity of Nrf2 that translocated into the nucleus
notably increased following treatment with curcumin, compared with
the control group. The maximum effect was observed at 2 h following
curcumin treatment and was most notable at the higher
concentrations of curcumin. These results suggest that curcumin
increased the transcription and translation of HO-1 through
activation of the Nrf2 transcription factor.
To confirm the anti-neuroinflammatory effects of
curcumin in HO-1 induction, the level of inflammatory mediators, NO
and TNF-α, were measured in Pam3CSK4-stimulated BV-2 microglial
cells incubated with or without the HO-1 inhibitor SnPP (20
µM) (Fig. 3G and H). The
cells were incubated with curcumin with or without SnPP for 0.5 h,
and then exposed to Pam3CSK4 for 16 h. The secretion of NO and
TNF-α were significantly increased following treatment with
Pam3CSK4 compared with the negative control; whereas there was a
significant decrease in NO and TNF-α production following treatment
with curcumin (20 µM) compared with cells treated with
Pam3CSK4 only. However, NO and TNF-α secretion were significantly
increased in cells co-treated with SnPP (20 µM) and
curcumin, compared with cells treated with curcumin only. These
results suggest that HO-1 is associated with the inhibitory effect
of curcumin on the Pam3CSK4-mediated release of inflammatory
mediators.
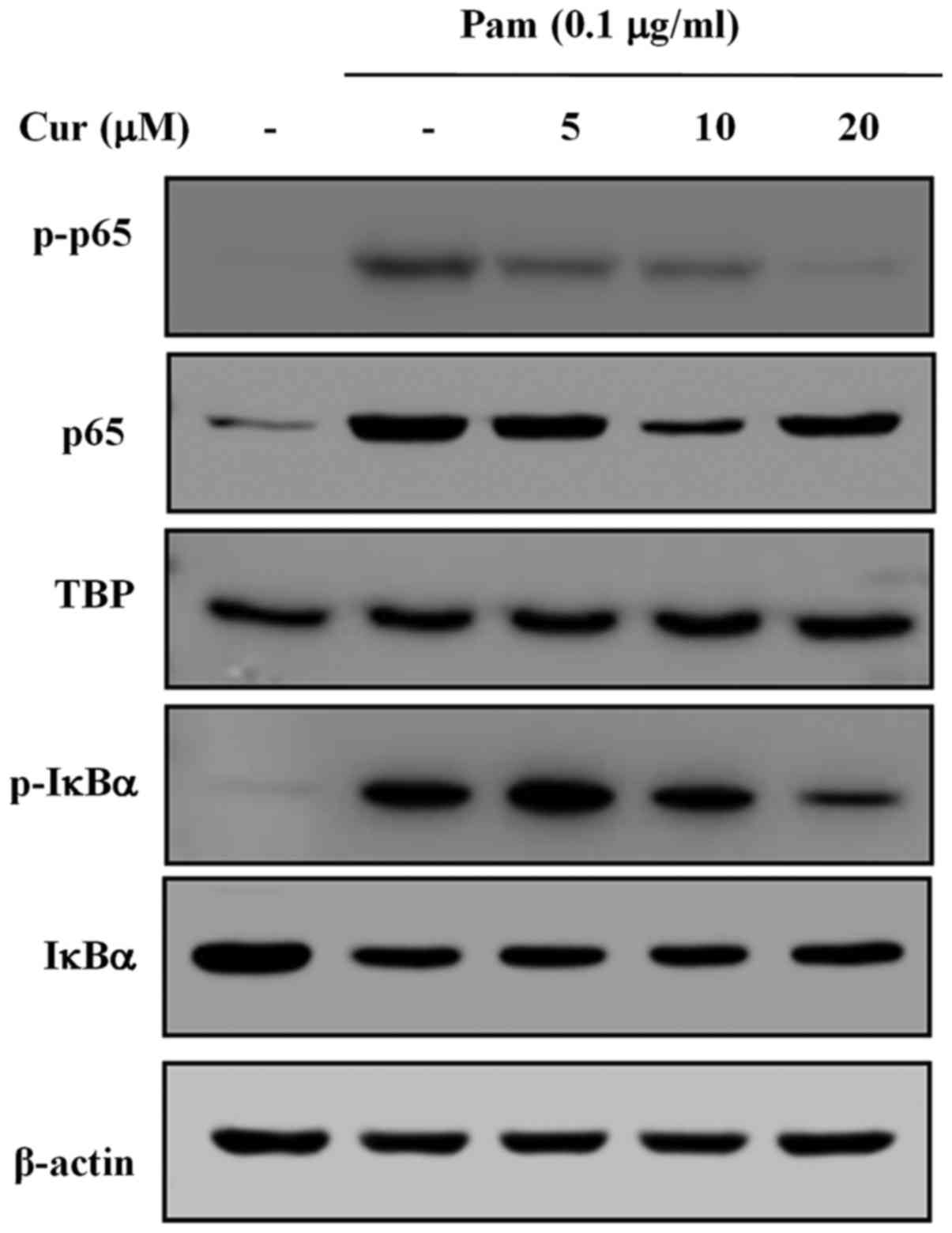
Treatment with curcumin reduces p65
translocation and IκBα degradation
Major cell signaling pathways, including those
associated with MAPKs, are activated by Toll-like receptors (TLRs)
in macrophage cells, which in turn leads to the induction of
transcription factors, such as NF-κB (23). The translocation of p65 and the
phosphorylation of IκBα in Pam3CSK4-stimulated BV-2 microglial
cells was investigated (Fig. 4).
The cells were stimulated with Pam3CSK4 (0.1 µg/ml) and
treated with curcumin at the indicated doses (0–20 µM). The
phosphorylation of p65 was markedly increased following stimulation
with Pam3CSK4, but gradually and dose-dependently decreased
following treatment with curcumin. Similarly, the phosphorylation
of IκBα was induced by treatment with Pam3CSK4, but recovered
dose-dependently when treated with curcumin. These results indicate
that treatment with curcumin reduces the translocation of p65 into
the nucleus and also reduces the degradation of IκBα.
Activation of MAPKs and the Akt signaling pathway
are crucial for the regulation of inflammatory mediators in
activated microglia (24). The
effect of curcumin on Pam3CSK4-induced phosphorylation of JNK,
ERK1/2, p38 and Akt in BV-2 cells was also investigated (Fig. 5). Exposure of BV-2 cells to
Pam3CSK4 for 30 min markedly increased the phosphorylation of JNK,
ERK, p38 and Akt. Treatment with curcumin notably reduced the
Pam3CSK4-induced phosphorylation of Akt and p38, which are
important upstream signaling molecules in inflammatory responses
mediated by activated microglial cells (25). The inhibition of p38 and JNK with
specific inhibitors significantly decreased the mRNA expression
levels of iNOS compared with the Pam3CSK4-treated cells, and
reduced the secretion of NO (Fig. 5B
and C).
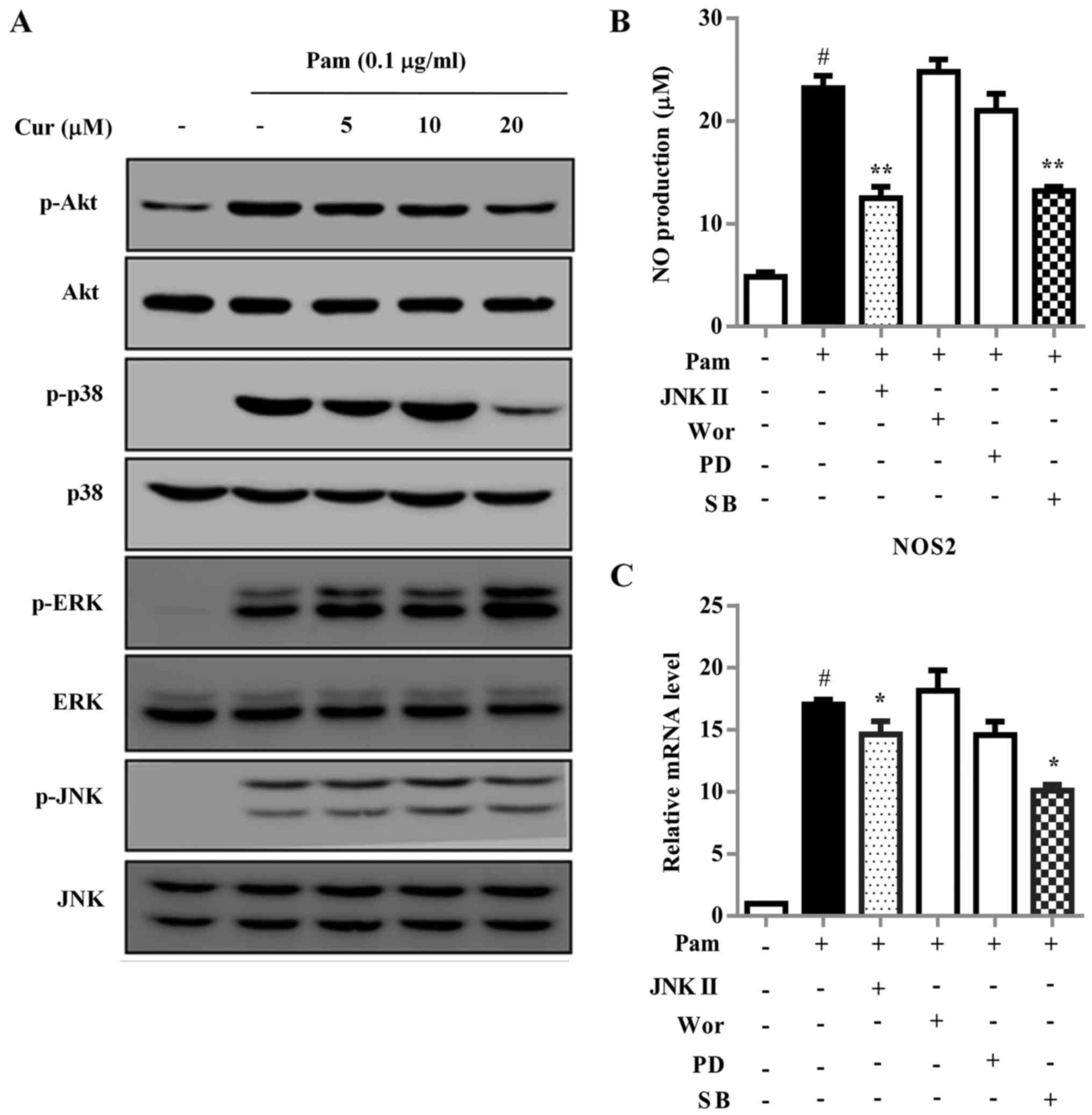 | Figure 5Cur inhibits neuroinflammation by
targeting the p38 mitogen-activated protein kinase signaling
pathway. (A) BV-2 cells were treated with Cur for 1 h followed by
stimulation with Pam for 0.5 h. Cell extracts were collected and
subjected to western blot analysis. BV-2 cells were treated with
JNK inhibitor (JNK II, 10 µM), Akt inhibitor (Wor, 5
µM), ERK inhibitor (PD98059, 10 µM) or p38 inhibitor
(SB230580, 10 µM) for 1 h, following treatment with Pam (0.1
µg/ml) for 16 h. The release of (B) NO and the expression
levels of inducible (C) NO synthase, encoded by NOS2, were
determined. Each bar represents the mean ± standard deviation from
three independent experiments. #P<0.01 vs. negative
control; *P<0.05 and **P<0.01 vs.
Pam-treated control. NO, nitric oxide; PD, PD98059; SB, SB203580;
Wor, Wortmannin; Pam, Pam3CSK4; Cur, curcumin; ERK, extracellular
signal-regulated kinase; JNK, c-Jun NH2-terminal protein kinase;
p-, phosphorylated; NOS2, nitric oxide synthase 2. |
Discussion
TLR2 is a transmembrane PRR that responds to
exogenous and endogenous ligands (3). TLRs are expressed in a variety of
immune cells and mediate neuroimmunity (26). The majority of exogenous ligands
recognized by TLR2 are lipoproteins or lipoteichoic acid from
gram-positive bacteria, whereas TLR4 predominantly identifies LPS
endotoxins from gram-negative bacteria (27). Bacterial endotoxins, endogenous
compounds and synthesized compounds, including peptidoglycan,
Pam3CSK4, lipoteichoic acid (LTA) and β-amyloid are collectively
known as danger-associated molecular patterns (DAMPs) (28). DAMPs are able to activate TLR2
under chronic pain conditions and neurodegenerative disorders
(28). A previous study using
TLR2 knock-out mice revealed that the expression of the TLR2
receptor is required for the development and persistence of
α-synuclein accumulation, α-synuclein-mediated neurodegeneration
and experimental spontaneous pain due to peripheral nerve injury
(26). Additionally, TLR2
activation is critical in the promoting brain injury (26). A previous study has revealed that
the activation of TLR2 in microglia lead to the development of
diseases associated with hypothalamic inflammation through the
NF-κB and COX-2 signaling pathways (29). Therefore, controlling the
activation of TLR2 and its downstream signaling pathways may be
beneficial for the attenuation of neuroinflammation associated
neuroinjuries. Activation of TLR2 heterodimers (with TLR1 or TLR6)
primarily requires the myeloid differentiation primary response
gene 88-dependent signaling pathways, including NF-κB and MAPKs
(28).
In the present study, the anti-neuroinflammatory
properties of curcumin were investigated in Pam3CSK4-induced
microglial cells. The activation of microglia by Pam3CSK4 markedly
mediated the M1/M2 ratio to imitate the M1 phenotype of microglial
cells identified in AD murine models (4). The present study revealed that
Pam3CSK4 notably increased the secretion of NO and TNF-α, and the
mRNA expression levels of iNOS in microglial cells, which are all
known markers of the M1 subtype of microglial cells (30). In addition, NO and PGE2
are considered to contribute to a number of physiological and
pathological processes, including ischemia, trauma, multiple
sclerosis, PD and AD (31).
Overexpression of NO and PGE2 is also associated with
the upregulation of iNOS and COX-2 (31). Inflammatory cytokines, including
TNF-α and IL-1β, have been demonstrated to be significantly
increased in neurodegenerative disorders (28). Recent study suggested that the
inhibition of these cytokines was beneficial for the treatment of
neurodegenerative diseases (28).
In the present study, it was identified that curcumin had a
significant anti-inflammatory effect through the inhibition of
inflammatory mediators. Furthermore, the suppression of
inflammatory molecules by curcumin was reversed following treatment
with SnPP, an HO-1 inhibitor. Curcumin reduced the secretion of
inflammatory molecules by upregulating HO-1 and Nrf2
expression.
The immunomodulatory role of HO-1 was first
described in peripheral macrophages, where HO-1 by-products,
biliverdin and carbon monoxide, inhibited NAD(P)H oxidase and TLR4
signaling, thus suppressed macrophage activation (16). Several previous studies have
employed various models to demonstrate the neuroprotective and
anti-neuroinflammatory effects of HO-1 against oxidative stress in
the CNS (14,16). Overexpression of HO-1 in microglia
and macrophages was identified as providing anti-inflammatory and
neuroprotective properties against secondary insults, including
oxidative stress in human and rat traumatic brain injuries
(32). Furthermore, a previous
study has also suggested a positive correlation between HO-1
expression and IL-10 release (33). In the present study, curcumin
induced the expression of HO-1 at the mRNA and protein level in a
dose-dependent manner, with maximal effects at 4 and 8 h,
respectively. HO-1 is a phase II antioxidant enzyme induced by
translocation of the transcription factor Nrf2 (34). The effect of Nrf2 has been
described in various in vivo models where Nrf2-knockout mice
demonstrated significant activation of microglial cells compared
with wild-type mice (35). In
addition, LPS induced an enhanced inflammatory response in Nrf2
knockout mice compared with the control mice (36). The results of the present study
demonstrated that curcumin increased the nuclear accumulation of
Nrf2 at 1 h following treatment and it reached a maximum at 2 h.
Therefore, the results indicated that HO-1 upregulation by the
Nrf2/ARE signaling pathway was associated with the inhibitory
effects of curcumin on the release of inflammatory mediators by
Pam3CSK4.
Curcumin also suppressed signaling pathways upstream
of the inflammatory mediators, such as NF-κB, which were associated
with the inflammatory responses triggered by TLR2 in BV-2 cells.
NF-κB is a master regulator of microglial inflammatory responses
(34). A previous study has
revealed crosstalk between NF-κB and Nrf2, which has a circular
regulatory effect (37). In
Nrf2−/− mouse embryonic fibroblast cells, the absence of
Nrf2 exacerbates NF-κB activity by enhancing the degradation of
IκBα, which leads to an increase in cytokine production (15,35). Additionally, the NF-κB subunit p65
may exert a negative effect on Nrf2 activity and ARE-linked gene
expression (37). During
neuroinflammation, NF-κB activation induces the expression of
inflammatory cytokines, including iNOS, NO, PGE2, COX-2
and ROS to trigger an over-activated inflammatory response, which
leads to progressive neuronal damage (31). Therefore, modulation of NF-κB
activation is considered to be a useful method of controlling
microglial activation. It has been previously reported that
curcumin inhibits NF-κB activation in various immune cell types,
thus it inhibits the production of inflammatory mediators (17). The results of the present study
also indicated that curcumin decreased nuclear translocation and
transactivation of NF-κB following Pam3CSK4 stimulation in
microglial cells.
Previous studies suggested that activation of the
phosphoinositide 3-kinase (PI3K)/Akt-dependent signaling pathway
promoted activation of M1 microglia, leading to the expression of
inflammatory mediators and neuronal injury (34,38). In addition, the MAPK signaling
pathway is also stimulated by increased oxidative stress, including
LPS, LTA and other agonists of TLR2 or TLR4 (24,39). The PI3K/Akt and MAPK signaling
pathways have been revealed to upregulate the gene expression of
iNOS and COX-2 in microglia activated by different stimuli, such as
LPS (24). A previous study
revealed that casticin inhibited the expression of COX-2 and iNOS
through inhibition of MAPK signaling pathways in macrophage
(39). The results of the present
study are consistent with these previous findings. Pretreatment of
microglial cells with curcumin notably decreased the phosphorylated
MAPKs and Akt, particularly p-p38 and Akt, whereas it did not
change the total MAPK and Akt levels. Furthermore, the p38 and JNK
inhibitors suppressed the secretion of NO and the mRNA expression
of iNOS. These findings illuminate the fundamental role of p38 MAPK
in the anti-neuroinflammatory effect of curcumin.
In conclusion, the results of the present study
revealed that curcumin exerted anti-neuroinflammatory effects in
Pam3CSK4-stimulated microglial cells. These results suggest that
the anti-neuroinflammatory role of curcumin is primarily through
inhibition of the p38 MAPK and NF-κB signaling pathways, and
induction of HO-1 by the Nrf2/ARE signaling pathway, thereby
decreasing the production of inflammatory mediators. These results
suggest that curcumin may be a novel candidate for the treatment of
chronic neuroinflammatory diseases by reducing the over-activation
of microglial cells.
Acknowledgments
The present study was supported by the Research
Grants of the National Natural Science Foundation of China (grant
no. 81401299), the Key Project of Department of Education of
Guangdong Province (grant no. 2015KTSCX120), the Shenzhen Peacock
Plan (grant nos. 827-000129, 827-000209, 827-000107 and
KQTD20140630100746562), the Shenzhen Research Grant (grant nos.
JCYJ20150324140036854 and JCYJ20160422091658982) and Shenzhen
Science and Technology Project (grant no. 20160422091658982).
Abbreviations:
|
AD
|
Alzheimer's disease
|
|
CNS
|
central nervous system
|
|
TNF-α
|
tumor necrosis factor-α
|
|
TLR
|
Toll-like receptor
|
|
MAPK
|
mitogen-activated protein kinase
|
|
JNK
|
c-Jun NH2-terminal protein kinase
|
|
ERK
|
extracellular signal-regulated
kinase
|
|
NF-κB
|
nuclear factor-κB
|
|
PGE2
|
prostaglandin E2
|
|
HO-1
|
heme oxygenase-1
|
|
Nrf2
|
nuclear factor (erythroid-derived
2)-like 2
|
References
|
1
|
Spangenberg EE and Green KN: Inflammation
in Alzheimer's disease: Lessons learned from microglia-depletion
models. Brain Behav Immun. 61:1–11. 2017. View Article : Google Scholar
|
|
2
|
De Hoz R, Salobrar-Garcia E, Salazar JJ,
Roias B, Ajoy D, Lopez-Cuenca I, Rojas P, Trivino A and Ramierz JM:
The role of microglial in retinal neurodegeneration: Alzheimer's
disease, Parkinson, and glaucoma. Front Aging Neurosci. 9:2142017.
View Article : Google Scholar
|
|
3
|
Larochelle A, Bellavance MA and Rivest S:
Role of adaptor protein MyD88 in TLR-mediated preconditioning and
neuro-protection after acute excitotoxicity. Brain Behav Immun.
46:221–231. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Tang Y and Le W: Differential role of M1
and M2 microglia in neurodegenerative diseases. Mol Neurobiol.
53:1181–1194. 2016. View Article : Google Scholar
|
|
5
|
Nakagawa Y and Chiba K: Diversity and
plasticity of microglial cells in psychiatric and neurological
disorders. Pharmacol Ther. 154:21–35. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kaur CG, Rathnasamy and Ling EA: Biology
of microglial in the developing Brain. J Neuropathol Exp Neurol.
76:736–753. 2017.PubMed/NCBI
|
|
7
|
Moss DW and Bates TE: Activation of murine
microglial cell lines by lipopolysaccharide and interferon-gamma
causes NO-mediated decreases in mitochondrial and cellular
function. Eur J Neurosci. 13:529–538. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Rangarajan P, Karthikeyan A and Dheen ST:
Role of dietary phenols in mitigating microglia-mediated
neuroinflammation. Neuromolecular Med. 18:453–464. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Acharyya S, Villalta SA, Bakkar N,
Bupha-Intr T, Janssen PM, Carathers M, Li ZW, Beg AA, Ghosh S,
Sahenk Z, et al: Interplay of IKK/NF-kappaB signaling in
macrophages and myofibers promotes muscle degeneration in Duchenne
muscular dystrophy. J Clin Invest. 117:889–901. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Sweeney SE and Firestein GS: Signal
transduction in rheumatoid arthritis. Curr Opin Rheumatol.
16:231–237. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Jangra A, Kwatra M, Singh T, Pant R,
Kushwah P, Ahmed S, Dwivedi D, Saroha B and Lahkar M: Edaravone
alleviates cisplatin-induced neurobehavioral deficits via
modulation of oxidative stress and inflammatory mediators in the
rat hippo-campus. Eur J Pharmacol. 791:51–61. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Koh K and Kim J, Jang YJ, Yoon K, Cha Y,
Lee HJ and Kim J: Transcription factor Nrf2 suppresses LPS-induced
hyperactivation of BV-2 microglial cells. J Neuroimmunol.
233:160–167. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Talalay P: Chemoprotection against cancer
by induction of phase 2 enzymes. Biofactors. 12:5–11. 2000.
View Article : Google Scholar
|
|
14
|
Zhang J, Fu B, Zhang X, Zhang L, Bai X,
Zhao X, Chen L, Cui L, Zhu C, Wang L, et al: Bicyclol upregulates
transcription factor Nrf2, HO-1 expression and protects rat brains
against focal ischemia. Brain Res Bull. 100:38–43. 2014. View Article : Google Scholar
|
|
15
|
Innamorato NG, Rojo AI, García-Yagüe AJ,
Yamamoto M, de Ceballos ML and Cuadrado A: The transcription factor
Nrf2 is a therapeutic target against brain inflammation. J Immunol.
181:680–689. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Parada E, Buendia I, Navarro E, Avendaño
C, Egea J and López MG: Microglial HO-1 induction by curcumin
provides antioxidant, antineuroinflammatory, and glioprotective
effects. Mol Nutr Food Res. 59:1690–1700. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Kunnumakkara AB, Bordoloi D, Padmavathi G,
Monisha J, Roy NK, Prasad S and Aggarwal BB: Curcumin, the golden
nutraceutical: Multitargeting for multiple chronic diseases. Br J
Pharmacol. 174:1325–1348. 2017. View Article : Google Scholar
|
|
18
|
Tsai YM, Chien CF, Lin LC and Tsai TH:
Curcumin and its nano-formulation: The kinetics of tissue
distribution and blood-brain barrier penetration. Int J Pharm.
416:331–338. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Garcia-Alloza M, Borrelli LA, Rozkalne A,
Hyman BT and Bacskai: Curcumin labels amyloid pathology in vivo,
disrupts existing plaques, and partially restores distorted
neurites in an Alzheimer mouse model. J Neurochem. 102:1095–1104.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Prakobwong S, Khoontawad J, Yongvanit P,
Pairojkul C, Hiraku Y, Sithithaworn P, Pinlaor P, Aggarwal BB and
Pinlaor S: Curcumin decreases cholangiocarcinogenesis in hamsters
by suppressing inflammation-mediated molecular events related to
multistep carcinogenesis. Int J Cancer. 129:88–100. 2011.
View Article : Google Scholar
|
|
21
|
Zhou J, Miao H, Li X, Hu Y, Sun H and Hou
Y: Curcumin inhibits placental inflammation to ameliorate
LPS-induced adverse pregnancy outcomes in mice via upregulation of
phosphorylated Akt. Inflamm Res. 66:177–185. 2017. View Article : Google Scholar
|
|
22
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
23
|
Zhou D, Huang C, Lin Z, Zhan S, Kong L,
Fang C and Li J: Macrophage polarization and function with emphasis
on the evolving roles of coordinated regulation of cellular
signaling pathways. Cell Signal. 26:192–197. 2014. View Article : Google Scholar
|
|
24
|
Jung JS, Choi MJ, Lee YY, Moon BI, Park JS
and Kim HS: Suppression of lipopolysaccharide-induced
neuroinflammation by Morin via MAPK, PI3K/Akt, and PKA/HO-1
signaling pathway modulation. J Agric Food Chem. 65:373–382. 2017.
View Article : Google Scholar
|
|
25
|
Brown GC: Nitric oxide and neuronal death.
Nitric Oxide. 23:153–165. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Dzamko N, Gysbers A, Perera G, Bahar A,
Shankar A, Gao J, Fu Y and Halliday GM: Toll-like receptor 2 is
increased in neurons in Parkinson's disease brain and may
contribute to alpha-synuclein pathology. Acta Neuropathol.
133:303–319. 2017. View Article : Google Scholar
|
|
27
|
Fallarino F, Gargaro M, Mondanell G,
Downer EJ, Hossain MJ and Gran B: Delineating the role of Toll-like
receptors in the neuro-inflammation model EAE. Methods Mol Biol.
1390:383–411. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hossain MJ, Tanasescu R and Gran B: Innate
immune regulation of autoimmunity in multiple sclerosis: Focus on
the role of Toll-like receptor 2. J Neuroimmunol. 304:11–20. 2017.
View Article : Google Scholar
|
|
29
|
Jin S, Kim JG, Park JW, Koch M, Horvath TL
and Lee BJ: Hypothalamic TLR2 triggers sickness behavior via a
microglia-neuronal axis. Sci Rep. 6:294242016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Geloso MC, Corvino V, Marchese E, Serrano
A, Michetti F and D' Ambrosi N: The dual role of microglia in ALS:
Mechanisms and therapeutic approaches. Front Aging Neurosci.
9:2422017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Xia Q, Hu Q, Wang H, Yang H, Gao F, Ren H,
Chen D, Fu C, Zheng L, Zhen X, et al: Induction of
COX-2-PGE2 synthesis by activation of the MAPK/ERK
pathway contributes to neuronal death triggered by TDP-43-depleted
microglia. Cell Death Dis. 6:e17022015. View Article : Google Scholar
|
|
32
|
Liu Y and Zhang Z, Luo B, Schluesener HJ
and Zhang Z: Lesional accumulation of heme oxygenase-1+
microglia/macrophages in rat traumatic brain injury. Neuroreport.
24:281–286. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Fernández P, Guillén MI, Gomar F and
Alcaraz MJ: Expression of heme oxygenase-1 and regulation by
cytokines in human osteoarthritic chondrocytes. Biochem Pharmacol.
66:2049–2052. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Jayasooriya RG, Lee KT, Choi YH, Moon SK,
Kim WJ and Kim GY: Antagonistic effects of acetylshikonin on
LPS-induced NO and PGE2 production in BV2 microglial
cells via inhibition of ROS/PI3K/Akt-mediated NF-κB signaling and
activation of Nrf2-dependent HO-1. In Vitro Cell Dev Biol Anim.
51:975–986. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Rojo AI, Innamorato NG, Martín-Moreno AM,
De Ceballos ML, Yamamoto M and Cuadrado A: Nrf2 regulates
microglial dynamics and neuroinflammation in experimental
Parkinson's disease. Glia. 58:588–598. 2010. View Article : Google Scholar
|
|
36
|
Kobayashi EH, Suzuki T, Funayama R,
Nagashima T, Hayashi M, Sekine H, Tanaka N, Moriguchi T, Motohashi
H, Nakayama K, et al: Nrf2 suppresses macrophage inflammatory
response by blocking proinflammatory cytokine transcription. Nat
Commun. 7:116242016. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Wardyn JD, Ponsford AH and Sanderson CM:
Dissecting molecular cross-talk between Nrf2 and NF-kappaB response
pathways. Biochem Soc Trans. 43:621–626. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Chantong B, Kratschmar DV, Lister A and
Odermatt A: Dibutyltin promotes oxidative stress and increases
inflammatory mediators in BV-2 microglia cells. Toxicol Lett.
230:177–187. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Liou CJ, Len WB, Wu SJ, Lin CF, Wu XL and
Huang WC: Casticin inhibits COX-2 and iNOS expression via
suppression of NF-kappaB and MAPK signaling in
lipopolysaccharide-stimulated mouse macrophages. J Ethnopharmacol.
158:310–316. 2014. View Article : Google Scholar
|