Introduction
Tongue squamous cell carcinoma (TSCC) is the most
common malignancy in the oral cavity. TSCC is more aggressive than
other cancers of the oral cavity because of the propensity for
rapid local invasion and spread. Unfortunately, the mortality rate
of TSCC has not improved significantly and the 5-year survival rate
has remained at <50% in the past 30 years despite increasing
advances in therapeutic modalities. The major cause of
TSCC-associated mortalities is local/regional relapse and
metastasis. Previous studies demonstrated that epithelial
mesenchymal transition (EMT) has a pivotal role in cancer invasion
and metastasis (1,2). EMT is characterized by loss of cell
polarity and adhesion, enhancement of cell motility and acquisition
of a mesenchymal phenotype. It has been confirmed that reduced
E-cadherin expression is a hallmark of EMT. The loss of E-cadherin
has been linked with enhanced cell invasion and metastasis in oral
SCC (3–5). Our previous studies demonstrated
that E-cadherin expression was decreased in the budding tumor cells
located ahead of invasive tumor front in TSCC, and expression was
positively correlated with lymph node metastasis (6). Further studies confirmed that
Snail2, histone-lysine N-methyltransferase EZH2 and microRNA-138
(7–9) regulate E-cadherin expression at the
transcriptional and post-transcriptional levels. However, the
molecular mechanisms underlining EMT process are still not fully
understood in TSCC.
β2-adrenergic receptor (β2-AR) is a seven
transmembrane receptor (also known as G protein-coupled receptor)
required for the physiological response to adrenaline and
noradrenaline. Notably, several studies indicated that the β2-AR
pathway has an important role in solid cancer initiation and
progression, including in breast cancer, malignant melanoma,
prostate cancer and gastric cancer (10–13). More recent studies demonstrated
that activation of β2-AR signaling was involved in EMT, and
contributed to invasion and metastasis in colorectal adenocarcinoma
and gastric cancer (14–16). However, the expression of β2-AR
and its effect on TSCC progression have not well been documented.
In this study, deregulation of β2-AR in TSCC was examined to
investigate its roles in EMT and TSCC progression.
Materials and methods
Reagents and antibodies
Anti-β2-AR (cat. no. ab182136) was purchased from
Abcam (Cambridge, UK), anti-interleukin-6 (IL-6; cat. no. 12153),
anti-E-cadherin (cat. no. 9101s), anti-vimentin (cat. no. 9102s),
anti-Snail (cat. no. 3879), anti-phospho-signal transducer and
activator of transcription 3 (Stat3; cat. no. 12640) and anti-GAPDH
(cat. no. 2118s) antibodies were purchased from Cell Signaling
Technology, Inc. (Danvers, MA, USA). Human IL-6 antibody (cat. no.
AF-206-NA) were purchased from R&D Systems, Inc. (Minneapolis,
MN, USA). Non-selective β-AR blocker, propranolol hydrochloride
(cat. no. 318-98-9), non-specific β-AR agonist, isoproterenol
hydrochloride (cat. no. 51-30-9), the β1-AR antagonist, metoprolol
tartrate (cat. no. 56392-17-7) and the β2-AR antagonist ICI-118,551
hydrochloride (cat. no. 72795-19-8), were purchased from
Sigma-Aldrich (Merck KGaA, Darmstadt, Germany). Anti-rabbit IgG,
HRP-linked secondary antibody (cat. no. 7074s) was purchased from
Cell Signaling Technology.
Patients and tissue samples
All tissue samples were collected between January
2007 and December 2009, obtained from the First Affiliated
Hospital, Sun Yat-Sen University (Guangzhou, China). Patients with
TSCC (n=75) were enrolled in this study. Adjacent non-cancerous
tissue samples (n=20) and TSCC specimens (n=75) were used for
immunohistochemical assessment. All tissue samples used in this
study were used following the guidelines set by the Institution
Review Board of Sun Yat-Sen University. All patients received
radical surgery and none received any form of adjuvant therapy
prior to surgery. The tumor extent was classified based on the TNM
system by Union for International Cancer Control, and the tumor
grade was classified following the World Health Organization
classification of histological differentiation (6). Survival was calculated according to
the date of surgery and the date of the last follow-up (or death).
Informed consent was signed by all patients for the use of their
tissue and clinical data in clinical analysis and research studies
prior to treatment.
Immunohistochemistry and staining
evaluation
Immunohistochemical staining was performed according
to standard protocols (7).
Samples were fixed in 4% paraformaldehyde for 24 h at room
temperature and embedded in paraffin. Briefly, 4 µm thick
tissue sections were dewaxed and hydrated routinely. Following
antigen retrieval, tissue sections were incubated with primary
antibodies at 4°C overnight. Samples were washed for five times
with phosphate-buffered saline, then incubated with secondary
antibody (1:200) at room temperature for 30 min. Tissues were
treated with 3,3′-diaminobenzidine (1:200) for 1 min at room
temperature and counterstained with hematoxylin for 10 sec at room
temperature. After dehydration, slides were observed and analyzed
using an Axioskop 40 (Carl Zeiss AG, Oberkochen, Germany)
microscope. Five fields were randomly selected for each specimen
under a light microscope at a magnification of ×400, and three
experienced observers analyzed the images. The scoring criteria
were according to a standard protocol described previously
(7). A final score >4 was
classified as high β2-AR expression and a score ≤4 was classified
as low β2-AR expression.
Cell cultures and treatments
Cal27 and SCC15 TSCC cell lines were obtained from
American Type Culture Collection (Manassas, VA, USA). Cells were
cultured in Dulbecco's modified Eagle's medium-F12 (Gibco: Thermo
Fisher Scientific, Inc., Waltham, MA, USA) supplemented with 10%
fetal bovine serum (HyClone; GE Healthcare Life Sciences, Logan,
UT, USA), 1% penicillin and streptomycin. All cells were incubated
at 37°C in a humidified atmosphere containing 5% CO2
with the culture media changed every 2 days. Cultured cells were
treated at 70 to 90% confluence and were serum-starved for 8 h
before treatment. Briefly, cells were treated for 48 h according
the groups as follows: i) Culture medium (control); ii) 1 µM
propranolol (Prop); iii) 1 µM propranolol + 1 µM
isoproterenol (Prop + ISO); iv) 1 µM metoprolol + 1
µM isoproterenol (Meto + ISO); v) 1 µM ICI-118,551 +
1 µM isoproterenol (ICI + ISO); and vi) 1 µM ISO.
Propranolol, metoprolol and ICI-118,551 were added in the culture
media 30 min before adding ISO.
IL-6/Stat3 signaling analysis
Cancer cells were cultured with medium containing
500 ng/ml IL-6 antibody (R&D Systems, Inc.) to neutralize and
block IL-6 signaling in TSCC cells.
Immunofluorescence and laser scanning
confocal microscopy
Samples were blocked with goat serum (Boster
Biological Technology, Wuhan, China) and incubated with the primary
antibody [E-cadherin (1:250), vimentin (1:250)] diluted in 5%
bovine serum albumin (BSA; Boster Biological Technology) at 4°C
overnight. Secondary antibody conjugated to fluorescein
isothiocyanate was used to bind to the primary antibody (1:200)
diluted in PBS at 37°C for 30 min. Nuclei were stained with DAPI
(1:250; Sigma-Aldrich; Merck KGaA) diluted in PBS at 37°C for 5
min. Samples were placed on glass coverslip. Cells were observed
using a confocal laser scanning microscope (Zeiss LSM 510 Meta;
Carl Zeiss AG).
Migration and invasion assay
Cell migration assay was evaluated by using wound
healing and Transwell membrane chambers (8 µm pore size;
Corning Inc., New York, NY, USA). The wound healing assay was
performed as previously described (16). Briefly, the cells were seeded in
12-well plates and treated with ISO at 90% confluence and normal
culture medium was added to the control cells. For Transwell
membrane chambers, cells were seeded into the upper chambers and
cultured with or without ISO for 36 h. For invasion assay, top
chambers coated with Matrigel (BD Biosciences, Franklin Lakes, NJ,
USA) at a concentration of 1.37 mg/ml. Upper chambers were fixed in
4% paraformaldehyde at room temperature for 30 min, stained using
0.1% crystal violet at room temperature for 5 min, then counted
using a Axioskop 40 (Carl Zeiss AG) microscope.
Western blot analysis
Cells were lysed on ice in 120 µl
radioimmunoprecipitation lysis buffer containing phosphatase
inhibitors (Sigma-Aldrich; Merck KGaA) for 30 min. Total proteins
(30 µg; determined by bicinchoninic acid assay) were loaded
and separated by 10% SDS-PAGE, and then transferred into
nitro-cellulose membrane (EMD Millipore, Billerica, MA, USA).
Membranes were blocked in 5% evaporated skimmed milk at room
temperature for 1 h and then incubated with primary antibody at 4°C
overnight. Primary antibodies (1:1,000) were diluted in 5%
evaporated skimmed milk at 4°C overnight. Then incubated the
membrane with horseradish peroxidase-conjugated secondary antibody
(1:2,000; diluted in 5% evaporated skimmed milk at room temperature
for 1 h; Cell Signaling Technology, Inc.) and observed using
chemiluminescence (EMD Millipore).
Statistical analysis
Data was presented as mean ± SD. All experiments
were performed at least three times and all statistical analyses
were performed using SPSS 13.0 software for Windows (SPSS, Inc.,
Chicago, IL, USA). The association between β2-AR expression and the
clinical pathological parameters was determined by χ2
test. The survival analysis was performed by the Kaplan-Meier
method. Student's t-test was used for statistical analysis.
P<0.05 was considered to indicate a statistically significant
difference. A Cox regression was used to analyze the association
between the survival time and the clinicopathological
variables.
Results
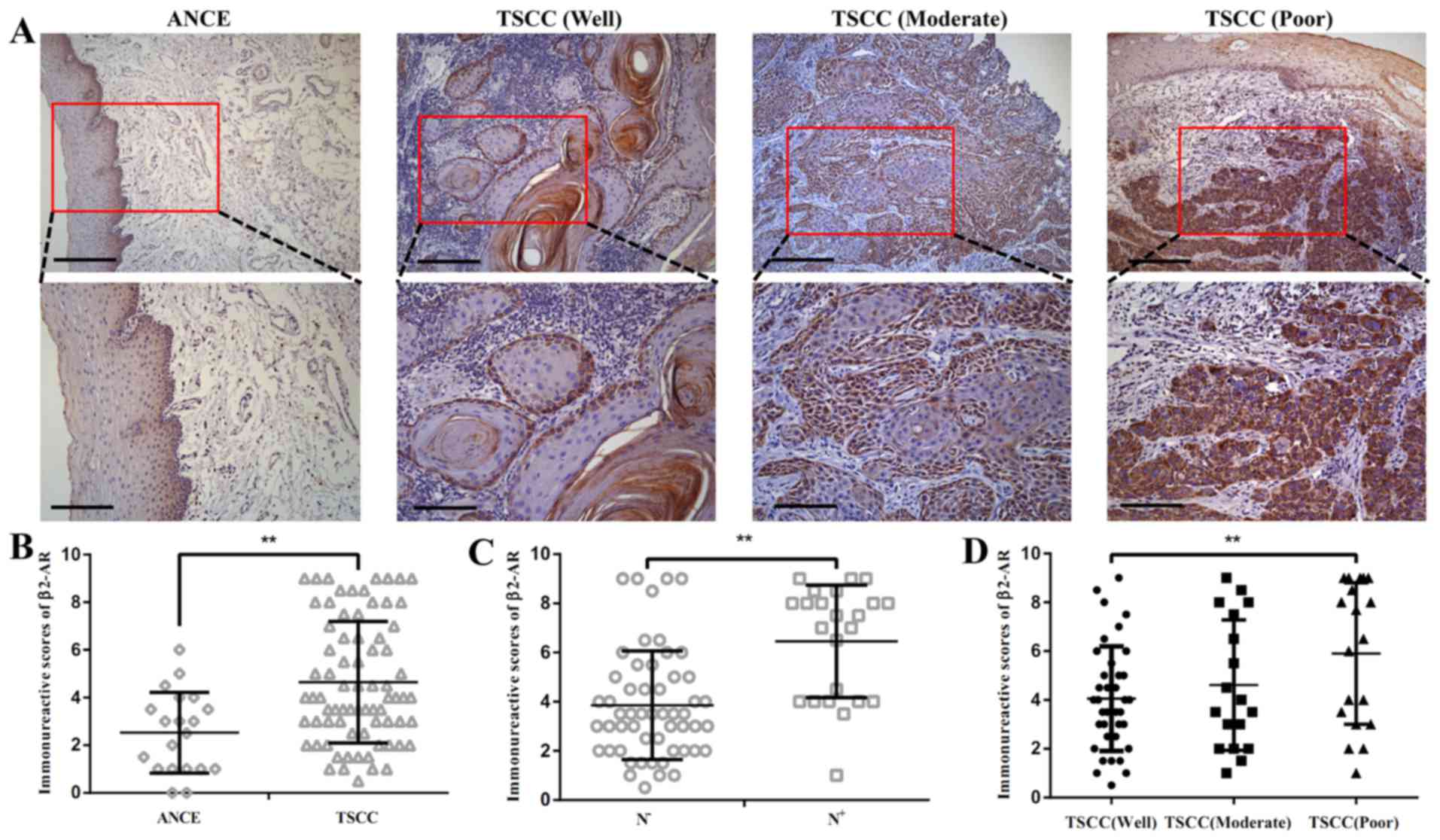
Expression of β2-AR in TSCC samples
The expression of p-β2-AR was detected by
immunohistochemistry in 75 cases with TSCC. The β2-AR was detected
in at low levels in adjacent non-cancerous epithelium (ANCE). In
primary tumor samples, predominant cytoplasm and membrane staining
of β2-AR was detected in cancer cells (Fig. 1A). The semi-quantitative analysis
revealed that β2-AR was overexpressed in TSCC compared with ANCE
(Fig. 1B). Enhanced expression of
β2-AR was also observed in TSCC with lymph node metastasis compared
with TSCC samples without lymph node metastasis (Fig. 1C). β2-AR expression was increased
SCC tissue with poor differentiation compared with well
differentiated TSCC tissue (Fig.
1D). Among 75 cases examined in the study, 36 (48%) cases
exhibited high β2-AR staining and 40 (52%) cases displayed low
β2-AR expression.
Association of β2-AR with
clinicopathological features in TSCC
Associations were tested between β2-AR expression
and clinical and pathological features in this TSCC cohort
(Table I). χ2 analysis
demonstrated that β2-AR expression was associated with tumor
differentiation (P=0.004) and N stage (P=0.013). There were no
associations between β2-AR expression and age, gender, T stage and
clinical stage.
 | Table IAssociation between β2-AR expression
and clinicopathological features in patients with tongue squamous
cell carcinoma. |
Table I
Association between β2-AR expression
and clinicopathological features in patients with tongue squamous
cell carcinoma.
| Clinicopathologic
feature | No. of cases | β2-AR expression
| P-value |
|---|
| High (%) | Low (%) |
|---|
| Gender | | | | |
| Male | 38 | 19 | 19 | 0.725 |
| Female | 37 | 17 | 20 | |
| Age (years) | | | | |
| ≥55 | 40 | 21 | 19 | 0.404 |
| <55 | 35 | 15 | 20 | |
| Differentiation | | | | |
| Well | 40 | 13 | 27 | 0.004 |
| Moderate+poor | 35 | 23 | 12 | |
| T stage | | | | |
| T1–2 | 68 | 34 | 34 | 0.280 |
| T3–4 | 7 | 2 | 5 | |
| N stage | | | | |
| N− | 52 | 20 | 32 | 0.013 |
| N+ | 23 | 16 | 7 | |
| Clinical stage | | | | |
| I–II | 48 | 20 | 28 | 0.143 |
| III–IV | 27 | 16 | 11 | |
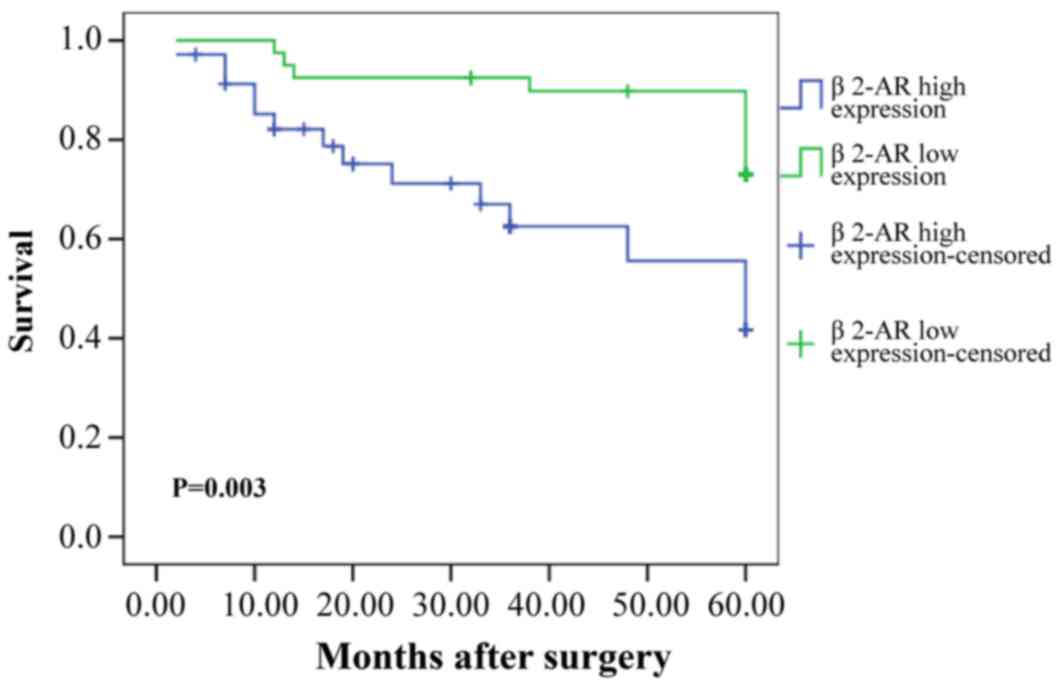
Prognostic value of β2-AR for patients
with TSCC
Kaplan-Meier analysis demonstrated a statistically
significant difference in 5-year survival rate between patients
with high β2-AR expression and low β2-AR expression (P=0.003;
Fig. 2). As demonstrated in
Table II, univariate analysis
indicated that tumor size, grade, pathological T stage, lymph node
metastasis, clinical stage, and β2-AR were all significant
prognostic factors for survival of patients with TSCC. Multivariate
analysis showed that lymph node metastasis, clinical stage, and
β2-AR were all independent prognostic factors for patient
survival.
| Table IICox proportional analysis of
variables affecting survival in patients with tongue squamous cell
carcinoma. |
Table II
Cox proportional analysis of
variables affecting survival in patients with tongue squamous cell
carcinoma.
| – Variable | Comparison | Univariate analysis
| Multivariate
analysis
|
|---|
| HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| Gender | Male vs.
female | 1.059
(0.475–2.359) | 0.889 | | |
| Age (years) | ≥55 vs. <55 | 0.996
(0.442–2.246) | 0.992 | | |
|
Differentiation | Well vs. moderate
or poor | 0.712
(0.319–1.588) | 0.406 | | |
| pT stage | T1–2 vs.
T3–4 | 2.392
(0.893–6.409) | 0.083 | | |
| pN stage | N0 vs.
N+ | 3.070
(1.372–6.871) | 0.006 | 0.878
(0.168–4.596) | 0.877 |
| pTNM stage | I–II vs.
III–IV | 3.039
(1.346–6.860) | 0.007 | 2.823
(0.582–13.696) | 0.198 |
| β2-AR
expression | High vs. low | 3.220
(1.396–7.428) | 0.006 | 2.780
(1.090–7.086) | 0.032 |
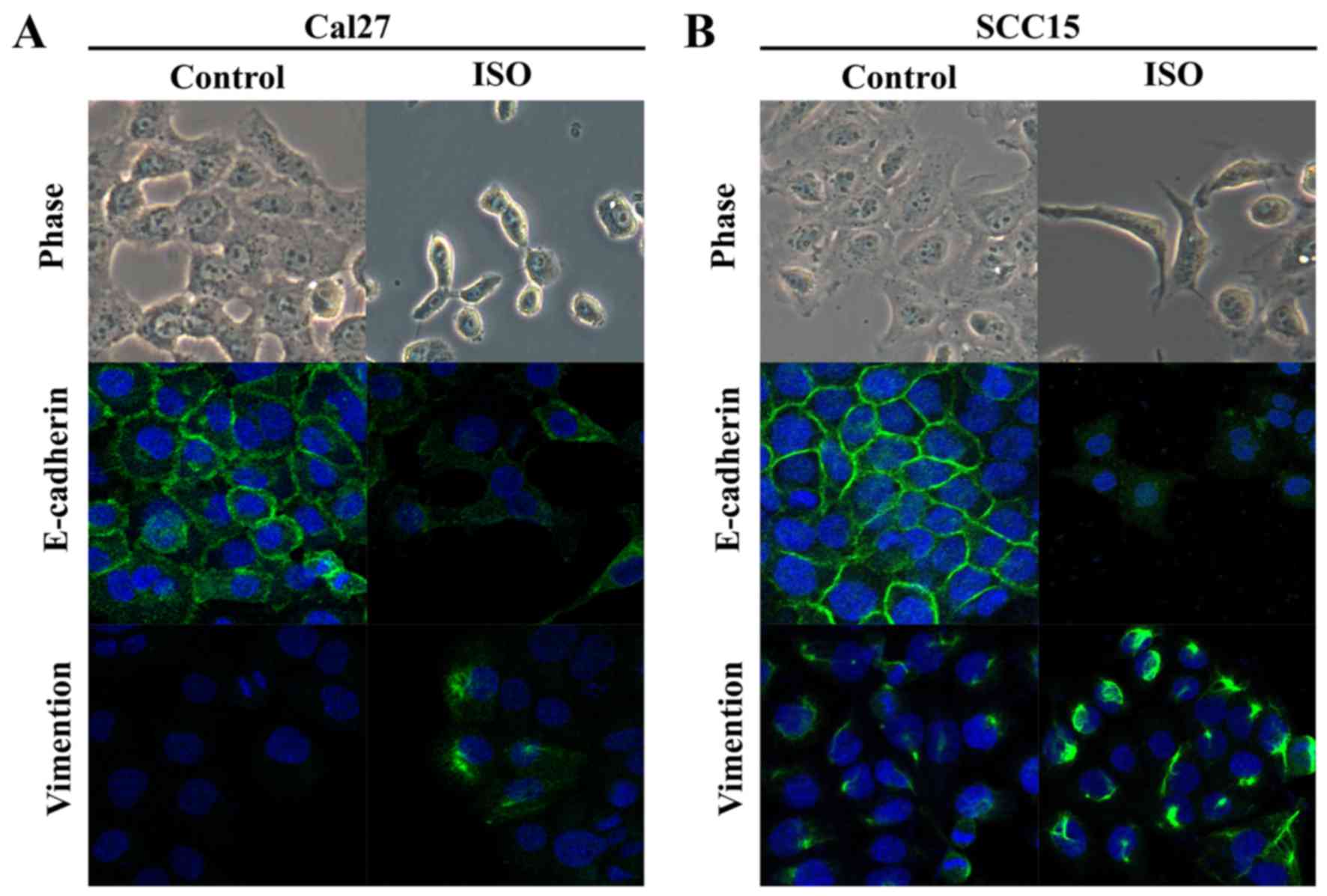
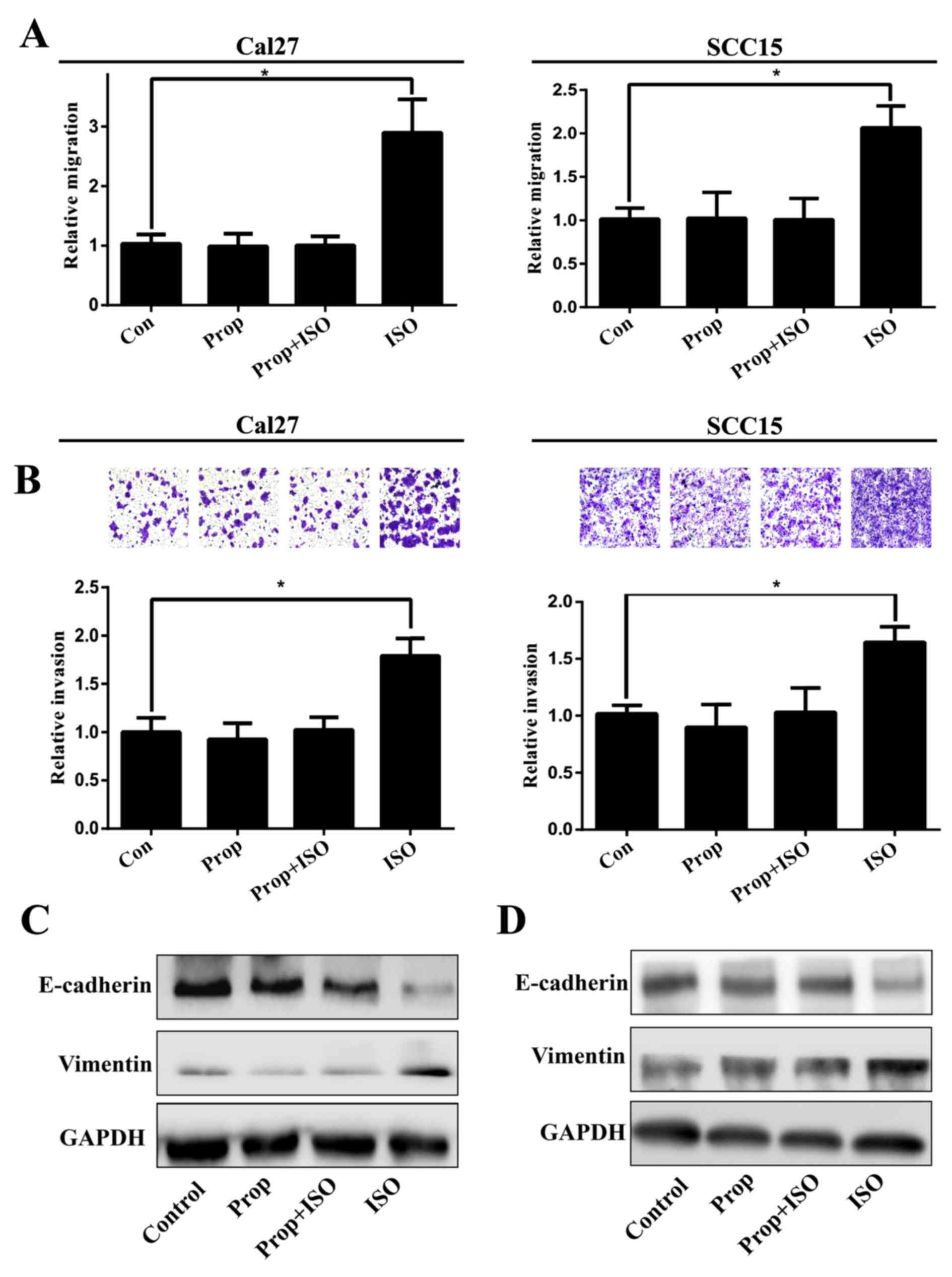
Effect of activation of β2-AR on EMT in
TSCC cells
TSCC cell lines Cal27 and SCC15 were treated with
ISO (β-AR agonist) to activate β2-AR signaling. The cell morphology
was dramatically changed from an epithelial phenotype to
mesenchymal phenotype with decreased E-cadherin expression and
increased vimentin expression (Fig.
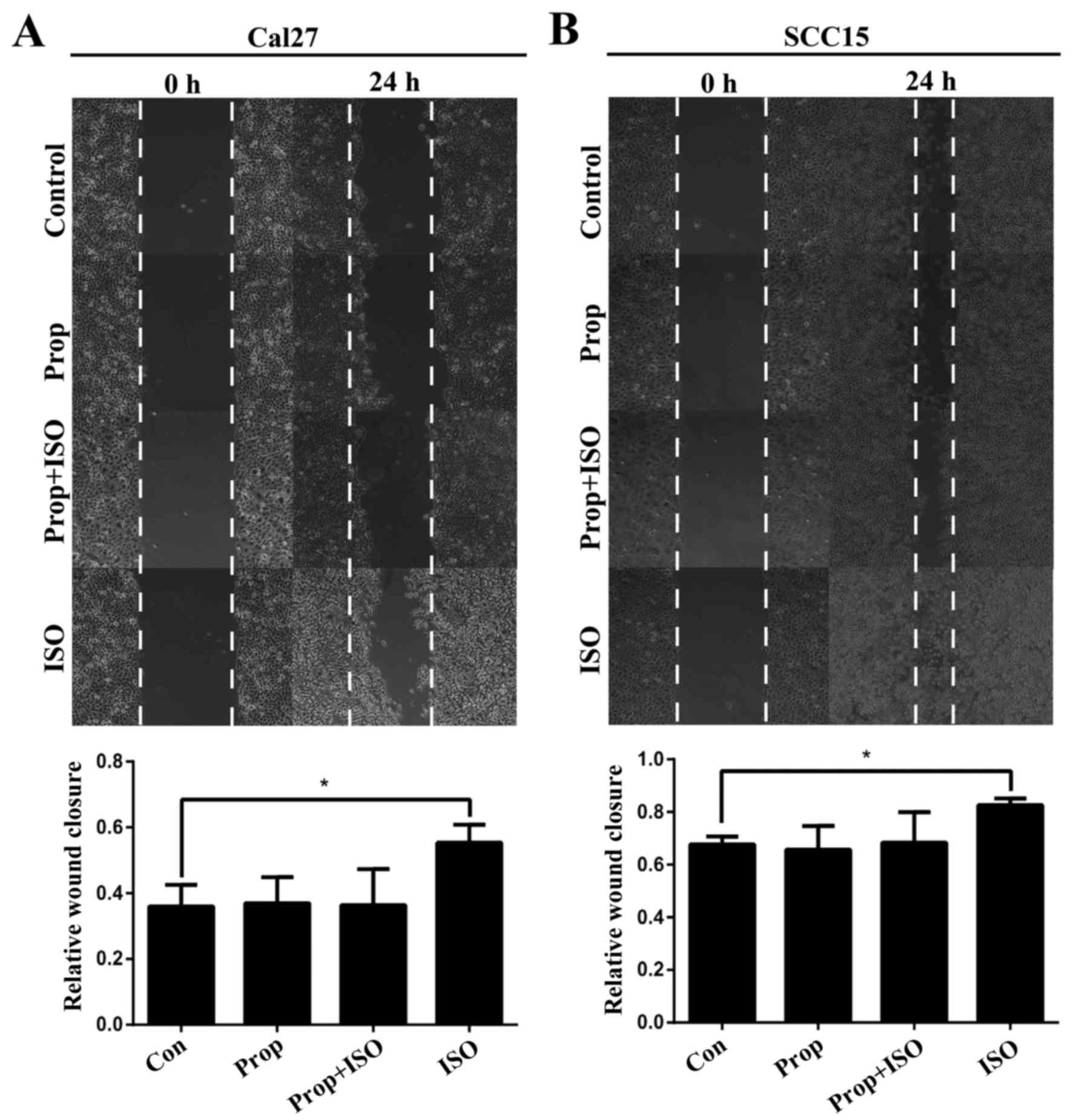
3A and B). A wound healing assay indicated that these changes
were accompanied by enhanced cell motility (Fig. 4A and B). Similar results were also
observed in Transwell assay coated with or without Matrigel
(Fig. 5A and B). Western blot
analyses further confirmed that E-cadherin expression was
downregulated and vimentin expression was upregulated by treatment
with ISO (Fig. 5C and D). By
contrast, western blot analysis indicated that ISO-mediated
activation of β2-AR (p-β2-AR), increased expression of vimentin and
Snail1, and reduced E-cadherin expression were inhibited by
co-treatment with ICI-118,551, but not metoprolol (Fig. 6A and B). The expression of IL-6,
Stat3, p-Stat3 and Snail1 were also examined in TSCC cells treated
by ISO with or without ICI-118,551. Activation of β2-AR promoted
the expression of IL-6, p-Stat3 and Snail1, while blockade of β2-AR
signaling with ICI-118,551 inhibited β2-AR-mediated activation of
IL-6/Stat3/Snail pathway (Fig.
7A). When cells were treated with anti-IL-6 monoclonal antibody
(mAb), ISO-induced EMT was partially reversed by blocking
IL-6/Stat3/Snail pathway (Fig.
7B) (24).
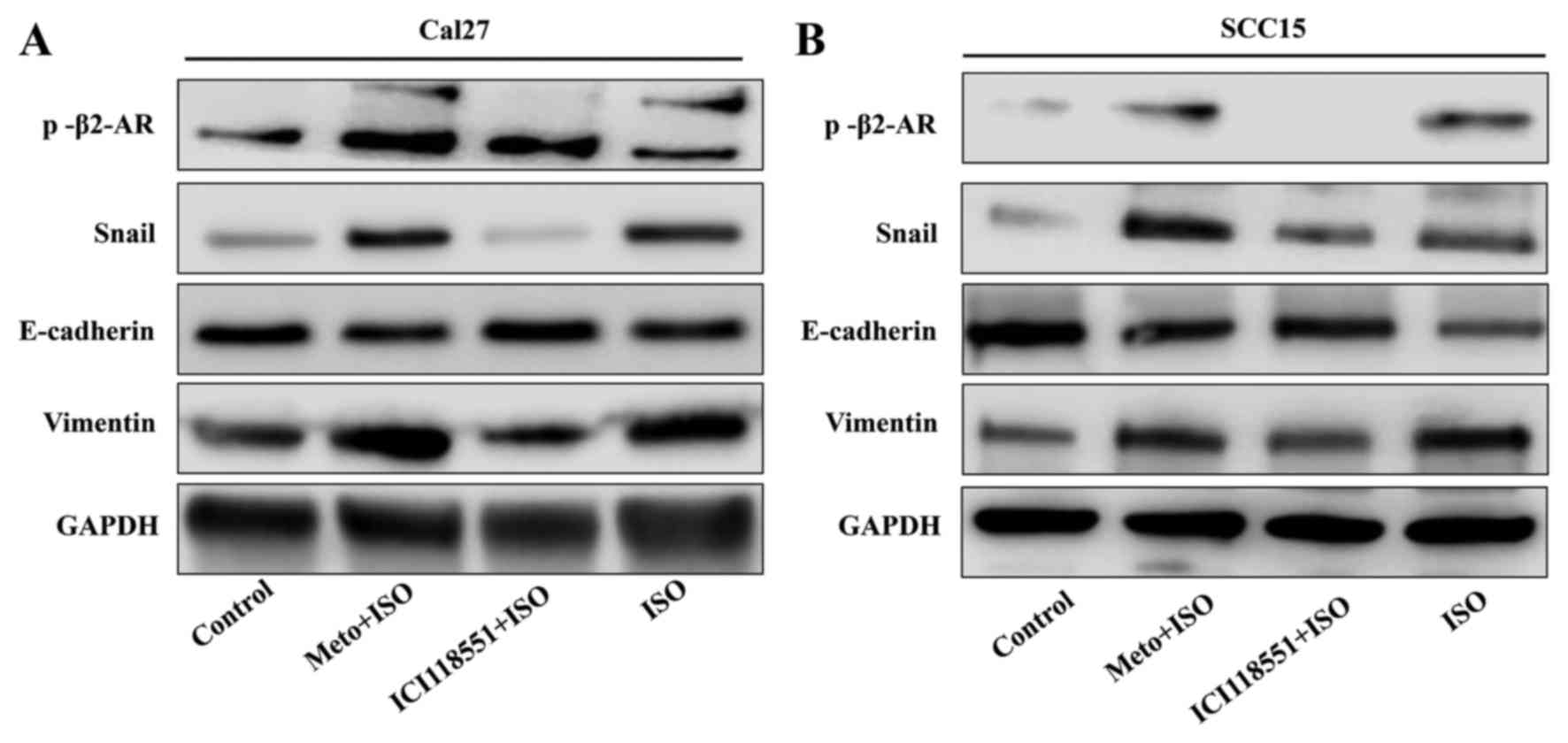 | Figure 6Western blot analysis indicates that
ISO induced epithelial mesenchymal transition through mediating
activation of β2-AR. (A) In Cal27 and (B) SCC15 cell lines,
p-β2-AR, vimentin and Snail1 were increased in ISO-treated groups,
while E-cadherin expression were decreased. Following treatment
with ICI-118,551 but not Meto, ISO induced the expression of
vimentin and Snail1, and E-cadherin was attenuated. p-, phospho;
β2-AR, β2-adrenergic receptor Meto, metoprolol; ISO,
isoproterenol. |
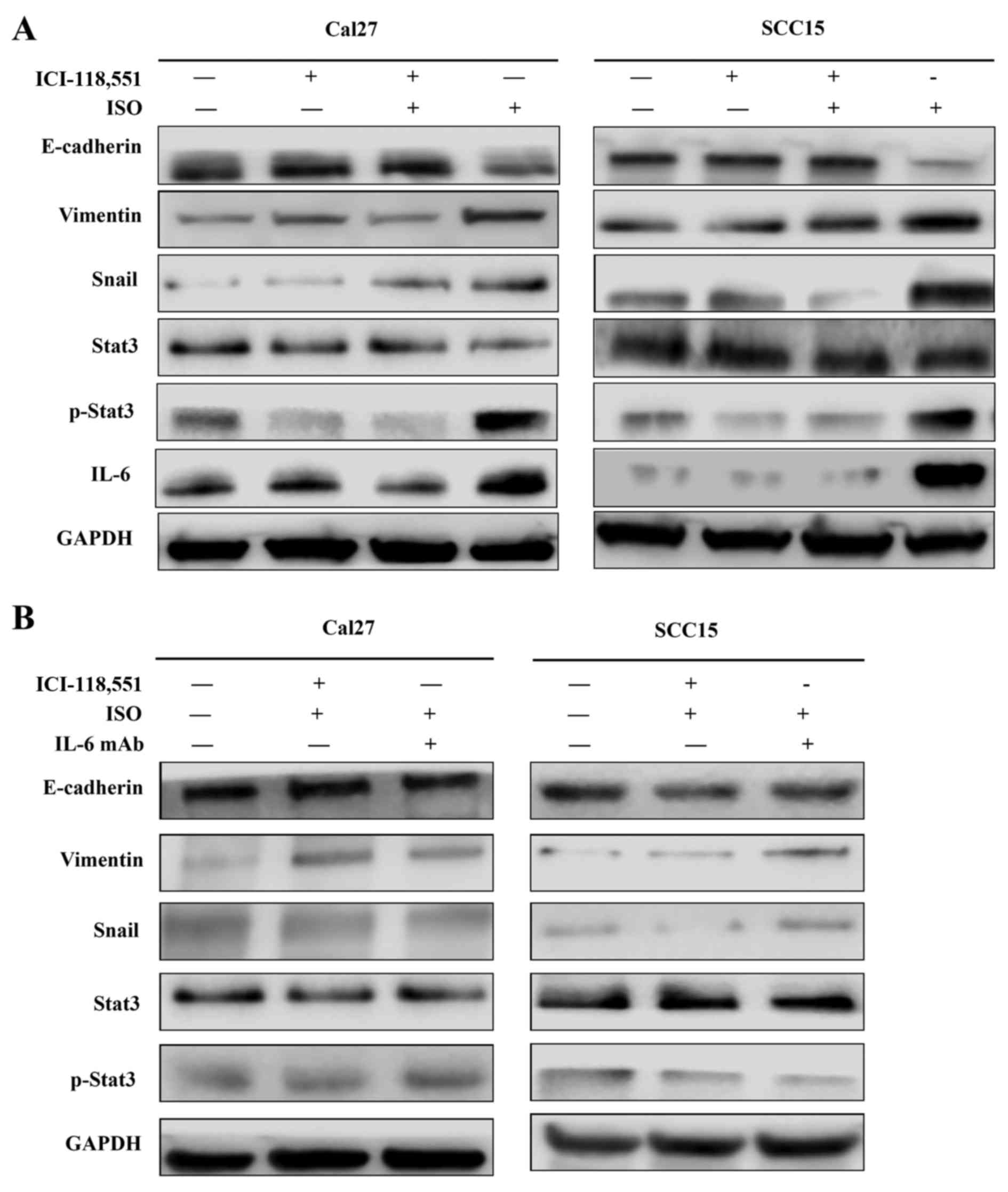 | Figure 7Activation of β2-AR promoted
epithelial mesenchymal transition in Cal27 and SCC15 cells through
initiating IL-6/Stat3/Snail1 pathway. (A) Expression of IL-6,
Stat3, p-Stat3 and Snail1 in Cal27 and SCC15 cells treated by ISO
with or without ICI-118,551. (B) Expression of IL-6, Stat3, p-Stat3
and Snail1 in the presence of IL-6 mAb, ISO and ICI-118,551. ISO,
isoproterenol; β2-AR, β2-adrenergic receptor; IL-6, interleukin-6;
mAb, monoclonal antibody; Stat3, signal transducer and activator of
transcription 3; p-, phospho. |
Discussion
Accumulating data demonstrate that EMT has an
important role in malignant cancer progression (17,18). Cancer cells can acquire the
ability to migrate and invade during EMT, which is orchestrated by
a complex molecular network. Recently, stress has been reported to
be correlated with cancer growth, drug resistance, recurrence,
progression and poor prognosis of patients (12). Stress activates the sympathetic
nervous system and results in increased production of
catecholamines, such as norepinephrine and epinephrine (12). The activation of β2-AR signaling
was indicated to be responsible for stress-mediated cancer
progression (19,20). The oncogenic role of β2-AR has
been suggested in several cancers type, including oral cancer
(21,22). However, the deregulation of β2-AR
in TSCC and its role in invasion and metastasis of TSCC has not
been fully investigated.
In the present study, the expression of β2-AR was
examined in TSCC tissue samples, and it was confirmed that β2-AR
expression was enhanced in TSCC tissue compared with non-cancerous
adjacent epithelium. This observation indicates that deregulation
of β2-AR signaling may contribute to TSCC tumorigenesis. Similar
results were previously observed in other studies (22). Further analysis demonstrated that
increased expression of β2-AR was positively associated with lymph
node metastasis, clinical stage and reduced overall survival.
Furthermore, overexpression of β2-AR was an independent poor
prognostic factor in patients with TSCC. These findings indicated
that activation of β2-AR signaling has a critical role in the
invasion and metastasis cascade in TSCC. Notably, Bravo-Calderón
et al (21) reported that
strong expression of β2-AR was a favorable prognostic factor for
patients with OSCC.
To confirm the observations above and explore the
functional relevance of β2-AR in invasion and metastasis of TSCC,
TSCC cells were treated with ISO (β-AR agonist) and/or propranolol
(non-selective β-AR antagonist) to activate or block β-AR signaling
pathway; a switch of TSCC cells from epithelial to mesenchymal
phenotype was observed in cancer cells, which suggested that EMT
was induced when cancer cells were treated with ISO. The switch was
also accompanied by decreased expression of E-cadherin, increased
expression of vimentin and enhanced cell migration and invasion.
These changes were also abolished by treatment with propranolol. To
further confirm that activation of β2-AR was responsible for the
ISO-induced EMT, metoprolol (selective β1-AR antagonist) and
ICI-118,551 (selective β2-AR antagonist) were used to inhibit β1-AR
and β2-AR signaling, respectively. Inhibition of β2-AR reversed
ISO-induced EMT. These findings confirmed that activation of β2-AR
induced EMT to promote migration and invasion in TSCC cells.
Similar results were also reported by Lu et al (15) and Shan et al (16). However, the molecular mechanisms
of β2-AR-mediated EMT in TSCC are not clear. Bernabé et al
(23) reported that β-AR
agonists, such as norepinephine, and ISO stimulation both increase
IL-6 secretion in human oral SCC cells lines. As previously
reported (24–26), the IL-6/Stat3 pathway has an
important role in EMT. To further clarify the potential mechanism
of β2-AR induced EMT, IL-6/Stat3/Snail1 pathways were investigated
in our study. In the current study, increased production of IL-6
was observed in Cal27 and SCC15 cells following ISO stimulation,
which was also associated with activation of Stat3 and increased
Snail1 expression to initiate EMT. Notably, these molecular changes
can be blocked by treatment with IL-6 mAb. These results suggested
that activation of IL-6/Stat3/Snail pathway is involved in β2-AR
mediated EMT in oral SCC.
In conclusion, the results of the present study
demonstrated that deregulation of β2-AR signaling may be involved
in TSCC progression. One of its major roles is promoting EMT to
enhance cancer cell migration and invasion in TSCC progression.
Furthermore, β2-AR-mediated EMT was regulated in TSCC, at least in
part, through activation of an IL-6/Stat3/Snail pathway. Further
studies are necessary to investigate the cross-talk between β2-AR
and IL-6 signaling.
Acknowledgments
This study was support by National Natural Science
Grant of China (nos. 81272949, 81172567 and 81372885), Guangdong
Province Nature Science Foundation (no. 20130319c).
References
|
1
|
Shaul YD, Freinkman E, Comb WC, Cantor JR,
Tam WL, Thiru P, Kim D, Kanarek N, Pacold ME, Chen WW, et al:
Dihydropyrimidine accumulation is required for the
epithelial-mesenchymal transition. Cell. 158:1094–1109. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Thiery JP, Acloque H, Huang RY and Nieto
MA: Epithelial-mesenchymal transitions in development and disease.
Cell. 139:871–890. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Jiang Y, Liao L, Shrestha C, Ji S, Chen Y,
Peng J, Wang L, Liao E and Xie Z: Reduced expression of E-cadherin
and p120-catenin and elevated expression of PLC-γ1 and PIKE are
associated with aggressiveness of oral squamous cell carcinoma. Int
J Clin Exp Pathol. 8:9042–9051. 2015.
|
|
4
|
Rajwar YC, Jain N, Bhatia G, Sikka N, Garg
B and Walia E: Expression and significance of cadherins and its
subtypes in development and progression of oral cancers: a review.
J Clin Diagn Res. 9:ZE05–ZE07. 2015.PubMed/NCBI
|
|
5
|
Soares MQ, Mendonça JA, Morais MO, Leles
CR, Batista AC and Mendonça EF: E-cadherin, β-catenin, and α2β1 and
α3β1 integrin expression in primary oral squamous cell carcinoma
and its regional metastasis. Histol Histopathol. 30:1213–1222.
2015.PubMed/NCBI
|
|
6
|
Wang C, Huang H, Huang Z, Wang A, Chen X,
Huang L, Zhou X and Liu X: Tumor budding correlates with poor
prognosis and epithelial-mesenchymal transition in tongue squamous
cell carcinoma. J Oral Pathol Med. 40:545–551. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Wang C, Liu X, Huang H, Ma H, Cai W, Hou
J, Huang L, Dai Y, Yu T and Zhou X: Deregulation of Snai2 is
associated with metastasis and poor prognosis in tongue squamous
cell carcinoma. Int J Cancer. 130:2249–2258. 2012. View Article : Google Scholar
|
|
8
|
Wang C, Liu X, Chen Z, Huang H, Jin Y,
Kolokythas A, Wang A, Dai Y, Wong DT and Zhou X: Polycomb group
protein EZH2-mediated E-cadherin repression promotes metastasis of
oral tongue squamous cell carcinoma. Mol Carcinog. 52:229–236.
2013. View
Article : Google Scholar
|
|
9
|
Liu X, Wang C, Chen Z, Jin Y, Wang Y,
Kolokythas A, Dai Y and Zhou X: MicroRNA-138 suppresses
epithelial-mesenchymal transition in squamous cell carcinoma cell
lines. Biochem J. 440:23–31. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu D, Deng Q, Sun L, Wang T, Yang Z, Chen
H, Guo L, Liu Y, Ma Y, Guo N, et al: A Her2-let-7-β2-AR circuit
affects prognosis in patients with Her2-positive breast cancer. BMC
Cancer. 15:8322015. View Article : Google Scholar
|
|
11
|
Yang EV, Kim SJ, Donovan EL, Chen M, Gross
AC, Webster Marketon JI, Barsky SH and Glaser R: Norepinephrine
upregulates VEGF, IL-8, and IL-6 expression in human melanoma tumor
cell lines: implications for stress-related enhancement of tumor
progression. Brain Behav Immun. 23:267–275. 2009. View Article : Google Scholar :
|
|
12
|
Barbieri A, Bimonte S, Palma G, Luciano A,
Rea D, Giudice A, Scognamiglio G, La Mantia E, Franco R, Perdonà S,
et al: The stress hormone norepinephrine increases migration of
prostate cancer cells in vitro an in vivo. Int J Oncol. 47:527–534.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Shi M, Yang Z, Hu M, Liu D, Hu Y, Qian L,
Zhang W, Chen H, Guo L, Yu M, et al: Catecholamine-induced
β2-adrenergic receptor activation mediates desensitization of
gastric cancer cells to trastuzumab by upregulating MUC4
expression. J Immunol. 190:5600–5608. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Zhang J, Deng YT, Liu J, Wang YQ, Yi TW,
Huang BY, He SS, Zheng B and Jiang Y: Norepinephrine induced
epithelial-mesenchymal transition in HT-29 and A549 cells in vitro.
J Cancer Res Clin Oncol. 142:423–435. 2016. View Article : Google Scholar
|
|
15
|
Lu YJ, Geng ZJ, Sun XY, Li YH, Fu XB, Zhao
XY and Wei B: Isoprenaline induces epithelial-mesenchymal
transition in gastric cancer cells. Mol Cell Biochem. 408:1–13.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Shan T, Cui X, Li W, Lin W, Li Y, Chen X
and Wu T: Novel regulatory program for norepinephrine-induced
epithelial-mesenchymal transition in gastric adenocarcinoma cell
lines. Cancer Sci. 105:847–856. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Ye X and Weinberg RA:
Epithelial-mesenchymal plasticity: a central regulator of cancer
progression. Trends Cell Biol. 25:675–686. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Ye X, Tam WL, Shibue T, Kaygusuz Y,
Reinhardt F, Ng Eaton E and Weinberg RA: Distinct EMT programs
control normal mammary stem cells and tumour-initiating cells.
Nature. 525:256–260. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Qin JF, Jin FJ, Li N, Guan HT, Lan L, Ni H
and Wang Y: Adrenergic receptor β2 activation by stress promotes
breast cancer progression through macrophages M2 polarization in
tumor microenvironment. BMB Rep. 48:295–300. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Braadland PR, Ramberg H, Grytli HH and
Taskén KA: β-Adrenergic receptor signaling in prostate cancer.
Front Oncol. 4:3752015. View Article : Google Scholar
|
|
21
|
Bravo-Calderón DM, Oliveira DT, Marana AN,
Nonogaki S, Carvalho AL and Kowalski LP: Prognostic significance of
beta-2 adrenergic receptor in oral squamous cell carcinoma. Cancer
Biomark. 10:51–59. 2011–2012. View Article : Google Scholar
|
|
22
|
Shang ZJ, Liu K and Liang DF: Expression
of beta2-adrenergic receptor in oral squamous cell carcinoma. J
Oral Pathol Med. 38:371–376. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Bernabé DG, Tamae AC, Biasoli ÉR and
Oliveira SH: Stress hormones increase cell proliferation and
regulates interleukin-6 secretion in human oral squamous cell
carcinoma cells. Brain Behav Immun. 25:574–583. 2011. View Article : Google Scholar
|
|
24
|
Yadav A, Kumar B, Datta J, Teknos TN and
Kumar P: IL-6 promotes head and neck tumor metastasis by inducing
epithelial-mesenchymal transition via the JAK-STAT3-SNAIL signaling
pathway. Mol Cancer Res. 9:1658–1667. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Kim MS, Lee WS, Jeong J, Kim SJ and Jin W:
Induction of metastatic potential by TrkB via activation of
IL6/JAK2/STAT3 and I3K/AKT signaling in breast cancer. Oncotarget.
6:40158–40171. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liu H, Ren G, Wang T, Chen Y, Gong C, Bai
Y, Wang B, Qi H, Shen J, Zhu L, et al: Aberrantly expressed Fra-1
by IL-6/STAT3 transactivation promotes colorectal cancer
aggressiveness through epithelial-mesenchymal transition.
Carcinogenesis. 36:459–468. 2015. View Article : Google Scholar : PubMed/NCBI
|