Introduction
Ovarian cancer is the most common form of
gynecological cancer and is a serious threat to women's health and
survival worldwide. Cis-diammine-dichloroplatinum (II), also known
as cisplatin, is a classic chemotherapy drug that is widely used
for the treatment of advanced cancers, including ovarian cancer
(1,2). Cisplatin exhibits a satisfactory
therapeutic effect during the early stages of administration;
however, patients often become resistant to cisplatin during
long-term administration. It has previously been reported that
overexpression of the anti-apoptotic B-cell lymphoma 2 (Bcl-2)
protein facilitates the pathophysiology and cisplatin resistance of
cancer by inhibiting apoptosis (3). Bcl-2 contains BH1, BH2, BH3 and BH4
domains. Bcl-2 has been reported to bind to Bcl-2-associated X
protein (Bax)/Bcl-2 homologous antagonist killer via its BH3 domain
and inhibit cytochrome c release from the mitochondria, thus
maintaining mitochondrial integrity. Notably, BH3-mimetic Bcl-2
inhibitors have attracted much attention and have been extensively
studied (4). The BH3-mimetics
ABT737 and ABT-263 have been reported to enhance cisplatin
sensitivity in cholangiocarcinoma and ovarian carcinoma cells
(5,6). However, the function of the Bcl-2
BH4 domain in ovarian carcinoma cells remains unclear.
Bcl-2 has been reported to induce a reduction in
endoplasmic reticulum (ER) Ca2+ load and release via its
BH4 domain (7). The ER is the
main store of intracellular Ca2+ and a crucial organelle
in eukaryotic cells. A large release of Ca2+ from the ER
can activate calpain-1, which is a Ca2+-dependent
cysteine protease responsible for the transition from caspase-4 to
cleaved caspase-4, which in turn activates ER stress-induced
apoptosis (8). Furthermore,
Ca2+ release from the ER can enhance mitochondrial
Ca2+ uptake, which is accompanied by a release of
cytochrome c and activation of the mitochondrial apoptotic
pathway (9). In our previous
study, it was demonstrated that the expression of Bcl-2 was higher
in cisplatin-resistant SKOV3/DDP ovarian cancer cells compared with
in cisplatin-sensitive SKOV3 ovarian cancer cells (10). A low dose of cisplatin induced
apoptosis, which was accompanied by cytoplasmic and mitochondrial
Ca2+ overload in SKOV3 cells, but not in SKOV3/DDP cells
(11). These results suggested
that Bcl-2 contributes to cisplatin resistance in ovarian cancer by
inhibiting ER Ca2+ release.
Bcl-2 inhibits ER Ca2+ release by
directly binding and inhibiting the inositol 1,4,5-trisphosphate
receptor (IP3R), which is the main ER Ca2+ release
channel, via its BH4 domain. IP3R transmits signals between the
cytoplasm and ER microenvironment, and can modulate the
mitochondrial Ca2+ signal (12). By deleting the BH4 domain, Liu
et al demonstrated that the BH4 domain was important for the
prosurvival activity of Bcl-2 (13). Stabilized TAT-fused IP3R-derived
peptide (TAT-IDPS) can specifically target the BH4
domain of Bcl-2 and reverse Bcl-2-mediated inhibition of
IP3-induced Ca2+ elevation, ultimately increasing
Ca2+-dependent cell apoptosis (14). TAT-IDPS has been
reported to induce apoptosis of diffuse large B-cell lymphoma cells
by promoting IP3-induced proapoptotic Ca2+ signaling
(14). These results indicated
that targeting the BH4 domain may induce conversion of Bcl-2 from a
prosurvival protein into a proapoptotic protein. Notably, Zhong
et al indicated that TAT-IDPS sensitivity is
positively correlated with Bcl-2 expression (15). Therefore, it may be hypothesized
that TAT-IDPS increases the sensitivity of cells to
cisplatin via IP3-dependent Ca2+ release in SKOV3/DDP
cells.
The present study aimed to evaluate the effects of
TAT-IDPS on the cytotoxicity of cisplatin in SKOV3/DDP
cells. The results demonstrated that TAT-IDPS increased
cisplatin-induced apoptosis by increasing Ca2+ release
from the ER to the cytoplasm and mitochondria. These experimental
results indicated that TAT-IDPS, combined with
chemotherapy, may be considered a viable treatment strategy for the
prevention and treatment of ovarian cancer.
Materials and methods
Antibodies and drugs
Anti-caspase-3 (sc-7272), anti-caspase-4 (sc-56056)
and anti-cleaved caspase-4 (sc-22173-R) antibodies (Abs) were
purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA).
Anti-cleaved caspase-3 (ab2302), anti-protein disulfide isomerase
(PDI; ab2792) and anti-CCAAT-enhancer-binding protein homologous
protein (CHOP; ab11419) Abs were purchased from Abcam (Hong Kong)
Ltd. (Hong Kong, China). Anti-β-actin (60008–1-Ig),
anti-Bcl-2-associated X protein (Bax; 50599-2-Ig), anti-Bcl-2
(12789-1-AP), anti-cytochrome c (10993-1-AP),
peroxidase-conjugated AffiniPure goat anti-mouse immunoglobulin
(Ig)G (H+L) (SA00001-1) and peroxidase-conjugated AffiniPure goat
anti-rabbit IgG (H+L) (SA00001-2) Abs were purchased from
Proteintech Group, Inc. (Chicago, IL, USA). The anti-calpain-1
catalytic subunit (#31038-1) Ab was purchased from Signalway
Antibody LLC (College Park, MD, USA). Cisplatin was purchased from
Sigma-Aldrich; Merck KgaA (Darmstadt, Germany) and was dissolved in
normal saline for in vitro use. TAT-IDPS was
synthesized and purified by ChinaPeptides Co., Ltd. (Shanghai,
China) and was dissolved in ddH2O. The sequence for
TAT-IDPS is RKKRRQRRRGGNVYTEIKCNSLLPLDDIVRV.
2-Aminoethyl diphenylborinate (2-APB; ab120124) was purchased from
Abcam (Hong Kong) Ltd.
Cell lines
Cisplatin-resistant ovarian cancer SKOV3/DDP cells
were obtained from the Chinese Academy of Medical Sciences
(Beijing, China) and Peking Union Medical College (Beijing, China).
Cells were cultured at 37°C in an atmosphere containing 5%
CO2 in RPMI-1640 culture medium (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) supplemented with 10% fetal
bovine serum (FBS; Gibco; Thermo Fisher Scientific, Inc.).
Cisplatin-resistant SKOV3/DDP cells were maintained in medium
containing 1 μg/ml cisplatin (Sigma-Aldrich; Merck KGaA) for
24 h to maintain resistance.
Cell viability assays
Cell viability was determined using the
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT)
assay. SKOV3/DDP cells were seeded into 96-well culture plates in
100 μl medium at a density of 1×104 cells/well.
After 2 days, agents [cisplatin (15 μg/ml) and/ or TAT-IDPS
(25 μM) in the presence or absence of 50 μM 2-APB]
were added to quadruplicate wells and incubated for 24 or 48 h at
37°C. For the MTT assay (Beyotime Institute of Biotechnology,
Shanghai, China), 20 μl/well MTT (5 mg/ml in PBS) was added
to each well; after 4 h at 37°C, dimethyl sulfoxide (150
μl/well; Beijing Chemical Industry Group Co., Beijing,
China) was added and the plates were agitated at room temperature
for 10 min. Absorbance was measured at a wavelength of 570 nm using
a microplate reader (BioTek Instruments, Inc., Winooski, VT,
USA).
Western blotting
Whole-cell protein extracts were isolated from
SKOV3/DDP cells using cell lysis buffer [50 mM Tris-HCl, pH 7.5;
150 mM NaCl; 1 mM sodium-EDTA, 1 mM EDTA; 1% (v/v) Triton X-100;
2.5 mM sodium pyrophos-phate; 1 mM β-glycerophosphate; 1 mM
Na3VO4; 1 mM NaF; 1 μg/ml leupeptin
and 1 mM phenylmethylsulfonyl fluoride] for western blotting.
Protein concentration was quantified using the Quick Start Bradford
protein assay kit (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
For western blot analysis, lysates (30 μg) were separated by
10% (w/v) SDS-PAGE and were transferred onto immobilon-P transfer
membranes (EMD Millipore, Billerica, MA, USA). The membranes were
blocked with 5% (w/v) non-fat dry milk in buffer [10 mM Tris-HCl
(pH 7.6), 100 mM NaCl, and 0.1% Tween-20] for 1 h at room
temperature and were then incubated with the desired primary Abs
[caspase-3 (dilution, 1:200); caspase-4 (dilution, 1:200); cleaved
caspase-4 (dilution, 1:200); cleaved caspase-3 (dilution, 1:1,000);
PDI (dilution, 1:1,000); CHOP (dilution, 1:1,000); β-actin
(dilution, 1:2,000); Bax (dilution, 1:1,000); cytochrome c
(dilution, 1:1,000); calpain-1 (dilution, 1:200)] overnight at 4°C,
followed by incubation with horseradish peroxidase-conjugated
secondary Abs (dilution, 1:2,000) for 1.5 h at room temperature.
Immunodetection was performed using enhanced chemiluminescence
reagents (PE0010-b; Solarbio, New York, NY, USA) and images were
captured using Syngene Bio Imaging (Syngene, Cambridge, UK).
Protein expression levels were normalized to those of β-actin and
the ratios of normalized protein to β-actin are presented as the
mean ± standard deviation from three independent experiments.
Protein expression levels were semi-quantified by densitometry
using Quantity One software (version 4.6.9; Bio-Rad Laboratories,
Inc.).
Immunofluorescence staining and confocal
laser microscopy
Cells (4×104 cells/well) were cultured on
coverslips overnight, following treatment with increasing doses of
cisplatin (5, 10 and 15 μg/ml) for 24 h at 37°C, and were
fixed with 4% (w/v) paraformaldehyde and stained with nuclear
Hoechst 33342 (1 μg/ml; Sigma-Aldrich; Merck KGaA) for 5
min. Subsequently, the cells were washed with PBS and were examined
under an Olympus FV1000 confocal laser microscope (Olympus
Corporation, Tokyo, Japan) to reveal cell chromatin condensation.
The expression of calpain-1 was examined by indirect
immunofluorescence. Cells (4×104 cells/ well) were
cultured on coverslips overnight, and were then treated with the
indicated dose of cisplatin with or without TAT-IDPS for
24 h at 37°C, after which the cells were rinsed with PBS three
times. Subsequently, cells were fixed with 4% (w/v)
paraformaldehyde for 20 min, permeabilized with 0.1% (v/v) Triton
X-100 for 5 min, blocked with bovine serum albumin (PA129262;
Thermo Fisher Scientific, Inc.), and incubated with anti-calpain-1
(dilution, 1:100) overnight at 4°C. Cells were then incubated with
Alexa Fluor 488 donkey anti-rabbit IgG (#R37602, 1:400 dilution;
Invitrogen; Thermo Fisher Scientific, Inc.) for 30 min at room
temperature, stained with Hoechst 33342 (1 μg/ml) for 5 min
at room temperature, and washed with PBS three times. After
mounting, cells were examined under an Olympus FV1000 confocal
laser microscope (Olympus Corporation). The same parameters of
illumination and detection were maintained digitally throughout the
experiments.
Calcium concentration analysis
The Ca2+-sensitive fluorescent dyes
Fluo-4/AM (Molecular Probes; Thermo Fisher Scientific, Inc.) and
Rhod-2/AM (AAT Bioquest, Sunnyvale, CA, USA) were used to measure
Ca2+ concentration according to the manufacturers'
protocols. Prior to exposure to various experimental conditions,
the cells were incubated with Fluo-4/AM or Rhod-2/AM for 30 min at
37°C. Cell samples were then analyzed by confocal laser microscopy.
All experiments were performed in triplicate.
Flow cytometry
Cell death was detected using the Muse®
Annexin V Dead Cell kit (EMD Millipore). SKVO3/DDP cells were
seeded in 6-well culture plates at a density of 2×105
cells/well. Following treatment under different experimental
conditions, cells were trypsinized and resuspended in RPMI-1640
medium supplemented with 10% FBS at a concentration of
1x106 cells/ml. In the dark, cells were incubated with
Annexin V at room temperature for 20 min. Finally, cells were
measured using the Muse® Cell Analyzer (version 1.5.0.0;
EMD Millipore). All experiments were performed in triplicate.
Mitochondrial membrane potential
(ΔΨm)
Alterations in ΔΨm during the early stages of
apoptosis were assessed using the Muse® MitoPotential
assay (MCH 100110; EMD Millipore). SKOV3/DDP cells were treated
with cisplatin alone or in combination with TAT-IDPS.
Subsequently, cells were centrifuged at 3,000 × g for 5 min at room
temperature, and the cell pellet was suspended in assay buffer
(1×105 cells/100 μl). MitoPotential dye working
solution was added, and the cell suspension was incubated at 37°C
for 20 min. Following the addition of Muse®
MitoPotential 7-aminoactinomycin D dye (propidium iodide) for 5
min, alterations in ΔΨm and in cellular plasma membrane
permeabilization were assessed on the basis of the fluorescence
intensities of both dyes, which were analyzed by flow cytometry
(Muse® Cell Analyzer version 1.5.0.0; EMD
Millipore).
Statistical analysis
Data are representative of three independent
experiments performed in triplicate and are presented as the mean ±
standard deviation. Data analysis was performed using one-way
analysis of variance, and Tukey's post-hoc test was used to
determine statistical significance of all pairwise comparisons of
interest. P<0.05 was considered to indicate a statistically
significant difference. Statistical analyses were performed using
the Complex Samples module in IBM SPSS Statistics 20.0 (SPSS, Inc.,
Chicago, IL, USA)
Results
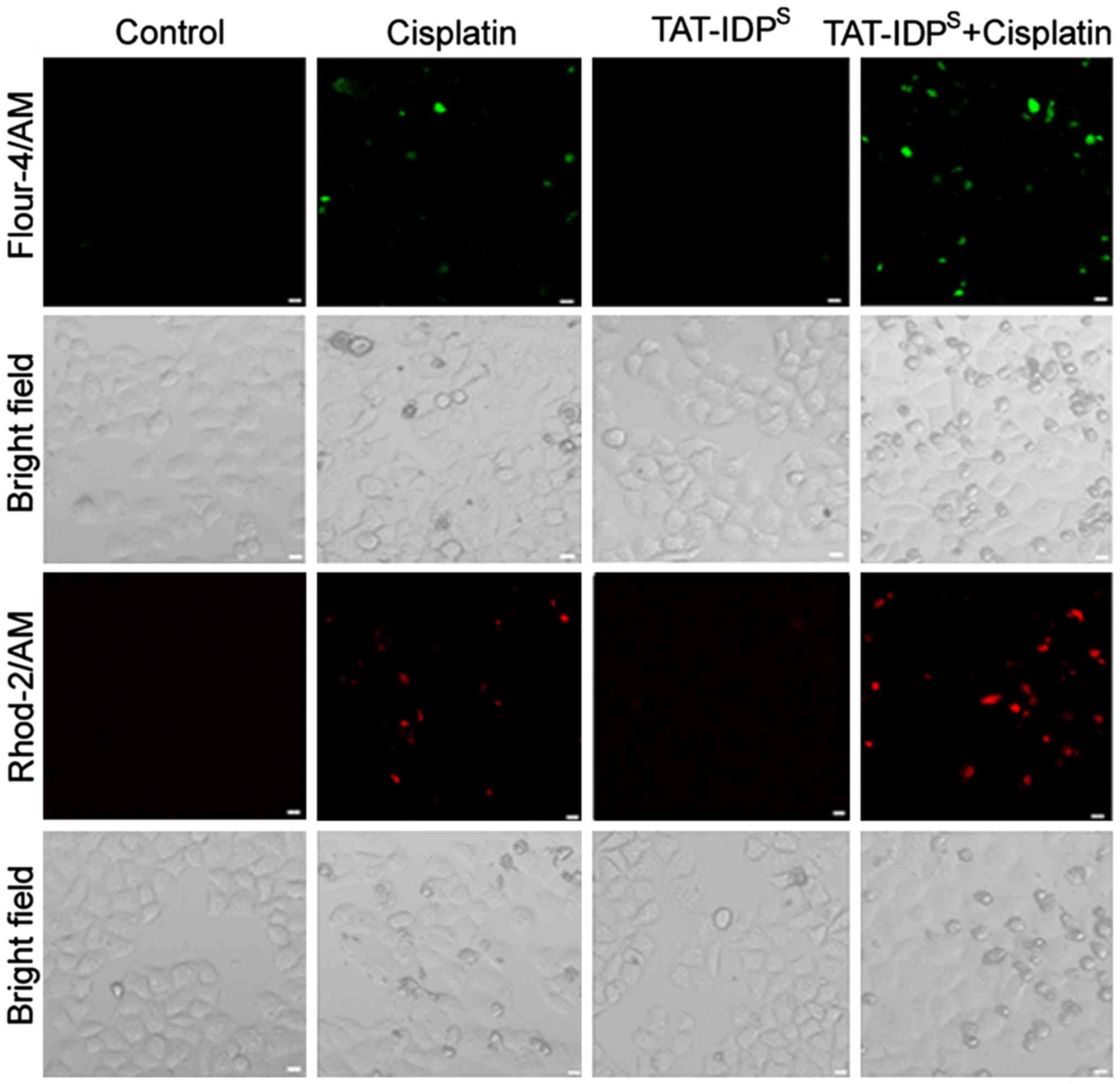
TAT-IDPS enhances
cisplatin-induced cytosolic Ca2+ elevation and
mitochondrial Ca2+ overload in SKOV3/DDP cells
TAT-IDPS can enhance IP3R-mediated
Ca2+ signals by disrupting the Bcl-2-IP3R interaction.
The present study explored whether TAT-IDPS increased
cisplatin-induced cytosolic and mitochondrial Ca2+
levels in SKOV3/DDP cells. SKOV3/DDP cells were treated with
cisplatin (15 μg/ml) and/or TAT-IDPS (25
μM) for 24 h and Ca2+ levels were then examined
in the mitochondria and cytoplasm. The Ca2+-sensitive
fluorescent dyes Fluo-4/AM and Rhod-2/AM were used to detect
cytosolic and mitochondrial Ca2+ levels by confocal
microscopy, respectively. Compared with in the control (untreated)
group, cisplatin induced an elevation in free Ca2+ in
the cytosol and mitochondria. Furthermore, cisplatin-induced
Ca2+ elevation in the cytosol and mitochondria was
further increased in the cells treated with cisplatin and
TAT-IDPS (Fig. 1).
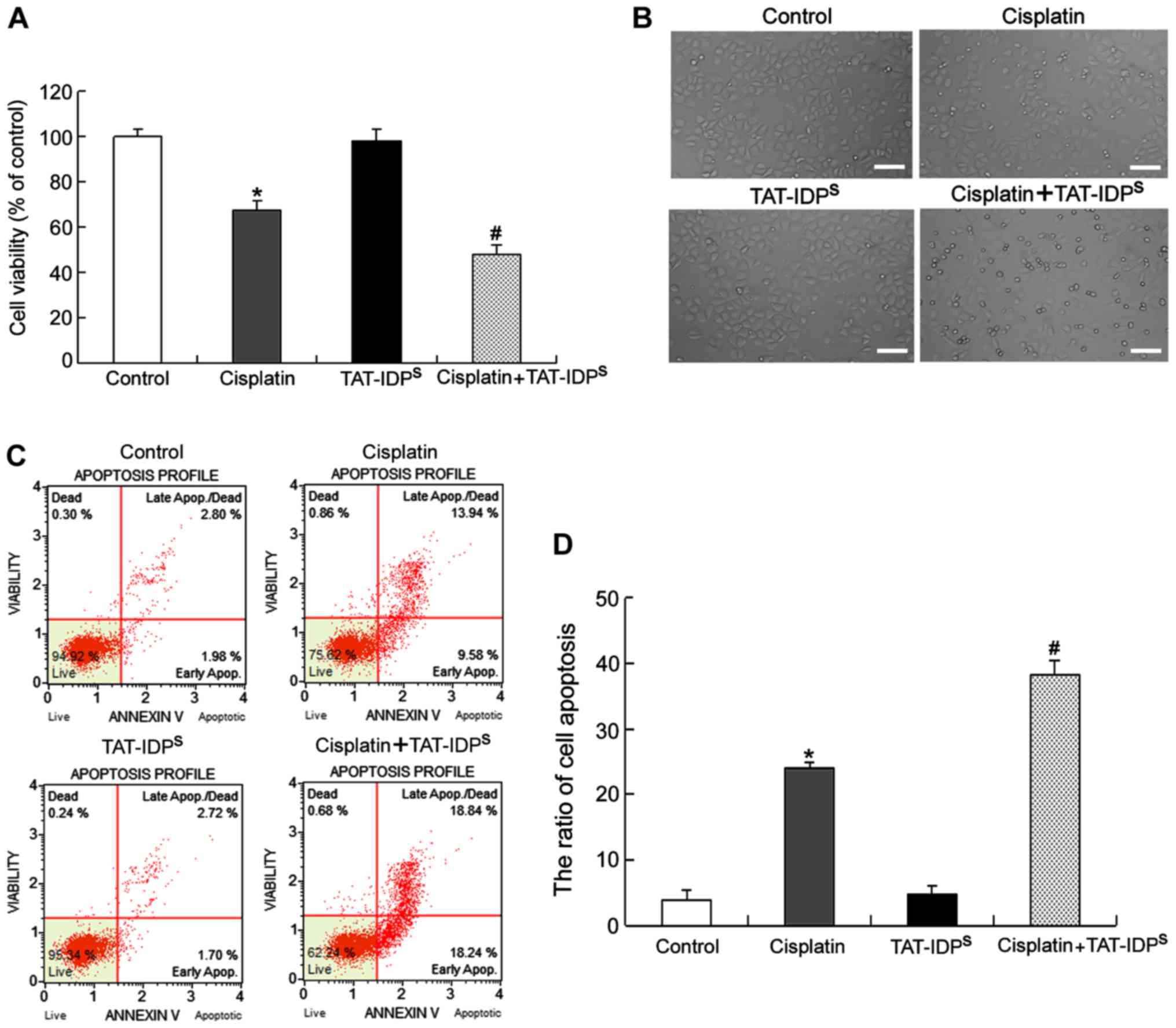
TAT-IDPS increases
cisplatin-induced apoptosis of SKOV3/DDP cells
To evaluate the effects of TAT-IDPS on
cisplatin-induced growth inhibition and apoptosis, SKOV3/DDP cells
were treated with cisplatin (15 μg/ml) and/or
TAT-IDPS (25 μM) for 24 h and growth inhibition
was examined using MTT assays. The results indicated that cisplatin
induced growth inhibition in SKOV3/DDP cells. In addition,
cotreatment with cisplatin and TAT-IDPS increased the
growth inhibitory effects of cisplatin, whereas TAT-IDPS
alone had no effect on cell viability compared with in the control
group (Fig. 2A). Under an optical
microscope, it was demonstrated that SKOV3/DDP cells treated with
cisplatin became round and fragmented compared with the control
cells and that TAT-IDPS enhanced this effect (Fig. 2B). Furthermore, flow cytometric
analysis demonstrated that the rate of apoptosis in SKOV3/DDP cells
treated with cisplatin and TAT-IDPS was higher than that
in cells treated with cisplatin alone (Fig. 2C and D). These results suggested
that TAT-IDPS increased cisplatin-induced growth
inhibition and apoptosis of SKOV3/DDP cells.
TAT-IDPS increases
cisplatin-induced ER stress-associated apoptosis by increasing
calpain-1 expression in SKOV3/DDP cells
Since calpain-1, which is a ubiquitous protease, can
be activated by cytoplasmic Ca2+ elevation (16), the present study detected the
expression of calpain-1 using confocal microscopy. The results
indicated that cotreatment with cisplatin and TAT-IDPS
for 24 h increased cisplatin-induced calpain-1 accumulation
(Fig. 3A). Calpain-1 has been
reported to enhance ER-associated apoptosis by cleaving its protein
substrates (16). Subsequently,
the present study investigated whether TAT-IDPS may
sensitize SKOV3/DDP cells to cisplatin-induced ER stress-associated
apoptosis. The expression levels of the following ER
stress-associated apoptotic proteins: Calpain-1, PDI, CHOP and
cleaved caspase-4, were detected by western blot analysis.
Cisplatin induced the expression of all four proteins; furthermore,
combined treatment with TAT-IDPS enhanced these effects
(Fig. 3B and C). These results
suggested that TAT-IDPS may increase cisplatin-induced
ER stress-associated apoptosis of SKOV3/DDP cells by increasing
calpain-1 expression.
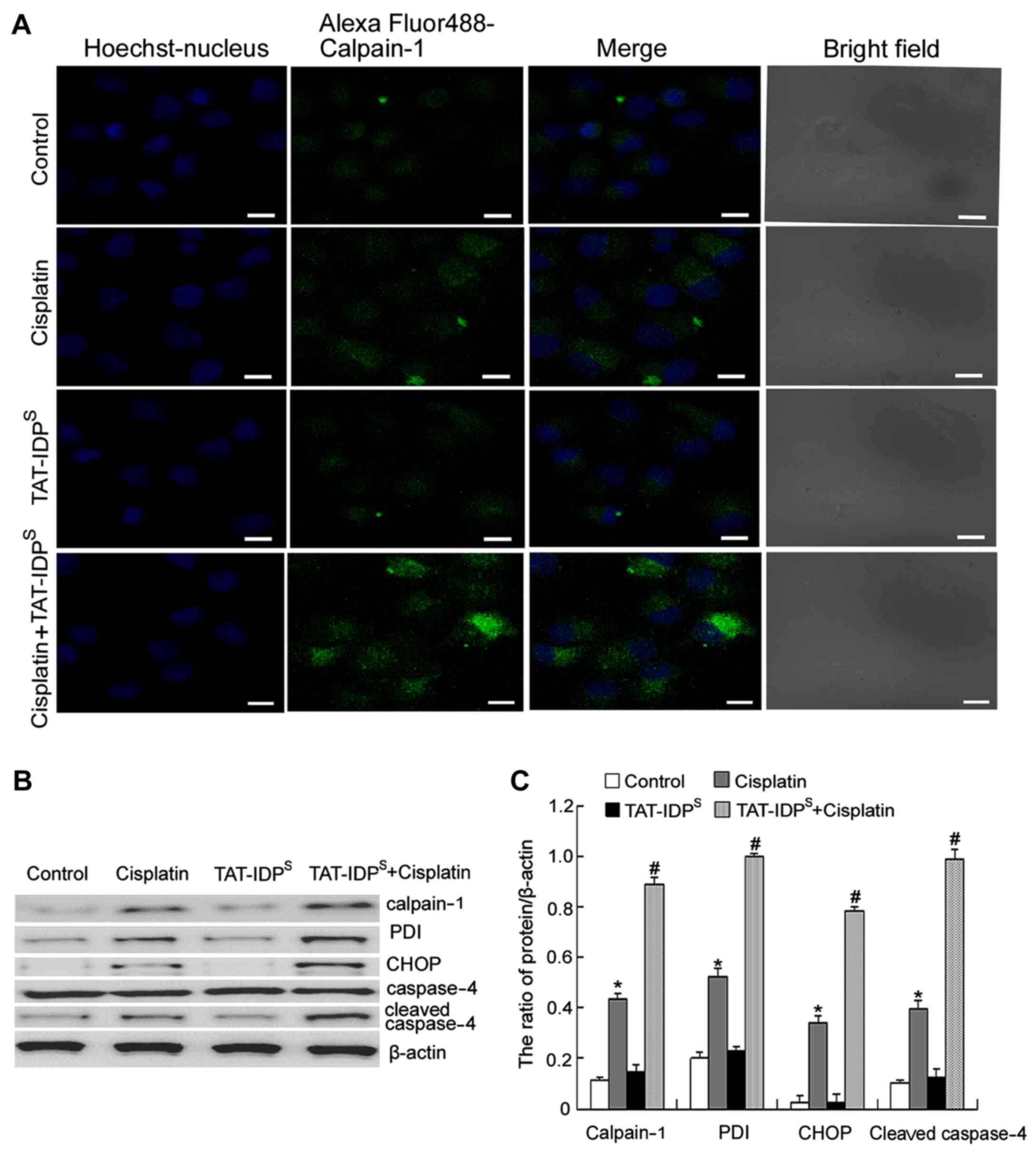 | Figure 3TAT-IDPS increases
cisplatin-induced endoplasmic reticulum stress-associated apoptosis
by increasing calpain-1 expression in SKOV3/DDP cells. (A)
Detection of calpain-1 in the cytoplasm of SKOV3/DDP cells exposed
to cisplatin (15 μg/ml) and/or TAT-IDPS (25
μM) for 24 h, under a confocal microscope (scale bar, 10
μm). (B) Western blot analysis was performed to detect the
expression levels of calpain-1, PDI, CHOP and cleaved caspase-4 in
SKOV3/DDP cells following treatment with cisplatin (15
μg/ml) and/or TAT-IDPS (25 μM) for 24 h. (C)
Semi-quantification of calpain-1, PDI, CHOP and cleaved caspase-4
expression. Data are presented as the mean ± standard deviation,
n=3. *P<0.05 vs. the control group;
#P<0.05 vs. the cisplatin group. CHOP,
CCAAT-enhancer-binding protein homologous protein; PDI, protein
disulfide isomerase; TAT-IDPS, TAT-fused IP3R-derived
peptide. |
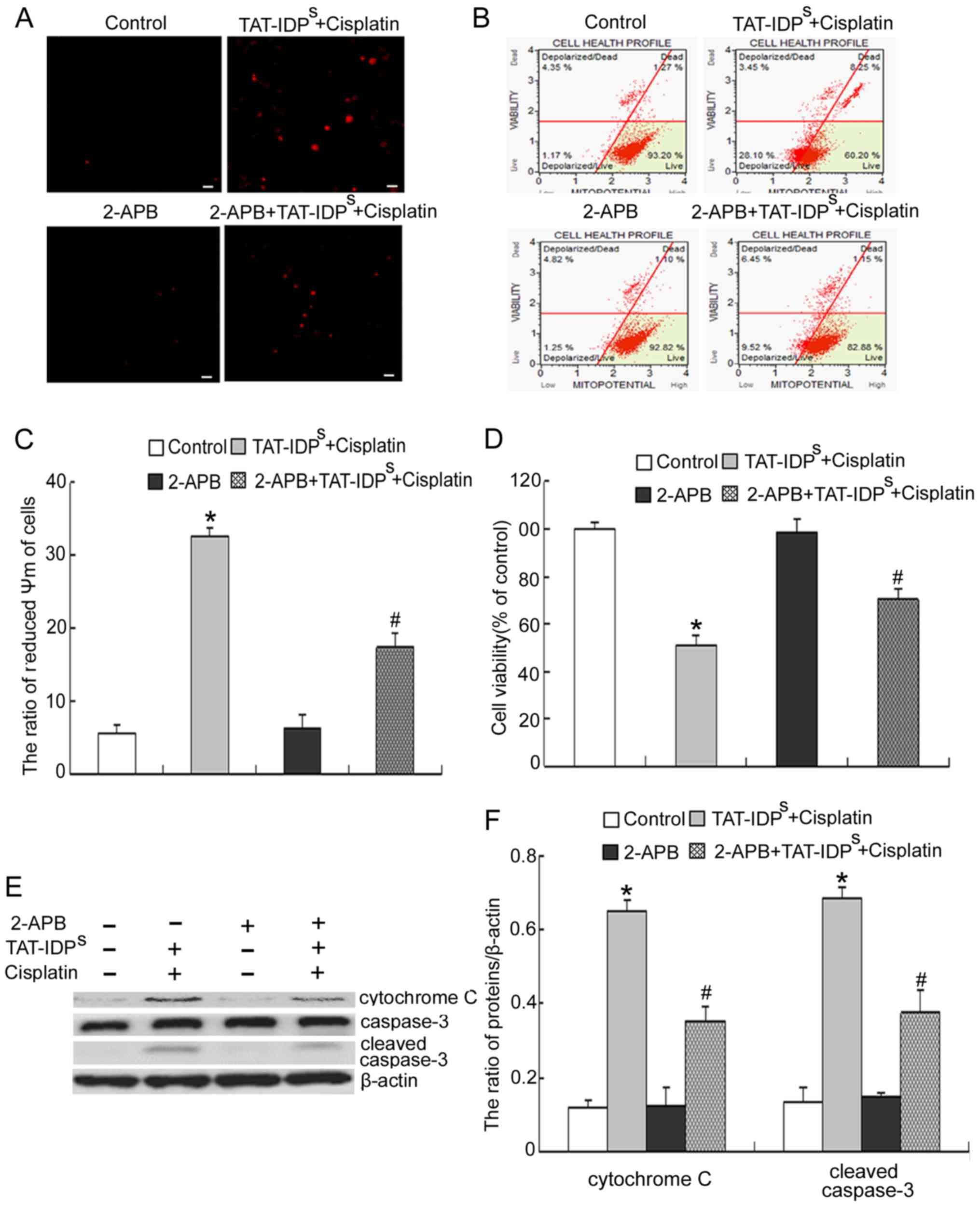
TAT-IDPS increases
cisplatin-induced mitochondria-mediated apoptosis of SKOV3/DDP
cells
Mitochondrial Ca2+ overload can lead to
opening of the mitochondrial inner membrane permeability transition
pore (MPTP) and can subsequently induce aberrant dissipation of
ΔΨm, thus resulting in cell apoptosis. Since an alteration in ΔΨm
is considered an early event in apoptosis (17), the present study monitored the
effects of TAT-IDPS and/or cisplatin on ΔΨm at 6 h.
SKOV3/DDP cells were treated with cisplatin (15 μg/ml)
and/or TAT-IDPS (25 μM) for 6 h, and alterations
in ΔΨm were determined using flow cytometry. TAT-IDPS
enhanced the cisplatin-induced decrease in ΔΨm (Fig. 4A and B). Finally, the expression
levels of mitochondrial apoptosis-associated proteins (cytochrome
c, Bcl-2, Bax and cleaved caspase-3) were detected using
western blotting. The results indicated that cotreatment with
cisplatin and TAT-IDPS for 24 h increased the
cisplatin-induced Bax/Bcl-2 ratio, and the cisplatin-induced
expression of cytochrome c and cleaved caspase-3 (Fig. 4C and D). These results suggested
that TAT-IDPS may further reduce the decrease in ΔΨm
induced by cisplatin, which may subsequently contribute to
mitochondria-mediated apoptosis of SKOV3/DDP cells.
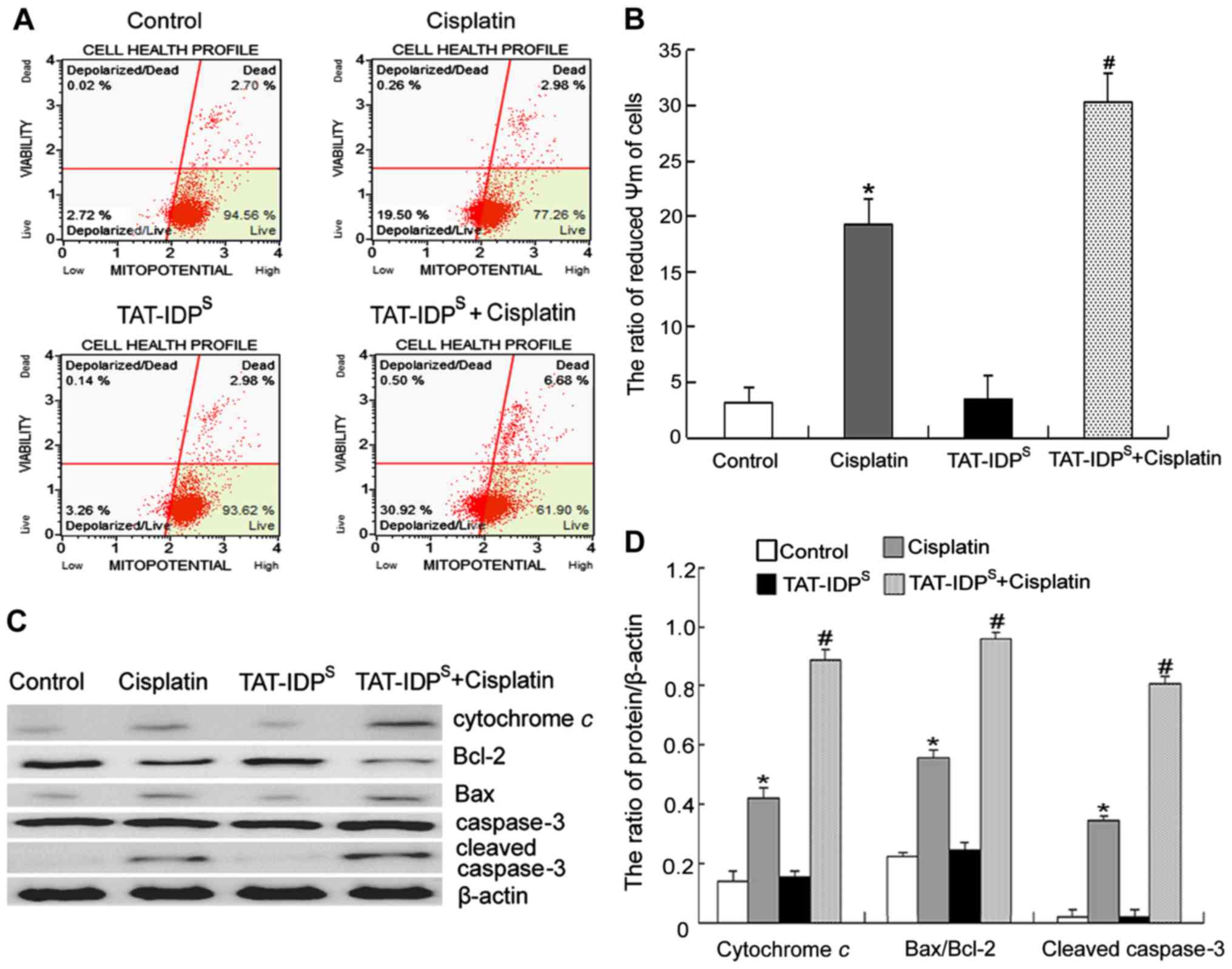 | Figure 4TAT-IDPS increases
cisplatin-induced mitochondria-mediated apoptosis of SKOV3/DDP
cells. (A) SKOV3/DDP cells were treated with cisplatin (15
μg/ml) and/or TAT-IDPS (25 μM) for 6 h.
ΔΨm was assessed by staining with MitoPotential dye and
7-aminoactinomycin D, and was analyzed using the Muse®
Cell Analyzer. (B) Quantification of ΔΨm. Data are presented as the
mean ± standard deviation, n=3. (C) Western blot analysis of the
expression of cytochrome c, Bcl-2, Bax and cleaved caspase-3
in SKOV3/DDP cells following treatment with cisplatin (15
μg/ml) and/or TAT-IDPS (25 μM) for 24 h.
(D) Semi-quantification of cytochrome c, Bcl-2, Bax, and
cleaved caspase-3 expression. Data are presented as the mean ±
standard deviation, n=3. *P<0.05 vs. the control
group; #P<0.05 vs. the cisplatin group. ΔΨm,
mitochondrial membrane potential; Bax, Bcl-2-associated X protein;
Bcl-2, B-cell lymphoma 2; TAT-IDPS, TAT-fused
IP3R-derived peptide. |
2-APB reduces the growth inhibition and
apoptosis induced by cotreatment with TAT-IDPS and
cisplatin
To further confirm that mitochondrial
Ca2+ overload caused by cotreatment with
TAT-IDPS and cisplatin derives from the ER, cells were
treated with 2-APB, an IP3R antagonist (18). Cisplatin and
TAT-IDPS-induced mitochondrial Ca2+ elevation
in SKOV3/DDP cells was markedly reduced in the presence of 2-APB
(Fig. 5A). In addition, 2-APB
reduced the decrease in ΔΨm induced by cisplatin and
TAT-IDPS (Fig. 5B and
C). Similarly, cisplatin and TAT-IDPS-induced growth
inhibition and apoptosis of SKOV3/DDP cells was markedly reduced in
the presence of 2-APB (Fig.
5D–F). Taken together, these results strongly indicated that
mitochondrial Ca2+ overload caused by cotreatment with
TAT-IDPS and cisplatin was derived from the ER.
Discussion
Bcl-2 is an anti-apoptotic protein that promotes
cell survival predominantly by inhibiting the release of cytochrome
c from the mitochondria via its BH3 domain. In addition,
Bcl-2 inhibits cell apoptosis by reducing Ca2+ release
from the ER via its BH4 domain. Recently, targeting the BH3 and BH4
domains to generate Bcl-2 inhibitors for cancer therapy has
attracted attention. Obatoclax is a BH3 mimetic that can induce
apoptosis of numerous types of cancer cell by interacting with the
Bcl-2 BH3 domain; this agent has been tested in a phase II clinical
trial in patients (19).
Furthermore, Deng et al reported that BDA-366, which is an
antagonist of the Bcl-2 BH4 domain, can induce apoptosis of human
myeloma U266 cells (20). The
results of the present study demonstrated that by targeting BH4,
TAT-IDPS increases cisplatin-induced apoptosis of
SKOV3/DDP cells (Fig. 2).
As a major intracellular Ca2+ store, the
ER serves a central role in intracellular Ca2+ signal
transduction, and controls cell death and survival. The release of
large amounts of Ca2+ from the ER promotes cell
apoptosis by affecting cytoplasmic and mitochondrial
Ca2+ homeostasis. There is strong evidence to suggest
that Bcl-2 directly inhibits proapoptotic Ca2+ release
from IP3R at the ER into the cytoplasm and mitochondria via its BH4
domain (21–23). TAT-IDPS can reverse the
Bcl-2-mediated inhibition of IP3-dependent Ca2+
elevation by targeting the BH4 domain, subsequently triggering
Ca2+-dependent apoptosis or increasing sensitivity
toward proapoptotic stimuli (15). The present study clearly
demonstrated that TAT-IDPS increased cisplatin-induced
cytoplasmic and mitochondrial Ca2+ elevation (Fig. 1).
Elevation of cytoplasmic Ca2+ levels can
lead to activation of a series of Ca2+-sensitive
enzymes. Calpain-1 is the target of cytoplasmic Ca2+,
which was originally termed μ-calpain based on the
Ca2+ concentration (μM range) required for its
optimal activity (24). Calpain-1
also participates in the apoptotic pathway triggered by ER stress.
Matsuzaki et al confirmed that Ca2+-dependent
calpain activation promoted neural cell death by inducing the
processing and activation of caspase-4 (25). According to the results of
confocal microscopy and western blot analyses, the present study
revealed that TAT-IDPS increased cisplatin-induced
calpain-1 expression (Fig. 3) and
increased cisplatin-induced ER stress-associated apoptosis of
SKOV3/DDP cells.
Mitochondria are exposed to high Ca2+
concentrations following the release of Ca2+ from the
ER. Accumulation of Ca2+ in the mitochondrial matrix can
lead to opening of the MPTP, which in turn causes mitochondrial
swelling and rupture, accompanied by dissipation of ΔΨm (26–28). The ΔΨm collapse further induces
rupture of the outer membrane and subsequent release of
mitochondrial apoptotic factors, including cytochrome c,
into the cytosol, finally leading to cell apoptosis (29,30). Our previous study demonstrated
that mitochondrial Ca2+ levels are closely associated
with cisplatin resistance in ovarian carcinoma cells (11). The present study demonstrated that
TAT-IDPS increased cisplatin-induced ΔΨm collapse and
mitochondrial-mediated apoptosis of SKOV3/DDP cells. Using 2-APB to
effectively block ER Ca2+ release, the present study
further confirmed that TAT-IDPS and cisplatin-induced
mitochondrial Ca2+ overload is derived from the ER
(Fig. 5A).
In conclusion, the results of the present study
indicated that TAT-IDPS increased the cisplatin-induced
elevation of cytoplasmic and mitochondrial Ca2+, which
further increased cisplatin-induced ER stress-mediated apoptosis by
increasing calpain-1 expression and activating the mitochondrial
apoptotic pathway. These results suggested that Ca2+
release from the ER may regulate cisplatin-induced SKOV3/DDP cell
apoptosis, providing a rationale for drugs that specifically
disrupt the Bcl-2-IP3R interaction as a novel tool for the
treatment of ovarian cancer.
Acknowledgments
The present study was supported by the National
Nature and Science Foundation of China (NSFC grant nos. 81372793,
81472419 and 81202552) and the Department of Education of Jilin
Province Project (grant no. 2016237). We thank Liwen Bianji for
editing the English in this manuscript.
References
|
1
|
Huang J, Liu K, Song D, Ding M, Wang J,
Jin Q and Ni J: Krüppel-like factor 4 promotes high-mobility group
box 1-induced chemotherapy resistance in osteosarcoma cells. Cancer
Sci. 107:242–249. 2016. View Article : Google Scholar :
|
|
2
|
Chen XX, Xie FF, Zhu XJ, Lin F, Pan SS,
Gong LH, Qiu JG, Zhang WJ, Jiang QW, Mei XL, et al:
Cyclin-dependent kinase inhibitor dinaciclib potently synergizes
with cisplatin in preclinical models of ovarian cancer. Oncotarget.
6:14926–14939. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Siddiqui WA, Ahad A and Ahsan H: The
mystery of BCL2 family: Bcl-2 proteins and apoptosis: an update.
Arch Toxicol. 89:289–317. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Lutz RJ: Role of the BH3 (Bcl-2 homology
3) domain in the regulation of apoptosis and Bcl-2-related
proteins. Biochem Soc Trans. 28:51–56. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Fan Z, Yu H, Cui N, Kong X, Liu X, Chang
Y, Wu Y, Sun L and Wang G: ABT737 enhances cholangiocarcinoma
sensitivity to cisplatin through regulation of mitochondrial
dynamics. Exp Cell Res. 335:68–81. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Wong M, Tan N, Zha J, Peale FV, Yue P,
Fairbrother WJ and Belmont LD: Navitoclax (ABT-263) reduces
Bcl-x(L)-mediated chemoresistance in ovarian cancer models. Mol
Cancer Ther. 11:1026–1035. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Han B, Park D, Li R, Xie M, Owonikoko TK,
Zhang G, Sica GL, Ding C, Zhou J, Magis AT, et al: Small-molecule
Bcl2 BH4 antagonist for lung cancer therapy. Cancer Cell.
27:852–863. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Nakajima A, Tsuji M, Inagaki M, Tamura Y,
Kato M, Niiya A, Usui Y and Oguchi K: Neuroprotective effects of
propofol on ER stress-mediated apoptosis in neuroblastoma SH-SY5Y
cells. Eur J Pharmacol. 725:47–54. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Krebs J, Agellon LB and Michalak M: Ca(2+)
homeostasis and endoplasmic reticulum (ER) stress: An integrated
view of calcium signaling. Biochem Biophys Res Commun. 460:114–121.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Liu N, Xu Y, Sun JT, Su J, Xiang XY, Yi
HW, Zhang ZC and Sun LK: The BH3 mimetic S1 induces endoplasmic
reticulum stress-associated apoptosis in cisplatin-resistant human
ovarian cancer cells although it activates autophagy. Oncol Rep.
30:2677–2684. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xu Y, Wang C, Su J, Xie Q, Ma L, Zeng L,
Yu Y, Liu S, Li S, et al: Tolerance to endoplasmic reticulum stress
mediates cisplatin resistance in human ovarian cancer cells by
maintaining endoplasmic reticulum and mitochondrial homeostasis.
Oncol Rep. 34:3051–3060. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Bonder DE and McCarthy KD: Astrocytic
Gq-GPCR-linked IP3R-dependent Ca2+ signaling does not
mediate neurovascular coupling in mouse visual cortex in vivo. J
Neurosci. 34:13139–13150. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Liu Z, Wild C, Ding Y, Ye N, Chen H, Wold
EA and Zhou J: BH4 domain of Bcl-2 as a novel target for cancer
therapy. Drug Discov Today. 21:989–996. 2016. View Article : Google Scholar :
|
|
14
|
Akl H, Monaco G, La Rovere R, Welkenhuyzen
K, Kiviluoto S, Vervliet T, Molgó J, Distelhorst CW, Missiaen L,
Mikoshiba K, et al: IP3R2 levels dictate the apoptotic sensitivity
of diffuse large B-cell lymphoma cells to an IP3R-derived peptide
targeting the BH4 domain of Bcl-2. Cell Death Dis. 4:e6322013.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zhong F, Harr MW, Bultynck G, Monaco G,
Parys JB, De Smedt H, Rong YP, Molitoris JK, Lam M, Ryder C, et al:
Induction of Ca2+-driven apoptosis in chronic
lymphocytic leukemia cells by peptide-mediated disruption of
Bcl-2-IP3 receptor interaction. Blood. 117:2924–2934. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Baudry M and Bi X: Calpain-1 and
Calpain-2: The Yin and Yang of Synaptic Plasticity and
Neurodegeneration. Trends Neurosci. 39:235–245. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Sun X, Xu H, Shen J, Guo S, Shi S, Dan J,
Tian F and Tian Y and Tian Y: Real-time detection of intracellular
reactive oxygen species and mitochondrial membrane potential in
THP-1 macrophages during ultrasonic irradiation for optimal
sonodynamic therapy. Ultrason Sonochem. 22:7–14. 2015. View Article : Google Scholar
|
|
18
|
Peppiatt CM, Collins TJ, Mackenzie L,
Conway SJ, Holmes AB, Bootman MD, Berridge MJ, Seo JT and Roderick
HL: 2-Aminoethoxydiphenyl borate (2-APB) antagonises inositol
1,4,5-trisphosphate-induced calcium release, inhibits calcium pumps
and has a use-dependent and slowly reversible action on
store-operated calcium entry channels. Cell Calcium. 34:97–108.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Schwartz-Roberts JL, Shajahan AN, Cook KL,
Warri A, Abu-Asab M and Clarke R: GX15-070 (obatoclax) induces
apoptosis and inhibits cathepsin D- and L-mediated autopha-gosomal
lysis in antiestrogen-resistant breast cancer cells. Mol Cancer
Ther. 12:448–459. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Deng J, Park D, Wang M, Nooka A, Deng Q,
Matulis S, Kaufman J, Lonial S, Boise LH, Galipeau J, et al:
BCL2-BH4 antagonist BDA-366 suppresses human myeloma growth.
Oncotarget. 7:27753–27763. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Akl H, La Rovere RM, Janssens A,
Vandenberghe P, Parys JB and Bultynck G: HA14-1 potentiates
apoptosis in B-cell cancer cells sensitive to a peptide disrupting
IP3 receptor/Bcl-2 complexes. Int J Dev Biol. 59:391–398. 2015.
View Article : Google Scholar
|
|
22
|
Parys JB: The IP3 receptor as a hub for
Bcl-2 family proteins in cell death control and beyond. Sci Signal.
7:pe42014. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Akl H, Vervloessem T, Kiviluoto S,
Bittremieux M, Parys JB, De Smedt H and Bultynck G: A dual role for
the anti-apoptotic Bcl-2 protein in cancer: Mitochondria versus
endoplasmic reticulum. Biochim Biophys Acta. 1843:2240–2252. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Pu X, Storr SJ, Ahmad NS, Chan SY, Moseley
PM, Televantou D, Cresti N, Boddy A, Ellis IO and Martin SG:
Calpain-1 is associated with adverse relapse free survival in
breast cancer: A confirmatory study. Histopathology. 68:1021–1029.
2016. View Article : Google Scholar
|
|
25
|
Matsuzaki S, Hiratsuka T, Kuwahara R,
Katayama T and Tohyama M: Caspase-4 is partially cleaved by calpain
via the impairment of Ca2+ homeostasis under the ER
stress. Neurochem Int. 56:352–356. 2010. View Article : Google Scholar
|
|
26
|
Rutter GA and Rizzuto R: Regulation of
mitochondrial metabolism by ER Ca2+ release: An intimate
connection. Trends Biochem Sci. 25:215–221. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
von Stockum S, Giorgio V, Trevisan E,
Lippe G, Glick GD, Forte MA, Da-Rè C, Checchetto V, Mazzotta G,
Costa R, et al: F-ATPase of Drosophila melanogaster forms
53-picosiemen (53-pS) channels responsible for mitochondrial
Ca2+-induced Ca2+ release. J Biol Chem.
290:4537–4544. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Fülöp L, Rajki A, Maka E, Molnár MJ and
Spät A: Mitochondrial Ca2+ uptake correlates with the
severity of the symptoms in autosomal dominant optic atrophy. Cell
Calcium. 57:49–55. 2015. View Article : Google Scholar
|
|
29
|
Hajnóczky G, Csordás G, Das S,
Garcia-Perez C, Saotome M, Sinha Roy S and Yi M: Mitochondrial
calcium signalling and cell death: Approaches for assessing the
role of mitochondrial Ca2+ uptake in apoptosis. Cell
Calcium. 40:553–560. 2006. View Article : Google Scholar
|
|
30
|
Giorgi C, Baldassari F, Bononi A, Bonora
M, De Marchi E, Marchi S, Missiroli S, Patergnani S, Rimessi A,
Suski JM, et al: Mitochondrial Ca(2+) and apoptosis. Cell Calcium.
52:36–43. 2012. View Article : Google Scholar : PubMed/NCBI
|