Introduction
Osteoarthritis (OA) is the most prevalent
degenerative disease of the articulating joints, and deterioration
of cartilage extracellular matrix (ECM) homeostasis is one of its
pathological characteristics. As the sole cell type in cartilage,
chondrocytes are responsible for regulating ECM anabolism and
catabolism. Therefore, chondrocyte activities, including autophagy,
apoptosis and proliferation, are crucial for preserving cartilage
homeostasis. For example, autophagy, a lysosomal degradation
process, is essential for survival, differentiation, development
and homeostasis (1). It has been
reported to control the secretion of type II collagen (Col 2), one
of the primary components of cartilage ECM. In the absence of
autophagy in growth-plate chondrocytes, Col 2 decreased due to the
abnormal accumulation of collagen molecules in the endoplasmic
reticulum (2). Increased
autophagy is an adaptive response to protect chondrocytes from
stresses (3). In addition, the
blockade of chondrocyte apoptosis and the promotion of chondrocyte
proliferation may be potential therapeutic strategies to treat OA.
Berberine may ameliorate cartilage degeneration from OA by
promoting cell survival and matrix production of chondrocytes in
interleukin (IL)-1β-stimulated articular chondrocytes and in a rat
OA model (4). Morroniside exerted
chondroprotective effects on OA chondrocytes by promoting cell
proliferation and ECM synthesis (5). Hyaluronic acid may suppress
chondrocyte apoptosis in a dose-dependent manner in IL-1β-induced
OA, which may be an underlying mechanism of the clinical action of
intra-articular injection of hyaluronic acid in the treatment of OA
(6).
The majority of cell signaling molecules are
involved in regulating OA chondrocyte autophagy, proliferation and
apoptosis, including protein kinase B (PKB/Akt) and mechanistic
target of rapamycin (mTOR) and extracellular signal-regulated
kinase (ERK). For example, mTOR targeting prevents physical harm to
joints in diabetic mice by autophagy activation (7). Cartilage-specific deletion of mTOR
upregulates autophagy and protects mice from OA (8). Akt and ERK contribute to the
chondroprotective effect of morroniside on OA chondrocytes by
promoting chondrocyte proliferation and ECM synthesis (5,9,10).
Therefore, present study predominantly focuses on elucidating the
role and regulatory mechanism of complicated cell signaling
molecules involved in chondrocyte events.
Various animal serums are added to chondrocyte
culture media as they contain numerous nutrient components and
cytokines. Notably, in the mid-1990s, a novel therapeutic strategy,
the intra-articular injection of autologous conditioned serum (ACS;
Orthokine) was developed, and has recently been demonstrated to be
safe and effective in a number of clinical studies, and is widely
adopted in Europe (11,12). ACS contains enriched
anti-inflammatory cytokines, including inter- leukin (IL)-1Ra,
IL-10 and IL-13, and various growth factors, while maintaining low
concentrations of pro-inflammatory cytokines, such as IL-1β and
tumor necrosis factor (TNF)-α. However, the elevated concentrations
of pro-inflammatory cytokines in ACS have no adverse effects on
proteoglycan turnover in the cartilage explant cultures (12,13). As a result, understanding the
regulatory mechanism of serum is needed to elucidate its effects on
OA therapy.
The present study aims to evaluate the effect of
different types of bovine fetal serum on human OA chondrocyte
proliferation, autophagy and ECM metabolism, and to further
investigate the regulatory mechanism associated with Akt and mTOR
signaling molecules.
Materials and methods
Reagents and antibodies
Inhibitors [mTOR inhibitor Rapamycin (cat. no. 9904)
and PI3K/Akt inhibitor LY294002 (cat. no. 9901)] and antibodies
against mTOR (cat. no. 2983), phosphorylated (p)-mTOR (Ser2448;
cat. on. 2971), Akt (cat. no. 4691), p-Akt (Ser473; cat. no. 4051),
B-cell lymphoma-2 (Bcl-2, cat. no. 2870), p-S6 (Ser235/236; cat.
no. 2211), Beclin 1 (cat. no. 4122), AMP-activated protein kinase
(AMPK)α (cat. no. 5831), p-AMPKα (Thr172; cat. no. 2535) and LC-3B
(cat. no. 3868) were purchased from Cell Signaling Technology Inc.
(Danvers, MA, USA). Anti-GAPDH antibody (cat. no. 0411) was
purchased from Santa Cruz Biotechnology, Inc. (Dallas, TX, USA).
Antibodies targeting Col 2 (cat. no. WH0001280M1) and aggrecan
(cat. no. SAB4500662) were purchased from Sigma-Aldrich (Merck
KGaA, Darmstadt, Germany). Anti-proliferating cell nuclear antigen
antibody (cat. no. BM0104) was purchased from Wuhan Boster
Biological Technology., Ltd. (Wuhan, Hubei, China). Secondary
antibodies (goat anti-mouse; cat. no. SA00001-1; and goat
anti-rabbit; cat. no. SA00001-2) were purchased from Proteintech
Group, Inc. (Chicago, IL, USA). Other commercially available
reagents of the highest grade were obtained.
Human OA chondrocyte isolation and
culture
Subsequent to receiving all patient consent and, in
accordance with the ethical guidelines of Zhongshan Hospital of
Xiamen University (Xiamen, China), human OA articular cartilage was
obtained from advanced OA patients who were under- going total knee
replacement surgery in Zhongshan Hospital of Xiamen University
(Xiamen, China). Under sterile conditions, the articular cartilage
on the femoral condyle and tibial plateau was minced into small
pieces and digested with 0.2% type II collagenase as described
previously (14,15). The isolated chondrocytes were
cultured in Dulbecco's modified Eagle's medium/F12 medium with 10%
fetal bovine serum (FBS; v/v) plus 1% penicillin/streptomycin at
37°C for 72 h in a water-saturated atmosphere of 5% CO2.
When chondrocytes were grown to 70–80% confluence, the cells were
plated in 60-mm Petri dishes (2×106 cells/dish) or
96-well plates (3×104 cells/well), followed by the
evaluation of chondrocytes using immunohistological techniques and
treatment with or without different inhibitors. Ethical approval
for the study was obtained from the Ethics Committee of the Medical
School, Xiamen University (Xiamen, China).
Cell viability assay
Cells were plated in 96-well plates
(2×105 cells/well) and starved with serum-free medium
for 24 h. The medium was then replaced with fresh medium with
different concentrations of serum in the presence or absence of
inhibitors for a further 24 h. The Cell Counting Kit-8 (CCK8) assay
(cat. no. C0037; Beyotime Institute of Biotechnology, Haimen,
China) was then performed, as described previously (16). The end product was quantified
spectrophotometrically at a wavelength of 450 nm. The optical
density values correspond to the number of viable cells (17).
Western blot analysis
Cells were collected by centrifugation (825 × g, 7
min) at room temperature and lysed in Radioimmunoprecipitation
assay lysis buffer containing 1% Triton X-100, 1% deoxycholate and
0.1% SDS (premixed solution; cat. no. P0013B, Byeotime Institute of
Biotechnology). Lysates were then centrifuged at 19,071 × g or 30
min at 4°C and the protein concentration was determined using a
Pierce Bicinchoninic acid Protein assay kit (Thermo Fisher
Scientific, Inc., Waltham, MA, USA). Protein extracts were
subjected to SDS-PAGE (8–12% according to the molecular weight of
the protein) and transferred to polyvinylidene fluoride membranes
(60 min, 100 V; EMD Millipore, Billerica, MA, USA) as described
previously (18). The membranes
were incubated with the indicated primary antibodies at 4°C
overnight and corresponding secondary antibodies at room
temperature for 1–2 h in succession. The signal was monitored using
a chemiluminescent detection system according to the manufacturer's
protocol (cat. no. WBKLS0500; EMD Millipore).
Laser-scanning confocal microscope
Chondrocytes were fixed in 4% paraformaldehyde after
harvesting. For staining of the endogenous LC-3B proteins, the
cells were incubated with anti-LC-3B antibody (dilution, 1:200)
followed by Cy3-conjugated secondary antibody (Merck KGaA) as
described previously (18). The
cells were stained with 50 mg/ml 4′,6-diamidino-2-phenylindole
(Sigma-Aldrich; Merck KGaA) to visualize the nuclei simultaneously
at room temperature for 5 min (18). The stained cells were finally
visualized under a confocal microscope (Leica TCS SP2 SE; Leica
Microsystems GmbH, Wetzlar, Germany).
Statistical analysis
Experimental data were formulated as the means ±
standard error of the mean of three independent samples.
Statistical analyses were performed with GraphPad Prism 5 software
(GraphPad Software, Inc., La Jolla, CA, USA), using one-way ANOVA.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Effect of different concentrations of
serum on cell proliferation and ECM synthesis in human OA
chondrocytes
To investigate the effect of serum on OA chondrocyte
proliferation, cultured human OA chondrocytes were starved for 24 h
and treated with different concentrations of serum (1, 5, 10 and
20%) for 24 h. The cell viability was measured using a CCK8 assay.
Fig. 1A indicates that
chondrocyte viability significantly increased in 5 and 20%
serum-treated groups without any alteration observed in the other
groups (P<0.05 vs. the serum-free group). Proliferation cell
nuclear antigen (PCNA), Bcl-2 and S6 are predominant biomarkers of
cell proliferation, and their expression levels were detected by
western blotting. PCNA, Bcl-2 p-S6 expression levels increased in
the 5% serum-treated group (Fig.
1B; P<0.05 vs. the serum-free group). 1, 10 or 20% serum led
to increased Bcl-2 and p-S6 expression levels (Fig. 1B, P<0.05 vs. the serum-free
group). In addition, the expression levels of the predominant
components of the ECM, aggrecan and Col 2, were detected by western
blotting analysis. The 5% serum simultaneously enhanced aggrecan
and Col 2 expression levels in the human OA chondrocytes (Fig. 1C; P<0.05 vs. the serum-free
group). The 10% serum only enhanced the expression level of Col 2
(Fig. 1C; P<0.05 vs. the
serum-free group). Consequently, only 5% serum simultaneously
significantly enhanced cell viability along with the increase in
PCNA, Bcl-2 and p-S6 expression levels in human OA chondrocytes,
and promoted ECM synthesis via the increase of aggrecan and Col 2
expression levels in human OA chondrocytes.
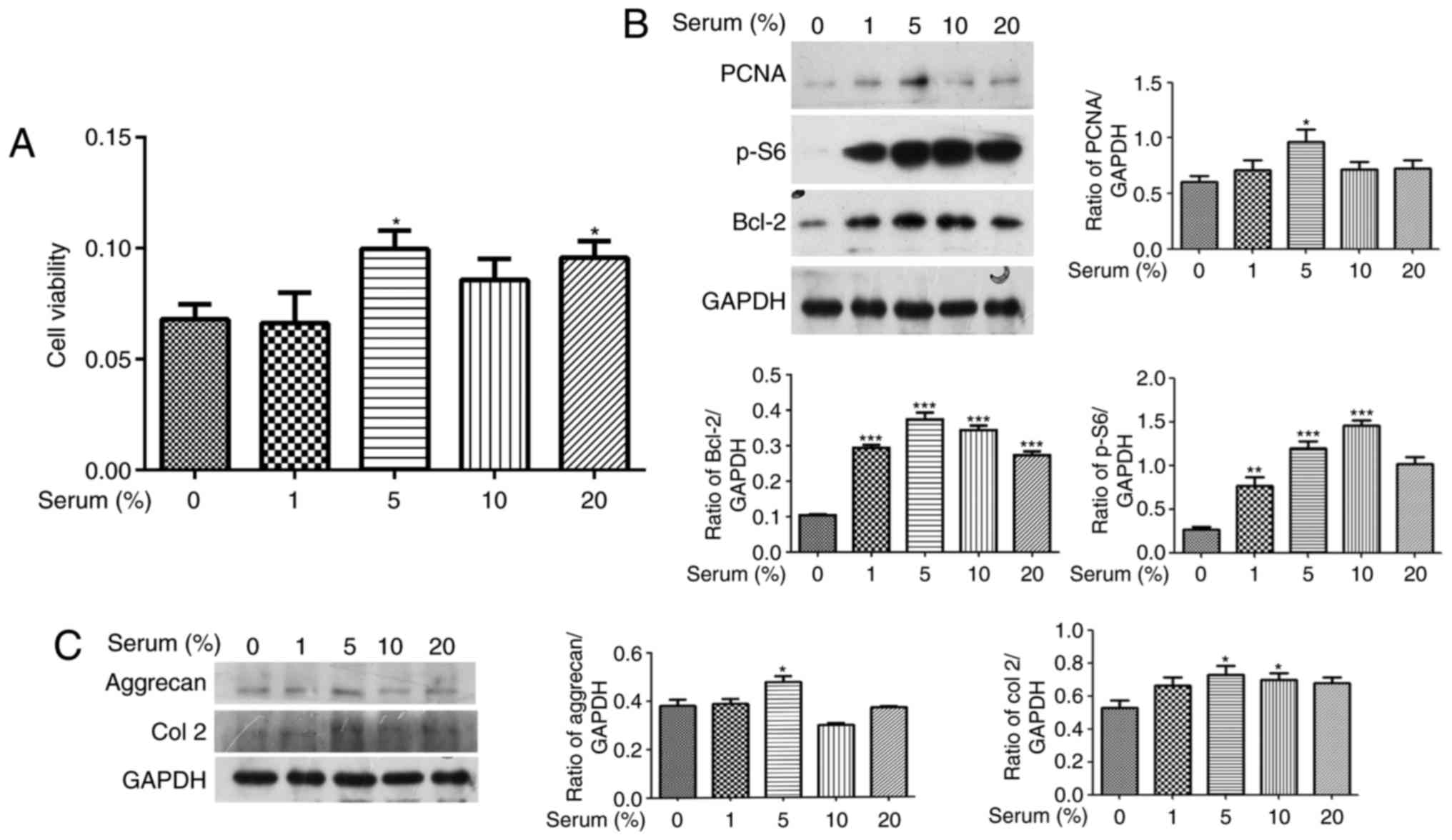 | Figure 1Effect of different concentrations of
serum on cell proliferation in human OA chondrocytes. Cultured
human OA chondrocytes were starved for 24 h and treated with
different concentrations of serum (1, 5, 10 and 20%) for 24 h. (A)
Cell viability was evaluated using a Cell Counting Kit 8 assay. (B)
The expression levels of PCNA, p-S6, and Bcl-2 were detected by
western blotting. The blots were normalized to an endogenous
protein (GAPDH). (C) The expression levels of aggrecan and Col 2
were detected with western blotting. The blots were normalized to
an endogenous protein (GAPDH). The data are representative of three
independent experiments, each yielding similar results
(*P<0.05, **P<0.01 and
***P<0.001 vs. serum-free group). OA, osteoarthritis;
PCNA, proliferation cell nuclear antigen; p-, phosphorylated;
Bcl-2, B-cell lymphoma 2; Col 2, type II collagen. |
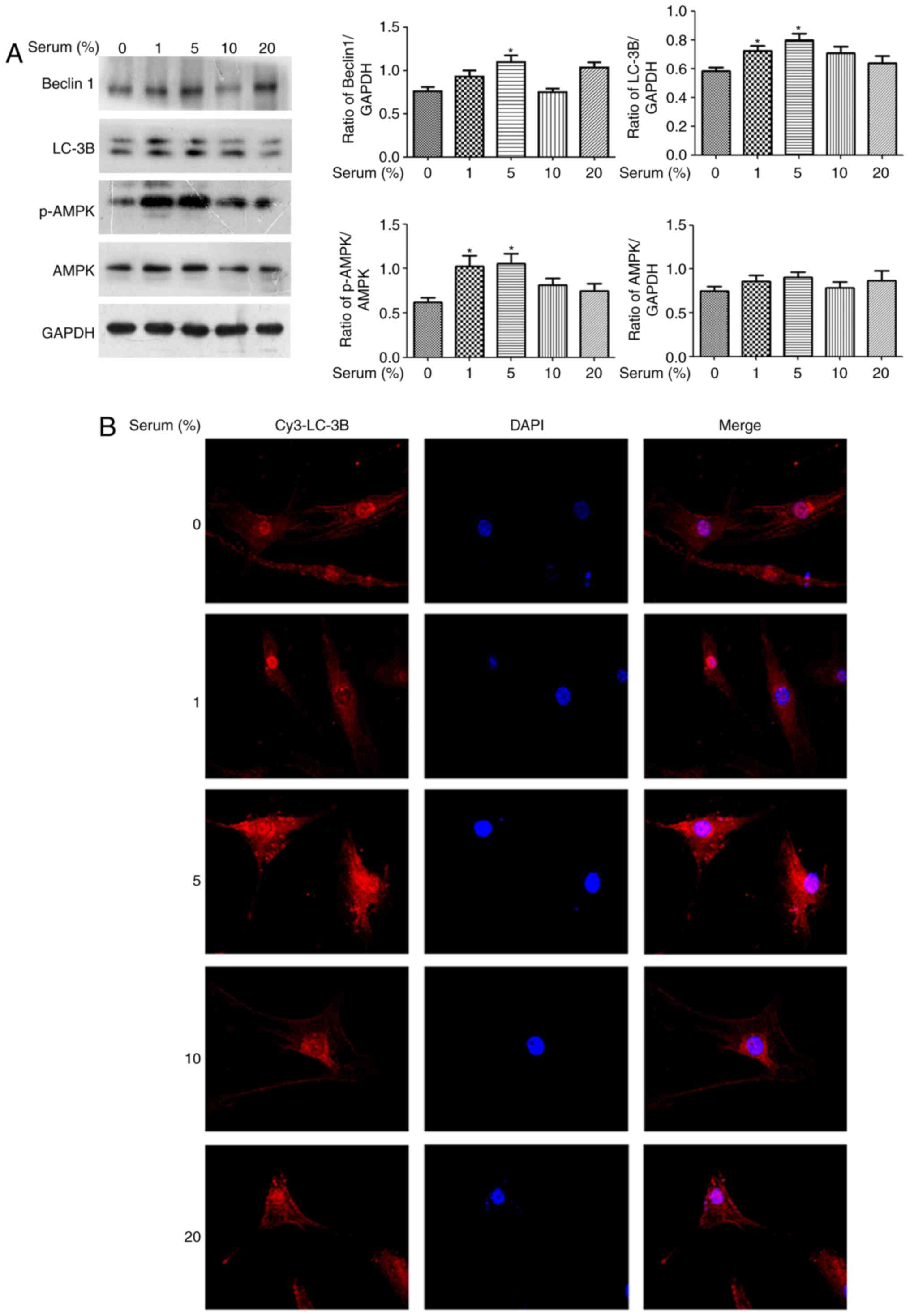
Effect of different concentrations of
serum on autophagy in human OA chondrocytes
As autophagy is essential for survival,
differentiation, development and homeostasis (1), the present study determined whether
serum may regulate autophagy of OA chondrocytes. Cultured human OA
chondrocytes were starved for 24 h and treated with different
concentrations of serum (1, 5, 10 and 20%) for 4 h. These
autophagy-associated markers, including LC-3B, Beclin 1 and AMPKα,
were detected by western blotting. Fig. 2A demonstrated that LC-3B II,
Beclin 1 and p-AMPKα expression levels increased in the 5%
serum-treated group (P<0.05 vs. the serum-free group). The 1%
serum led to increased LC-3B II and p-AMPKα expression levels.
Additionally, the results of laser-scanning confocal microscopy
demonstrated that the endogenous LC-3B expression level with
anti-LC-3B antibody in 5% serum-treated OA chondrocytes was higher
than the other concentration serum-treated groups, compared with
the serum-free group (Fig. 2B).
Overall, it was identified that only 5% serum could simultaneously
enhance the expression levels of LC-3B, Beclin 1 and p-AMPKα in
human OA chondrocytes.
Akt and mTOR participate in regulating
cell proliferation and autophagy in 5% serum-treated human OA
chondrocytes
Akt and mTOR have been implicated in OA
pathogenesis; thus, the phosphorylation levels of mTOR and Akt were
investigated in different concentrations of serum-treated human OA
chondrocytes. Fig. 3A
demonstrates that 5% serum caused a significant decrease in p-mTOR
andincrease in p-Akt, while the other concentrationss did not alter
the p-mTOR and p-Akt expression levels (P<0.05 vs. the
serum-free group). To further investigate the role of mTOR and Akt
on cellular proliferation and autophagy in 5% serum-treated human
OA chondrocytes, the starved cells were treated with 50 nM
rapamycin (mTOR inhibitor) or 10 mM LY294002 (phosphoinositide
3-kinase; PI3K/Akt inhibitor) for 2 h, respectively. This was
followed by 5% serum treatment at 37°C for 4 h in a water-saturated
atmosphere of 5% CO2. The results of Fig. 3B demonstrate that rapamycin
reduced PCNA and p-S6 expression levels in the 5% serum-treated
group (P<0.01 vs. the untreated 5% serum-treated group), while
the Bcl-2 expression level did not significantly change. LY294002
significantly inhibited the level of Bcl-2 expression in the 5%
serum-treated groups without any alterations to the PCNA and p-S6
expression levels (Fig. 3B;
P<0.001 vs. the untreated 5% serum-treated group). Furthermore,
Fig. 3C demonstrated that
rapamycin led to the increase in LC-3B II expression level in the
5% serum-treated group without any alteration to the p-AMPKα and
Beclin 1 expression levels (P<0.0001 vs. the untreated 5%
serum-treated group). Beclin 1 and p-AMPKα expression levels
increased in the LY294002-treated 5% serum-treated group, while
LC-3B II did not significantly change (P<0.01 vs. the untreated
5% serum-treated group). Therefore, the addition of either
rapamycin or LY294002 modulated cell proliferation and autophagy in
human OA chondrocytes, by targeting different signaling
molecules.
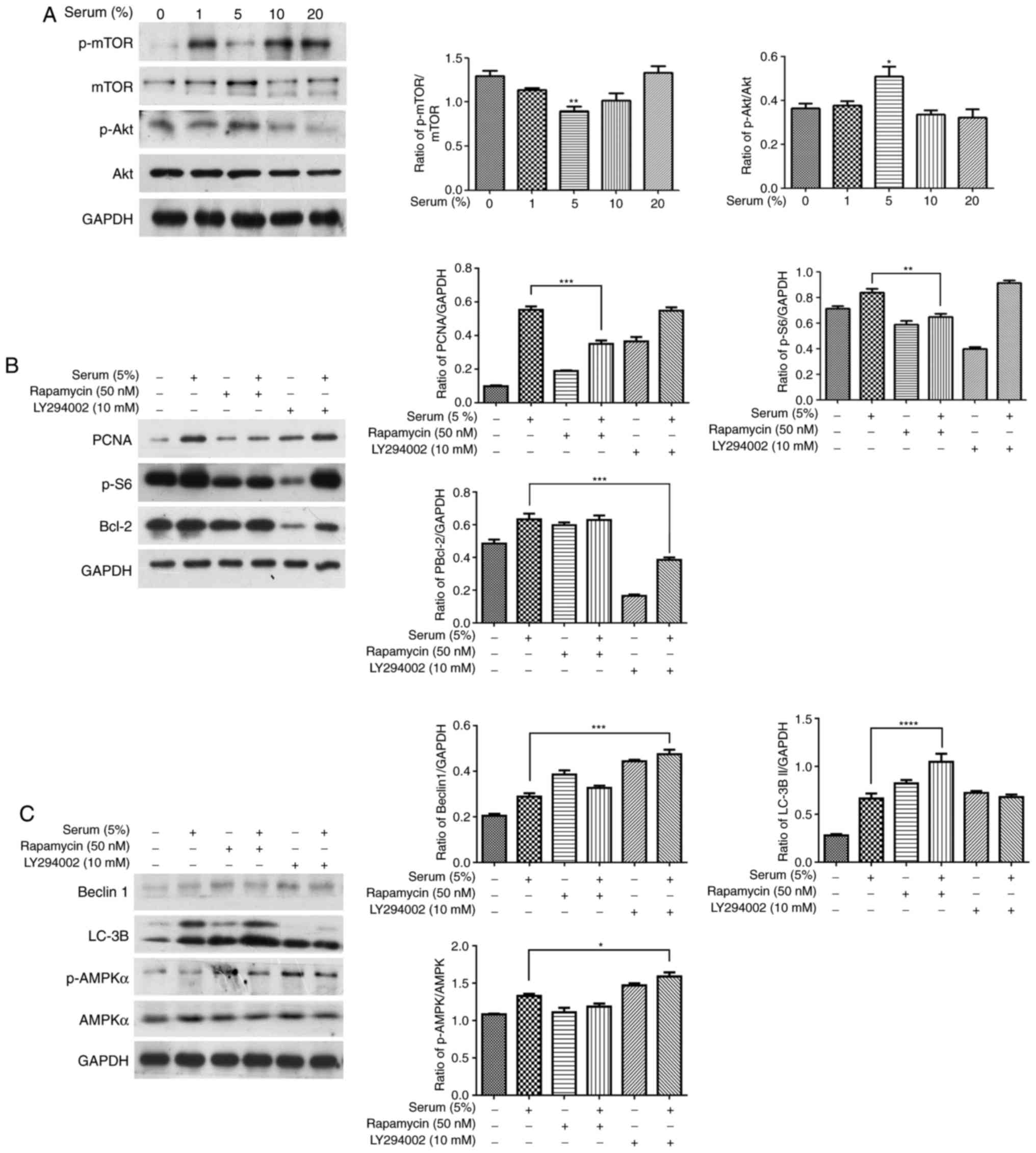 | Figure 3Effect of Akt and mTOR inhibitors on
serum-associated cell proliferation and autophagy in human OA
chondrocytes. (A) Cultured human OA chondrocytes were starved for
24 h and treated with different concentrations of serum (1, 5, 10
and 20%) for 24 h. The expression levels of Akt, p-Akt, mTOR and
p-mTOR were detected with western blotting. (B and C) The starved
chondrocytes were pretreated with 50 nM rapamycin (mTOR inhibitor)
or 10 mM LY294002 (PI3K/Akt inhibitor) for 2 h, respectively,
followed with 5% serum for 4 h. The expression levels of PCNA,
p-S6, Bcl-2, Beclin 1, LC-3B, p-AMPKα and AMPKα were detected by
western blotting. The blots were normalized to an endogenous
protein (GAPDH). The data are representative of three independent
experiments, each yielding similar results (*P<0.05,
**P<0.01, ***P<0.001 and
****P<0.0001 vs. serum-free group or untreated 5%
serum-treated group). mTOR, mechanistic target of rapamycin; OA,
osteoarthritis; p-, phosphorylated; PI3K, phosphoinositide
3-kinase; PCNA, proliferation cell nuclear antigen; Bcl-2, B-cell
lymphoma 2. |
Discussion
In the present study, the effect of different
concentrations of serum on cell events was investigated in human OA
chondrocytes. It is demonstrated that only 5% serum was able to
simultaneously regulate cell proliferation, ECM synthesis and
autophagy induction in human OA chondrocytes, compared with the
other concentrations. Notably, decreased mTOR and increased Akt
expression levels were observed in the 5% serum-treated group.
Furthermore, either mTOR or Akt inhibition modulated cell
proliferation and autophagy by targeting different signaling
molecules in human OA chondrocytes. Combined with the findings of
previous studies, it is proposed that 5% serum promotes cell
proliferation via the Akt/Bcl-2 axis and induces autophagy via the
mTOR/LC3B axis in human OA chondrocytes.
Although 10% serum is always used to culture
chondrocytes, certain studies indicate that chondrocytes cultured
with serum-free medium exhibit no proliferation difference when
compared with those cultured with 10% FBS (19). Human nasal chondrocytes
proliferate at comparable rates in different serum conditions, with
no statistically significant differences observed between the lower
percentage of autologous serum (2%) and 10% FBS (20). In the present study, treatment
with 5% serum, not 10%, simultaneously resulted in increased cell
proliferation, autophagy and ECM synthesis in human OA
chondrocytes. Therefore, it is proposed that 5% serum may be more
suitable for culturing chondrocytes than 10% serum, at least in
human OA chondrocytes.
Under high nutrient conditions, mTOR suppresses cell
autophagy by phosphorylating autophagy related 13 (Atg13) to
prevent the interaction between Atg13 and unc-51 like autophagy
activating kinase (ULK), or phosphorylating ULK Ser757 to disrupt
the interaction between AMPK and ULK (21). Conversely, activated mTORC1, due
to the inactivation of TSC complex subunit 2 with Akt, triggers the
mTOR/S6 axis to regulate protein synthesis, promoting cell
proliferation (22–24). Therefore, activated mTOR may
perform different functions in cellular processes, including
promoting cell proliferation and suppressing autophagy induction.
In the present study, the cell viability, PCNA, Bcl-2, aggrecan and
Col 2 expression levels increased in the 5% serum-treated group in
Fig. 1, indicating that 5% serum
may promote cell proliferation and ECM synthesis in human OA
chondrocytes. However, 5% serum led to a significantly decreased
p-mTOR expression level, implying that mTOR/S6-associated
proliferation may be blocked and that decreased levels of mTOR
expression, by 5% serum, did not inhibit cell proliferation. By
contrast, the increase of LC-3B II, Beclin 1 and p-AMPKα expression
levels in the 5% serum-treated group indicated that 5% serum may
induce autophagy. Furthermore, the inhibitor of mTOR caused a
significant increase in LC-3B II expression levels, demonstrating
that decreased mTOR in 5% serum-treated chondrocytes may be
involved in autophagy induction of OA chondrocytes. Thus, mTOR may
be involved in autophagy induction in 5% serum-treated human OA
chondrocytes, but not cell proliferation. The regulation of mTOR on
chondrocyte autophagy is well known. Chondrocyte autophagy is
stimulated by hypoxia-inducible factor-1-dependent AMPK activation
and mTOR suppression (25).
Inhibition of the PI3K/AKT/mTOR signaling pathway can promote
autophagy of articular chondrocytes and attenuate inflammatory
response in rats with OA (26).
The intra-articular injection of rapamycin, an mTOR inhibitor,
reduced mTOR expression levels, leading to a delay in articular
cartilage degradation in an OA murine model (27). Therefore, the present study
proposed that mTOR may predominantly be involved in regulating
autophagy induction in 5% serum-treated chondrocytes.
The activation of Akt increases in normal
chondrocytes more than in OA chondrocytes, and is required for
basal and insulin-induced Col 2 expression in chondrocytes
(28). Elevated production of
tribbles pseudokinase 3, an inhibitor of Akt activation, has been
reported to increase in OA chondrocytes (29). In addition, previous studies
demonstrated that increased Akt expression levels by extracellular
stimuli may protect articular cartilage from OA deterioration by
promoting cell survival in human or rat OA chondrocytes (4,5).
Thus, Akt is a positive element for chondroprotection. Consistent
with these studies, 5% serum significantly enhanced p-Akt
expression levels concomitantly with Bcl-2 depression caused by
LY294002, exhibiting the promoting effect of Akt on
serum-associated cell proliferation of human OA chondrocytes.
Therefore, 5% serum is hypothesized to promote cell proliferation,
to a lesser extent, by triggering the Akt/Bcl-2 signal axis.
However, Hahn-Windgassen et al (30) demonstrated that Akt is a negative
regulator of AMPK. Metformin, an AMPK activator, decreased the
phosphorylation of Akt in bladder cancer cells (31). Notably, Galasso et al
(32) reported that in OA human
cartilage, increased phosphatase and tensin homolog (Akt
inhibitor), AMPK and autophagy reflected the chondrocyte responses
observed during starvation and steroid depletion (32). The present data that LY294002 may
upregulate p-AMPKα in 5% serum-treated human OA chondrocytes was
consistent with the above-mentioned studies. Therefore, Akt may
reduce autophagy induction by suppressing p-AMPKα. However, the
induced autophagy of chondrocytes eventually increased in the 5%
serum-treated group, implying that increased Akt may predominantly
promote proliferation, rather than exert inhibitory effects on
autophagy induction in 5% serum-treated human OA chondrocytes,
which may be due to being interfered with by other signaling
pathways, such as the mTOR/LC-3B axis.
The PI3K/Akt signaling pathway is an important
activator of mTOR, although Akt-independent regulation of mTOR in
response to mitogens, nutrient availability and conditions that
deplete intracellular energy in cancer have been reported (33). Multiple signaling pathways are
involved in regulating mTOR independently of Akt, such as
LKB1/AMPK, AMPK/mTORC1, and calmodulin-dependent protein kinase
kinase β/AMPK signaling pathways (33). Furthermore, mTOR inhibition may
result in feedback activation of Akt in cancer (33–35), consistent with the present data of
the opposite effect of 5% serum on Akt and mTOR in human OA
chondrocytes (data not shown). Future studies are required to
determine the interaction between Akt and mTOR on cell processes in
5% serum-treated human OA chondrocytes.
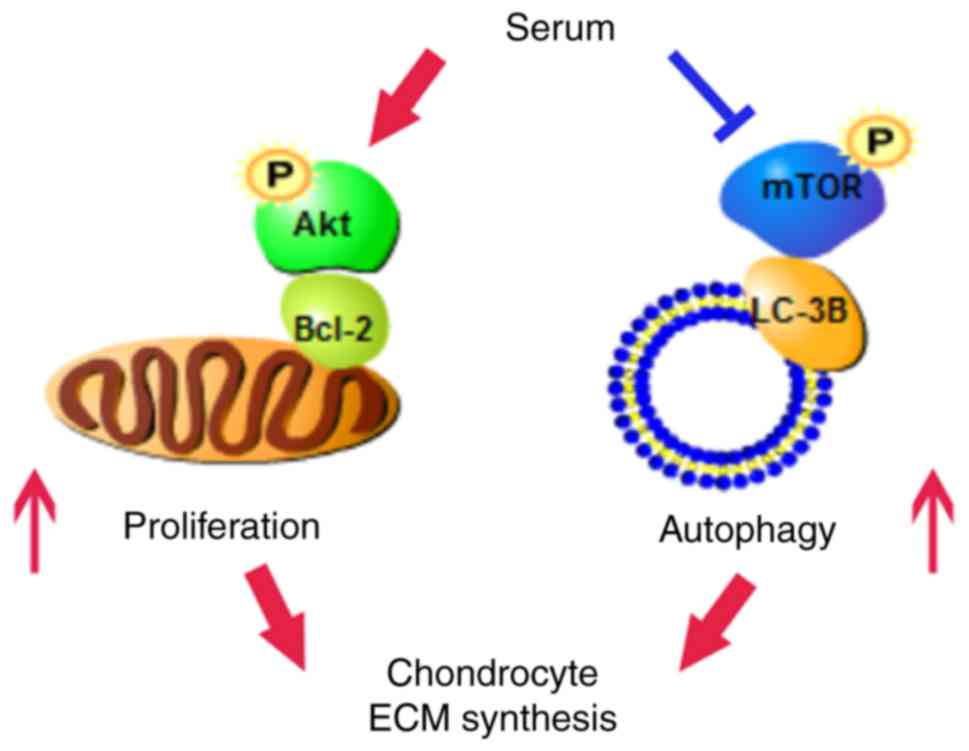
In conclusion, 5% serum may simultaneously promote
cell proliferation and autophagy induction of OA chondrocytes with
the increase of ECM synthesis by upregulating Akt and
downregulating mTOR signaling molecules. Furthermore, decreased
mTOR expression levels promoted autophagy induction in human OA
chondrocytes via the mTOR/LC-3B axis, and increased Akt enhanced
cell proliferation via the Akt/Bcl-2 axis (Fig. 4). Consequently, the effect of
increased Akt and decreased mTOR expression levels on human OA
chondrocytes should be considered simultaneously for the
preclinical and clinical treatment of OA.
Acknowledgments
The authors would like to thank Dr Honghai Zhao and
Dr Shaojie Wang (Zhongshan Hospital, Xiamen University) for
supplying the materials in the manuscript. The present study was
supported by the National Science Foundation of China (grant nos.
81572189 and 81371952) and the Natural Science Foundation of
Xiamen, China (grant no. 3502Z20159014).
References
|
1
|
Levine B and Kroemer G: Autophagy in the
pathogenesis of disease. Cell. 132:27–42. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Cinque L, Forrester A, Bartolomeo R,
Svelto M, Venditti R, Montefusco S, Polishchuk E, Nusco E, Rossi A,
Medina DL, et al: FGF signalling regulates bone growth through
autophagy. Nature. 528:272–275. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sasaki H, Takayama K, Matsushita T, Ishida
K, Kubo S, Matsumoto T, Fujita N, Oka S, Kurosaka M and Kuroda R:
Autophagy modulates osteoarthritis-related gene expression in human
chondrocytes. Arthritis Rheum. 64:1920–1928. 2012. View Article : Google Scholar
|
|
4
|
Zhao H, Zhang T, Xia C, Shi L, Wang S,
Zheng X, Hu T and Zhang B: Berberine ameliorates cartilage
degeneration in interleukin-1β-stimulated rat chondrocytes and in a
rat model of osteoarthritis via Akt signalling. J Cell Mol Med.
18:283–292. 2014. View Article : Google Scholar
|
|
5
|
Cheng L, Zeng G, Liu Z, Zhang B, Cui X,
Zhao H, Zheng X, Song G, Kang J and Xia C: Protein kinase B and
extracellular signal-regulated kinase contribute to the
chondroprotective effect of morroniside on osteoarthritis
chondrocytes. J Cell Mol Med. 19:1877–1886. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Zhou PH, Liu SQ and Peng H: The effect of
hyaluronic acid on IL-1beta-induced chondrocyte apoptosis in a rat
model of osteoarthritis. J Orthop Res. 26:1643–1648. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Ribeiro M, López de Figueroa P,
Nogueira-Recalde U, Centeno A, Mendes AF, Blanco FJ and Caramés B:
Diabetes-accelerated experimental osteoarthritis is prevented by
autophagy activation. Osteoarthritis Cartilage. 24:2116–2125. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Zhang Y, Vasheghani F, Li YH, Blati M,
Simeone K, Fahmi H, Lussier B, Roughley P, Lagares D, Pelletier JP,
et al: Cartilage-specific deletion of mTOR upregulates autophagy
and protects mice from osteoarthritis. Ann Rheum Dis. 74:1432–1440.
2015. View Article : Google Scholar
|
|
9
|
Cui X, Wang S, Cai H, Lin Y, Zheng X,
Zhang B and Xia C: Overexpression of microRNA-634 suppresses
survival and matrix synthesis of human osteoarthritis chondrocytes
by targeting PIK3R1. Sci Rep. 6:231172016. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Chen J, Crawford R and Xiao Y: Vertical
inhibition of the PI3K/Akt/mTOR pathway for the treatment of
osteoarthritis. J Cell Biochem. 114:245–249. 2013. View Article : Google Scholar
|
|
11
|
Baltzer AW, Moser C, Jansen SA and Krauspe
R: Autologous conditioned serum (Orthokine) is an effective
treatment for knee osteoarthritis. Osteoarthritis Cartilage.
17:152–160. 2009. View Article : Google Scholar
|
|
12
|
Moser C: Response to: cytokine profile of
autologous conditioned serum for treatment of osteoarthritis, in
vitro effects on cartilage metabolism and intra-articular levels
after injection. Arthritis Res Ther. 12:4102010. View Article : Google Scholar
|
|
13
|
Rutgers M, Saris DB, Dhert WJ and Creemers
LB: Cytokine profile of autologous conditioned serum for treatment
of osteoarthritis, in vitro effects on cartilage metabolism and
intra-articular levels after injection. Arthritis Res Ther.
12:R1142010. View
Article : Google Scholar : PubMed/NCBI
|
|
14
|
Attur MG, Dave M, Cipolletta C, Kang P,
Goldring MB, Patel IR, Abramson SB and Amin AR: Reversal of
autocrine and paracrine effects of interleukin 1 (IL-1) in human
arthritis by type II IL-1 decoy receptor. Potential for
pharmacological intervention. J Biol Chem. 275:40307–40315. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Zeng G, Cui X, Liu Z, Zhao H, Zheng X,
Zhang B and Xia C: Disruption of phosphoinositide-specific
phospholipases Cγ1 contributes to extracellular matrix synthesis of
human osteoarthritis chondrocytes. Int J Mol Sci. 15:13236–13246.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Li C, Cai S, Wang X and Jiang Z:
Identification and characterization of ANO9 in stage II and III
colorectal carcinoma. Oncotarget. 6:29324–29334. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Zheng CH and Levenston ME: Fact versus
artifact: Avoiding erroneous estimates of sulfated
glycosaminoglycan content using the dimethylmethylene blue
colorimetric assay for tissue-engineered constructs. Eur Cell
Mater. 29:224–236. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Zhang B and Xia C:
12-O-tetradecanoylphorbol-1, 3-acetate induces the negative
regulation of protein kinase B by protein kinase Calpha during
gastric cancer cell apoptosis. Cell Mol Biol Lett. 15:377–394.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Shao XX, Duncan NA, Lin L, Fu X, Zhang JY
and Yu CL: Serum-free media for articular chondrocytes in vitro
expansion. Chin Med J (Engl). 126:2523–2529. 2013.
|
|
20
|
Wolf F, Haug M, Farhadi J, Candrian C,
Martin I and Barbero A: A low percentage of autologous serum can
replace bovine serum to engineer human nasal cartilage. Eur Cell
Mater. 15:1–10. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Jain MV, Paczulla AM, Klonisch T, Dimgba
FN, Rao SB, Roberg K, Schweizer F, Lengerke C, Davoodpour P,
Palicharla VR, et al: Interconnections between apoptotic,
autophagic and necrotic pathways: Implications for cancer therapy
development. J Cell Mol Med. 17:12–29. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Laplante M and Sabatini DM: mTOR signaling
in growth control and disease. Cell. 149:274–293. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Inoki K, Li Y, Xu T and Guan KL: Rheb
GTPase is a direct target of TSC2 GAP activity and regulates mTOR
signaling. Genes Dev. 17:1829–1834. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tee AR, Manning BD, Roux PP, Cantley LC
and Blenis J: Tuberous sclerosis complex gene products, Tuberin and
Hamartin, control mTOR signaling by acting as a GTPase-activating
protein complex toward Rheb. Curr Biol. 13:1259–1268. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Bohensky J, Leshinsky S, Srinivas V and
Shapiro IM: Chondrocyte autophagy is stimulated by HIF-1 dependent
AMPK activation and mTOR suppression. Pediatr Nephrol. 25:633–642.
2010. View Article : Google Scholar :
|
|
26
|
Xue JF, Shi ZM, Zou J and Li XL:
Inhibition of PI3K/AKT/mTOR signaling pathway promotes autophagy of
articular chondrocytes and attenuates inflammatory response in rats
with osteoarthritis. Biomed Pharmacother. 89:1252–1261. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Takayama K, Kawakami Y, Kobayashi M, Greco
N, Cummins JH, Matsushita T, Kuroda R, Kurosaka M, Fu FH and Huard
J: Local intra-articular injection of rapamycin delays articular
cartilage degeneration in a murine model of osteoarthritis.
Arthritis Res Ther. 16:4822014. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Rosa SC, Rufino AT, Judas F, Tenreiro C,
Lopes MC and Mendes AF: Expression and function of the insulin
receptor in normal and osteoarthritic human chondrocytes:
Modulation of anabolic gene expression, glucose transport and
GLUT-1 content by insulin. Osteoarthritis Cartilage. 19:719–727.
2011. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Cravero JD, Carlson CS, Im HJ, Yammani RR,
Long D and Loeser RF: Increased expression of the Akt/PKB inhibitor
TRB3 in osteoarthritic chondrocytes inhibits insulin-like growth
factor 1-mediated cell survival and proteoglycan synthesis.
Arthritis Rheum. 60:492–500. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Hahn-Windgassen A, Nogueira V, Chen CC,
Skeen JE, Sonenberg N and Hay N: Akt activates the mammalian target
of rapamycin by regulating cellular ATP level and AMPK activity. J
Biol Chem. 280:32081–32089. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Peng M, Huang Y, Tao T, Peng CY, Su Q, Xu
W, Darko KO, Tao X and Yang X: Metformin and gefitinib cooperate to
inhibit bladder cancer growth via both AMPK and EGFR pathways
joining at Akt and Erk. Sci Rep. 6:286112016. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Galasso O, Panza S, Santoro M, Goldring
MB, Aquila S and Gasparini G: PTEN elevation, autophagy and
metabolisc reprogramming may be induced in human chondrocytes
during steroids or nutrient depletion and osteoarthritis. J Biol
Regul Homeost Agents. 29(Suppl 4): S1–S14. 2015.
|
|
33
|
Memmott RM and Dennis PA: Akt-dependent
and -independent mechanisms of mTOR regulation in cancer. Cell
Signal. 21:656–664. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
O'Reilly KE, Rojo F, She QB, Solit D,
Mills GB, Smith D, Lane H, Hofmann F, Hicklin DJ, Ludwig DL, et al:
mTOR inhibition induces upstream receptor tyrosine kinase signaling
and activates Akt. Cancer Res. 66:1500–1508. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yori JL, Lozada KL, Seachrist DD, Mosley
JD, Abdul-Karim FW, Booth CN, Flask CA and Keri RA: Combined
SFK/mTOR inhibition prevents rapamycin-induced feedback activation
of AKT and elicits efficient tumor regression. Cancer Res.
74:4762–4771. 2014. View Article : Google Scholar : PubMed/NCBI
|