Introduction
Human lens epithelium is the most metabolically
active cell layer of the lens, and is the initial cell layer
exposed to environmental and oxidative insult (1). Oxidative stress has an important
role in the degradation, oxidation, cross-linking and aggregation
of lens proteins, and also triggers lens epithelial cell apoptosis.
The apoptosis of lens epithelial cells has been proposed as a
common basis for the initiation of noncongenital cataract
formation, with oxidative stress as a major contributor to cataract
formation (2–4). Exposure to oxidative stress results
in lens opacification in experimental animal models (5,6)
and cultured lens systems (7–9).
Hydrogen peroxide (H2O2) is
one of the most physiologically relevant oxidants of the lens and
aqueous humor, and has been reported to deplete glutathione and
damage ion pump activity in lens epithelial cells (LECs) (10,11). Elevated levels of
H2O2 are reported in the aqueous humor of
patients with cataract and can lead to opacification of the lens
in vitro (12,13). Overall, previous studies have
demonstrated that H2O2-induced apoptosis in
human lens epithelium cells is a useful model of cataractogenesis
(14–16).
It has been confirmed that loss of transparency
during human cataract formation is involved in a variety of complex
metabolic and physiologic mechanisms, and an increase in
antioxidant levels in the lens may prevent or ameliorate oxidative
damage, and reduce cataract risk (7). Therefore, it is important to develop
protective strategies against apoptosis in human lens epithelial
cells to prevent cataractogenesis.
Chlorogenic acid (CGA) is one of the most abundant
poly-phenol compounds in coffee, strawberries, pineapple, apple,
sunflower and blueberries (17).
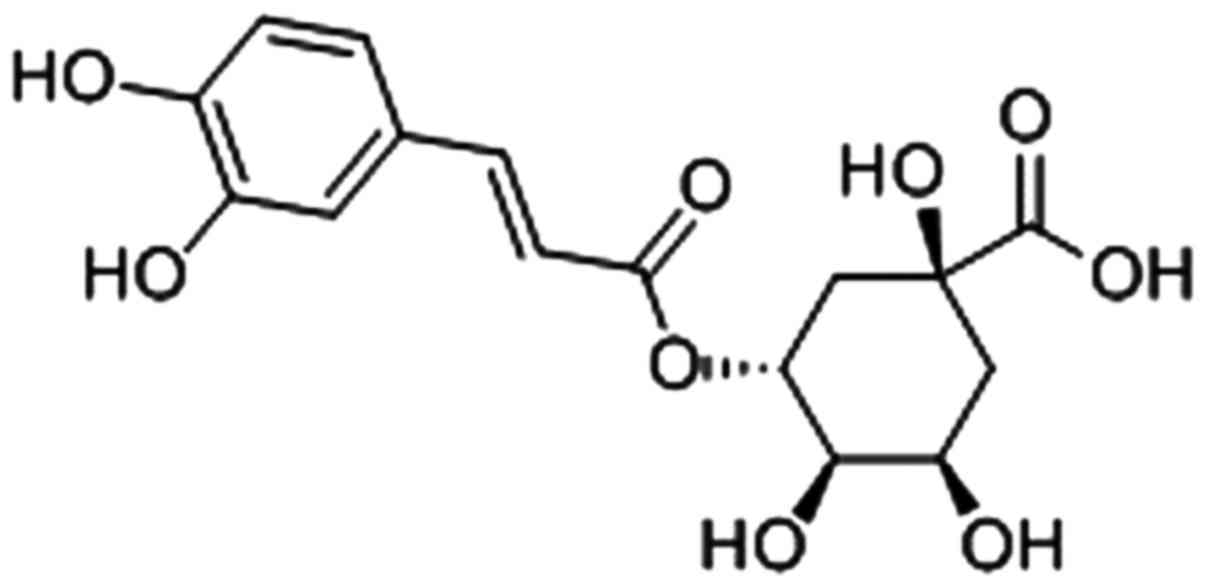
The molecular structure of CGA is presented in Fig. 1. It can exert various biological
properties and modulatory effects on lipid and glucose metabolism
under metabolic dysregulation conditions, such as antioxidant,
antiangiogenic, anticarcinogenic and antiglycation (18–21). Kim et al (22) reported that CGA may provide a
potential therapeutic approach for prevention of diabetic
complications, such as cataracts. Akila et al (23) reported that CGA is an effective
protective agent to maintain the activities of enzymic
antioxidants, including superoxide dismutase, catalase, glutathione
peroxidase and glutathione-S-transferase. Similarly, Ye et
al (24) demonstrated that
CGA efficiently protected kidney function against oxidative stress
in a rat model of diabetic nephropathy.
Based on these observations, we hypothesize that CGA
may protect human lens epithelial cells (hLECs) against oxidative
stress-induced apoptosis, and may offer benefits in the treatment
of cataract associated with oxidative stress. In the present study,
H2O2-treated hLECs and rabbit lenses were
used as models to examine the protective effect of CGA on LECs
exposed to H2O2-mediated oxidative
stress.
Materials and methods
Materials
Human HLE-B3 lens epithelial cell line was obtained
from the American Type Culture Collection (Manassas, VA, USA), and
chlorogenic acid (purity, 98.7%) was purchased from the National
Institute for the Control of Pharmaceutical and Biological Products
of China (Beijing, China). MTT and H2O2 (30%)
were purchased from Sigma-Aldrich (Merck KGaA, Darmstadt, Germany).
A 500 µM H2O2 solution was prepared in
phosphate-buffered saline (PBS) immediately prior to application.
The Annexin V/propidium iodide (PI) apoptotic detection kit was
purchased from Nanjing KeyGen Biotech Co., Ltd. (Nanjing, China).
All other chemicals used were purchased from Sigma-Aldrich (Merck
KGaA) unless otherwise stated.
Cell culture and treatment
HLE-B3 cells were cultured in RPMI-1640 medium
(HyClone; GE Healthcare Life Sciences, Logan, UT, USA) containing 1
g/l glucose, 10% fetal bovine serum (HyClone; GE Healthcare Life
Sciences), 100 U/ml penicillin and 100 µg/ml streptomycin
(HyClone; GE Healthcare Life Sciences) under a humidified
atmosphere with 5% CO2 at 37°C. The cells were seeded
into a 60 mm culture dish (Corning Incorporated, Corning, NY, USA).
When at 75–80% confluence, the cells were treated with
H2O2 (10–500 µM) for 24 h or
pretreated with CGA for 2 h prior to
H2O2-treatment. At the indicated time-points,
the cells were collected for different assays.
Cell viability assay
The concentrations of CGA and
H2O2 were optimized using MTT assay. Briefly,
hLECs were cultured in 96-well plates and treated with a broad
range of concentrations for each reagent
(H2O2 or CGA) for 24 h. To examine the effect
of CGA, hLECs were incubated with 100 µM
H2O2 in the absence or presence of different
concentrations of CGA for 24 h. Subsequently, 20 µl MTT
solution (5 mg/ml) was added into each well. After another 4 h
incubation at 37°C, the medium was removed and the formazan
crystals formed by oxidation of the MTT dye were dissolved with 150
µl DMSO. The absorbance was measured at 490 nm using the
spectrophotometer [Unico (shanghai) Science Instruments Co., Ltd.,
Shanghai, China]and the cell survival ratio was expressed as a
percentage of the blank.
Measurement of intracellular reactive
oxygen species (ROS)
To obtain further evidence for the protective effect
of CGA against H2O2 induced oxidative stress,
alterations of intracellular ROS levels were determined. The
production of intracellular ROS was measured using
2′,7′-dichlorofluorescin diacetate (DCFH-DA; Invitrogen, Carlsbad,
CA, USA) by flow cytometry. Briefly, hLECs were incubated either
with 100 µM H2O2 alone or treated with
different concentrations (10, 30 and 50 µM) of CGA for 2 h
prior to treatment with 100 µM H2O2
for 24 h. After harvest of hLECs, the cells were incubated with
DCFH-DA solution (10 µM) in the dark at 37°C for 30 min,
washed with PBS (pH 7.4), and analyzed within 30 min using a flow
cytometer (Accuri C6; Accuri Cytometers Inc., Ann Arbor, MI, USA).
The specific fluorescence signals that correspond to DCFH-DA were
collected with a 525 nm band pass filter. For each determination,
2.0×104 cells were counted.
Apoptosis assa
For quantification of the apoptosis rate in hLECs,
cells were cultured on a 6-well plate at 5.0×105
cells/well and treated with 100 µM
H2O2 with or without CGA (0, 10, 30 and 50
µM) for 24 h. Subsequently, hLECs were collected and stained
using Annexin V/PI kit and assessed by a flow cytometer (Accuri C6;
Accuri Cytometers), following the instructions of the
manufacturer.
Revere transcription-quantitative
polymerase chain reaction (RT-qPCR)
The effect of CGA on the gene expression of BCL2
associated X, apoptosis regulator (Bax) and BCL2, apoptosis
regulator (Bcl-2) mRNA using RT-qPCR in the presence and absence of
H2O2. The cells (6×105) were
incubated either with 100 µM H2O2 for
24 h or with CGA for 2 h prior to treatment with 100 µM
H2O2. Total RNA was extracted with TRIzol
reagent (Life Technologies; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) and cDNA was generated using a Superscript cDNA
kit (Thermo Fisher Scientific, Inc.) based on the manufacturer's
protocol. Following quantification with a microspectrophotometer
(Beijing Kaiao Technology Development Co., Ltd., Beijing, China),
cDNA was synthesized using total RNA. qPCR was performed with
SYBR-Green Master Mix (Takara Biotechnology Co., Ltd., Dalian,
China) in a Stratagene Mx3000P sequence detection system (Agilent
Technologies, Inc., Santa Clara, CA, USA). GAPDH was used as a
positive control, and a negative control without template RNA was
also included. The primer sequences are presented in Table I. The reactions were performed in
a total volume of 20 µl using the SensiMix One-Step kit
(http://www.quantace.com; Quantace, Finchley, UK).
The conditions of PCR amplification for Bcl-2 and Bax were as
follows: 95°C for 10 min, followed by 40 cycles of a 95°C
denaturation for 15 sec, annealing at 55°C for 30 sec, and 72°C
extension for 50 sec. Each experiment was carried out four times
and the ΔΔCq values were calculated by normalizing the gene
expression levels to the expression of GAPDH (25). The relative expression level of
each gene was expressed as a fold change.
 | Table IPrimer sequences for Bcl-2, Bax and
GAPDH. |
Table I
Primer sequences for Bcl-2, Bax and
GAPDH.
| Target gene | Primer
sequence |
|---|
| Bcl-2 | F:
5′-GAGTGGATGACCGTCTACCTG-3′ |
| R:
5′-CCTGAGACCTTCTGCTTTCG-3′ |
| Bax | F:
5′-TTTTGCTTCAGGGTTTCATC-3′ |
| R:
5′-GACACTCGCTCAGCTTCTTG-3′ |
| GAPDH | F:
5′-CCATGTTCGTCATGGGTG TGAACCA-3′ |
| R:
5-GCCAGTAGAGGCAGGGATGATGTTC-3′ |
ELISA
The levels of Bcl-2/Bax (anti-/pro-apoptotic)
proteins were determined using commercially available ELISA kits
[BCL-2 kit (JYM0302Hu), BAX kit (JYM0265Hu); Colorfulgene
Biological Technology, Co., Ltd., Wuhan, China]. The hLECs
(6×105) were incubated with 100 µM
H2O2 for 24 h alone or treated with CGA for 2
h prior to treatment with 100 µM H2O2
for 24 h. The cells were then collected and cell concentration was
diluted to 106/ml with PBS (pH 7.2–7.4). Following three
freeze-thaw cycles, damaged cells were centrifuged at 5,000 × g at
4°C for 20 min. The supernatant was carefully collected and stored
at −80°C prior to use. The protein in the cell lysate was
determined using the ELISA kit according to the manufacturer's
instructions. Subsequently, the plates were read at 450 nm using a
microplate reader (BioTek ELX800; BioTek Instruments, Inc.,
Winooski, VT, USA). The protein level of each sample was determined
by comparison to a standard curve.
Lens organ culture and treatment
All animal procedures were in accordance with the
Association for Research in Vision and Ophthalmology Statement for
the Use of Animals in Ophthalmic and Vision Research (26), and the animal experiments were
approved by Shandong University of Traditional Chinese Medicine
Animal Care and Ethics Committee (Jinan, China). In the present
study, New Zealand White rabbits (n=30; weighing 1.8–2.0 kg) aged
10–12 weeks were euthanized with an overdose of sodium
pentobarbital injection through the marginal ear vein. The eyes
were removed and the lenses were carefully dissected by a posterior
approach. Lenses were immediately transferred into a 24-well
culture plate containing 2 ml Dulbecco's modified Eagle's medium
(HyClone, Beijing, China), 1 g/l glucose, 100 U/ml penicillin and
100 µg/ml streptomycin (HyClone; GE Healthcare Life
Sciences). Approximately, 24 h after the preparation of organ
cultures, transparent lenses were selected for further
experimentation. During the experiment, H2O2
and CGA were maintained at indicated concentrations for a period of
12 h, and the medium was changed. Conditioned medium was stored for
further culture. Lenses were cultured in a 5% CO2
incubator at 37°C for 48 h, and were images using a
stereomicroscope under a cross background (1.0×1.0 cm). Each sample
contained three lenses and lens opacity was analyzed using
ImageJ-1.46 software (National Institutes of Health, Bethesda, MD,
USA). This experiment was repeated three times independently.
Flow cytometry with Annexin V/PI
staining
The apoptosis rate of lens epithelial cells was
assessed by flow cytometry with Annexin V/PI staining. The lenses
exposed to 500 µM H2O2 were cultured
with various concentrations (i.e., 0, 10, 30 and 50 µM) of
CGA for 48 h. At the indicated time-points (0, 12, 24 and 48 h,
respectively), lens epithelial explants were carefully detached
from rabbit lens under an operation microscope (YZ20P5; 66 Vision
Tech Co., Ltd., Suzhou, China). Lens epithelial explants were then
sheared, digested with trypsin, and RPMI-1640 medium was added to
terminate the digestion (27).
The cell suspension was then passed through a cell strainer and
centrifuged (300 × g) at 4°C for 5 min. The cells were resuspended
in RPMI-1640 medium and collected by centrifugation at 300 × g at
4°C for 10 min. The collected cells were washed with cold PBS and
stained with Annexin V/PI apoptotic detection kit. Finally, the
cells were resuspended in 500 µl PBS for further analysis
using a flow cytometer (Accuri C6; Accuri Cytometers, Inc.).
Statistical analysis
All experiments were repeated three times and data
are expressed as the mean ± standard deviation, and were analyzed
by one-way analysis of variance, followed by Tukey's honest
significant difference post hoc test, using SPSS software (version
17.0; SPSS Inc., Chicago, IL, USA). P<0.05 was considered to
indicate a statistically significant difference.
Results
Cell viability assessed by MTT assay
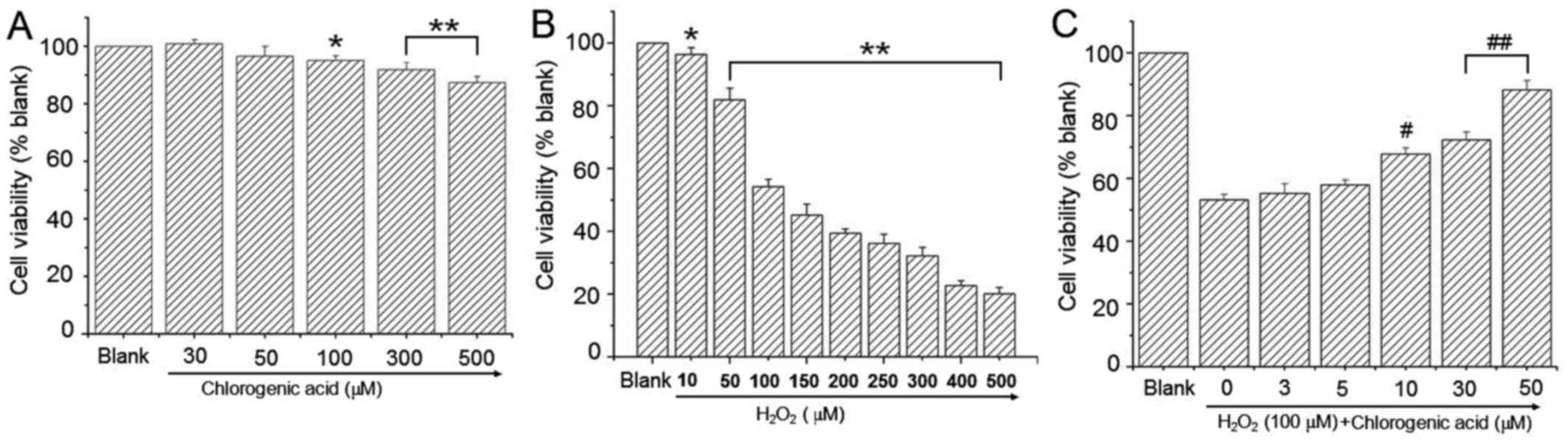
To determine the effect of CGA on hLECs, cells were
treated with a broad range of CGA concentrations for 24 h. Cell
viability was presented as a percentage of the blank value. The
results indicated that CGA had no cytotoxicity in hLEC cells when
the CGA concentration was <100 µM (Fig. 2A). H2O2-treated cells exhibited
lower cell viability at H2O2 concentrations ≥50 µM (Fig. 2B). Co-treatment with CGA resulted
in a dose-dependent reduction in cytotoxicity induced by H2O2 (100
µM; Fig. 2C).
Effect of CGA on intracellular ROS
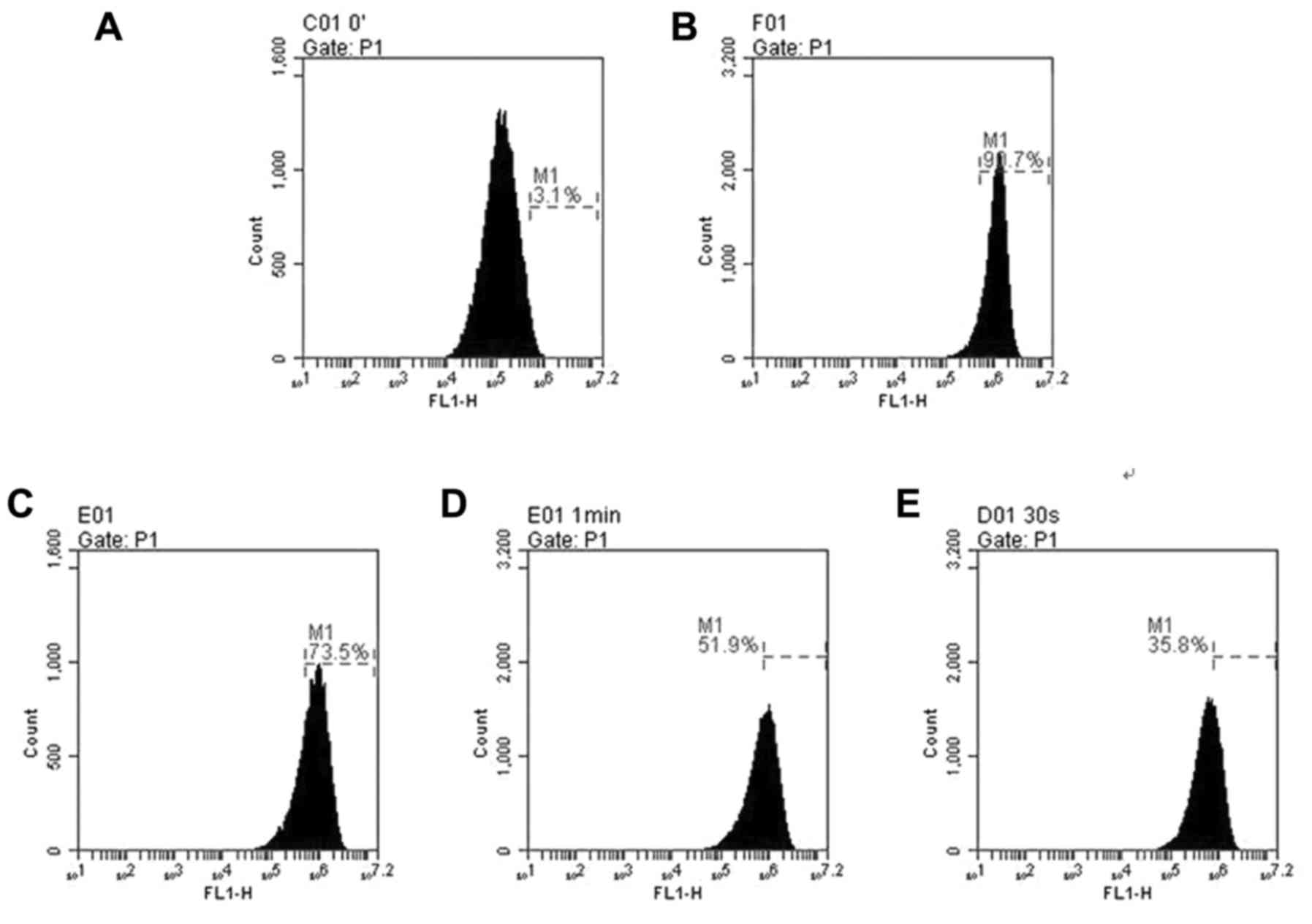
hLECs treatment with 100 µM H2O2 alone for 24
h resulted in the production of ROS with a ~3-fold increase
compared with 50 µM CGA-treated cells (Fig. 3). Pretreatment with CGA prior to
H2O2 exposure markedly reduced the ROS levels. When hLECs were
treated with CGA (0, 10, 30 and 50 µM) for 2 h prior to
treatment with 100 µM H2O2 for 24 h,
the ROS generation reduced from 90.7±7.75 to 73.5±5.98%, 51.9±4.74
and 35.8±3.53% (Fig. 3B–E),
respectively. These findings demonstrated that with the increase of
concentrations of CGA, the intracellular ROS level induced by
H2O2 was reduced and the ROS reduction was in
a concentration-dependent manner. In addition, there was a
significant difference in the level of
H2O2-induced ROS compared with that of
untreated cells (Fig. 3).
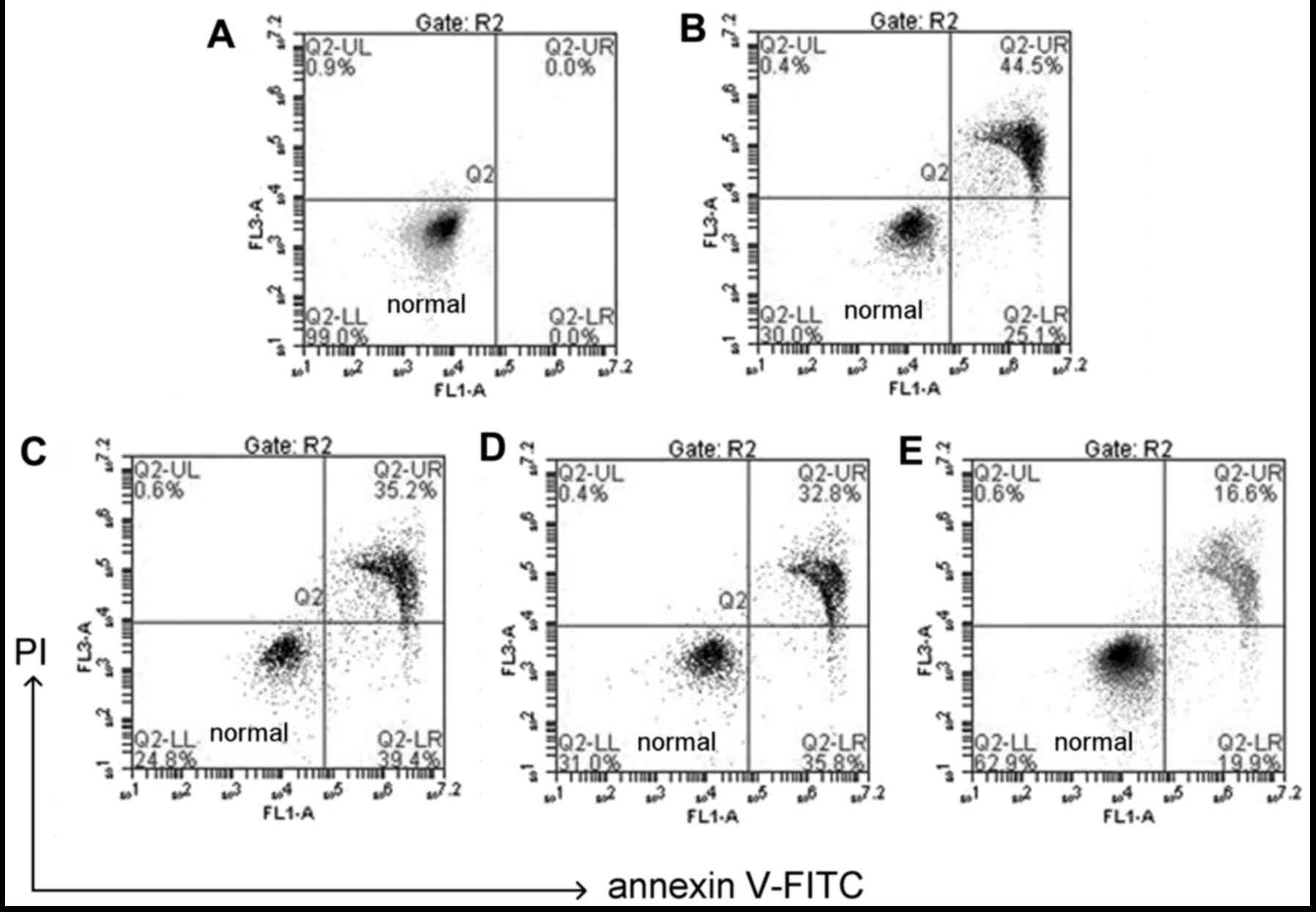
CGA prevents
H2O2-induced apoptotic changes in hLECs
The use of Annexin V/PI double staining allows
distinction between live cell populations, cells entering early
apoptosis and those in late-stage apoptosis/necrosis. Q2-LL
quadrant indicates healthy cells, Q2-UR quadrant indicates necrosis
cells (Fig. 4). Any late-stage
apoptosis cells were considered as necrosis as this technique is
not sensitive enough to differentiate between the two. The data in
Fig. 4 demonstrated that
following treatment with 100 µM H2O2
and co-treatment with CGA (10, 30 and 50 µM) for 24 h, the
late apoptotic rate of HLE-B3 cells decreased from 44.5 to 35.2,
32.8 and 16.6%, respectively (Fig.
4).
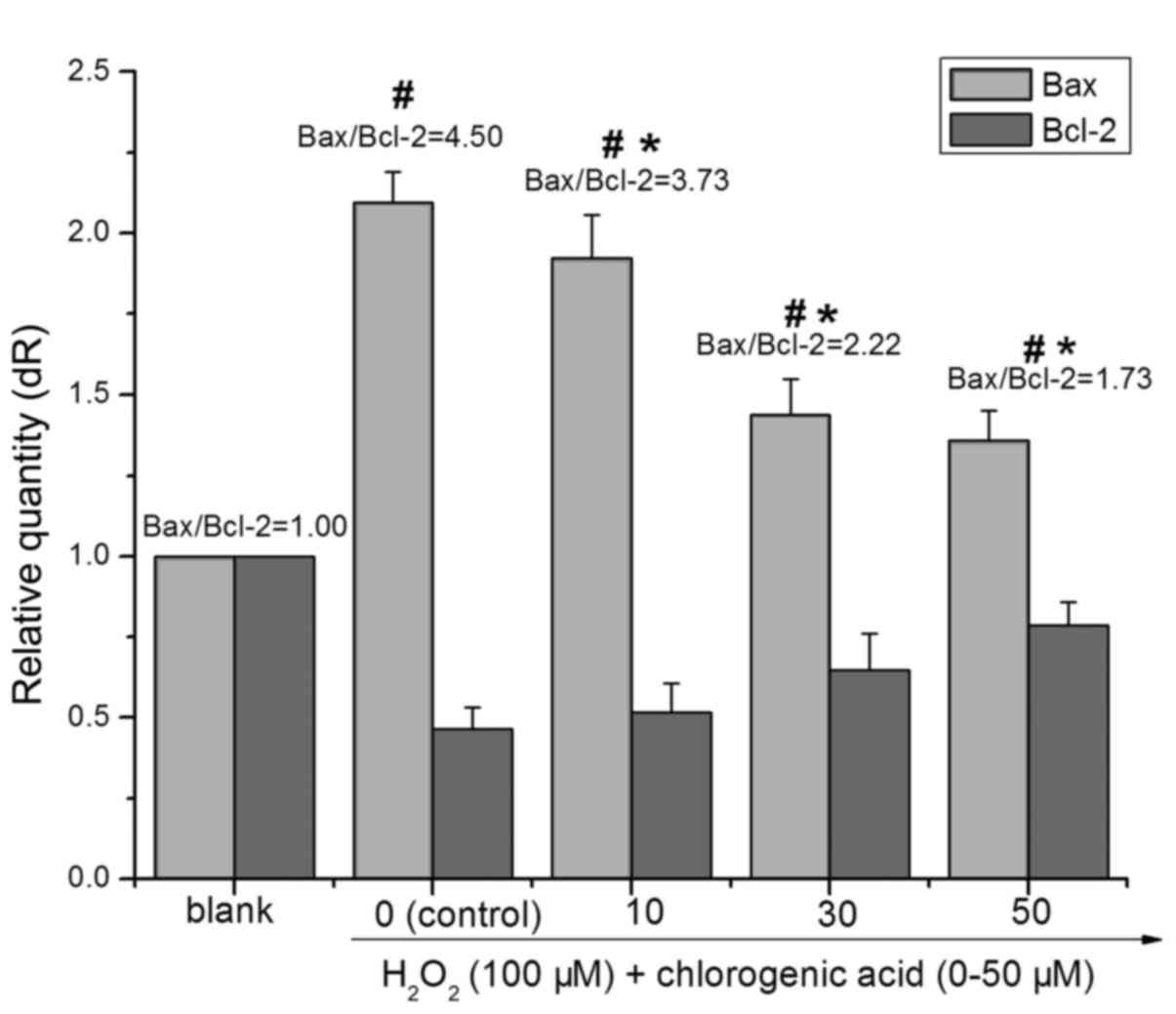
CGA modulates the expression of
Bcl-2/Bax
The members of the Bcl-2 protein family are pivotal
role in the regulation of the mitochondrial apoptotic pathway
(28). Both pro-apoptotic and
anti-apoptotic Bcl-2 family members can affect the execution of
apoptosis. As presented in Fig.
5, treatment with 100 µM H2O2 for
24 h decreased the expression of Bcl-2 (0.47-fold higher compared
with blank) and increased the expression of Bax (2.09-fold higher
compared with blank), whereas pretreatment with CGA inhibited the
downregulation of Bcl-2 and upregulation of Bax. The ratio of
Bax/Bcl-2 in the 100 µM H2O2-treated
group was significantly higher than that in the blank group
(P<0.01). With elevated CGA concentrations, the ratio of
Bax/Bcl-2 in the CGA-treated (i.e., 10, 30 and 50 µM) groups
decreased significantly compared with the
H2O2-treated group (P<0.01). Following
treatment with 100 µM H2O2 and
co-treatment with different concentrations of CGA (10, 30 and 50
µM) for 24 h the ratios of Bax/Bcl-2 were 4.50±0.61,
3.73±0.46, 2.22±0.47 and 1.73±0.33, respectively.
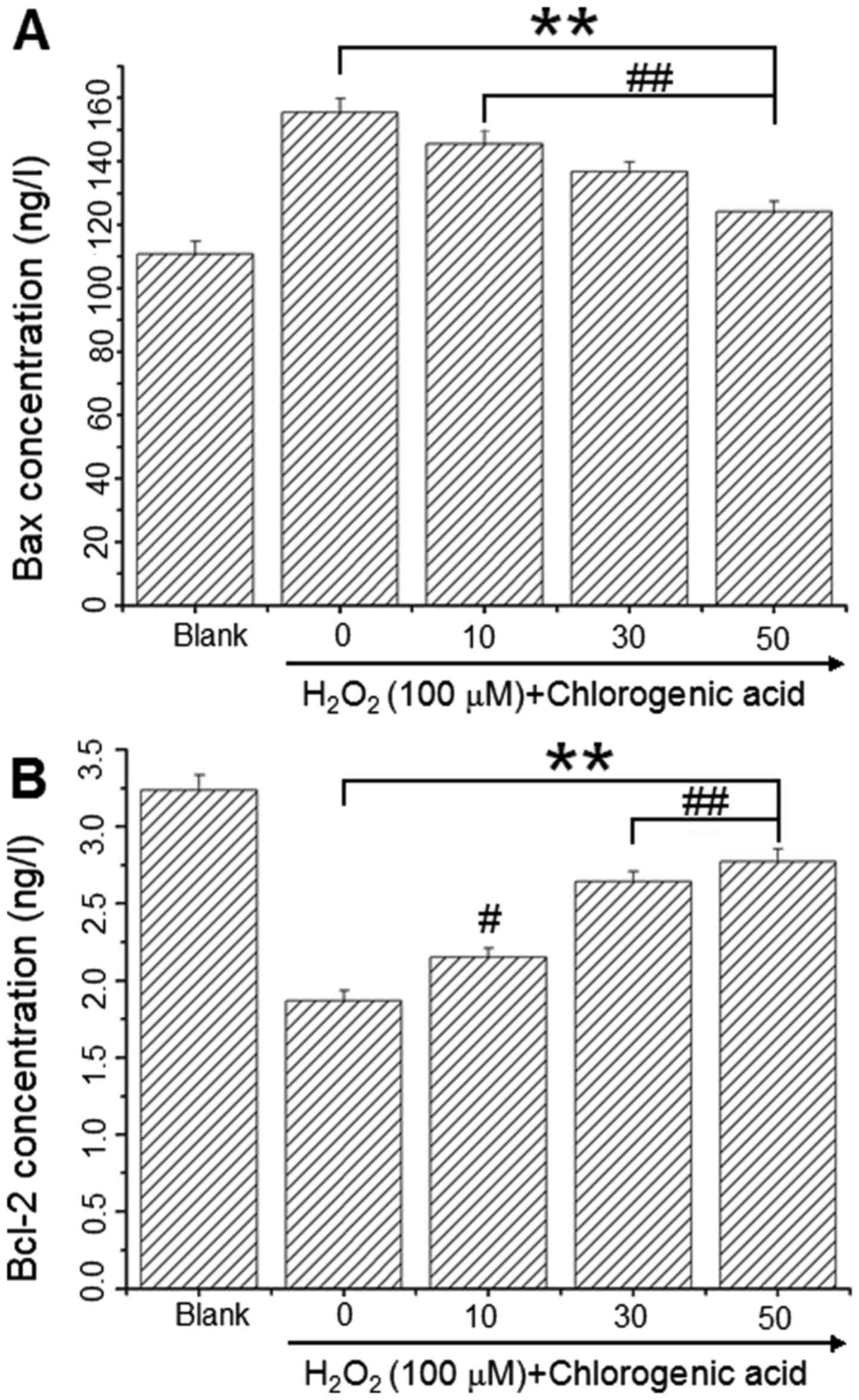
CGA downregulates Bcl-2 expression and
upregulates Bax expression
Bcl-2 and Bax levels were assessed using
commercially available ELISA kits to determine whether these key
regulators of apoptosis were involved in the
H2O2-induced apoptosis mechanism. As
presented in Fig. 5, Bax protein
levels were 110.91±3.97, 155.49±4.30, 145.59±4.17, 136.78±3.07 and
124.19±3.11 ng/l (Fig. 6A) and
Bcl-2 levels were 3.24±0.10, 1.87±0.07, 2.15±0.06, 2.64±0.09 and
2.77±0.10 ng/l (Fig. 6B) in the
blank, 100 µM H2O2 and
H2O2 + CGA 10, 30 and 50 µM groups,
respectively. These results demonstrated that CGA increased Bcl-2
protein expression and decreased Bax protein expression in hLECs
following incubation with 100 µM H2O2
for 24 h.
CGA prevents
H2O2-induced lens opacity
The protective effects of CGA against
cataractogenesis of rabbit lenses induced with
H2O2 (500 µM) were investigated
further. As presented in Fig. 7,
loss of transparency in lenses exposed to 500 µM
H2O2 was first noted in the equatorial
region, spreading throughout the superficial cortex by 24 h and
into deeper regions by 48 h. By contrast, there was little change
in transparency of untreated lenses during the entire exposure
period. The opacities of rabbit lenses incubated in
H2O2 (500 µM) media containing various
concentrations of CGA were measured every 12 h and compared with
the control and blank samples. The opacities of the lenses began to
increase after 12 h treatment with H2O2, and
were gradually improved by CGA treatment in a dose-dependent
manner.
At the indicated time-point, lens epithelial
explants were detached. LECs were harvested and stained with
Annexin V/PI for further analysis using a flow cytometer. Lenses
treated with 500 µM H2O2 for 12, 24
and 48 h, caused a time-dependent increase of apoptosis rates
(51.8±3.81, 77.2±4.12 and 94.57±4.77% of control value,
respectively) whereas co-treatment with different concentrations of
CGA [10 µM (III), 30 µM (IV), 50 µM (V)] the
decreased the apoptosis rates (Fig.
8).
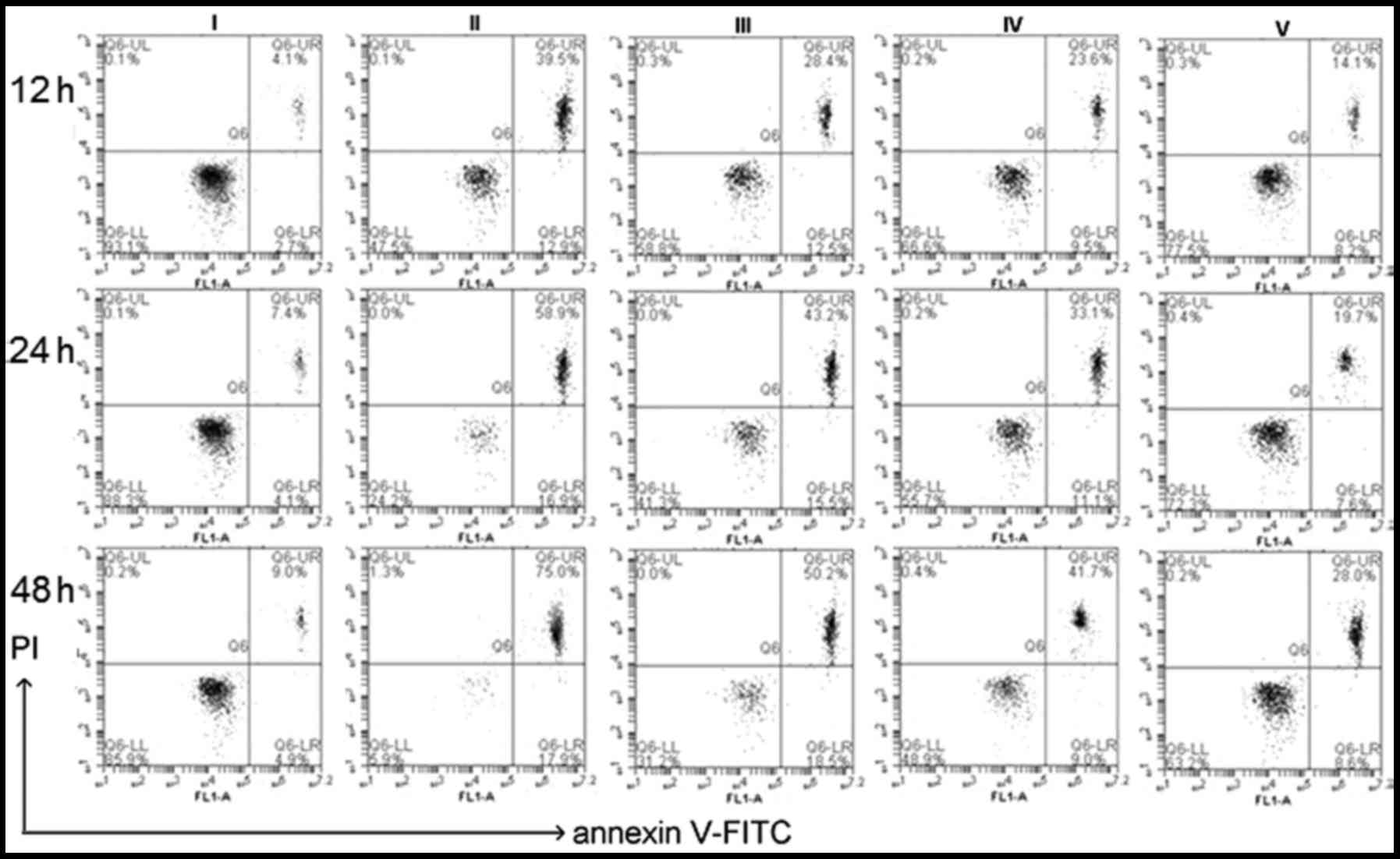 | Figure 8Apoptotic analysis of isolated lens
epithelial cells by flow cytometry after staining with Annexin
V/PI. Cells were treated untreated (blank; I) or treated with 500
µM H2O2 (control) plus different
concentrations of chlorogenic acid [0 µM (II), 10 µM
(III), 30 µM (IV), 50 µM (V)]. At the indicated
time-points (i.e., 12, 24 and 48 h), the apoptosis rates were (II):
51.8±3.81, 77.2±4.12 and 94.57±4.77%; (III): 42.3±3.76, 58.3±3.67
and 65.9± 4.12%; (IV): 33.7±2.95, 42.3±3.29 and 50.43±4.13%; (V):
22.4±1.09, 28.5±2.54 and 39.2±3.71% of control value. Data are
expressed as the mean ± standard deviation. P<0.01 blank vs. II,
III, IV, V; P<0.01 control vs. I, III, IV, V; one-way analysis
of variance and followed by Tukey's honest significant difference
post hoc test). PI, propidium iodide; FITC, fluorescein
isothiocyanate. |
Discussion
As the most anterior part of the lens, LECs are the
primary site of external insult that ultimately leads to cataracts
(29). LECs are also the most
metabolically-active part of the lens and are responsible for
maintaining homeostasis and transparency. Li et al (3) have reported that apoptosis in lens
epithelium may be a common cellular basis for noncongenital
cataract formation, and that blocking apoptosis may prevent
cataract formation. To investigate the protective effects of CGA on
H2O2-induced apoptosis in hLECs, we performed
a dose-response experiment using a range of concentrations (10, 30
and 50 µM). H2O2 was used as the
oxidant model of classical oxidative stress. The results
demonstrated that all concentrations of CGA exerted a protective
effect against oxidative stress and an inhibitory effect against
apoptosis, as measured by the cell morphology and cell viability
studies. In addition, CGA inhibited cytotoxicity in hLECs caused by
100 µM H2O2 compared with
H2O2 treatment alone. Apoptosis is an
intracellular suicide mechanism that cause morphologic changes and
biochemical responses. In the presence of CGA, the proportions of
apoptotic cells were significantly decreased. Thus, it is suggested
that CGA may have a potentially beneficial role in the prevention
of cataract formation.
It is well established that the proto-oncogene Bcl-2
can prevent apoptosis induced by a variety of factors. Regarding
the mechanism by which Bcl-2 prevents cell death, one theory
suggests that it acts by protecting cells from oxidative stress.
Mao et al (30) reported
that through downregulation of the αB-crystallin gene, Bcl-2
reduces the tolerance of rabbit lens epithelial cells against
H2O2-induced apoptosis. Bcl-2 and Bax
proteins are widely regarded as the most important apoptotic
regulators, and their relative levels determine the fate of cells.
Bcl-2 protein expression in the mitochondrial outer membrane
inhibits cytochrome translocation into the cytosol, which is a
critical step in the apoptotic process. By contrast, Bax is a
pro-apoptotic antagonist of Bcl-2, and has been characterized as a
Bcl-2 binding protein that shares significant sequence homology
with Bcl-2 (31). An altered
ratio of anti-apoptotic to pro-apoptotic Bcl-2 family genes is
critical in determining whether apoptosis is performed. In the
current study, RT-qPCR analysis revealed that the Bax/ Bcl-2 ratio
was significantly increased by the treatment with 100 µM
H2O2, and this increase was inhibited by
pretreatment with CGA (10, 30 and 50 µM). This result
indicates that the Bcl-2 family may have a critical role in
regulating hLEC death induced by H2O2, and
that CGA is able to protect against
H2O2-stimulated apoptosis through modulation
of Bax/Bcl-2 expression.
Rabbit lenses were cultured ex vivo in an
attempt to mimic the potential in vivo pathological
environment present in patients with cataracts (12). Superoxide damages the lens,
leading to loss of transparency and the formation of a cataract
in vivo and ex vivo. Charakidas et al
(32) suggested that the
accumulation of small-scale epithelial losses during a lifetime may
induce alterations in lens fiber formation and homeostasis,
resulting in loss of lens transparency. Therefore, it is important
to develop protective strategies for apoptosis of hLECs. As
demonstrated in the current study, damage to lenses exposed to
H2O2 ex vivo was initially observed in
the equatorial region, spreading throughout the superficial cortex
by 24 h and into the deeper regions by 48 h. The
H2O2-induced opacity of lenses was improved
following treatment with CGA. Evidence from these ex vivo
experiments indicated that the observed lens opacity can be induced
by oxidative stress, and can be ameliorated by CGA.
In conclusion, the present study demonstrates that
H2O2 can induce human lens epithelial cell
apoptosis and lens opacification, whereas CGA can effectively
attenuate human lens epithelial cell apoptosis and lens opacity
under oxidative stress mediated by H2O2. CGA,
a potent antioxidant, can effectively protect HLE-B3 cells against
H2O2-induced oxidative stress and apoptosis
via Bcl-2/Bax signaling pathway, suggesting that CGA may be applied
clinically as a potential protective treatment for cataract
formation.
Acknowledgments
This study was supported by the National Natural
Science Foundation of China (grant nos. 81403438 and 81001577), and
the Natural Science Foundation of Shandong province (grant nos.
ZR2014HP059 and ZR2014HL048).
References
|
1
|
Long AC, Colitz CM and Bomser JA:
Apoptotic and necrotic mechanisms of stress-induced human lens
epithelial cell death. Exp Biol Med (Maywood. 229:1072–1080. 2004.
View Article : Google Scholar
|
|
2
|
Kasai H, Fukada S, Yamaizumi Z, Sugie S
and Mori H: Action of chlorogenic acid in vegetables and fruits as
an inhibitor of 8-hydroxydeoxyguanosine formation in vitro and in a
rat carcinogenesis model. Food Chem Toxicol. 38:467–471. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Li SY, Chang CQ, Ma FY and Yu CL:
Modulating effects of chlorogenic acid on lipids and glucose
metabolism and expression of hepatic peroxisome
proliferator-activated receptor-alpha in golden hamsters fed on
high fat diet. Biomed Environ Sci. 22:122–129. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Surveswaran S, Cai YZ, Xing J, Corke H and
Sun M: Antioxidant properties and principal phenolic phytochemicals
of Indian medicinal plants from Asclepia and Periplocoideaedeae.
Nat Prod Res. 24:206–221. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Truscott RJ: Age-related nuclear
cataract-oxidation is the key. Exp Eye Res. 80:709–725. 2005.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Varma SD, Devamanoharan PS and Morris SM:
Prevention of cataracts by nutritional and metabolic antioxidants.
Crit Rev Food Sci Nutr. 35:111–129. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Gupta SK, Trivedi D, Srivastava S, Joshi
S, Halder N and Verma SD: Lycopene attenuates oxidative stress
induced experimental cataract development: An in vitro and in vivo
study. Nutrition. 19:794–799. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Spector A: Oxidative stress-induced
cataract: Mechanism of action. FASEB J. 9:1173–1182.
1995.PubMed/NCBI
|
|
9
|
Spector A, Wang GM, Wang RR, Li WC and
Kleiman NJ: A brief photochemically induced oxidative insult causes
irreversible lens damage and cataract. II. Mechanism of action. Exp
Eye Res. 60:483–493. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Csukas S, Costarides A, Riley MV and Green
K: Hydrogen peroxide in the rabbit anterior chamber: Effects on
glutathione, and catalase effects on peroxide kinetics. Curr Eye
Res. 6:1395–1402. 1987. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Spector A, Ma W, Sun F, Li D and Kleiman
NJ: The effect of H2O2 and tertiary butyl hydroperoxide upon a
murine immortal lens epithelial cell line, alpha TN41. Exp Eye Res.
75:573–582. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Spector A and Garner WH: Hydrogen peroxide
and human cataract. Exp Eye Res. 33:673–681. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Reddan JR, Giblin FJ, Dziedzic DC,
Wirebaugh BM and Peters JL: Hydrogen peroxide affects specific
epithelial subpopulations in cultured rabbit lenses. Invest
Ophthalmol Vis Sci. 36:289–299. 1995.PubMed/NCBI
|
|
14
|
Jia Z, Song Z, Zhao Y, Wang X and Liu P:
Grape seed proanthocyanidin extract protects human lens epithelial
cells from oxidative stress via reducing NF-кB and MAPK protein
expression. Mol Vis. 17:210–217. 2011.PubMed/NCBI
|
|
15
|
Tang X, Yao K, Zhang L, Yang Y and Yao H:
Honokiol inhibits H(2)O(2)-induced apoptosis in human lens
epithelial cells via inhibition of the mitogen-activated protein
kinase and Akt pathways. Eur J Pharmacol. 650:72–78. 2011.
View Article : Google Scholar
|
|
16
|
Yang J, Cai L, Zhang S, Zhu X, Zhou P and
Lu Y: Silica-based cerium (III) chloride nanoparticles prevent the
fructose-induced glycation of α-crystallin and
H2O2-induced oxidative stress in human lens
epithelial cells. Arch Pharm Res. 37:404–411. 2014. View Article : Google Scholar
|
|
17
|
Gugliucci A and Bastos DH: Chlorogenic
acid protects paraoxonase 1 activity in high density lipoprotein
from inactivation caused by physiological concentrations of
hypochlorite. Fitoterapia. 80:138–142. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Gugliucci A, Bastos DH, Schulze J and
Souza MF: Caffeic and chlorogenic acids in Ilex paraguariensis
extracts are the main inhibitors of AGE generation by methylglyoxal
in model proteins. Fitoterapia. 80:339–344. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kim C, Yu HG and Sohn J: The
anti-angiogenic effect of chlorogenic acid on choroidal
neovascularization. Korean J Ophthalmol. 24:163–168. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kuszak Li WC, Dunn JR, Wang K, Ma RR, Wang
W, Spector GM, Leib A, Cotliar M, Weiss AMM, et al: Lens epithelial
cell apoptosis appears to be a common cellular basis for
non-congenital cataract development in humans and animals. J Cell
Biol. 130:169–181. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Spector A, Wang GM, Wang RR, Li WC and
Kuszak JR: A brief photochemically induced oxidative insult causes
irreversible lens damage and cataract. I. Transparency and
epithelial cell layer. Exp Eye Res. 60:471–481. 1995c. View Article : Google Scholar
|
|
22
|
Kim YS, Kim NH, Lee YM and Kim JS:
Preventive effect of chlorogenic acid on lens opacity and
cytotoxicity in human lens epithelial cells. Biol Pharm Bull.
34:925–928. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Akila P and Vennila L: Chlorogenic acid a
dietary polyphenol attenuates isoproterenol induced myocardial
oxidative stress in rat myocardium: An in vivo study. Biomed
Pharmacother. 84:208–214. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Ye HY, Li ZY, Zheng Y, Chen Y, Zhou ZH and
Jin J: The attenuation of chlorogenic acid on oxidative stress for
renal injury in streptozotocin-induced diabetic nephropathy rats.
Arch Pharm Res. 39:989–997. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
26
|
Guide for Care and Use of Laboratory
Animals: NIH Publication No. 80-23. Washington, D.C.: National
Academy Press; 1996
|
|
27
|
Koushan SS, Behzad B, Masoumeh M, Vahid K
and Fatemeh ZS: Studies on the cytotoxic activities of Punica
granatum L. var. spinosa (apple punice) extract on prostate cell
line by induction of apoptosis. ISRN Pharm. 2012:1–6. 2012.
|
|
28
|
Burlacu A: Regulation of apoptosis by
Bcl-2 family proteins. J Cell Mol Med. 7:249–257. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wu ZM, Yin XX, Ji L, Gao YY, Pan YM, Lu Q
and Wang JY: Ginkgo biloba extract prevents against apoptosis
induced by high glucose in human lens epithelial cells. Acta
Pharmacol Sin. 29:1042–1050. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Mao YW, Xiang H, Wang J, Korsmeyer S,
Reddan J and Li DW: Human bcl-2 gene attenuates the ability of
rabbit lens epithelial cells against
H2O2-induced apoptosis through
down-regulation of the α B-crystallin gene. J Biol Chem.
276:43435–43445. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Soluk Tekkeşın M, Mutlu S and Olgaç V:
Expressions of bax, bcl-2 and Ki-67 in odontogenic keratocysts
(Keratocystic Odontogenic Tumor) in comparison with ameloblastomas
and radicular cysts. Turk Patoloji Derg. 28:49–55. 2012.
|
|
32
|
Charakidas A, Kalogeraki A, Tsilimbaris M,
Koukoulomatis P, Brouzas D and Delides G: Lens epithelial apoptosis
and cell proliferation in human age-related cortical cataract. Eur
J Ophthalmol. 15:213–220. 2005.PubMed/NCBI
|