Introduction
Gastric carcinoma is associated with the highest
morbidity rates worldwide and is considered a lethal cancer; the
mortalicy rate in China due to gastric carcinoma accounts for 40%
of the mortality cases of gastric carcinoma worldwide (1). Traditional radical surgery and
chemotherapy can extend the 5-year survival rate of gastric
carcinoma patients to approximately 35%; however, this rate for
patients with advanced-stage disease is only approximately 15%
(2). Therefore, compared to
traditional therapies, the targeted therapy of cancer may enhance
the treatment efficacy of cancers, including gastric carcinoma to
an extremely promising stage. Recently, the exploration of novel
specific cancer biological targets and markers with high efficiency
has become an international hotspot in the targeted therapy of
cancer.
MicroRNAs (miRNAs or miRs) are endogenous non-coding
micromolecular RNAs with approximately 21–25 nucleotides in length
with highly conserved sequences in evolution. miRNAs can regulate
gene transcription by binding to the non-coding domains of target
mRNAs and then degrading the target genes or inhibiting target gene
expression. It has been indicated that miRNAs have potent
biological functions and are able to regulate approximately30% of
mRNAs (3,4). Recent studies have revealed that
miRNAs play important roles in the genesis and development of
gastric carcinoma, and they are also therapeutic targets in gastric
carcinoma (3–5). As a type of miRNAs associated with
cancer, the miR-200 family can block the development and metastasis
of cancer by suppressing cancer neovascularization. The members of
the miR-200 family include miR-200a, miR-200b, miR-200c, miR-141
and miR-429. However, the expression levels of the miR-200 family
members in various tissues differ and their functions and
regulatory mechanisms are diverse as well (6,7).
Moreover, the miR-200 family members have been found to be low
expressed at low levels in gastric carcinoma tissues and to be was
closely associated with a poor prognosis and unfavorable clinical
parameters of patients, which renders the miR-200 family members
effective biomarkers for gastric carcinoma diagnosis and prognosis
(8). However, the specific
functions and mechanisms of action of the miR-200 family members in
gastric carcinoma remain unclear. Therefore, in this study,
miR-200c, a member of the miR-200 family, was employed as the study
object. We investigated the expression of miR-200c in gastric
carcinoma cell lines and examined its regulatory effects and
molecular mechanisms of action as regards the proliferation,
apoptosis and invasion of gastric carcinoma cells.
Materials and methods
Materials and cell culture
The BCA kit, Hoechst staining kit and HRP-labeled
goat anti-rabbit IgG (H+L) (A0208) were obtained from Beyotime
Institute of Biotechnology (Jiangsu, China). Luciferase assay
reagent (LAR) and passive lysis buffer (PLB) were from Promega
(Madison, WI, USA). Methyl thiazolyl tetrazolium (MTT) was
purchased from Gibco (Grand Island, NY, USA). Rabbit
anti-endothelin receptor A (EDNRA) polyclonal antibody (ab117529)
was from Abcam (Cambridge, MA, USA). The transwell insert was from
Corning, Inc. (Corning, NY, USA). Crystal violet was from Sigma
(St. Louis, MO, USA). TRIzol and the one-step RT-PCR kit were from
Takara (Dalian, China). The Lipofectamine™ 2000 kit, the wild-type
(Wt) vector pGL3-EDNRA 3′UTR-Wt and the mutant (Mut) vector
pGL3-EDNRA 3′UTR-Mut were obtained from Invitrogen (Carlsbad, CA,
USA). siRNA targeting EDNRA (siRNA EDRNA) and siRNA negative
control (NC) were prepared by Invitrogen and miR-200c mimics and
miR-200c NC were synthesized by RiboBio (Guangdong, China) based on
the primers listed in Table
I.
 | Table ISequences of primers for siRNA EDNRA,
siRNA NC, miR-200c mimics and miR-200c NC. |
Table I
Sequences of primers for siRNA EDNRA,
siRNA NC, miR-200c mimics and miR-200c NC.
| Gene | Primer sequences
(5′→3′) |
|---|
| siRNA | F:
UGGCAGUGUCUUAGCUGGUUG |
| EDNRA | R:
ACCAGCUAAGACACUGCCAUU |
| siRNA | F:
UUCUCCGAACGUGUCACGUTT |
| NC | R:
ACGUGACACGUUCGGAGAATT |
| miR-200c | F:
UACAGUACUGACCUGUCCACUCUGG |
| mimics | R:
CCGCCUUUGUGCUUCUGUUCUUCGU |
| miR-200c | F:
UUCUCCGAACGUGUCACGUTT |
| NC | R:
ACGUGACACGUUCGGAGAATT |
The human gastric carcinoma cell lines, BGC-823,
SGC-7901 and HGC-27, and the immortalized gastric mucosa epithelial
cell line, GES-1, were obtained from the Type Culture Collection of
the Chinese Academy of Sciences (Shanghai, China) and cultured in
DMEM containing 10% (v/v) fetal bovine serum (FBS) (both from
HyClone, Logan, UT, USA).
Determination of miR-200c and EDNRA
expression levels by RT-PCR and western blot analysis
For RT-PCR, total RNA from the BGC-823, SGC-7901,
HGC-27 and GES-1 cells was extracted using TRIzol reagent according
to the manufacturer's instructions and the RNA purity was evaluated
by determining the absorbance at 260 and 280 nm with UV-vis
spectrophotometry. RNA was then reverse transcribed into cDNA and
amplified with the one-step RT-PCR kit. The primers as shown in
Table II were added to the 25
μl of the PCR reaction system. The PCR parameters were set
as follows: denaturation for 45 sec at 94°C, annealing for 45 sec
at 59°C, elongation for 60 sec at 72°C and 35 circles. The
amplification products were loaded to conduct 2% (w/v) agarose gel
electrophoresis. Electrophoresis strips were analyzed on a gel
imaging system (ChemiDoc™ XRS; Bio-Rad, Hercules, CA, USA).
| Table IISequences of primers used for
RT-PCR. |
Table II
Sequences of primers used for
RT-PCR.
| Gene | Primer sequences
(5′→3′) |
|---|
| miR-200c | F:
GTAAATCGGTGTGTGTGTCGCGGGTCG |
| R:
CAAACCCTCCGAAACCCCTCGACCG |
| EDNRA | F:
GCTCTAGATAGGTAGCAACGTGGCTTT |
| R:
GCTCTAGAGCCCCAAAACTTGTCAAC |
| GAPDH | F:
AGCCACATCGCTCAGACA |
| R:
TGGACTCCACGACGTACT |
For western blot analysis, the BGC-823, SGC-7901,
HGC-27 and GES-1 cells were seeded into 6-well plates. After 48 h,
the cells were collected and mixed with moderate RIPA buffer. The
mixture was centrifuged and the supernatant was carefully collected
to obtain the total protein. The protein content was determined
using the BCA kit. The protein was degenerated, quantified and
loaded to conduct sodium dodecyl sulfate-polyacrylamide gel
electrophoresis (SDS-PAGE) electrophoresis for 1–2 h. The protein
was then transferred to a membrane by a wet method for 30–40 min.
The membrane was incubated with rabbit anti-EDNRA polyclonal
antibody buffer (1:100 dilution) at 4°C overnight. After a rinse,
the membrane was immersed in HRP-labeled goat anti-rabbit IgG (H+L)
buffer (1:200 dilution) at room temperature for 1–2 h. Finally,
after rinsing again, enhanced chemiluminescence (ECL) solution
dripped onto the membrane and the membrane was exposed on a gel
imaging system (ChemiDoc™ XRS; Bio-Rad). Quantity One software
(Bio-Rad) was utilized to evaluate the gray values of proteins
strips.
After the siRNA EDNRA, siRNA NC, miR-200c mimics and
miR-200c NC were respectively transfected into the BGC-823 cells as
described below, the miR-200c expression level was assayed by
RT-PCR, and EDNRA expression was evaluated by RT-PCR and western
blot analysis once again.
Cell transfection
When the confluence of the BGC-823 cells reached
50–70%, siRNA EDNRA, siRNA NC, miR-200c mimics or miR-200c NC were
respectively transfected into the cells using Lipofectamine™ 2000
kit according to the manufacturer's instructions. After 6 h, the
culture medium was replaced with DMEM containing 10% (v/v) FBS and
the cells were cultured at 37°C and 5% CO2 for a further
48 h. EDNRA expression was then determined by western blot analysis
and RT-PCR and miR-200c expression was measured by RT-PCR.
Determination of cell viability
The BGC-823 cells were seeded into 96-well plates.
When the cell confluence reached 50%, siRNA EDNRA, siRNA NC,
miR-200c mimics, or miR-200c NC were respectively transfected into
the cells as described above. After 48 h, 20 μl of MTT (5
mg/ml) were added to each well for 4 h. The culture medium was then
discarded and 150 μl of dimethyl sulfoxide were added to
each well. Vibration was conducted to fully dissolve the crystal
substances. Lastly, the plates were subjected to absorbance
measurement at 560 nm on a microplate reader (Infinite F200/M200;
Tecan, Männedorf, Switzerland). Relative cell viability was
calculated based on the absorbance (relative cell viability =
(ODtreatment−ODblank) /
(ODcontrol−ODblank) ×100%) and cells without
any treatment served as the controls.
Determination of cell apoptosis
The BGC-823 cells were seeded into 6-well plates.
When the cell confluence reached 50%, siRNA EDNRA, siRNA NC,
miR-200c mimics, or miR-200c NC were respectively transfected into
the cells as described above. After 48 h, the cells were stained
and fixed using the Hoechst staining kit according to the
manufacturer's instructions. The cells were then examined under a
microscope (AF6000; Leica, Wetzlar, Germany).
Determination of cell invasive
capacity
Matrigel was tiled uniformly onto the micromembrane
(8 μm) of the Transwell insert to prepare the gel. The
BGC-823 cells were seeded into 6-well plates. When the cell
confluence reached 50%, siRNA EDNRA, siRNA NC, miR-200c mimics, or
miR-200c NC were respectively transfected into the cells as
described above. After 48 h, the cells were digested and placed
into the upper chamber of the Transwell insert. The lower chamber
of the Transwell insert was filled with DMEM. After 48 h, the
Transwell insert was removed, fixed with paraformaldehyde and
stained with crystal violet. The cells in the lower chamber were
then conted under a light microscope (AF6000; Leica) in 5
individual visual fields. The average cell number in each visual
field represented the invasive capacity of the cells.
Luciferase reporter gene expression
analysis
The miR-200c mimics and EDNRA recombinant vectors
were co-transfected into the BGC-823 cells. The cells were divided
into the following groups: the miR-200c mimics + Wt EDNRA, miR-200c
NC + Wt EDNRA, miR-200c mimics + Mut EDNRA and the miR-200c NC +
Mut EDNRA groups. The dual-luciferase reporting system (Promega)
was employed to detect the activity of transfected luciferase
according to the manufacturer's instructions. Briefly, the
transfected cells were washed with phosphate-buffered solution
(PBS) 3 times and mixed with PLB. Following full lysis, LAR was
added to the mixture. The fluorescence intensity was then
determined with a dual-reporting luciferase detector. Following the
addition of stop buffer, the fluorescence intensity was determined
again. The relative fluorescence intensity was calculated according
to the following formula: relative fluorescence intensity =
fluorescence intensity of firefly luciferase/fluorescence intensity
of sea pansy luciferase.
Statistical analysis
All experiments were repeated at least 3 times and
data are expressed as the means ± standard deviation (SD).
Statistical analysis was conducted using an ANOVA followed by
Tukey's post hoc test with a SPSS 17.0 software. Significant
differences were defined at P<0.05.
Results
Original miR-200c expression levels in
gastric carcinoma cells and normal gastric cells
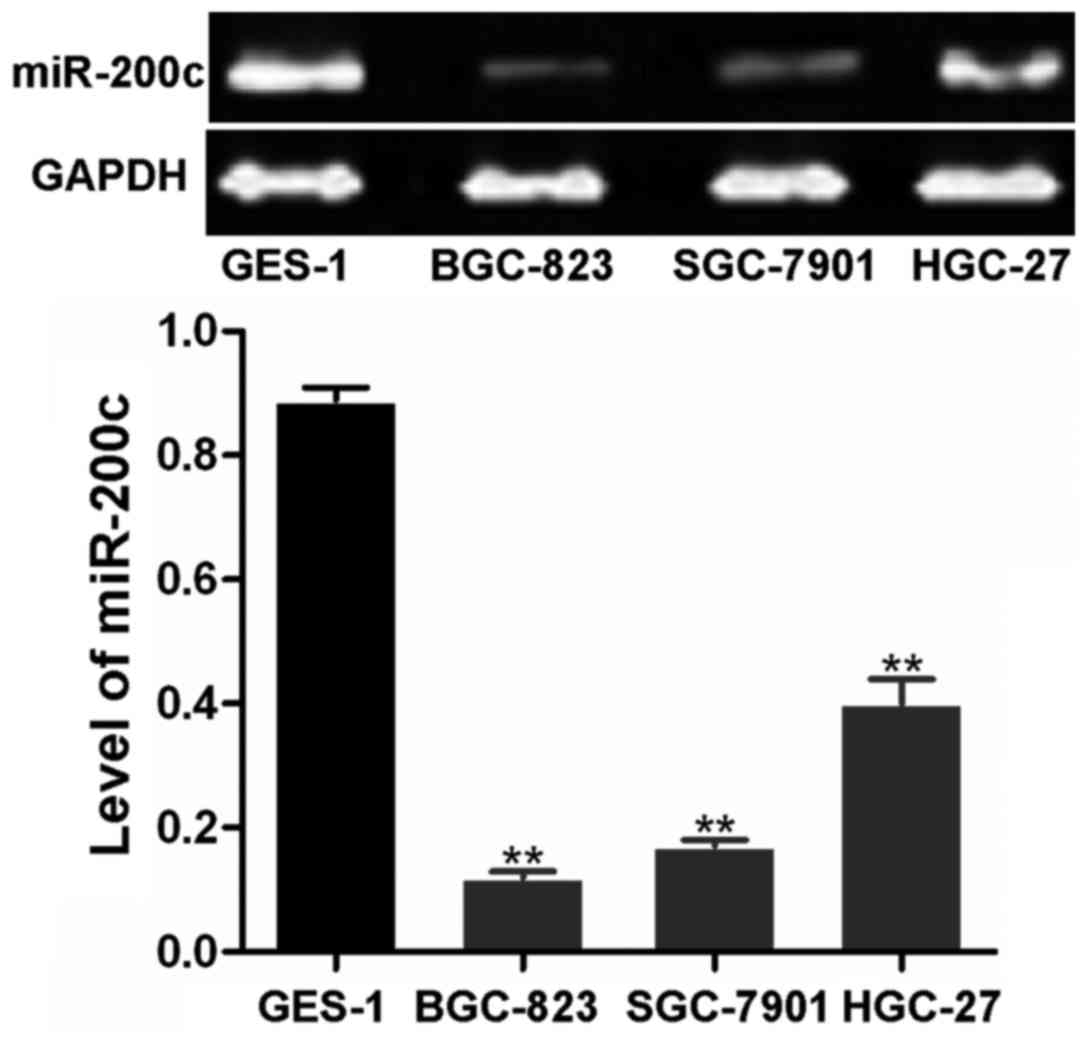
The miR-200c expression levels in the SGC-7901,
BGC-823 and HGC-27 cells were markedly lower than those in the
GES-1 cells (Fig. 1). It was
suggested that compared to the normal gastric cells, miR-200c
expression was downregulated in the gastric carcinoma cells. The
BGC-823 cells had the lowest expression of miR-200c, and thus they
were employed for the cell model of gastric carcinoma in the
following experiments.
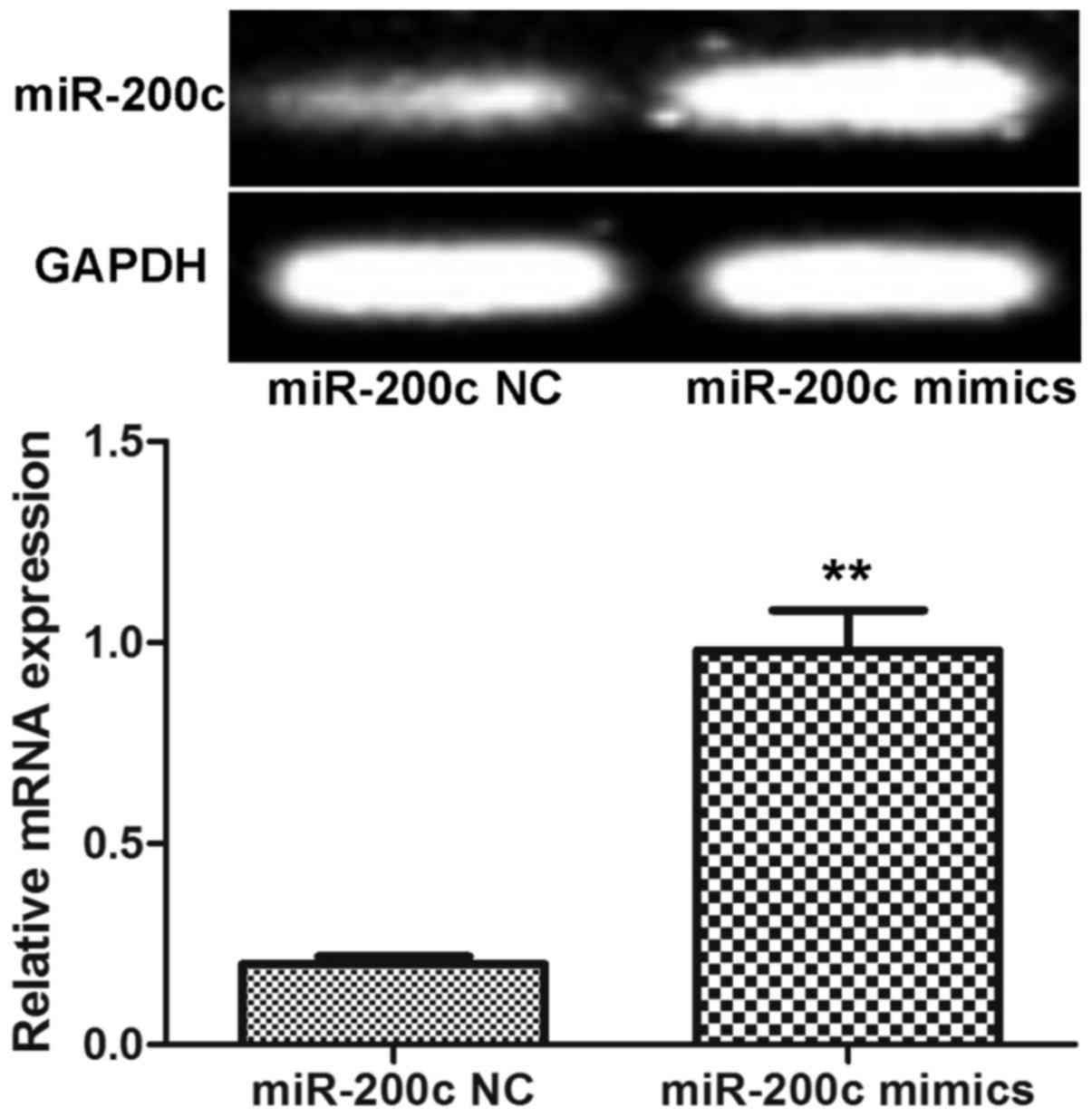
miR-200c expression level in gastric
carcinoma cells transfected with miR-200c mimics
At 6 h after the miR-200c mimics and miR-200c NC
were transfected into the BGC-823 cells, RT-PCR demonstrated that
the strip in the miR-200c mimics group was markedly lighter than
that in the miR-200c NC group (Fig.
2). Moreover, as compared with the miR-200c NC-transfected
cells (0.20±0.02), miR-200c expression following transfection with
miR-200c mimics (0.98±0.10) was significantly upregulated by almost
4-fold (P<0.01). These results confirmed the successful
transfection of the miR-200c mimics into the BGC-823 cells.
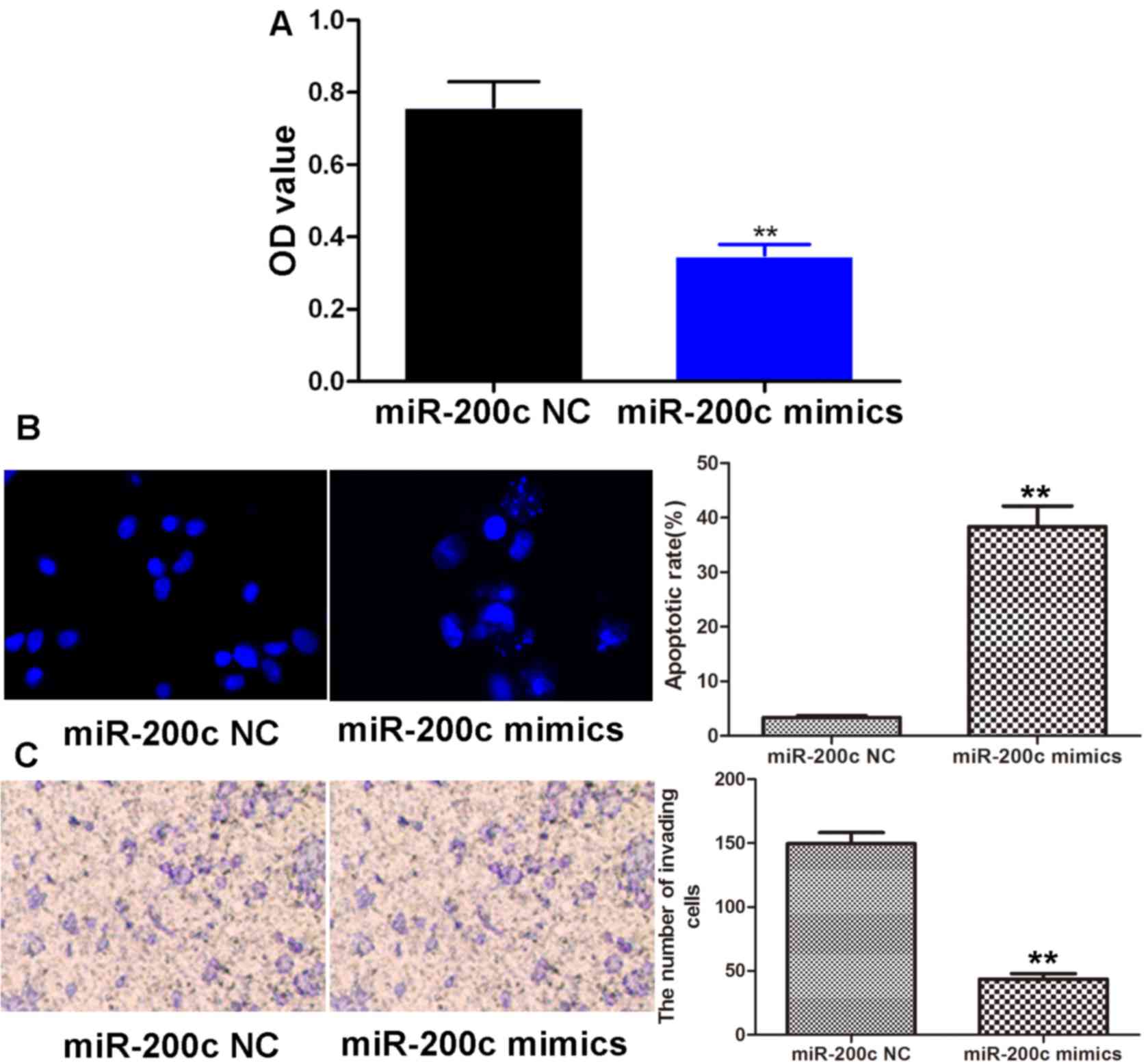
Effects of miR-200c mimics on the
viability, apoptosis and invasive capacity of gastric carcinoma
cells
We then examined the viability, apoptosis and
invasive capacity of the BGC-823 cells following transfection with
miR-200c mimics or miR-200c NC MTT assay, Hoechst staining and
Transwell assay, respectively. We found that compared to the
miR-200c NC-transfected cells, transfection with miR-200c mimics
significantly reduced the viability of the BGC-823 cells
(P<0.01; Fig. 3A). As shown in
Fig. 3B, the bright green-colored
cells represented the apoptotic cells. It was be observed that the
bright green-colored cells in the miR-200c mimics group were
notably greater in number than those in the miR-200c NC group.
Quantitatively, the apoptotic rate following transfection with
miR-200c mimics (38.46±3.85%) increased by >10-fold as compared
to transfection with miR-200c NC (3.34±0.33%, P<0.01).
Furthermore, as shown in Fig. 3C,
the number of invading cells in the miR-200c mimics group
(43.50±4.34) was markedly lower than that in the miR-200c NC group
(149.52±8.74, P<0.01). In summary, miR-200c mimics were capable
of effectively suppressing the proliferation, promoting the
apoptosis suppressing the invasive capacity of the gastric
carcinoma cells.
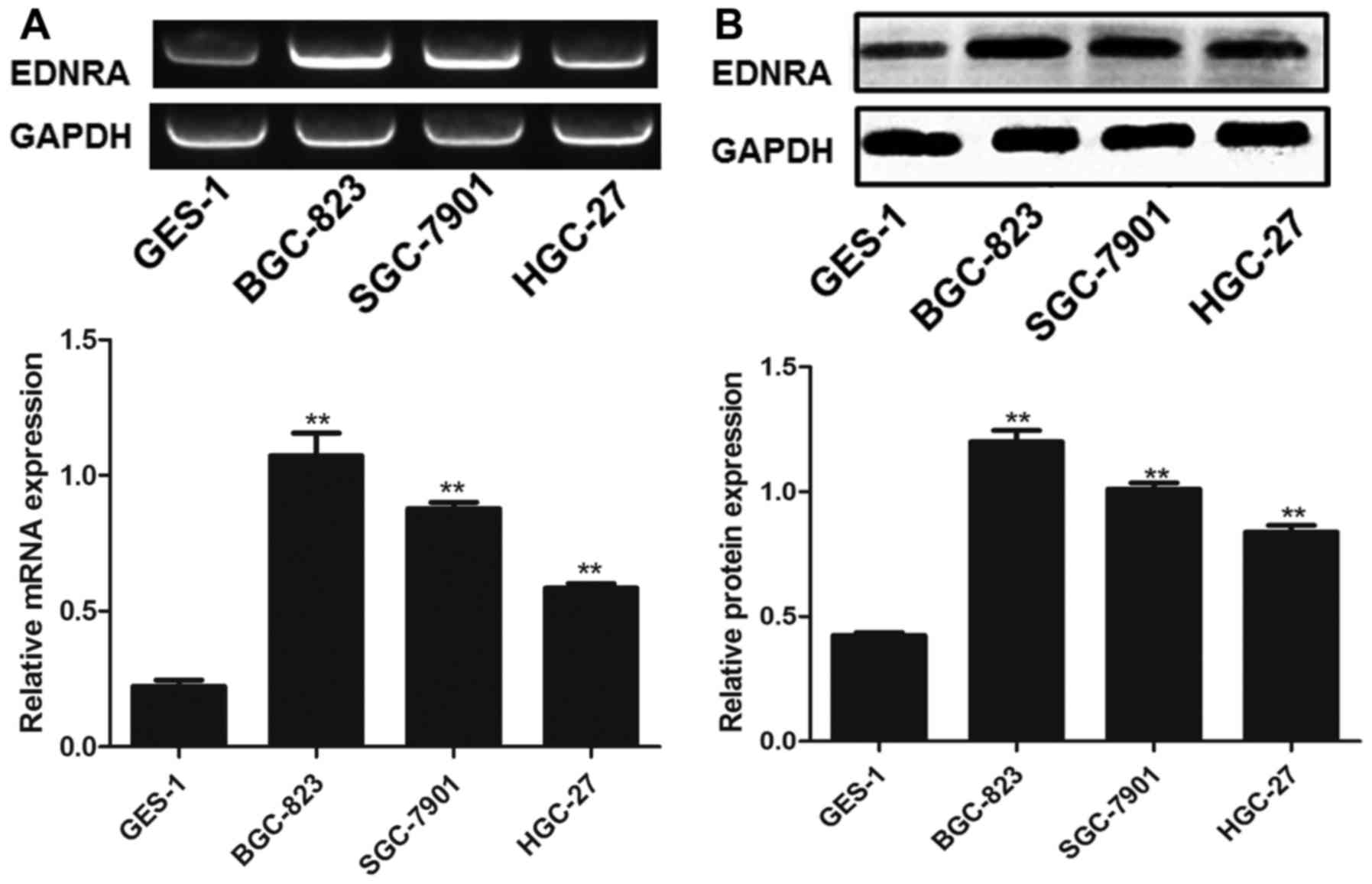
Original EDNRA expression levels in
gastric carcinoma cells and normal gastric cells
We examined the original EDNRA expression levels in
the SGC-7901, BGC-823, HGC-27 and GES-1 cells by RT-PCR and western
blot analysis. As shown in Fig.
4, the results revealed that the mRNA and protein levels of
EDNRA in the SGC-7901, BGC-823 and HGC-27 cells were significantly
higher than those in the GES-1 cells (all P<0.01). It was thus
indicated that EDNRA was overexpressed in diverse gastric carcinoma
cells, particularly in the BGC-823 cells.
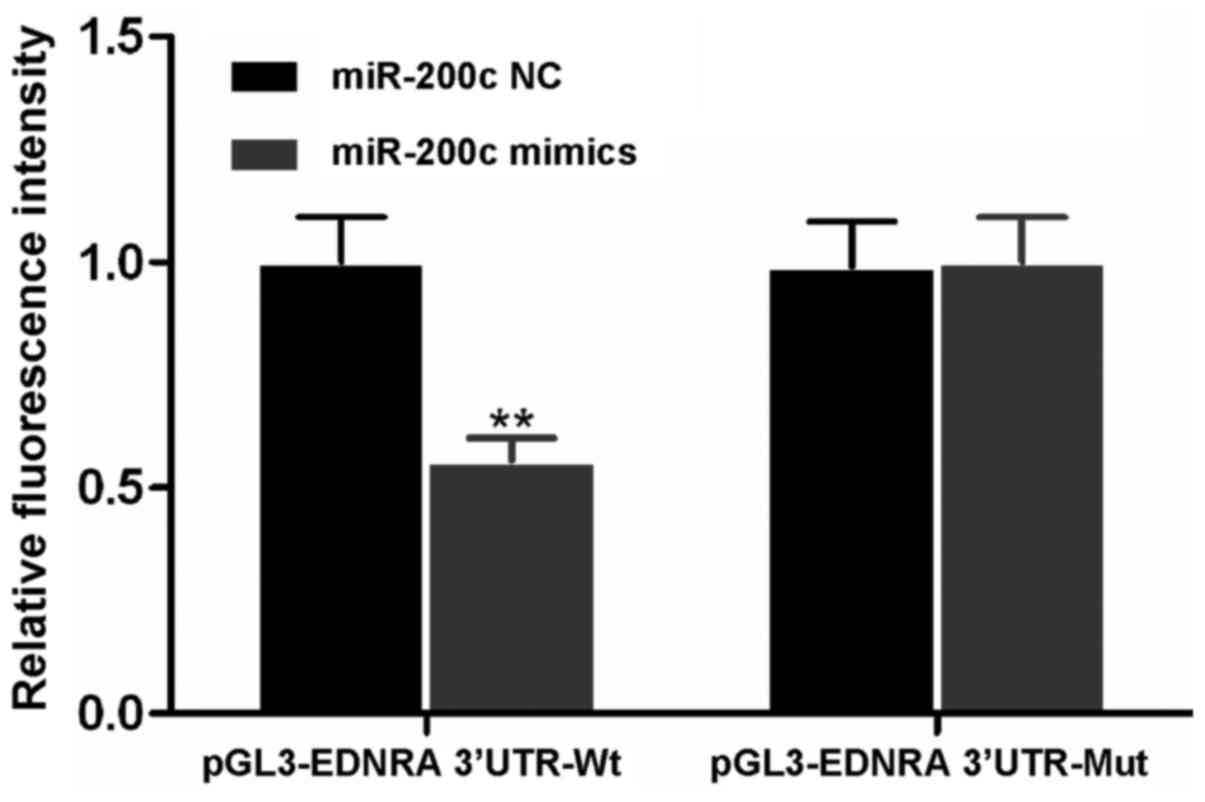
Luciferase reporter gene expression
analysis
The miR-200c mimics, miR-200c NC, Wt vector
pGL3-EDNRA 3′UTR-Wt, and Mut vector pGL3-EDNRA 3′UTR-Mut were
transfected into the BGC-823 cells. We found that the fluorescence
intensities in the co-transfection groups transfected with miR-200c
NC and either of the two vectors were similar (Fig. 5). There was no significant
difference in the fluorescence intensity between the cells
transfected with the miR-200c mimics or miR-200c NC and the
pGL3-EDNRA 3′UTR-Mut vector (P>0.05). However, following
co-transfection with miR-200c mimics and the pGL3-EDNRA 3′UTR-Wt
vector, the fluorescence intensity was significantly decreased as
compared with the other 3 co-transfection groups (P<0.01). These
results suggested that EDNRA was the downstream target gene of
miR-200c and that EDNRA may be regulated by miR-200c.
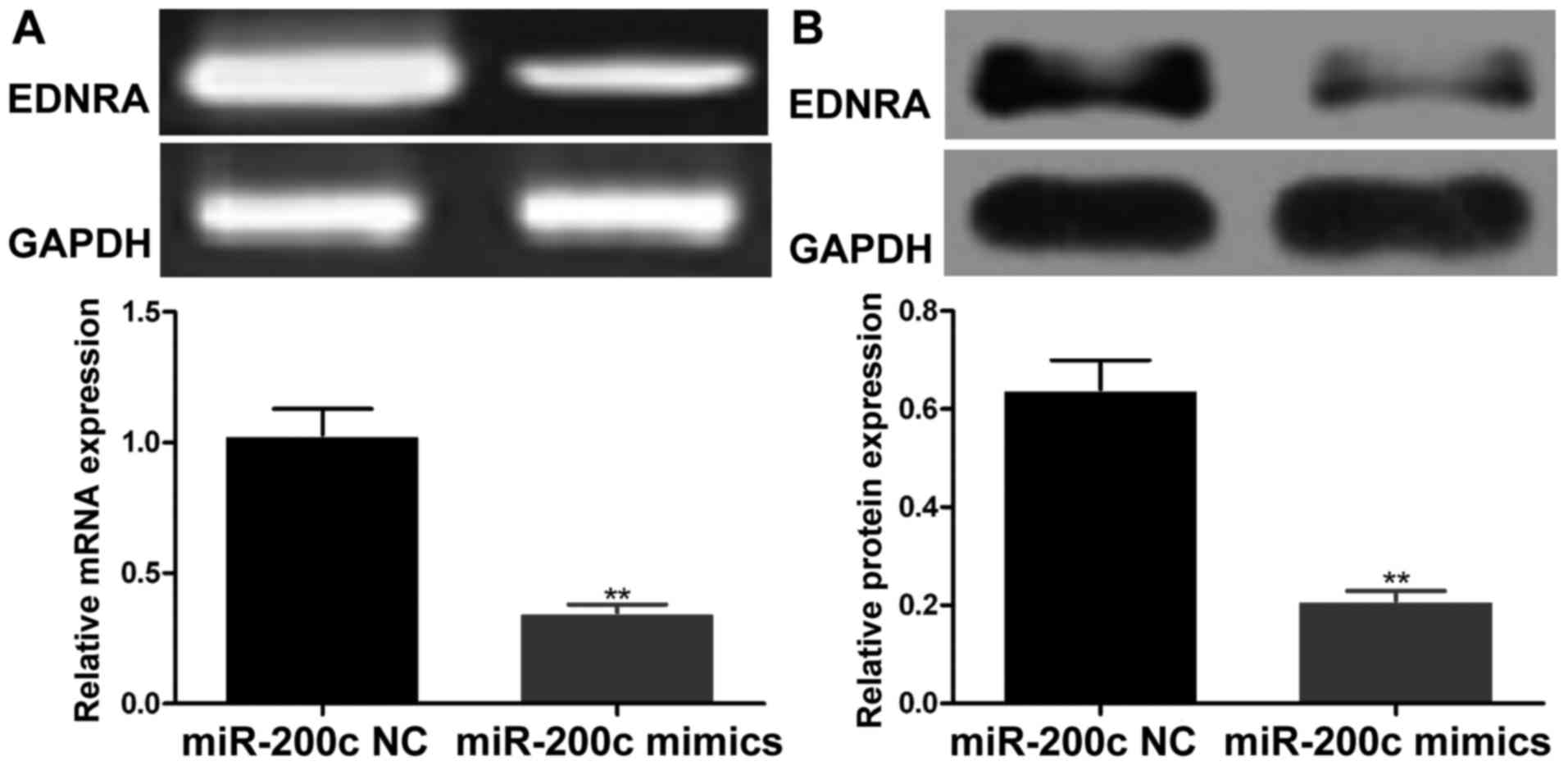
Effects of miR-200c mimics on the EDNRA
expression levels in gastric carcinoma cells
RT-PCR and western blot analysis respectively
revealed that compared to the miR-200c NC-transfected cells,
transfection with miR-200c mimics markedly downregulated the mRNA
and protein expression levels of EDNRA in the BGC-823 cells
(P<0.01; Fig. 6).
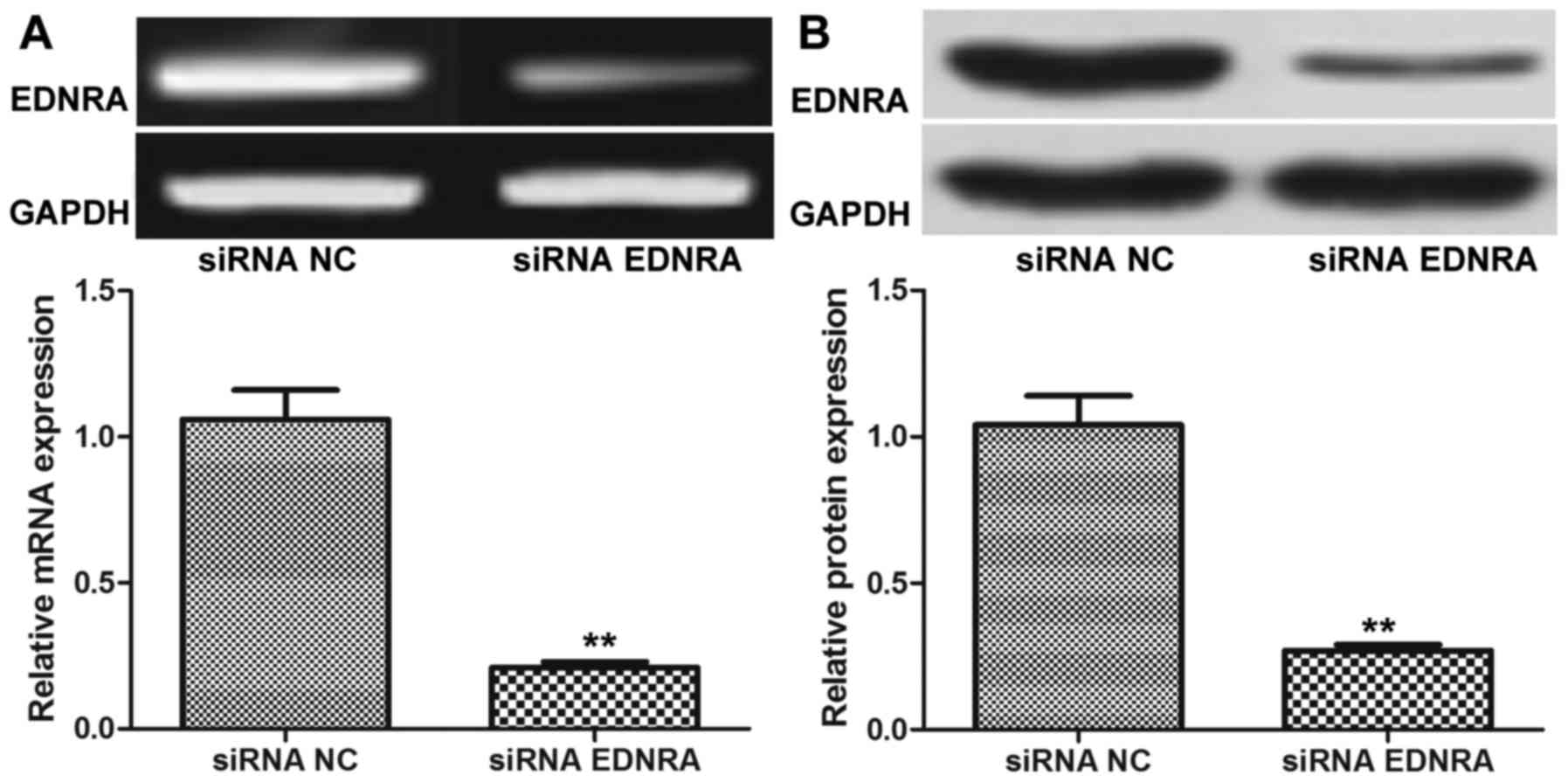
EDNRA expression levels in siRNA
EDNRA-transfected gastric carcinoma cells
After the siRNA EDNRA or siRNA NC were transfected
into the BGC-823 cells, the blots in the siRNA EDNRA group in both
RT-PCR and western blot analysis were markedly fainter in
comparison to those in the siRNA NC group (Fig. 7). As shown by quantitative western
blot analysis, the cells in the siRNA EDNRA group (0.21±0.02)
produced significantly less EDNRA protein than the cells in the
siRNA NC group (1.06±0.10, P<0.01). This similar difference
between siRNA EDNRA and siRNA NC was also found in the quantitative
RT-PCR analysis for the EDNRA mRNA levels (0.27±0.02 vs. 1.04±0.10,
P<0.01). It was thus suggested that siRNA EDNRA downregulated
the expression of EDNRA in the gastric carcinoma cells, which was
similar to the effects of miR-200c mimics on EDNRA expression.
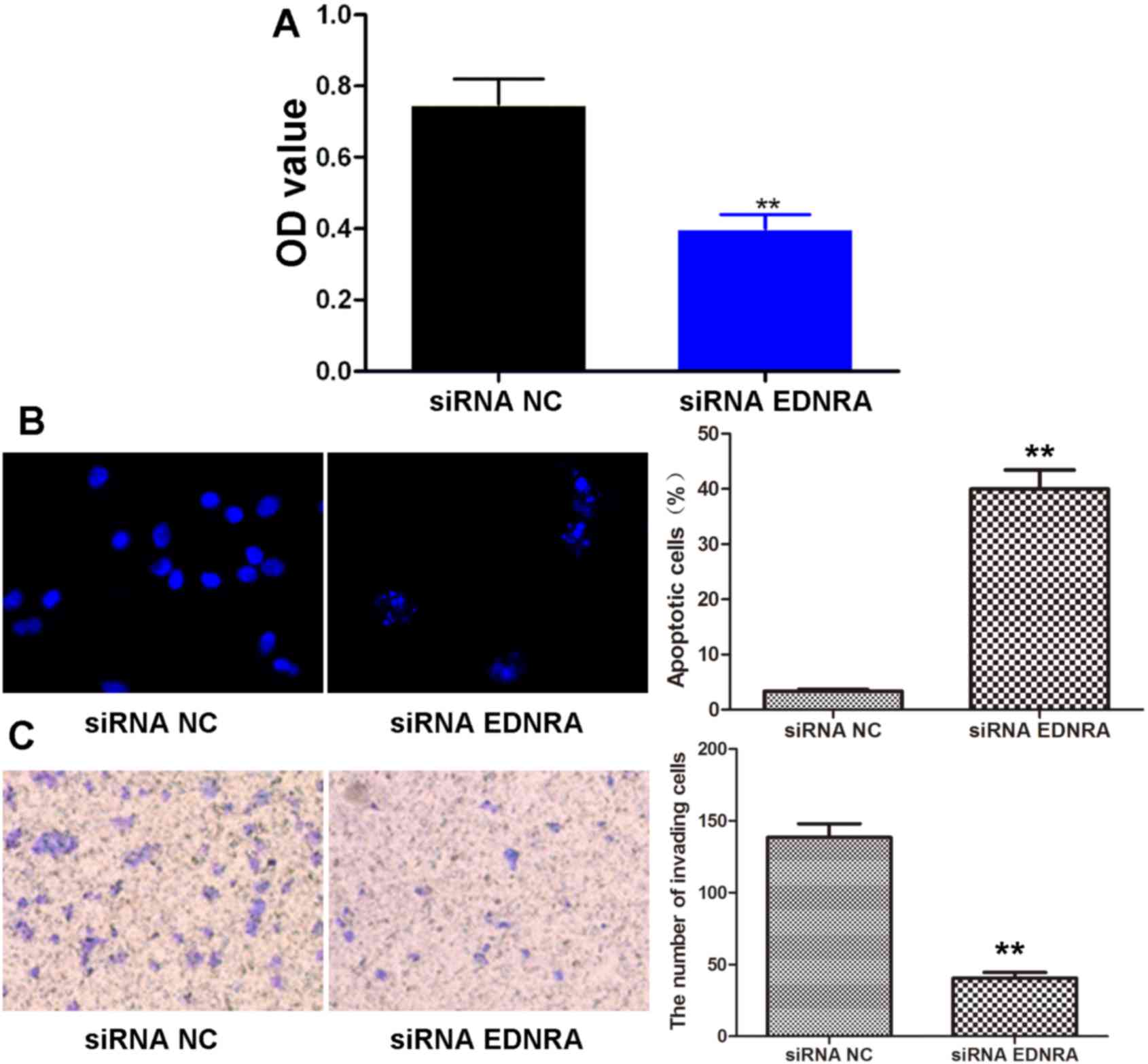
Effects of siRNA EDNRA on the viability,
apoptosis and invasive capacity of gastric carcinoma cells
We then examined the viability, apoptosis and
invasive capacity of the BGC-823 cells following transfection with
siRNA EDNRA or siRNA NC by MTT assay, Hoechst staining and
Transwell assay, respectively. We found that compared to
transfection with siRNA NC, transfection with siRNA EDNRA
significantly decreased the viability of the BGC-823 cells
(P<0.01; Fig. 8A). As shown in
Fig. 8B, the bright green-colored
apoptotic cells in the siRNA EDNRA group were markedly greater in
number than those in the siRNA NC group. Quantitatively, the
apoptotic rate following transfection with siRNA EDNRA
(40.46±3.99%) was increased by >11-fold as compared to
transfection with siRNA NC (3.37±0.32%, P<0.01). Moreover, as
shown in Fig. 8C, the number of
invading cells in the siRNA EDNRA group (40.51±4.03) was
significantly less than that in the siRNA NC group (138.53±9.58,
P<0.01). Thus, siRNA EDNRA were able to efficiently block the
proliferation, enhance the apoptosis and inhibit the invasion of
gastric carcinoma cells.
Discussion
As a member of the miR-200 family, miR-200c is
located on human chromosome 12p13 and it is adjacent to and
clustered with miR-141, another member of the miR-200 family.
miR-200c was screened from the miRNA of breast cancer cells, lung
cancer cells, fibrosarcoma cells, melanoma cells and normal human
bronchial epithelial cells by Hurteau et al (9) in 2006 and Zinc finger E-box-binding
homeobox 1 (ZEB1) was confirmed to its target gene through
bioinformatics (10,11). Subsequently, Ceppi et al
revealed the differential expression of miR-200c between lung
cancer and normal lung tissue (12). Thereafter, it has been
successively demonstrated the abnormal expression of miR-200c in
liver cancer, renal cancer, pancreatic cancer, bladder cancer and
ovarian cancer tissues and cells (13 and refs therein). Moreover, miR-200c
was found to regulate the growth, proliferation, apoptosis,
metastasis and invasion of cancer cells by affecting the expression
of target genes, such as ZEB1, 1-phosphatidylinositol-3-phosphate
5-kinase (FAB1), homeobox D10 (HOXD10) and BMI1, and to be closely
associated with epithelial mesenchymal transition (EMT) in cancer
(14,15). In recent years, miR-200c has also
been found to be associated with the genesis and development of
gastric carcinoma. Blanco-Calvo et al demonstrated that the
expression of miR-200c in gastric carcinoma tissues was
significantly inferior to that in normal tissues (16). Chang et al adopted the
RT-PCR method to reveal that the expression levels of miR-200
family members, including miR-200c in 46 cases of gastric cancer
were lower than those in the pericarcinomatous tissues, and that
they were strongly linked to pathological patterns and angiogenesis
(17). Consequently, it was
suggested that miR-200c may be the judging indicator of gastric
carcinoma invasion and may also be an effective indicator of
patient prognosis. Although these studies have demonstrated the
close association between miR-200c and the genesis and development
of gastric carcinoma, the specific mechanisms involved remain
unknown. Therefore, in this study, we investigated the molecular
mechanisms involved in the influence of miR-200c on the biological
behaviors of gastric carcinoma cells, such as proliferation,
apoptosis and invasion. Firstly, we revealed the downregulated
expression of miR-200c in 3 gastric carcinoma cell lines in
comparison to a normal gastric cell line. This result was in line
with that of a previous study (18). After considering the lowest
miR-200c expression level, histological grade and the toxicity of
the transfection agents, the BGC-823 cells were employed as the
model gastric carcinoma cells in the following experiments. We then
successfully transfected the miR-200c mimics and miR-200c NC into
gastric carcinoma cells using Lipofectamine 2000 which was verified
by RT-PCR. MTT assay and Hoechst staining also demonstrated that as
compared to transfection with miR-200c NC, transfection with
miR-200c mimics markedly suppressed the proliferation and promoted
the apoptosis of gastric carcinoma cells, which was consistent with
the effects of miR-200c mimics on breast cancer cells, renal cancer
cells and colon cancer cells in previous studies (19–21). Furthermore, Transwell assay
revealed that transfection with miR-200c mimics notably suppressed
the invasive ability of the gastric carcinoma cells, which was in
accordance with the roles of miR-200c mimics in bladder cancer
cells, colon cancer cells and head and neck squamous cell carcinoma
(22–24). Thus, miR-200c mimics were able to
strongly inhibit the proliferation and invasion, and promote the
apoptosis of gastric carcinoma cells.
Similar to numerous miRNAs, miR-200c affected the
biological behavior of cancer cells by regulating the target genes.
Tang et al reported that exogenous miR-200c inhibited the
growth and invasion of cancer cells by targeting DNMT3A, DNA
methyltransferase 3 beta (DNMT3B) and SP1 (25). Saito et al demonstrated
that miR-200c affected EMT in cancer cells by targeting ZEB and
then regulating E-cadherin expression (26). Zhou et al revealed that the
upregulation of miR-200c expression affected the metastasis and
invasion of gastric carcinoma through the targeted downregulation
of ZEB1/2 expression (27).
Moreover, the alterations in EMT regulated by ZEB1 have been shown
to be associated with the EMT-related gene, EDNRA (28), and the EDNRA signal can regulate
EMT (29). As a G-protein coupled
receptor, EDNRA contains 7 transmembrane domains and mediates
signaling pathways regulated by G-protein coupled receptor kinases.
Its affinities with different endothelin are distinctive with a
strongest affinity with endo-thelin-1 (ET-1). Thus, it has a potent
vasoconstrictive effect. The gene polymorphism of EDNRA is closely
related to cardiovascular disease, immune disease and cancer
(30,31). Furthermore, the overexpression of
EDNRA in bladder cancer cells, epithelial ovarian cancer, and
prostate cancer has been shown to be closely associated with the
genesis and development of cancer, and ET-1/EDNRA regulates EMT and
chemotherapeutic drug resistance in epithelial ovarian cancer cells
(32–34). However, the specific mechanisms of
action of EDNRA in gastric carcinoma have not yet been revealed.
Furthermore, EDNRA may be the target gene of miR-200c (35). Therefore, the specific mechanisms
of action of EDNRA in gastric carcinoma and the association between
miR-200c and EDNRA were investigated in this study. Based on RT-PCR
and western blot analysis, respectively, we first demonstrated the
mRNA and protein overexpression of EDNRA in gastric carcinoma cells
in comparison with that of normal gastric cells, and the resutls
were in agreement with the expression tendency of EDNRA in other
cancer tissues (32,33).
We then investigated whether EDNRA is the target
gene of miR-200c by utilizing luciferase reporter gene assay. The
fluorescence intensity in the co-transfection group of miR-200c
mimics and Wt vector pGL3-EDNRA 3′UTR-Wt was markedly inferior to
that in the other 3 co-transfection groups, suggesting that EDNRA
was the target gene of miR-200c. This interesting result was also
confirmed by the results of RT-PCR and western blot analysis,
showing that transfectoin with miR-200c mimics markedly
downregulated the mRNA and protein expression of EDNRA in gastric
carcinoma cells. In consequence, we speculated that miR-200c may
affect the biological behavior of gastric carcinoma cells, such as
proliferation, apoptosis and invasion by regulating the expression
of the downstream target gene, EDNRA. However, as miR-200c may have
multiple target genes, we transfected siRNA EDNRA into the gastric
carcinoma cells to evaluate the effects of siRNA EDNRA on EDNRA
expression and the biological behavior of gastric carcinoma cells.
Following transfection with siRNA EDNRA, the results of RT-PCR and
western blot analysis revealed that the mRNA and protein expression
levels of EDNRA were downregulated, which was similar to the
effects of miR-200c mimics on EDNRA expression. In addition, after
the silencing of EDNRA by siRNA EDNRA, the proliferative and
invasive capacity of the gastric carcinoma cells was markedly
suppressed and apoptosis was promoted; these effects were similar
to those observed with transfection with miR-200c mimics. Thus,
this confirms our previous speculation that miR-200c mimics
disrupted the proliferation, apoptosis and invasion of gastric
carcinoma cells by targeting EDNRA.
In conclusion, our findings demonsrated that
miR-200c expression was downregulated in gastric carcinoma cells,
while this downregulation was reversed by transfection with
miR-200c mimics. EDNRA was overexpressed in gastric carcinoma
cells, while its overexpression was downregulated by transfection
with siRNA EDNRA or miR-200c. EDNRA was validated to be the
downstream target gene of miR-200c. miR-200c regulated the
biological behavior of gastric carcinoma cells by regulating their
proliferation, apoptosis and invasion via the downregulation of the
expression of EDNRA. These results may provide new insight into the
diagnosis and treatment of gastric carcinoma by providing an
effective biomarker (miR-200c and EDNRA).
Acknowledgments
The authors are thankful for the financial support
from the Provincial Non-profit Technology Application Research
Project of Zhejiang Province (2014C33140) and the Science and
Technology Research Project of Jinhua, Zhejiang (2014-3-076).
References
|
1
|
Halkia E, Kopanakis N, Nikolaou G and
Spiliotis J: Cytoreductive surgery and HIPEC for peritoneal
carcinomatosis. A review on morbidity and mortality. J BUON.
20(Suppl 1): S80–S87. 2015.PubMed/NCBI
|
|
2
|
Spych M, Serbiak B, Rychter A,
esien-Lewandowicz E, Gottwald L and Fijuth J: Post-operative
radiochemotherapy in patients with gastric cancer: one department's
experience of 56 patients. Br J Radiol. 84:457–463. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Mizuguchi Y, Takizawa T, Yoshida H and
Uchida E: Dysregulated miRNA in progression of hepatocellular
carcinoma: A systematic review. Hepatol Res. 46:391–406. 2016.
View Article : Google Scholar
|
|
4
|
Wang QX, Zhu YQ, Zhang H and Xiao J:
Altered MiRNA expression in gastric cancer: A systematic review and
meta-analysis. Cell Physiol Biochem. 35:933–944. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Reddy KB: MicroRNA (miRNA) in cancer.
Cancer Cell Int. 15:382015. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Feng X, Wang Z, Fillmore R and Xi Y:
MiR-200, a new star miRNA in human cancer. Cancer Lett.
344:166–173. 2014. View Article : Google Scholar :
|
|
7
|
Shi C and Zhang Z: The prognostic value of
the miR-200 family in ovarian cancer: A meta-analysis. Acta Obstet
Gynecol Scand. 95:505–512. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Song F, Yang D, Liu B, Guo Y, Zheng H, Li
L, Wang T, Yu J, Zhao Y, Niu R, et al: Integrated microRNA network
analyses identify a poor-prognosis subtype of gastric cancer
characterized by the miR-200 family. Clin Cancer Res. 20:878–889.
2014. View Article : Google Scholar
|
|
9
|
Hurteau GJ, Spivack SD and Brock GJ:
Potential mRNA degradation targets of hsa-miR-200c, identified
using informatics and qRT-PCR. Cell Cycle. 5:1951–1956. 2006.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kumar S, Nag A and Mandal CC: A
comprehensive review on miR-200c, a promising cancer biomarker with
therapeutic potential. Curr Drug Targets. 16:1381–1403. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang KC, Xi HQ, Cui JX, Shen WS, Li JY,
Wei B and Chen L: Prognostic role of miR-200c in various
malignancies: A systematic review and meta-analysis. Int J Clin Exp
Med. 8:1931–1943. 2015.PubMed/NCBI
|
|
12
|
Ceppi P, Mudduluru G, Kumarswamy R, Rapa
I, Scagliotti GV, Papotti M and Allgayer H: Loss of miR-200c
expression induces an aggressive, invasive, and chemoresistant
phenotype in non-small cell lung cancer. Mol Cancer Res.
8:1207–1216. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Mutlu M, Raza U, Saatci Ö, Eyüpoğlu E,
Yurdusev E and Şahin Ö: miR-200c: A versatile watchdog in cancer
progression, EMT, and drug resistance. J Mol Med (Berl).
94:629–644. 2016. View Article : Google Scholar
|
|
14
|
Tan G, Qiu M, Chen H and Liu J: The
relationship between miR-200c and the progression of tumor. Chin J
Cancer Biother. 22:120–124. 2015.In Chinese.
|
|
15
|
Fan H and Ye M: Research progress of
micro-RNA200s in tumor metastasis. Chin Cancer. 21:840–844. 2012.In
Chinese.
|
|
16
|
Blanco-Calvo M, Tarrío N, Reboredo M,
Haz-Conde M, García J, Quindós M, Figueroa A, Antón-Aparicio L,
Calvo L and Valladares-Ayerbes M: Circulating levels of GDF15, MMP7
and miR-200c as a poor prognostic signature in gastric cancer.
Future Oncol. 10:1187–1202. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Chang L, Guo F, Huo B, Lv Y, Wang Y and
Liu W: Expression and clinical significance of the microRNA-200
family in gastric cancer. Oncol Lett. 9:2317–2324. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Cui FB, Liu Q, Li RT, Shen J, Wu PY, Yu
LX, Hu WJ, Wu FL, Jiang CP, Yue GF, et al: Enhancement of
radiotherapy efficacy by miR-200c-loaded gelatinase-stimuli
PEG-Pep-PCL nanoparticles in gastric cancer cells. Int J
Nanomedicine. 9:2345–2358. 2014.PubMed/NCBI
|
|
19
|
Radisky DC: miR-200c at the nexus of
epithelial-mesenchymal transition, resistance to apoptosis, and the
breast cancer stem cell phenotype. Breast Cancer Res. 13:1102011.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Gao C, Peng FH and Peng LK: MiR-200c
sensitizes clear-cell renal cell carcinoma cells to sorafenib and
imatinib by targeting heme oxygenase-1. Neoplasma. 61:680–689.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Chen J, Wang W, Zhang Y, Hu T and Chen Y:
The roles of miR-200c in colon cancer and associated molecular
mechanisms. Tumour Biol. 35:6475–6483. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mahdavinezhad A, Mousavi-Bahar SH,
Poorolajal J, Yadegarazari R, Jafari M, Shabab N and Saidijam M:
Evaluation of miR-141, miR-200c, miR-30b expression and
clinicopathological features of bladder cancer. Int J Mol Cell Med.
4:32–39. 2015.PubMed/NCBI
|
|
23
|
Tanaka S, Hosokawa M, Ueda K and Iwakawa
S: Effects of decitabine on invasion and exosomal expression of
miR-200c and miR-141 in oxaliplatin-resistant colorectal cancer
cells. Biol Pharm Bull. 38:1272–1279. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Tamagawa S, Beder LB, Hotomi M, Gunduz M,
Yata K, Grenman R and Yamanaka N: Role of miR-200c/miR-141 in the
regulation of epithelial-mesenchymal transition and migration in
head and neck squamous cell carcinoma. Int J Mol Med. 33:879–886.
2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tang H, Deng M, Tang Y and Xie X, Guo J,
Kong Y, Ye F, Su Q and Xie X: miR-200b and miR-200c as prognostic
factors and mediators of gastric cancer cell progression. Clin
Cancer Res. 19:5602–5612. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Saito T, Kurashige J, Nambara S, Komatsu
H, Hirata H, Ueda M, Sakimura S, Uchi R, Takano Y, Shinden Y, et
al: A long non-coding RNA activated by transforming growth factor-β
is an independent prognostic marker of gastric cancer. Ann Surg
Oncol. 22(Suppl 3): S915–S922. 2015. View Article : Google Scholar
|
|
27
|
Zhou X, Wang Y, Shan B, Han J, Zhu H, Lv
Y, Fan X, Sang M, Liu XD and Liu W: The downregulation of
miR-200c/141 promotes ZEB1/2 expression and gastric cancer
progression. Med Oncol. 32:4282015. View Article : Google Scholar
|
|
28
|
Ohta H, Aoyagi K, Fukaya M, Danjoh I, Ohta
A, Isohata N, Saeki N, Taniguchi H, Sakamoto H, Shimoda T, et al:
Cross talk between hedgehog and epithelial-mesenchymal transition
pathways in gastric pit cells and in diffuse-type gastric cancers.
Br J Cancer. 100:389–398. 2009. View Article : Google Scholar
|
|
29
|
Rosanò L, Cianfrocca R, Tocci P, Spinella
F, Di Castro V, Caprara V, Semprucci E, Ferrandina G, Natali PG and
Bagnato A: Endothelin A receptor/β-arrestin signaling to the Wnt
pathway renders ovarian cancer cells resistant to chemotherapy.
Cancer Res. 74:7453–7464. 2014. View Article : Google Scholar
|
|
30
|
López-Mejías R, Genre F, García-Bermúdez
M, Ubilla B, Castañeda S, Llorca J, González-Juanatey C, Corrales
A, Miranda-Filloy JA, Pina T, et al: Lack of association between
ABO, PPAP2B, ADAMST7, PIK3CG, and EDNRA and carotid intima-media
thickness, carotid plaques, and cardiovascular disease in patients
with rheumatoid arthritis. Mediators Inflamm. 2014:7562792014.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Low SK, Takahashi A, Cha PC, Zembutsu H,
Kamatani N, Kubo M and Nakamura Y: Genome-wide association study
for intracranial aneurysm in the Japanese population identifies
three candidate susceptible loci and a functional genetic variant
at EDNRA. Hum Mol Genet. 21:2102–2110. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Laurberg JR, Jensen JB, Schepeler T, Borre
M, Ørntoft TF and Dyrskjøt L: High expression of GEM and EDNRA is
associated with metastasis and poor outcome in patients with
advanced bladder cancer. BMC Cancer. 14:6382014. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Pflug BR, Zheng H, Udan MS, D'Antonio JM,
Marshall FF, Brooks JD and Nelson JB: Endothelin-1 promotes cell
survival in renal cell carcinoma through the ET(A) receptor. Cancer
Lett. 246:139–148. 2007. View Article : Google Scholar
|
|
34
|
Kar SP, Beesley J, Amin Al Olama A,
Michailidou K, Tyrer J, Kote-Jarai Z, Lawrenson K, Lindstrom S,
Ramus SJ, Thompson DJ, et al: Genome-wide meta-analyses of breast,
ovarian and prostate cancer association studies identify multiple
new susceptibility loci shared by at least two cancer types. Cancer
Discov. 6:1052–1067. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Wang S, Xiao Y, Liu Z, Yang X, Xiong X,
Zhu W, Shi H and Luo D: Network-guided genetic screening for
metastasis-related microRNA-200c in breast cancer. Tumor.
33:111–118. 2013.In Chinese.
|