Introduction
Neuropathic pain caused by nerve injury remains an
intractable disease due to a lack of satisfactory treatment
(1–3). In 2009, Ciaramitaro et al
(4) carried out a multicenter
prospective study on the prevalence of neuropathic pain after
traumatic brachial plexus injury. From the 107 patients enrolled,
56% had neuropathic pain. Neuropathic pain impaired the quality of
life and caused depression. Brachial plexus avulsion (BPA) induces
a characteristic persistent oppression and intermittent shooting
pain, which is often difficult to cure (5,6).
The pain could be experienced as burning or a feeling of
compression. Pain after BPA is resistant to most traditional pain
relief treatments (7) due to a
lack of understanding of the cellular or molecular mechanisms in
the occurrence and development of pain (8,9).
As long-term sensitivity changes in neuropathic pain
are associated with changes in gene regulation, it is worthy of
note to determine if the microRNAs (miRNAs/miRs) that regulate
genes taking part in the nociceptive pathways affect the occurrence
and development of pain (10).
The key roles of miRNAs in nervous system development and
pathophysiology is increasingly evident (11), however, certain facts remain to be
elucidated. Understanding the gene regulatory events in
miRNA-mediated neuropathic pain may provide a pathway for
identifying biomarkers or finding novel therapeutic targets
(12). Since pathophysiological
changes in pain are associated with altered expression in
pain-associated proteins, miRNA may be a promising tool for
controlling inflammatory and neuropathic pain by regulating gene
and protein expression in pain pathways (13). Few studies have been performed on
pain following brachial plexus injury (14–16).
In the present study, miRNA expression profiling was
performed of the thalamus (T), the anterior cingulate (AC), the
dorsal horn of the spinal cord (DHSC) and the blood (B) in a
neuropathic pain model 4 weeks after complete brachial plexus
avulsion (BPA) surgery. The combination of experimental and
bioinformatics methods was applied to identify the biological and
cytological functions that were influenced by the changes in miRNA
expression (17). These data
provide further evidence in support of the hypothesis that
post-complete BPA pain may be regulated by miRNA.
Materials and methods
Study approval
Animal handling and procedures used in this study
were in agreement with the guidelines of the Animal Care and Use
Committee of Fudan University (Shanghai, China). Neuropathic pain
was induced in male Sprague-Dawley rats (n=40; weight, 200–250 g;
age, 8 weeks; supplied by the Department of Laboratory Animal
Science, Fudan University, Shanghai, China) by CBPA. The rats were
kept in an environment with a temperature of 20°C and humidity of
50% and were maintained on a 12/12-h light/dark cycle and allowed
free access to food and water. The animal use protocol was reviewed
and approved by the Animal Ethics and Welfare Committee of Huashan
Hospital affiliated to Fudan University.
Animal behavioral tests
Mechanical allodynia
The rats were placed on a metal mesh floor, covered
by a transparent plastic box and raised 30 cm above the floor. The
plantar surface of the paw was stimulated with a series of
ascending force von Frey monofilaments. The threshold was taken as
the lowest force that evoked a brisk withdrawal response to one of
five repetitive stimuli (7). A
withdrawal response was considered valid only when the paw was
completely removed from the platform.
Cold allodynia
Cold allodynia was measured by an acetone spray
test, as described by Choi et al (18). Briefly, 250 ml acetone was
squirted onto the mid-plantar surface of the paw. The withdrawal
responses were evaluated on a scale of 0–3 points: 0 points, the
paw was not moved; 1 point, a response in which the paw had little
or no weight born on it; 2 points, a response in which the paw was
elevated and was not in contact with any surface; and 3 points, a
vigorous response in which the rat licked, bit or shook the
paw.
All behavioral tests were performed by the same
technician who was blinded to the study groups and identification
of animals in order to avoid subjective differences in
interpretation, which could occur with different observers
(19).
Surgery procedure
Following the animal behavioral tests, the rats were
anesthetized with sodium pentobarbital injected intraperitoneally.
The CBPA rat model (20) was used
to induce neuropathic pain. The brachial plexus was approached
through a horizontal supraclavicular incision. The
sternocleidomastoid muscle was cut off and the omohyoid muscle was
pulled aside, leaving the transverse cervical vessels intact. The
brachial plexus was located in the scalene fissure, including
upper, middle and lower trunks. The complete brachial plexus was
grasped with forceps and extracted from the spinal cord by
traction. The tissue layers were then brought together and the skin
was closed with 4-0 silk sutures.
These rats were maintained on a 12/12 h light/dark
cycle and allowed free access to food and water for one month. At
the end of the 4 weeks, paw withdrawal threshold to mechanical
stimuli was assessed using von Frey filaments and cold allodynia
was measured by an acetone spray test, as previously described.
According to the comparison of the behavioral test
results (both mechanical allodynia and cold allodynia tests) prior
to and following surgery, the rats were divided into two groups:
The pain and control group. In the pain group, the paw withdrawal
thresholds decreased and cold allodynia points rose following
surgery, while the two results did not significantly change
following surgery in the control group.
At the completion of behavioral testing, each group
(n=3) was euthanized using CO2. The flow rate for
CO2 euthanasia displaced 20% of the chamber volume/min
according to the 2013 edition of the American Veterinary Medical
Association Guidelines for the Euthanasia of Animals. The following
tissues were collected from these rats for microRNA analysis: DHSC,
T, AC and B.
miRNA microarray
Total RNA was isolated using an miRNeasy mini kit
and RNase-Free DNase set (both Qiagen, Inc., Valencia, CA, USA).
RNA quality was checked with denaturing agarose gel (1.5%)
electrophoresis and nucleic acid staining. Samples with 28S and 18S
rRNA bands were resolved into two discrete bands that had no
significant smearing below each band, and the 28S rRNA band
intensity, which was approximately twice that of the 18S rRNA band,
was used for subsequent procedures. Total RNA (1 µg) was
labeled with Affymetrix® FlashTag™, Biotin HSR RNA
Labeling kits (Affymetrix, Inc., Santa Clara, CA, USA). Next,
samples were hybridized to a GeneChip® miRNA 4.0 array
(Affymetrix, Inc.) at 60 rpm, at 48°C for 16 h. Fluorescent images
of microarray slides were scanned using a GeneChip®
Scanner 3000 7G (Affymetrix, Inc.). All CEL files from the miRNA
4.0 chips were normalized using Expression Console software
(version 1.3.1; Affymetrix, Inc.). Affymetrix miRNA arrays are
designed to contain all miRNAs in miRBase release 20 (http://tools.thermofisher.com/content/sfs/brochures/miRNA_4-0_and_4-1_datasheet.pdf).
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) assay
All procedures were performed according to the
corresponding manufacturer's protocols. Expression of the miRNAs
was examined with the SYBR-Green RT-PCR kit (Invitrogen; Thermo
Fisher Scientific, Inc., Waltham, MA, USA). Results were normalized
to U6 expression. RT was performed with 1 mg total RNA using the
ThermoScript RT-PCR system (Invitrogen; Thermo Fisher Scientific.
Inc.) for first-strand complementary DNA (cDNA) synthesis. For
qPCR, cDNA was amplified using the MyiQ Real-Time PCR Detection
system (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Quantitative cycle values were obtained from each amplification
curve using iQ5 Optical System Software (version 2.1) provided by
the manufacturer (Bio-Rad Laboratories, Inc.). The
2−ΔΔct method was used (21). Primer sequences will be provided
upon request. The primers used were as follows: rno-miR-3573-5p
forward, 5′-ACACTCCAGCTGGGTGAGGGGCAGTGATAGAAAGGA-3′ and reverese,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTC AGTTGAGTCCTTTC-3′; rno-miR-3074
forward, 5′-ACACTCCAGCTGGGGATATCAGCTCAGTAGGCACCG-3′ and reverse,
CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGCGGTGCC-3′; rno-miR-25-5p
forward, 5′-ACACTCCAGCTGGGAGGCGGAGACACGGGCAATTGC-3′ and reverse,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGGCAATTGC-3′; rno-miR-702-3p
forward, 5′-ACACTCCAGCTGGGTGCCCACCCTTTACCCCACTCCA-3′ and reverse,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGTGGAGTG-3′; rno-miR-30c-1-3p
forward, 5′-ACACTCCAGCTGGGCTGGGAGAGGGTTGTTTACTCC-3′ and reverse,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGGGAGTAAA-3′; rno-miR-93-3p
forward, 5′-ACA CTCCAGCTGGGACTGCTGAGCTAGCACTTCCCGA-3′ and reverse,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGTCGGGAAG-3′; rno-miR-873-5p
forward, 5′-ACACTCCAGCTGGGGCAGGAACTTGTGAGTCTCCT-3′ and reverse,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGAGGAGAC-3′; rno-miR-455-3p
forward, 5′-ACACTCCAGCTGGGGCAGTCCACGGGCATATA-3′ and reverse,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGAGTGTATA-3′; rno-miR-32-3p
forward, 5′-ACACTCCAGCTGGGTGGACGGAGAACTGAT-3′ and reverse, 5′-CTCA
ACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGACCCTTAT-3′; rno-miR-184 forward,
5′-ACACTCCAGCTGGGGCAATTTAGTGTGTGTGA-3′ and reverse,
5′-CTCAACTGGTGTCGTGGAGTCGGCAATTCAGTTGAGAATATCAC-3′; universal miR
reverse, 5′-TGTCGTGGAGTCGGCAATTC-3′; U6 forward,
5′-CTCGCTTCGGCAGCACATA-3′ and reverse,
5′-ATATGGAACGCTTCACGAATTTGC-3′. The thermocycling conditions were
as follows: stage 1 (1 cycle), pre-degeneration at 95°C for 5 sec;
stage 2 (40 cycles), denaturation at 95°C for 5 sec and primer
annealing at 61°C for 30 sec; stage 3 (1 cycle), sufficient
extention at 72°C for 1 min and cooling down at 4-10°C.
Bioinformatic evaluation
For the differential regulation miRNAs, the
predicted, but not (yet) verified mRNA targets were investigated.
Since hundreds were found for each, Gene Ontology (GO) analysis
(http://www.geneontology.org/page/go-database) was
performed using the GO enrichment analysis software tool kit
(22,23) and pathway analysis was performed
using Kyoto Encyclopedia of Genes and Genomes (KEGG)
pathway/brite/module mapping tools (24) (http://www.kegg.jp/kegg/pathway.html#mapping) to
define which gene molecular function terms were enriched that
characterize the collection of target genes in comparison with the
genome in general.
Statistical analysis
Due to the small sample size (n=3), the random
variance model t-test was adopted to filter the differentially
expressed microRNAs between the control and pain groups using
GraphPad 5.0. Following the significance analysis and false
discovery rate analysis, differentially expressed genes were
selected according to their P-values. P<0.05 was considered to
indicate a statistically significant difference.
Results
miRNA microarray
To functionally investigate a possible link between
miRNA expression and the post-brachial plexus injury neuropathic
pain, the differential expression of miRNAs between groups in 4
different tissues, DHSC, T, AC and B, was analyzed. The expression
of 728 miRNAs was detected in each tissue and divided into three
types according to the changes: Decrease, increase and no change.
The statistical results of the microRNA array analysis of the 4
different tissues are shown in Table
I (P<0.05; n=3). The statistical analysis software used was
GraphPad 5.0.
 | Table ImiRNAs with statistical differences
in expression between the pain and control groups. |
Table I
miRNAs with statistical differences
in expression between the pain and control groups.
| miRNA | P-value | FDR | Fold-change | Regulation
trend | Sequence length,
nt | Sequence
(5′-3′) |
|---|
| DHSC group | | | | | | |
| miR-3573-5p | 0.0068 | 1.000 | 0.70 | Down | 22 |
UGAGGGGCAGUGAUAGAAAGGA |
| miR-3074 | 0.041413 | 1.000 | 0.90 | Down | 22 |
GAUAUCAGCUCAGUAGGCACCG |
| miR-1193-3p | 0.025148 | 1.000 | 1.23 | Up | 22 |
UAGGUCACCCGUUUUACUAUCC |
| miR-410-5p | 0.040567 | 1.000 | 1.23 | Up | 21 |
AGGUUGUCUGUGAUGAGUUCG |
| miR-340-5p | 0.018692 | 1.000 | 1.48 | Up | 22 |
UUAUAAAGCAAUGAGACUGAUU |
| AC group | | | | | | |
| miR-208a-3p | 0.018404 | 0.997 | 0.37 | Down | 18 |
AUAAGACGAGCAAAAAGC |
| miR-6216 | 0.028229 | 0.997 | 0.71 | Down | 23 |
GAUACACAGAGGCAGGAGGAGAA |
| miR-3580-3p | 0.034532 | 0.997 | 0.72 | Down | 22 |
UGACUAGGGUAGUAUGAGUAGA |
| miR-205 | 0.045209 | 0.997 | 1.41 | Up | 23 |
UCCUUCAUUCCACCGGAGUCUGU |
| miR-25-5p | 0.030307 | 0.997 | 1.46 | Up | 22 |
AGGCGGAGACACGGGCAAUUGC |
| miR-702-3p | 0.035705 | 0.997 | 1.49 | Up | 23 |
UGCCCACCCUUUACCCCACUCCA |
| miR-501-3p | 0.024475 | 0.997 | 1.65 | Up | 23 |
AAUGCACCCGGGCAAGGAUUUGG |
| let-7f-5p | 0.035378 | 0.997 | 1.70 | Up | 22 |
UGAGGUAGUAGAUUGUAUAGUU |
| miR-381-5p | 0.032536 | 0.997 | 1.80 | Up | 22 |
AGCGAGGUUGCCCUUUGUAUAU |
| miR-30c-1-3p | 0.011214 | 0.997 | 1.87 | Up | 22 |
CUGGGAGAGGGUUGUUUACUCC |
| miR-671 | 0.028852 | 0.997 | 1.87 | Up | 21 |
UCCGGUUCUCAGGGCUCCACC |
| miR-184 | 0.006879 | 0.997 | 2.42 | Up | 22 |
UGGACGGAGAACUGAUAAGGGU |
| T group | | | | | | |
| miR-34b-5p | 0.04851 | 0.971 | 0.62 | Down | 23 |
AGGCAGUGUAAUUAGCUGAUUGU |
| miR-181d-3p | 0.0342 | 0.971 | 0.65 | Down | 20 |
CCACCGGGGGAUGAAUGUCA |
| miR-484 | 0.027441 | 0.971 | 0.66 | Down | 22 |
UCAGGCUCAGUCCCCUCCCGAU |
| miR-370-5p | 0.016922 | 0.971 | 0.70 | Down | 24 |
CAGGUCACGUCUCUGCAGUUACAC |
| miR-9b-5p | 0.008269 | 0.971 | 0.78 | Down | 19 |
UUCGGUUAUCUAGCUUUAU |
| miR-1912-3p | 0.017277 | 0.971 | 0.79 | Down | 19 |
CACAGAACAUGCAGUGAGA |
| miR-759 | 0.018461 | 0.971 | 0.81 | Down | 22 |
GCAGAGUGCAAACAAUUUUGAC |
| miR-463-5p | 0.036635 | 0.971 | 0.81 | Down | 21 |
UACCUAAUUUGUUGUCCAUCA |
| miR-193-3p | 0.045474 | 0.971 | 0.81 | Down | 22 |
AACUGGCCUACAAAGUCCCAGU |
| miR-802-5p | 0.040079 | 0.971 | 0.83 | Down | 21 |
UCAGUAACAAAGAUUCAUCCU |
| miR-218a-5p | 0.019683 | 0.971 | 0.84 | Down | 21 |
UUGUGCUUGAUCUAACCAUGU |
| miR-31b | 0.031174 | 0.971 | 0.84 | Down | 19 |
CUAUGCCAGCAUCUUGCCU |
| miR-3593-5p | 0.033797 | 0.971 | 0.87 | Down | 22 |
UGGCCUCCGCAGGGUUGAAGCU |
| miR-3570 | 0.039038 | 0.971 | 1.15 | Up | 22 |
GGUACAAUCAACGGUCGAUGGU |
| miR-3588 | 0.018922 | 0.971 | 1.19 | Up | 22 |
UCACAAGUUAGGGUCUCAGGGA |
| miR-664-3p | 0.039461 | 0.971 | 1.30 | Up | 22 |
UAUUCAUUUACUCCCCAGCCUA |
| miR-488-3p | 0.025485 | 0.971 | 1.41 | Up | 21 |
UUGAAAGGCUGUUUCUUGGUC |
| miR-30c-1-3p | 0.009129 | 0.971 | 2.03 | Up | 22 |
CUGGGAGAGGGUUGUUUACUCC |
| miR-106b-3p | 0.018801 | 0.971 | 2.08 | Up | 22 |
CCGCACUGUGGGUACUUGCUGC |
| miR-93-3p | 0.012217 | 0.971 | 2.69 | Up | 23 |
ACUGCUGAGCUAGCACUUCCCGA |
| miR-28-5p | 0.041407 | 0.971 | 2.85 | Up | 22 |
AAGGAGCUCACAGUCUAUUGAG |
| miR-873-5p | 0.02098 | 0.971 | 2.93 | Up | 21 |
GCAGGAACUUGUGAGUCUCCU |
| B group | | | | | | |
| miR-455-3p | 0.028562 | 0.997 | 0.32 | Down | 22 |
GCAGUCCACGGGCAUAUACACU |
| miR-32-3p | 0.030898 | 0.997 | 0.47 | Down | 22 |
GCAAUUUAGUGUGUGUGAUAUU |
| miR-466b-2-3p | 0.032782 | 0.997 | 0.57 | Down | 21 |
AUAUACAUACACACAUACACA |
| miR-702-3p | 0.035054 | 0.997 | 0.62 | Down | 23 |
UGCCCACCCUUUACCCCACUCCA |
| miR-742-5p | 0.035279 | 0.997 | 0.72 | Down | 21 |
UACUCACAUGGUUGCUAAUCA |
| miR-195-3p | 0.021132 | 0.997 | 0.73 | Down | 23 |
CCAAUAUUGGCUGUGCUGCUCCA |
| miR-509-3p | 0.045567 | 0.997 | 0.77 | Down | 22 |
UGAUUGACAUGUCUGCAGUGGA |
In the DHSC group, 332 miRNAs exhibited decreased
expression, 378 miRNAs exhibited increased expression and 18 miRNAs
exhibited no change. The statistical analysis showed that 2 miRNAs
were downregulated and 3 miRNAs were upregulated significantly.
In the T group, 305 miRNAs exhibited decreased
expression, 409 miRNAs exhibited increased expression and 14 miRNAs
exhibited no change. The statistical analysis showed that 13 miRNAs
were downregulated and 9 miRNAs were upregulated significantly.
In the AC group, 348 miRNAs exhibited decreased
expression, 359 miRNAs exhibited increased expression and 21 miRNAs
exhibited no change. The statistical analysis showed that 3 miRNAs
were downregulated and 9 miRNAs were upregulated significantly.
In the B group, 362 miRNAs exhibited decreased
expression, 352 miRNAs exhibited increased expression and 14 miRNAs
exhibited no change. The statistical analysis showed that 7 miRNAs
were downregulated and no miRNAs were upregulated
significantly.
PCR verification
To validate the microarray results, RT-qPCR was
performed for these differentially regulated (diff-reg) miRNAs in
the 4 groups. It was found that the relative expression of 10
miRNAs among them were significantly altered, which coincided with
the results of the microarray. The 10 miRNAs were miR-3573-5p
(DHSC), miR-3074 (DHSC), miR-30c-1-3p (AC and T), miR-702-3p (AC),
miR-184 (AC), miR-25-5p (AC), miR-873-5p (T), miR-93-3p (T),
miR-455-3p (B) and miR-32-3p (B). miR-30c-1-3p with a significant
change appeared in both the AC and T groups. The other miRNAs could
not be verified by PCR. The statistical analysis showed that
miR-3573-5p (DHSC), miR-3074 (DHSC), miR-455-3p (B) and miR-32-3p
(B) were downregulated, and that miR-30c-1-3p (AC and T),
miR-702-3p (AC), miR-184 (AC), miR-25-5p (AC), miR-873-5p (T) and
miR-93-3p (T) were upregulated significantly (Table II).
| Table IImiRNAs with differential expression
by PCR verification between pain and control groups. |
Table II
miRNAs with differential expression
by PCR verification between pain and control groups.
| miRNA | P-value | FDR | Fold-change | Regulation
trend | Sequence length,
nt | Sequence
(5′-3′) |
|---|
| DHSC group | | | | | | |
|
rno-miR-3573-5p | 0.0067996 | 1.000 | 0.70 | Down | 22 |
UGAGGGGCAGUGAUAGAAAGGA |
| rno-miR-3074 | 0.0414127 | 1.000 | 0.90 | Down | 22 |
GAUAUCAGCUCAGUAGGCACCG |
| AC group | | | | | | |
| rno-miR-25-5p | 0.0303073 | 0.997 | 1.46 | Up | 22 |
AGGCGGAGACACGGGCAAUUGC |
|
rno-miR-702-3p | 0.0357045 | 0.997 | 1.49 | Up | 23 |
UGCCCACCCUUUACCCCACUCCA |
|
rno-miR-30c-1-3p | 0.0112141 | 0.997 | 1.87 | Up | 22 |
CUGGGAGAGGGUUGUUUACUCC |
| rno-miR-184 | 0.0068785 | 0.997 | 2.42 | Up | 22 |
UGGACGGAGAACUGAUAAGGGU |
| T group | | | | | | |
|
rno-miR-30c-1-3p | 0.0091288 | 0.971 | 2.03 | Up | 22 |
CUGGGAGAGGGUUGUUUACUCC |
| rno-miR-93-3p | 0.0122168 | 0.971 | 2.69 | Up | 23 |
ACUGCUGAGCUAGCACUUCCCGA |
|
rno-miR-873-5p | 0.02098 | 0.971 | 2.93 | Up | 21 |
GCAGGAACUUGUGAGUCUCCU |
| B group | | | | | | |
|
rno-miR-455-3p | 0.0285622 | 0.997 | 0.32 | Down | 22 |
GCAGUCCACGGGCAUAUACACU |
| rno-miR-32-3p | 0.0308978 | 0.997 | 0.47 | Down | 22 |
GCAAUUUAGUGUGUGUGAUAUU |
Bioinformatics analysis of the diff-reg
miRNAs
For these diff-reg miRNAs with PCR verification, the
predicted, but not (yet) verified, mRNA targets were
investigated.
In the DHSC group, there were 27 intersection genes
(Table III), which were
involved in neuropathic pain according to GO and pathway analyses.
GO analysis showed that the downregulation function of miR-3573-5p
and miR-3074 associated with neuropathic pain included 'axon
guidance', 'synaptic transmission', 'synapse maturation',
'excitatory post-synaptic membrane potential', 'neuron apoptotic
process', 'macrophage activation involved in immune response',
'cell chemotaxis', 'neuron migration' and 'neuron differentiation'.
The target genes of miR-3573-5p and miR-3074 took part in several
pathways, including the 'calcium signaling pathway', 'cholinergic
synapse', 'GABAergic synapse', 'glutamatergic synapse', the 'HIF-1
signaling pathway', the 'MAPK signaling pathway', the 'mTOR
signaling pathway', the 'notch signaling pathway', the 'PI3K-Akt
signaling pathway', 'synaptic vesicle cycle' and the 'Wnt signaling
pathway'. Downregulation of miR-3573-5p and miR-3074 would result
in upregulation of the target genes, which inhibited neuron
apoptotic process, promoted long-term potentiation and synapse
plasticity, and strengthened cell proliferation and
differentiation.
| Table IIIIntersection genes involved in
neuropathic pain in GO and pathway analyses. |
Table III
Intersection genes involved in
neuropathic pain in GO and pathway analyses.
| Group | Genes |
|---|
| DHSC | Grin1, Grm4, Grm6,
Itga5, Notch1, P2rx1, Prkcg, Shank1, Peg12, Shank3, Srf, Stx1b,
Stxbp1, Syk, Vegfa, Vhl, Wnt8b, Wif1, Adra1b, Cacna1a, Cd38, Chat,
Ddit4, Efna2, Fgfr3, Gad1, Gng11 |
| AC | Camk2b, Csf1,
Hdac1, Kdr, Mapk3, Prkcg, Tgfa, Vegfa |
| T | Camk2b, Itgb1,
Prkcg |
| B | Scn1a, Kdr,
Nos3 |
In the T group, there were 3 intersection genes,
which were involved in neuropathic pain according to GO and pathway
analyses (Table III). GO
analysis showed that the upregulation function of miR-30c-1-3p,
miR-873-5p and miR-93-3p associated with neuropathic pain included
'neurotransmitter uptake', 'neuron apoptotic process', 'sensory
perception of pain', 'long-term memory' and 'neuron projection
development'. The target genes of miR-30c-1-3p, miR-873-5p and
miR-93-3p took part in several pathways, including the 'calcium
signaling pathway', 'cholinergic synapse', 'dopaminergic synapse',
the 'ErbB signaling pathway', 'GABAergic synapse', 'glutamatergic
synapse', the 'hedgehog signaling pathway', the 'HIF-1 signaling
pathway', 'long-term potentiation', the 'MAPK signaling pathway',
the 'mTOR signaling pathway', the 'neurotrophin signaling pathway',
the 'notch signaling pathway', the 'PI3K-Akt signaling pathway',
'retrograde endocannabinoid signaling', 'serotonergic synapse' and
the 'Wnt signaling pathway'. Upregulation of these miRNAs would
result in downregulation of the target genes, which promoted neuron
apoptotic apoptosis, reduced long-term potentiation and synapse
plasticity, and inhibited cell proliferation and
differentiation.
In the AC group, there were 8 intersection genes,
which were involved in neuropathic pain according to GO and pathway
analyses (Table III). GO
analysis indicated that the upregulation function of miR-30c-1-3p,
miR-702-3p, miR-184 and miR-25-5p associated with neuropathic pain
included 'neuron apoptotic process', 'regulation of neuronal
synaptic plasticity', 'axon guidance', 'synaptic transmission
(glutamatergic)', 'axon extension', 'axonogenesis', 'long-term
synaptic potentiation', 'sensory perception of pain', 'response to
cold', 'response to stress', 'neuron differentiation', 'neuron
migration', 'neuron cell-cell adhesion' and 'long-term memory'. The
target genes of miR-30c-1-3p, miR-702-3p, miR-184 and miR-25-5p
took part in several pathways, including the 'calcium signaling
pathway', 'cholinergic synapse', the 'ErbB signaling pathway',
'GABAergic synapse', 'glutamatergic synapse', the 'hedgehog
signaling pathway', the 'HIF-1 signaling pathway', 'long-term
depression', the 'mTOR signaling pathway', the 'MAPK signaling
pathway', 'dopaminergic synapse', 'long-term potentiation',
'serotonergic synapse', the 'neurotrophin signaling pathway', the
'notch signaling pathway', the 'PI3K-Akt signaling pathway',
'retrograde endocannabinoid signaling' and the 'Wnt signaling
pathway'. Upregulation of these miRNAs would result in
downregulation of the target genes, which promoted neuron apoptotic
apoptosis, reduced long-term potentiation and synapse plasticity,
and inhibited cell proliferation and differentiation.
In the B group, there were 3 intersection genes,
which were involved in neuropathic pain according to GO and pathway
analyses (Table III). GO
analysis indicated that the downregulation function of miR-455-3p
and miR-32-3p associated with neuropathic pain included 'neuron
projection morphogenesis', 'cell migration', 'cell proliferation'
and 'apoptotic process'. The target genes of miR-455-3p and
miR-32-3p took part in four pathways: The 'calcium signaling
pathway', the 'HIF-1 signaling pathway', the 'PI3K-Akt signaling
pathway' and the 'VEGF signaling pathway'. Downregulation of
miR-455-3p and miR-32-3p would induce upregulation of the target
genes, which inhibited cell apoptosis, and promoted
vascularization, cell migration and proliferation.
In the post-brachial plexus injury neuropathic pain
model, the central nerve groups (AC and T) and the peripheral nerve
group (DHSC) exhibited contrasting effects on synapse plasticity
and neuron apoptosis. The former showed downregulation of synapse
plasticity, cell proliferation and differentiation, while the later
showed upregulation.
miR-30c-1-3p with a significant change appeared in
the AC and T groups, which significantly increased in the pain
model. The functions of miR-30c-1-3p associated with neuropathic
pain included the 'calcium signaling pathway', 'cholinergic
synapse', 'dopaminergic synapse', the 'HIF-1 signaling pathway',
the 'ErbB signaling pathway', 'long-term potentiation', the
'neurotrophin signaling pathway', the 'Wnt signaling pathway',
'GABAergic synapse', 'glutamatergic synapse', the 'MAPK signaling
pathway', the 'mTOR signaling pathway', 'retrograde endocannabinoid
signaling', 'serotonergic synapse' and 'long-term depression'. The
predicted genes of miR-30c-1-3p were calcium/calmodulin dependent
protein kinase IIβ (Camk2b) and protein kinase Cγ (Prkcg), which
were validated by PCR to exhibit significant downregulation in the
AC and T groups. Camk2b and Prkcg took part in the 'calcium
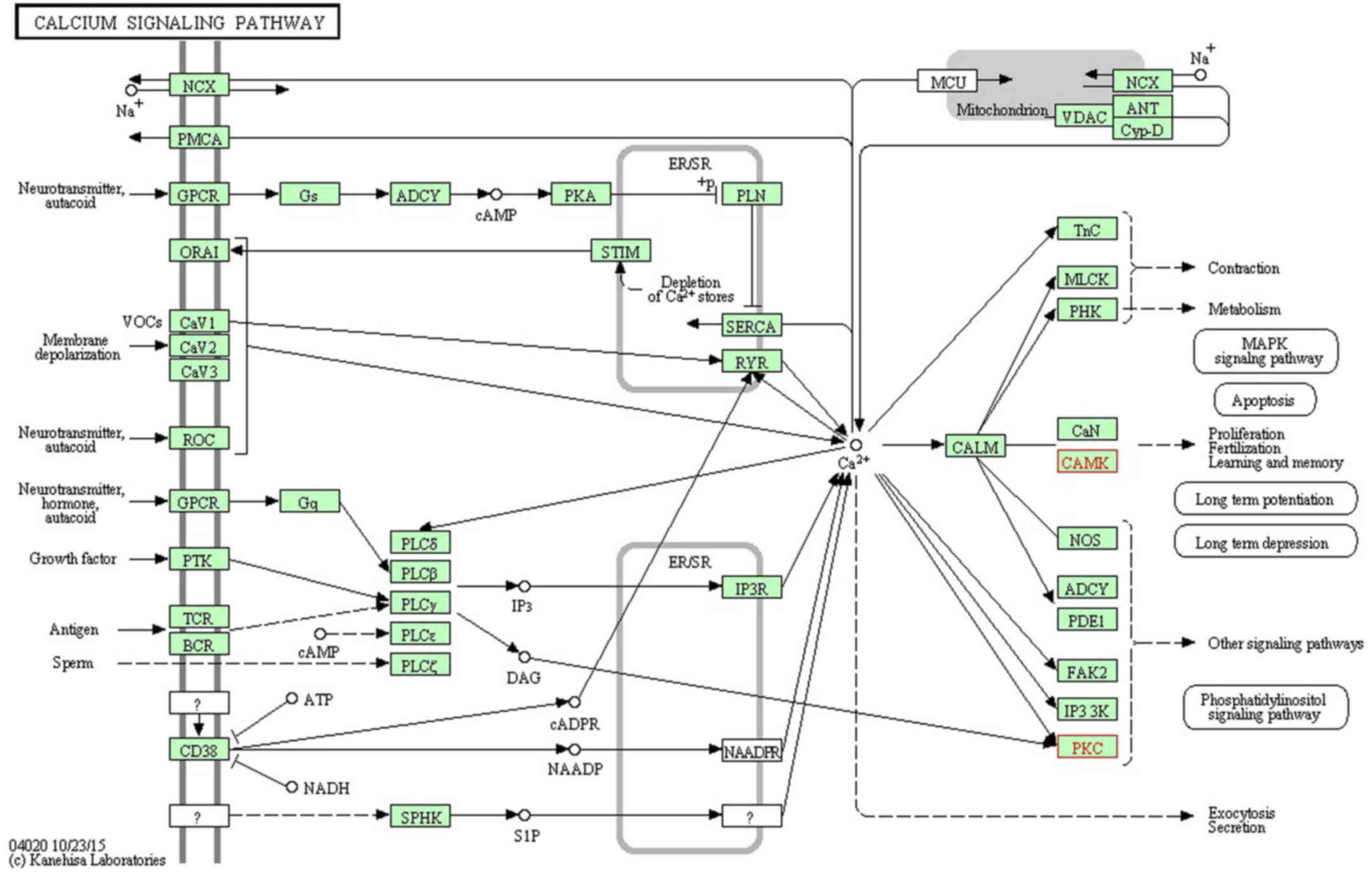
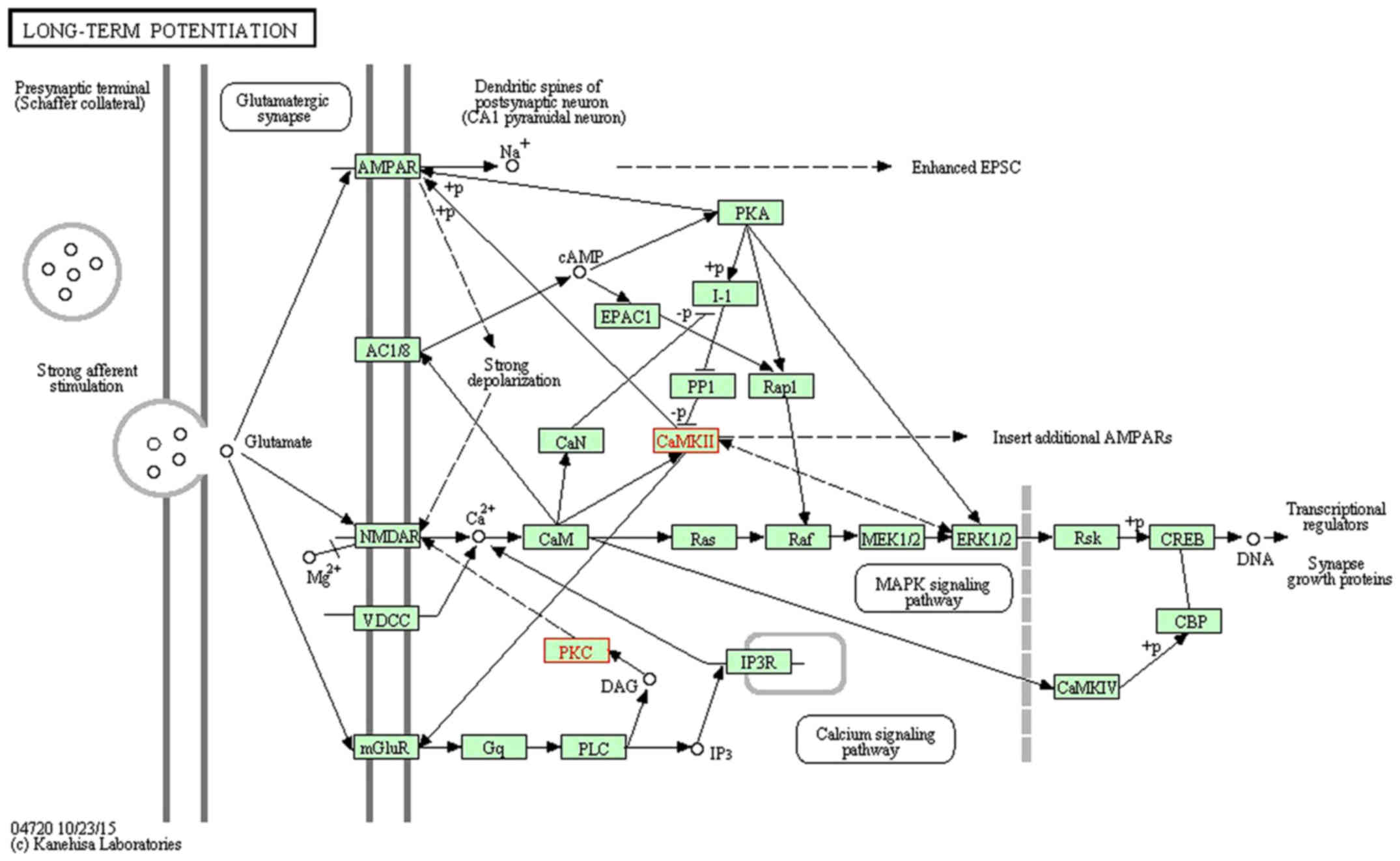
signaling pathway' (www.kegg.jp/dbget-bin/www_bget?map04020; Fig. 1), 'long-term potentiation'
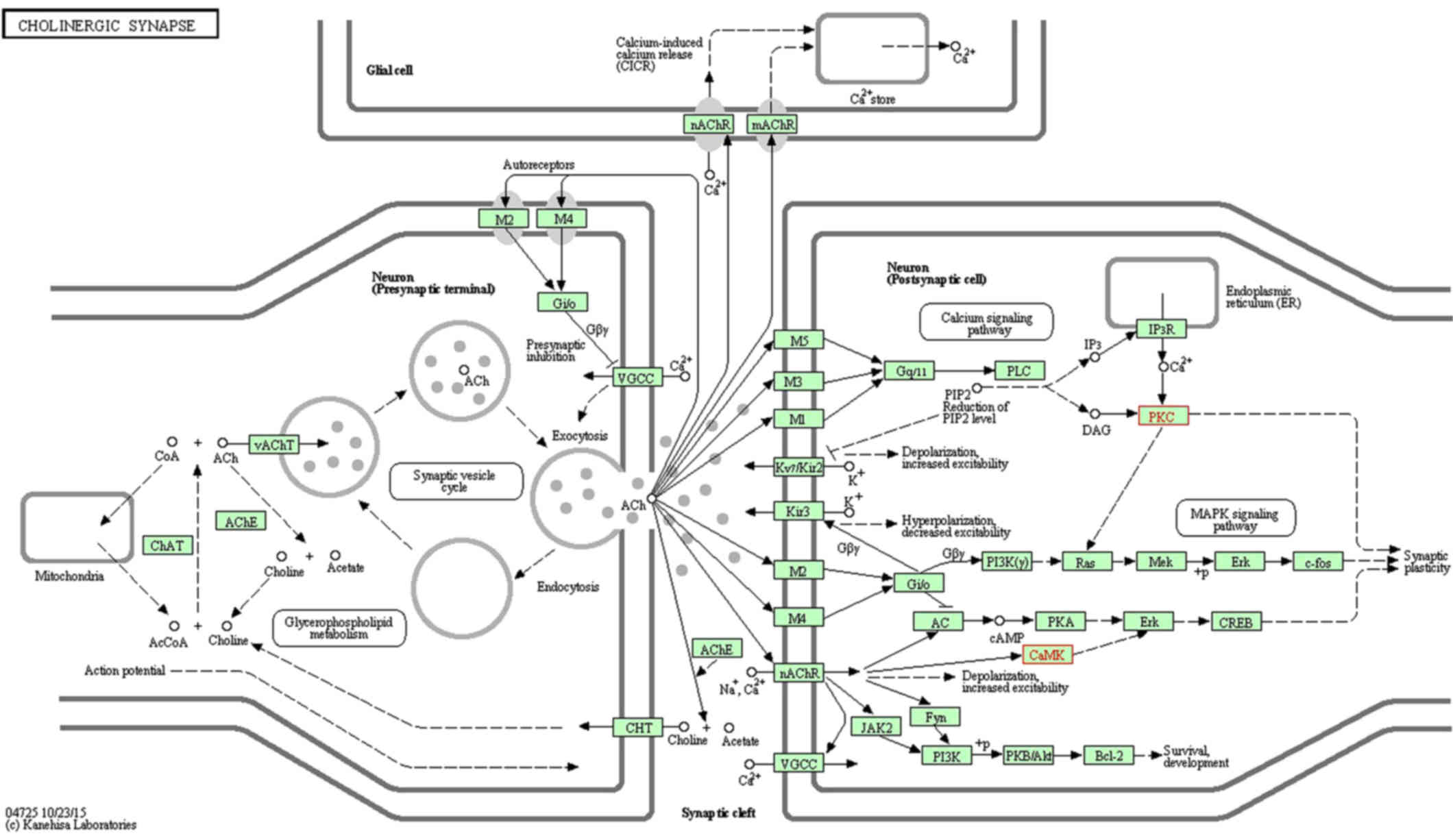
(www.kegg.jp/dbget-bin/www_bget?map04720; Fig. 2) and 'cholinergic synapse'
(www.kegg.jp/dbget-bin/www_bget?map04725; Fig. 3). All figures were obtained using
the KEGG pathway mapping tool in the KEGG database (24–26). The downregulation of Camk2b and
Prkcg would inhibit cell proliferation and synapse growth protein
formation, which made synapse plasticity decrease.
Discussion
It has previously been revealed that descending
facilitatory modulation is a key mechanism underlying the induction
and maintenance of neuropathic pain (27). It is well documented that spinal
nociception is powerfully modulated by an endogenous descending
inhibitory system (27), although
descending control is bi-directional via inhibitory and
facilitatory systems. When the brachial plexus was avulsed, the
peripheral nerves were separated from the DHSC, which resulted in
blockage of the descending inhibitory system. Sustained activation
of descending facilitation increased the excitability of spinal
nociceptive neurons and promoted long-term potentiation and synapse
plasticity, which in turn evoked neuroplasticity of certain
supraspinal structures, including the anterior cingulate cortex
(ACC) and rostral ventral medulla (8–30),
which underly certain states of neuropathic pain.
The roles of the ACC and T in pain conditions have
been consistently demonstrated for the past several decades
(31,32). Numerous animal studies have been
performed to identify the T and ACC as potential mediators of the
pain experience, which are not only involved in the transmission of
pain sensation, but also serve a role in processing pain-related
emotions (27). The ACC and T
widely connect with the descending modulation system.
However, the miRNAs within the ACC and T that
mediated the perception of nociceptive signals following brachial
plexus injury remain poorly understood. In the present study, 4
miRNAs in the ACC and 3 miRNAs in the T exhibited significant
upregulation, and were associated with neuropathic pain. The
functions of the miRNAs in the ACC and T were synergetic in the
aspect of positive regulation of neuron apoptotic process, and
inhibition of long-term potentiation and synapse plasticity.
Previous studies reported that peripheral injury induced long-term
potentiation of excitatory synaptic responses in the ACC neurons
(33,34).
miR-30c-1-3p and its predicted genes (Camk2b and
Prkcg) existed in the AC and T groups, with significant changes, in
the present study. Waggener et al (35) reported that calmodulin-dependent
protein kinase II regulated oligodendrocyte maturation and central
nervous system myelination. Fang et al (36) reported that Camk2b protected
neurons from homocysteine-induced apoptosis with the involvement of
the hypoxia-inducible factor 1α signal pathway. Miletic et
al (37) demonstrated that
Prkcg mediated the phosphorylation of glutamate receptor 1 in the
postsynaptic density of spinal dorsal horn neurons, accompanied
with neuropathic pain. Yeh et al (38) observed the expression of dorsal
spinal protein kinase C γ-subunit and other pain-related molecules
in the spinal area in neuropathic pain model animals. Together,
these studies indicated that miR-30c-1-3p may serve an extremely
important role in neuropathic pain.
Besides central sensitization, peripheral
sensitization is also associated with neuropathic pain following
brachial plexus avulsion, which may result from ectopic discharge
(39), scar stimulation (40) and excitory conduction short
circuit due to the lack of myelin sheaths, among others (41). The concrete mechanisms of these
causes include five aspects: Neuronal apoptosis, axonal
regeneration, immune cell infiltration, glial cell accumulation and
chemokine transfer (42). The
present study indicated that 2 microRNAs in the DHSC group and 2
microRNAs in the B group exhibited significant down-regulation in
the pain model. GO analysis showed that these microRNAs had
functions in 'axon guidance', 'synaptic transmission', 'synapse
maturation', 'excitatory postsynaptic membrane potential', 'the
neuronal apoptotic process', 'macrophage activation involved in the
immune response', 'cell chemotaxis', 'neuron migration' and 'neuron
differentiation', 'neuron projection morphogenesis', 'cell
migration', 'cell proliferation' and 'apoptosis', which facilitated
the peripheral sensitization and induced neuropathic pain. The
functions of the microRNAs in the DHSC and B groups were synergetic
in the aspect of inhibition of cell apoptosis and promotion of cell
proliferation, which contrasted with the functions of the
differentially expressed miRNAs in the AC and T groups. When
sustained activation of descending facilitation system occurred, a
number of neurotrophic factors and neurotransmitters were exhausted
and synapse growth proteins decreased, which resulted in neuron
apoptosis increasing and synapse plasticity decreasing in the AC
and T.
There were 10 miRNAs with significant expressional
changes in the pain group, namely miR-3573-5p (DHSC), miR-3074
(DHSC), miR-30c-1-3p (AC and T), miR-702-3p (AC), miR-184 (AC),
miR-25-5p (AC), miR-873-5p (T), miR-93-3p (T), miR-455-3p (B) and
miR-32-3p (B). Previous studies on the aforementioned miRNAs
associated with neuropathic pain have included a study by Lu et
al (43), which found that
miR-702-3p was differentially expressed in rat cortex in the
anesthetic treatment group, which indicated the miRNA may be
associated with neuropathic pain. McAdams et al (44) reported that morphine decreased the
expression of miR-455-3p in the hippocampus of stressed neonatal
mice and that the miRNA was involved in neurodevelopment,
neurotransmission and inflammation. Gong et al (45) investigated the differentially
expressed miRNAs in the lumbar spinal dorsal horn of mice with
streptozotocin-induced diabetic neuropathic pain (DNP) and found
that aberrant expression of miR-184 may contribute to the
pathogenesis of DNP, and was a potential target for therapeutic
interventions following DNP. McKiernan et al (46) identified miR-184 as a novel
contributor to neuronal survival following mild and severe
seizures. Furthermore, Liu et al (47) found that high levels of miR-184
promoted proliferation, but inhibited differentiation of adult
neural stem/progenitor cells, and that the miRNA regulated the
expression of numblike, a known regulator of brain development.
In future research, the predicted genes (Camk2b and
Prkcg) should be verified as the targets of miR-30c-1-3p by
cytology. miR-30c-1-3p could be transfected into neuron cells
through plasmids and then the amount of target proteins could be
measured by PCR and western blot analysis. If Camk2b and Prkcg were
validated as the targets of miR-30c-1-3p, an attempt would be made
to inject an miR-30c-1-3p-expressing virus into the ACC of the rat
and then behavioral tests would be performed to determine whether
the rat was in pain or not.
In conclusion, in the present study, post-CBPA pain
was regulated intricately by miRNAs from different tissues in
different pathways. There were 10 miRNAs with significant changes
in expression in 4 different tissues in the post-BPA pain model.
The central nerve groups (AC and T) and the peripheral nerve groups
(DHSC) exhibited contrasting effects on synapse plasticity and
neuron apoptosis. miR-30c-1-3p and its predicted genes (Camk2b and
Prkcg) existed in the AC and T groups with significant espressional
changes.
Acknowledgments
The authors would like to thank Dr Jiali Li (Fudan
University) for providing assistance with the data analysis. The
present study was sponsored by the Ministry of Science and
Technology of China (973 program; grant no. 2014CB542204), the
Natural Science Foundation of Shanghai (grant no. STCSM
16ZR1404200) and the National Natural Science Foundation of China
(grant no. NSFC 81572127).
References
|
1
|
Mendell JR and Sahenk Z: Clinical
practice. Painful sensory neuropathy. N Engl J Med. 348:1243–1255.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Sawynok J: Topical and peripherally acting
analgesics. Pharmacol Rev. 55:1–20. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Hansson PT and Dickenson AH:
Pharmacological treatment of peripheral neuropathic pain conditions
based on shared commonalities despite multiple etiologies. Pain.
113:251–254. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Ciaramitaro P, Padua L, Devigili G, Rota
E, Tamburin S, Eleopra R, Cruccu G and Truini A; Neuropathic pain
special interest group of the Italian Neurological Society:
Prevalence of neuropathic pain in patients with traumatic brachial
plexus injury: A multicenter prospective hospital-based study. Pain
Med. March 3–2017.Epub ahead of print. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Anand P and Birch R: Restoration of
sensory function and lack of long-term chronic pain syndromes after
brachial plexus injury in human neonates. Brain. 125:113–122. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Berman JS, Birch R and Anand P: Pain
following human brachial plexus injury with spinal cord root
avulsion and the effect of surgery. Pain. 75:199–207. 1998.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Baruah S, Devi BI, Bhat DI and Shukla D:
Drezotomy in the management of post brachial plexus injury
neuropathic pain: Preliminary results. Indian J Neurotrauma.
11:27–29. 2014. View Article : Google Scholar
|
|
8
|
Aley KO and Levine JD: Different
peripheral mechanisms mediate enhanced nociception in
metabolic/toxic and traumatic painful peripheral neuropathies in
the rat. Neuroscience. 111:389–397. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Erichsen HK and Blackburn-Munro G:
Pharmacological characterisation of the spared nerve injury model
of neuropathic pain. Pain. 98:151–161. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Aldrich BT, Frakes EP, Kasuya J, Hammond
DL and Kitamoto T: Changes in expression of sensory organ-specific
microRNAs in rat dorsal root ganglia in association with mechanical
hypersensitivity induced by spinal nerve ligation. Neuroscience.
164:711–723. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Bhalala OG, Srikanth M and Kessler JA: The
emerging roles of microRNAs in CNS injuries. Nat Rev Neurol.
9:328–339. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Erson AE and Petty EM: MicroRNAs in
development and disease. Clin Genet. 74:296–306. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Tan PH, Pao YY, Cheng JK, Hung KC and Liu
CC: MicroRNA-based therapy in pain medicine: Current progress and
future prospects. Acta Anaesthesiol Taiwan. 51:171–176. 2013.
View Article : Google Scholar
|
|
14
|
Parry CB: Pain in avulsion lesions of the
brachial plexus. Pain. 9:41–53. 1980. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Narakas AO: The effects on pain of
reconstructive neurosurgery in 160 patients with traction and/or
crush injury to the brachial plexus. Phantom and Stump Pain.
Siegfried J and Zimmerman M: 1st edition. Springer-Verlag; Berlin:
pp. 126–147. 1981, View Article : Google Scholar
|
|
16
|
Bruxelle J, Travers V and Thiebaut JB:
Occurrence and treatment of pain after brachial plexus injury. Clin
Orthop Relat Res. 237:87–95. 1988.
|
|
17
|
von Schack D, Agostino MJ, Murray BS, Li
Y, Reddy PS, Chen J, Choe SE, Strassle BW, Li C, Bates B, et al:
Dynamic changes in the MicroRNA expression profile reveal multiple
regulatory mechanisms in the spinal nerve ligation model of
neuropathic pain. PLoS One. 6:e176702011. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Choi Y, Yoon YW, Na HS, Kim SH and Chung
JM: Behavioral signs of ongoing pain and cold allodynia in a rat
model of neuropathic pain. Pain. 59:369–376. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Li X, Kroin JS, Kc R, Gibson G, Chen D,
Corbett GT, Pahan K, Fayyaz S, Kim JS, van Wijnen AJ, et al:
Altered spinal microRNA-146a and the microRNA-183 cluster
contribute to osteoarthritic pain in knee joints. J Bone Miner Res.
28:2512–2522. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang L, Yuzhou L, Yingjie Z, Jie L and Xin
Z: A new rat model of neuropathic pain: Complete brachial plexus
avulsion. Neurosci Lett. 589:52–56. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2−ΔΔCT method. Methods. 25:402–408. 2001. View Article : Google Scholar
|
|
22
|
Ashburner M, Ball CA, Blake JA, Botstein
D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT,
et al: The Gene Ontology Consortium: Gene ontology: Tool for the
unification of biology. Nat Genet. 25:25–29. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
The Gene Ontology Consortium: Expansion of
the Gene Ontology knowledgebase and resources. Nucleic Acids Res.
45:D331–D338. 2017. View Article : Google Scholar :
|
|
24
|
Kanehisa M and Goto S: KEGG: Kyoto
Encyclopedia of Genes and Genomes. Nucleic Acids Res. 28:27–30.
2000. View Article : Google Scholar
|
|
25
|
Kanehisa M, Furumichi M, Tanabe M, Sato Y
and Morishima K: KEGG: new perspectives on genomes, pathways,
diseases and drugs. Nucleic Acids Res. 45:D353–D361. 2017.
View Article : Google Scholar :
|
|
26
|
Kanehisa M, Sato Y, Kawashima M, Furumichi
M and Tanabe M: KEGG as a reference resource for gene and protein
annotation. Nucleic Acids Res. 44:D457–D462. 2016. View Article : Google Scholar :
|
|
27
|
Zhang L, Zhang Y and Zhao ZQ: Anterior
cingulate cortex contributes to the descending facilitatory
modulation of pain via dorsal reticular nucleus. Eur J Neurosci.
22:1141–1148. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Wei F, Li P and Zhuo M: Loss of synaptic
depression in mammalian anterior cingulate cortex after amputation.
J Neurosci. 19:9346–9354. 1999.PubMed/NCBI
|
|
29
|
Porreca F, Ossipov MH and Gebhart GF:
Chronic pain and medullary descending facilitation. Trends
Neurosci. 25:319–325. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Robinson D, Calejesan AA and Zhuo M:
Long-lasting changes in rostral ventral medulla neuronal activity
after inflammation. J Pain. 3:292–300. 2002. View Article : Google Scholar
|
|
31
|
Vogt BA: Pain and emotion interactions in
subregions of the cingulate gyrus. Nat Rev Neurosci. 6:533–544.
2005. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Zhuo M: Cortical excitation and chronic
pain. Trends Neurosci. 31:199–207. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Wei F and Zhuo M: Potentiation of sensory
responses in the anterior cingulate cortex following digit
amputation in the anaesthetised rat. J Physiol. 532:823–833. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Xu H, Wu LJ, Wang H, Zhang X, Vadakkan KI,
Kim SS, Steenland HW and Zhuo M: Presynaptic and postsynaptic
amplifications of neuropathic pain in the anterior cingulate
cortex. J Neurosci. 28:7445–7453. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Waggener CT, Dupree JL, Elgersma Y and
Fuss B: CaMKIIβ regulates oligodendrocyte maturation and CNS
myelination. J Neurosci. 33:10453–10458. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Fang M, Feng C, Zhao YX and Liu XY: Camk2b
protects neurons from homocysteine-induced apoptosis with the
involvement of HIF-1α signal pathway. Int J Clin Exp Med.
7:1659–1668. 2014.
|
|
37
|
Miletic G, Hermes JL, Bosscher GL, Meier
BM and Miletic V: Protein kinase C gamma-mediated phosphorylation
of GluA1 in the postsynaptic density of spinal dorsal horn neurons
accompanies neuropathic pain, and dephosphorylation by calcineurin
is associated with prolonged analgesia. Pain. 156:2514–2520. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Yeh CY, Chung SC, Tseng FL, Tsai YC and
Liu YC: Biphasic effects of chronic intrathecal gabapentin
administration on the expression of protein kinase C gamma in the
spinal cord of neuropathic pain rats. Acta Anaesthesiol Taiwan.
49:144–148. 2011. View Article : Google Scholar
|
|
39
|
Kim CH, Oh Y, Chung JM and Chung K: The
changes in expression of three subtypes of TTX sensitive sodium
channels in sensory neurons after spinal nerve ligation. Brain Res
Mol Brain Res. 95:153–161. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Ma L, Uchida H, Nagai J, Inoue M, Chun J,
Aoki J and Ueda H: Lysophosphatidic acid-3 receptor-mediated
feed-forward production of lysophosphatidic acid: an initiator of
nerve injury-induced neuropathic pain. Mol Pain. 5:642009.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Campbell JN and Meyer RA: Mechanisms of
neuropathic pain. Neuron. 52:77–92. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Nickel FT, Seifert F, Lanz S and Maihöfner
C: Mechanisms of neuropathic pain. Eur Neuropsychopharmacol.
2:81–91. 2012. View Article : Google Scholar
|
|
43
|
Lu Y, Jian MY, Ouyang YB and Han RQ:
Changes in rat brain microRNA expression profiles following
sevoflurane and propofol anesthesia. Chin Med J (Engl).
128:1510–1515. 2015. View Article : Google Scholar
|
|
44
|
McAdams RM, McPherson RJ, Beyer RP,
Bammler TK, Farin FM and Juul SE: Dose-dependent effects of
morphine exposure on mRNA and microRNA (miR) expression in
hippocampus of stressed neonatal mice. PLoS One. 10:e01230472015.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Gong Q, Lu Z, Huang Q, Ruan L, Chen J,
Liang Y, Wang H, Yue Y and Feng S: Altered microRNAs expression
profiling in mice with diabetic neuropathic pain. Biochem Biophys
Res Commun. 456:615–620. 2015. View Article : Google Scholar
|
|
46
|
McKiernan RC, Jimenez-Mateos EM, Sano T,
Bray I, Stallings RL, Simon RP and Henshall DC: Expression
profiling the microRNA response to epileptic preconditioning
identifies miR-184 as a modulator of seizure-induced neuronal
death. Exp Neurol. 237:346–354. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Liu C, Teng ZQ, Santistevan NJ, Szulwach
KE, Guo W, Jin P and Zhao X: Epigenetic regulation of miR-184 by
MBD1 governs neural stem cell proliferation and differentiation.
Cell Stem Cell. 6:433–444. 2010. View Article : Google Scholar : PubMed/NCBI
|