Introduction
Gastric cancer (GC) is one of the major causes of
malignant tumor-associated mortality worldwide (1). Although the application of surgery
has markedly improved the prognosis for early-stage GC over the
past decade, most patients who are either diagnosed at an advanced
stage or who have relapsed after an apparently curative operation
still succumb to this cancer type, with an overall 5-year survival
rate of ~20% (2,3). Toxicity of combination chemotherapy
and drug resistance remain challenges in successfully treating GC
patients (4,5). In addition, drug resistance is
frequently observed in GC and other tumor types, but the details of
this resistance have remained to be fully elucidated (6,7).
In recent years, microRNAs (miRNAs/miRs), which are
evolutionarily conserved small non-coding RNAs, were identified to
have important roles in cell differentiation, apoptosis,
proliferation and metabolism by inhibiting translation of mRNA or
by promoting its degradation (8–12).
Mounting evidence suggests that miRNAs are involved in
tumorigenesis and progression, while they may have tumor suppressor
or oncogenic roles (13–15). Furthermore, the influence of
miRNAs on the tumor cell response to chemotherapeutic drugs has
been reported in various malignancies (16–18). However, the role of miRNAs in drug
resistance of GC has remained to be fully elucidated (19).
Recently, miR-21-5p has been of significant
interest, as studies have demonstrated that it is frequently
overex-pressed in a wide variety of cancer types, including breast
cancer (20), prostate cancer
(21), lung cancer (22,23), GC (24), pancreatic cancer (25), head and neck squamous cell
carcinoma (26) and leukemia
(27). In addition, it has been
reported that dysregulation of miR-21-5p may be a predictor of
tumor response to 5-fluorouracil, docetaxel and temozolomide
(28–30). Although doxorubicin (DOX) is an
unconventional chemotherapeutic drug for GC, it is effective in
certain cancer types, including leukemia, lymphoma and breast
cancer. However, the functions and mechanisms of miR-21-5p in
doxorubicin (DOX) drug resistance and relapse of GC have remained
elusive.
The present study reported that the upregulation of
miR-21-5p in clinical GC tissues is associated with an advanced
clinical stage and a poor prognosis. Furthermore, miR-21-5p was
identified to be upregulated in DOX-resistant GC cells. Inhibiting
the expression of miR-21-5p may effectively reverse DOX resistance.
In addition, the present study investigated the mechanisms of the
roles of miR-21-5p in GC. The results suggested that miR-21-5p has
an important role in DOX resistance of GC, with implications for
cancer therapy.
Patients and methods
Patients and clinical samples
Tissue samples from 46 patients with GC (tumor
tissues and matched adjacent normal tissues as a control) and their
clinical information were collected between January 2012 and
December 2014 at the First Affiliated Hospital of Nanchang
University (Nanchang, China). All patients received complete
resection (D2 type) and did not receive chemotherapy or
radiotherapy prior to surgery. In addition, certain patients
received corresponding chemotherapy after surgery. The
chemotherapeutic drugs included 5-fluorouracil, capecitabine and
cisplatin. Patient characteristics are displayed in Table I. The study protocol was approved
by the Ethics Committee of the First Affiliated Hospital of
Nanchang University (Nanchang, China). Written informed consent was
obtained from all participants.
 | Table ICharacteristics of gastric cancer
patients of the present study (n=46). |
Table I
Characteristics of gastric cancer
patients of the present study (n=46).
| Characteristic | Median (range) | n (%) |
|---|
| Age at diagnosis
(years) | 58 (46–71) | |
| <60 | | 20 (43.5) |
| ≥60 | | 26 (56.5) |
| Sex | | |
| Male | | 25 (54.3) |
| Female | | 21 (45.7) |
|
Differentiation | | |
| Well | | 16 (34.8) |
| Moderate | | 16 (34.8) |
| Poor | | 14 (30.4) |
| Metastasis | | |
| Yes | | 22 (47.8) |
| No | | 24 (52.2) |
|
Tumor-nodes-metastasis stage | | |
| I+II | | 30 (65.2) |
| III+IV | | 16 (34.8) |
| After therapy
(n=30)a | | |
| Complete
remission | | 17 (56.7) |
| Relapse or
death | | 13 (43.3) |
Cell culture and DOX-resistant cell
selection
The SGC7901 human gastric adenocarcinoma cell line
(cat. no. TCHU46) and the HEK-293T human embryonic kidney cell line
(cat. no. CRL-3216) were purchased from the Shanghai Institute of
Biomedical Sciences (Shanghai, China) and the American Type Culture
Collection (Manassas, VA, USA), respectively. Three DOX-resistant
SGC7901 cell lines (SGC7901/DOX) were generated from parental
SGC7901 cells by gradually treating them with 0.5, 1 and 2
μg/ml DOX for >1 month, as previously reported by Davies
et al (31). Hence, these
cells were named SGC7901/DOX-0.5 μg/ml, SGC7901/DOX-1
μg/ml and SGC7901/DOX-2 μg/ml. SGC7901 and the
selected SGC7901/DOX cell lines were cultured in Dulbecco's
modified Eagle's medium (Gibco; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) containing 10% fetal bovine serum (HyClone; GE
Healthcare, Little Chalfont, UK) at 37°C in a humidified atmosphere
containing 5% CO2 for further use.
RNA extraction and reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
analysis
Total RNA was extracted from fresh GC tissue using
RNAiso Plus (Takara Bio Inc., Otsu, Shiga Prefecture, Japan), and
the RNA concentration and quality were detected with a
NanoDrop-1000 spectrophotometer. RT-qPCR was then performed to
detect miR-21-5p and its target mRNA expression levels. For
RT-qPCR, 0.5 μg total RNA were used to generate
complementary DNA using the M-MLV Reverse Transcriptase kit (cat.
no. RR037A; Takara Bio Inc.). RT was for 30 min at 16°C, 45 min at
42°C and 5 min at 85°C. Then qPCR was performed to amplify the
targets using the LightCycler® 480 real-time PCR system
(Roche Diagnostics, Basel, Switzerland) with specific mRNA/miRNA
primers and SYBR® Premix Ex Taq™ II reagents (cat. no.
RR820A; Takara Bio Inc.) according to the manufacturer's protocol.
Thermocycling conditions were as follows: 95°C for 3 min followed
by 40 cycles of 95°C for 15 sec and 60°C for 30 sec. The relative
expression levels of mRNA and miRNA were determined using the
2−ΔΔCq method (32),
with GAPDH and U6 used as internal controls separately. All PCRs
were performed in triplicate. The sequences for all primers used in
PCR are listed in Table II.
| Table IIPrimers and other oligonucleotides
sequences. |
Table II
Primers and other oligonucleotides
sequences.
| Name | Sequence
(5′-3′) | Product length
(bp) |
|---|
| miR-21-5p-RT |
GTCGTATCCAGTGCAGGGTCCGAGGTATTCG
CACTGGATACGACTCAACA | |
| miR-21-5p-F |
GCACCTAGCTTATCAGACTGA | 22 |
| miR-21-5p-R |
GTGCAGGGTCCGAGGT | |
| U6-F C |
TCGCTTGGCAGCACATATACT | 90 |
| U6-R |
ACGCTTCACGAATTTGCGTGTC | |
| PTEN-3′UTR-F |
GCTTTGGTGGTAGTACTCTGGTTGTTAAGCT
AGGTATTTTGAGACTACTTCCCCATCAC | 59 |
| PTEN-3′UTR-R |
GTGATGGGGAAGTAGTCTCAAAATACCTAGC
TTAACAACCAGAGTACTACCACCAAAGC | |
| MUT-PTEN-F |
GCTTTGGTGGTAGTACTCTGGTTGTATTCGAT
GGTATTTTGAGACTACTTCCCCATCAC | 59 |
| MUT-PTEN- R |
GTGATGGGGAAGTAGTCTCAAAATACCATCG
AATACAACCAGAGTACTACCACCAAAGC | |
|
TIMP3-3′UTR(1)-F |
TTGCATATACCCACATGGGGACATAAGCTAA
TTTTTTTACAGGACACAGAATTCTGT | 59 |
|
TIMP3-3′UTR(1)-R |
ACAGAATTCTGTGTCCTGTAAAAAAATTAGC
TTATGTCCCCATGTGGGTATATGCAA | |
| MUT-TIMP3(1)-F |
TTGCATATACCCACATGGGGACAATTCGATAT
TTTTTTACAGGACACAGAATTCTGT | 59 |
| MUT-TIMP3(1)-R |
ACAGAATTCTGTGTCCTGTAAAAAAATATCG
AATTGTCCCCATGTGGGTATATGCAA | |
|
TIMP3-3′UTR(2)-F
C |
GATGTCAGAGGGCGGTTTTGAGCTTTCTATA
AGCTATAGCTTTGTTTATTTCACCCGT | 59 |
|
TIMP3-3′UTR(2)-R |
ACGGGTGAAATAAACAAAGCTATAGCTTATA
GAAAGCTCAAAACCGCCCTCTGACATCG | |
| MUT-TIMP3(2)-F C |
GATGTCAGAGGGCGGTTTTGAGCTTTCTAA
TTCGATTAGCTTTGTTTATTTCACCCGT | 59 |
| MUT-TIMP3(2)-R |
ACGGGTGAAATAAACAAAGCTAATCGAATTA
GAAAGCTCAAAACCGCCCTCTGACATCG | |
| PTEN-F |
GCCCAGACTGCATACGAT | 294 |
| PTEN-R CC |
TAATCTATTTGCCATCAA | |
| TIMP3-F |
TACCTGCCTTGCTTTGTG | 257 |
| TIMP3-R C |
ATCTGGGAAGAGTTAGTGTC | |
| Pre-miR-21-5p-F
C |
GCGGATCCGCATGGCAACACCAGTCGA | 42 |
| Pre-miR-21-5p-R
CC |
GCTCGAGTTGTCAGACAGCCCATCGA | |
| GAPDH-F |
GCTGAACGGGAAGCTCACTG | 169 |
| GAPDH-R |
GTGCTCAGTGTAGCCCAGGA | |
Cell transfection
All small molecular RNAs, including miR-21-5p mimics
and scrambled oligonucleotides miR-negative control (NC), miR-21-5p
antisense oligonucleotide (anti-miR-21-5p) and its counterpart
control (anti-miR-NC), and small interfering (si)RNA for the target
mRNA were purchased from GenePharma Biotech (Shanghai, China).
These small RNAs were delivered into cultured cells
using Lipofectamine® 3000 (Invitrogen; Thermo Fisher
Scientific, Inc.). In brief, 3×105 cells were seeded in
6-well plates and grown to 75% confluence. All RNA transfections
were performed at a final concentration of 50 nM unless otherwise
indicated according to the manufacturer's protocol. The
transfection efficiency, which was assessed using a fluorescein
amidite-conjugated siRNA (non-homologous to any human genome
sequences; cat. no. A07001; GenePharma Biotech, Shanghai, China)
and fluorescence-activated cell sorting (FACS) analysis, was
determined to be ~80% for GC cells (data not shown).
Measurement of drug sensitivity using the
cell counting kit-8 (CCK-8) assay
The transfected cells were cultured in 96-well
plates at 5×103 cells per well in 100 μl medium
and were maintained overnight, followed by exposure to various
concentrations of DOX for 48 h. Cell viability was monitored using
a CCK-8 (Dojindo, Kumamoto, Japan). In brief, 10 μl CCK-8
stain was added to each well of the plate, followed by incubation
for 2 h and measurement of the absorbance at 450 and 630 nm
(reference wavelength) using a microplate reader. The concentration
causing 50% inhibition of growth (IC50) of the drug was
estimated from a relative survival curve. All experiments were
performed three times.
Apoptosis assay
Apoptosis was assessed using an Annexin
V-fluorescein isothiocyanate (FITC) apoptosis kit (cat. no.
K101-100; Biovision, Milpitas, CA, USA). Cells were washed with
ice-cold PBS and re-suspended in 1X Annexin V binding solution at a
concentration of 1×106 cells/ml. The cells were then
collected and stained with 5 μl Annexin V-FITC and 5
μl propidium iodide for 15 min at room temperature in the
dark prior to analysis with a FACSCalibur instrument (BD
Biosciences, Franklin Lakes, NJ, USA). The collected data were
analyzed using FlowJo V10.1 software (BD Biosciences).
Target gene prediction
Four computer programs/algorithms, namely Targetscan
(http://www.targetscan.org), starBase
v2.0 (http://starbase.sysu.edu.cn/),
miRBase (http://www.mirbase.org/index.shtml) and PICTAR
(http://pictar.mdc-berlin.de), were used
to predict the targets of miR-21-5p. To overcome the limitations of
each program, the putative target genes predicted by at least three
of the programs were selected for further experimental
analyses.
Construction of the vector and luciferase
reporter assay
The expression vector for miR-21-5p
(pcDNA-miR-21-5p) was generated by cloning the genomic fragments
encompassing the corresponding miR-21-5p precursor and its 5′- and
3′-flanking sequences into the pcDNA6.2 plasmid (cat. no. K246020;
Invitrogen; Thermo Fisher Scientific, Inc., Waltham, MA, USA).
Fragments of the 3′-untranslated region (UTR) of potential target
mRNAs containing putative binding sites for miR-21-5p were inserted
downstream of the stop codon of Firefly luciferase in the
psi-Check2 vector (cat. no. C8021; Promega Corp., Madison, WI, USA)
as previously described (33). To
yield mutant constructs, mutations were introduced in the potential
miR-21-5p binding sites using the QuikChange Site-directed
Mutagenesis kit (cat. no. 210515; Stratagene, San Diego, CA, USA).
The luciferase reporter constructs and pcDNA-miR-21-5p (or empty
plasmid) were co-transfected into HEK-293T cells in a 48-well plate
for reporter assays using Lipofectamine™ 3000 (Invitrogen; Thermo
Fisher Scientific, Inc.), respectively. Renilla and Firefly
luciferase activities were assayed using the
Dual-Luciferase® Reporter 1000 assay system (cat. no.
E1960; Promega Corp.). The effect of miR-21-5p on target gene
expression was detected by calculating the relative value of
Firefly luciferase activity normalized to Renilla luciferase
activity.
Western blot analysis
Cells were harvested at 48 h after transfection,
washed with PBS and lysed in radioimmuno-precipitation assay buffer
(Thermo Fisher Scientific, Inc.) supplemented with protease
inhibitors (cat. no. 04693132001; Roche Diagnostics) for 30 min at
4°C prior to centrifugation. Subsequently, the concentration was
measured using the BCA Protein assay kit (cat. no. 23225; Pierce;
Thermo Fisher Scientific, Inc.). Equal amounts of protein (20
μg) were separated by 10% SDS-PAGE and then transferred onto
polyvinylidene difluoride membranes (EMD Millipore, Billerica, MA,
USA). Membranes were blocked for 2 h at room temperature by using
bovine serum albumin (cat. no. SW3015-500 ml; Beijing Solarbio
Science and Technology Co., Ltd., Beijing, China), the membranes
were incubated overnight at 4°C with anti-phosphatase and tensin
homologue (PTEN) antibody (cat. no. 14642; 1:1,000), anti-tissue
inhibitor of matrix metalloproteinases 3 (TIMP3) antibody (cat. no.
5673; 1:1,000), anti-tubulin antibody (cat. no. 2146; 1:2,000 all
from Cell Signaling Technology, Inc., Danvers, MA, USA), or
anti-GAPDH antibody (cat. no. 60004-1-l g; 1:5,000) followed by
horseradish peroxidase-linked secondary antibodies (cat nos.
SA00006-2 and SA00001-1; 1:5,000; all from Proteintech, Wuhan,
China) for 1 h at room temperature. Proteins were then visualized
with a chemiluminescence detection system (cat. no. WBKLSO100; EMD
Millipore). The experiments were performed at least three
times.
Statistical analysis
Statistical analyses were performed with GraphPad
Prism 6.0 software (GraphPad Inc., La Jolla, CA, USA). The
cytological experiments were performed at least in triplicate and
all values are expressed as the mean ± standard deviation.
Differences between groups were analyzed using Student's t-test
when only two groups were present, and in the case of more than two
groups, one-way analysis of variance followed by a post-hoc
multiple comparisons test (least-significant differences test) was
applied. Unless otherwise noted, differences in mRNA/miRNA
expression in clinical samples between two groups were analyzed
using the Mann-Whitney U test for independent unpaired samples, and
paired samples were analyzed using the Wilcoxon test. The
χ2 or Fisher's exact test were used to analyze
categorical variables. Correlations were determined using Spearman
correlation analysis. P<0.05 was considered to indicate a
statistically significant difference.
Results
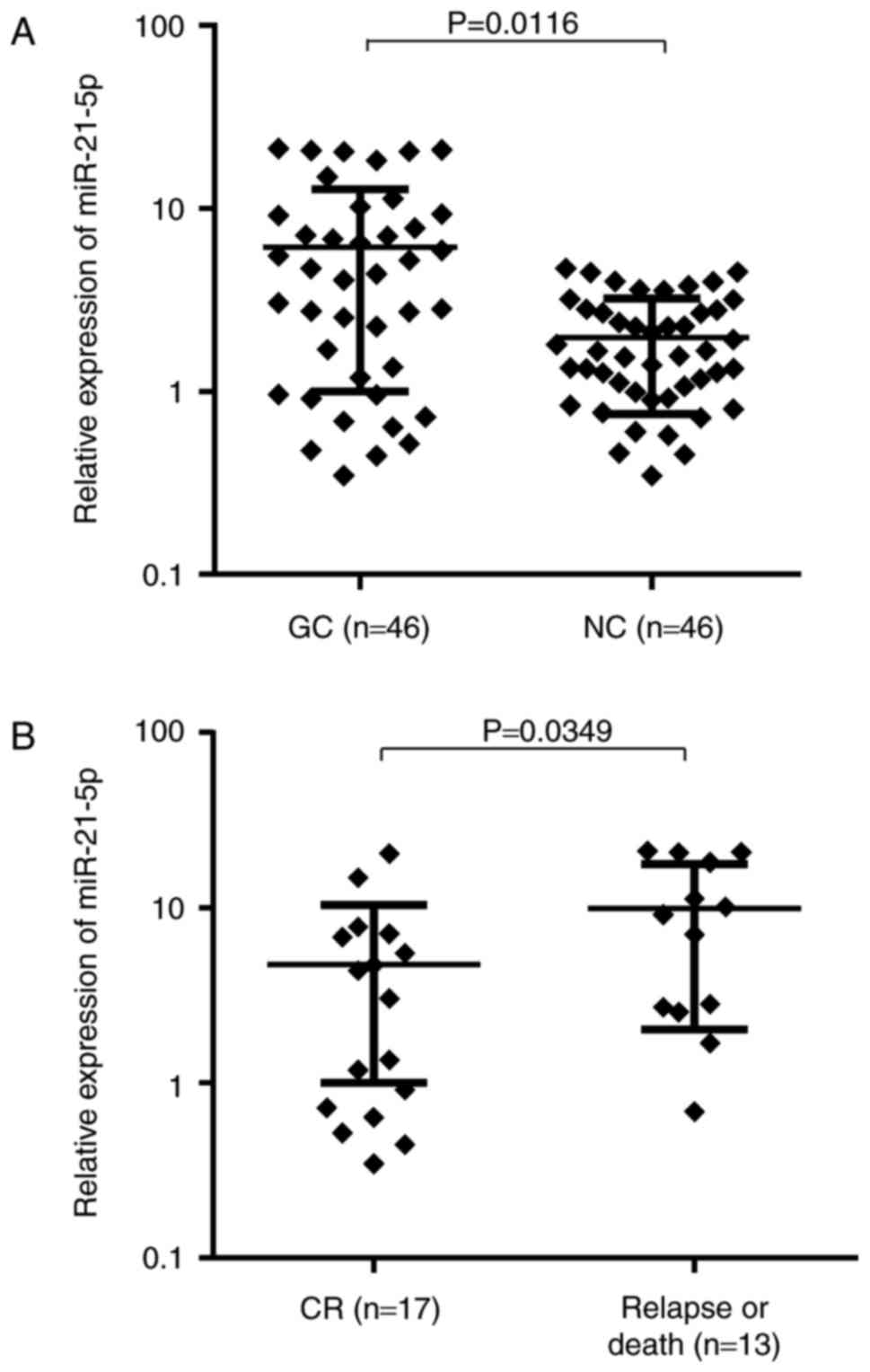
Upregulation of miR-21-5p in clinical GC
tissue is correlated with an advanced clinical stage and poor
prognosis
In a previous study (34), differential miRNA expression
profiles revealed dysregulation of miR-21-5p in GC tissues. To
further confirm this result, the present study examined miR-21-5p
expression levels in primary tumor samples from a GC patient cohort
(n=46). miR-21-5p expression in GC tissue was 3.91-fold (range,
0.031–72.429) higher than that in matched adjacent NC tissues
(P=0.0116; Fig. 1A). The median
values of relative miR-21-5p expression in the GC and NC tissues
were 4.03 and 1.27, respectively. Of these 46 GC patients, 24
(52.2%) exhibited a tumor tissue-specific upregulation of miR-21-5p
expression levels by >2-fold. To determine whether miR-21-5p
expression correlates with clinical characteristics of GC patients,
the patients were stratified into high and low expression groups
according to the median value of miR-21-5p expression. As indicated
in Table III, a statistically
significant correlation between miR-21-5p expression and available
pathological parameters was identified, including differentiation
status, lymph node metastasis and tumor-nodes-metastasis (TNM)
stage of GC.
| Table IIICharacteristics of gastric cancer
patients in association with miR-21-5p expression levels. |
Table III
Characteristics of gastric cancer
patients in association with miR-21-5p expression levels.
| Parameter | Total (n) | miR-21-5p
| P-value |
|---|
| Low expression
(n) | High expression
(n) |
|---|
| Age at diagnosis
(years) | | | | 0.234 |
| <60 | 20 | 12 | 8 | |
| ≥60 | 26 | 11 | 15 | |
| Sex | | | | 0.375 |
| Male | 25 | 14 | 11 | |
| Female | 21 | 9 | 12 | |
|
Differentiation | | | | 0.017a |
| Well | 16 | 9 | 7 | |
| Moderate | 16 | 10 | 6 | |
| Poor | 14 | 2 | 12 | |
| Metastasis | | | | 0.013a |
| Yes | 22 | 4 | 18 | |
| No | 24 | 14 | 10 | |
|
Tumor-nodes-metastasis stage | | | | 0.047a |
| I+II | 30 | 20 | 10 | |
| III+IV | 16 | 5 | 11 | |
Of the 46 primary patients, 16 (34.8%) received
treatment and were then lost to follow-up. The remaining 30 GC
patients (65.2%) with complete follow-up data (mean follow-up time,
39 months) were divided into two groups based on prognosis. The
miR-21-5p expression levels at diagnosis in the patients who
suffered from relapse or death (n=13) was significantly higher than
that of the remaining 17 patients in continuous complete remission
(CR; P=0.0349; Fig. 1B). These
results collectively indicate that higher miR-21-5p expression in
GC patients is correlated with an advanced clinical stage and a
poor prognosis.
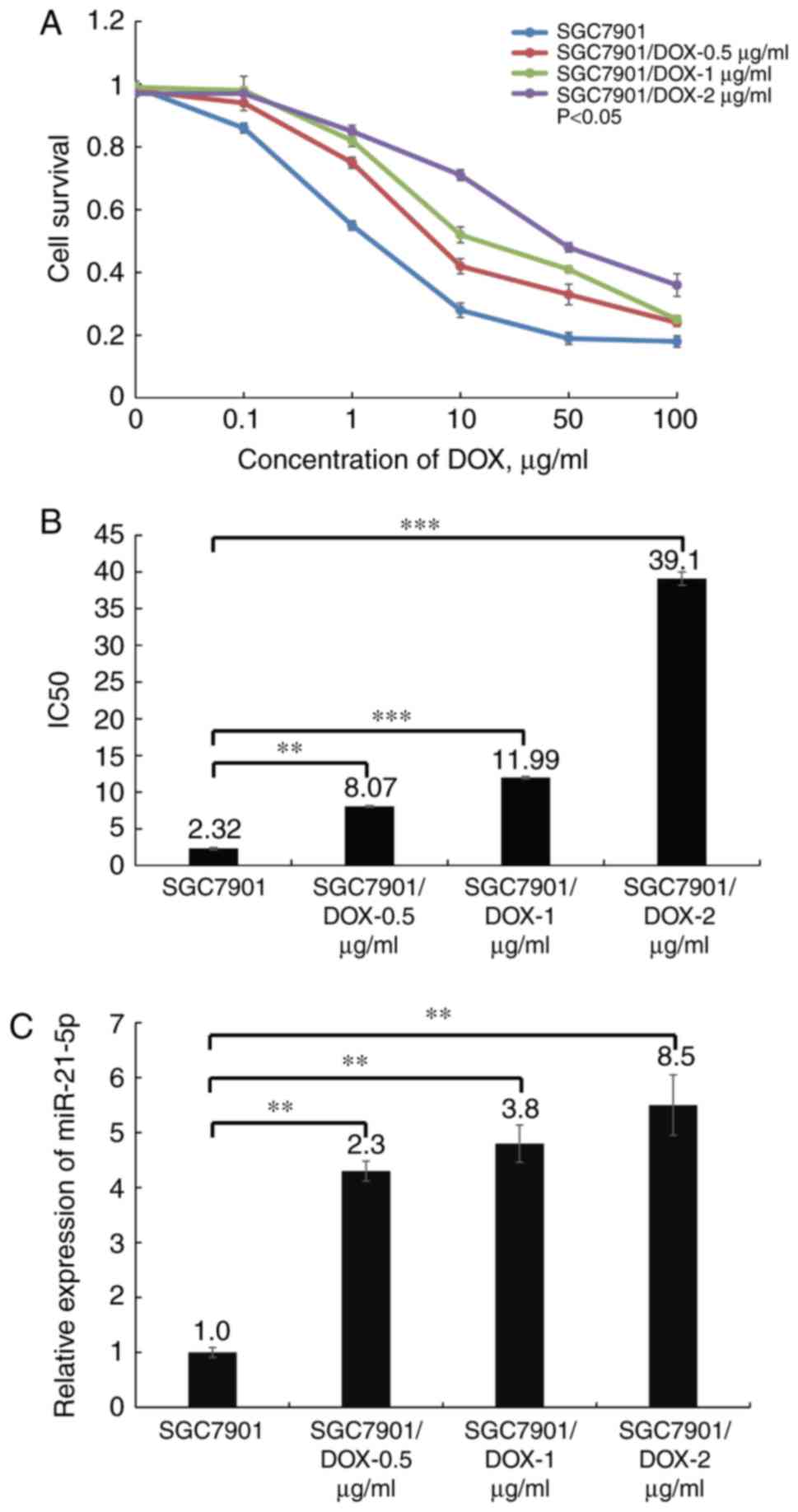
miR-21-5p expression is dysregulated in
DOX-resistant SGC7901/DOX cells
The survival curves of the three SGC7901/DOX cell
lines and the parental cell line SGC7901 are illustrated in
Fig. 2A. The results indicated
that the three SGC7901/DOX cell lines, SGC7901/DOX-0.5
μg/ml, SGC7901/DOX-1 μg/ml and SGC7901/DOX-2
μg/ml had a 1.4-, 2.1- and 3.3-fold acquired resistance to
DOX, respectively, based on their IC50 values
(P<0.05; Fig. 2B). Next, the
miR-21-5p expression levels were determined in the three
DOX-resistant cell lines to further elucidate whether miR-21-5p
expression affects DOX resistance. The results indicated that
miR-21-5p was least expressed in the SGC7901 cells, while the
expression levels gradually increased in the three DOX-resistant
cell lines with increasing DOX tolerance levels (P<0.05;
Fig. 2C), suggesting that
miR-21-5p may be associated with DOX resistance of GC cells. As
SGC7901/DOX-2 μg/ml exhibited the greatest resistance to
DOX, this cell line was used for the subsequent experiments.
Suppression of miR-21-5p expression
sensitizes SGC7901/DOX cells to DOX
To elucidate the association between miR-21-5p and
DOX resistance in GC cells, miR-21-5p antagomir (anti-miR-21) was
transfected into SGC7901/DOX-2 μg/ml to specifically inhibit
the expression of miR-21-5p. The miR-21-5p expression in the miR-21
inhibitor group was reduced to 44.8% of that in the corresponding
control group (P<0.05; Fig.
3A), and the anti-miR-21-transfected group of SGC7901/DOX-2
μg/ml cells exhibited a significantly lower survival rate
upon exposure to DOX compared with that of the control-transfected
group. Furthermore, the IC50 value was determined to be
1.443±0.017 μg/ml for the anti-miR-21 group and 3.985±0.064
μg/ml for the anti-miR-NC group (P<0.05; Fig. 3B). In addition, it was
demonstrated that in SGC7901/DOX-2 μg/ml cells, transfection
with anti-miR-21 enhanced DOX-induced apoptosis following DOX
treatment at 1 μg/ml for 48 h (P<0.05; Fig. 3C). These results indicate that
knockdown of miR-21-5p increased DOX sensitivity in SGC7901/DOX-2
μg/ml cells; thus, it was hypothesized that antagonism of
miR-21-5p partially reversed DOX resistance in these cells.
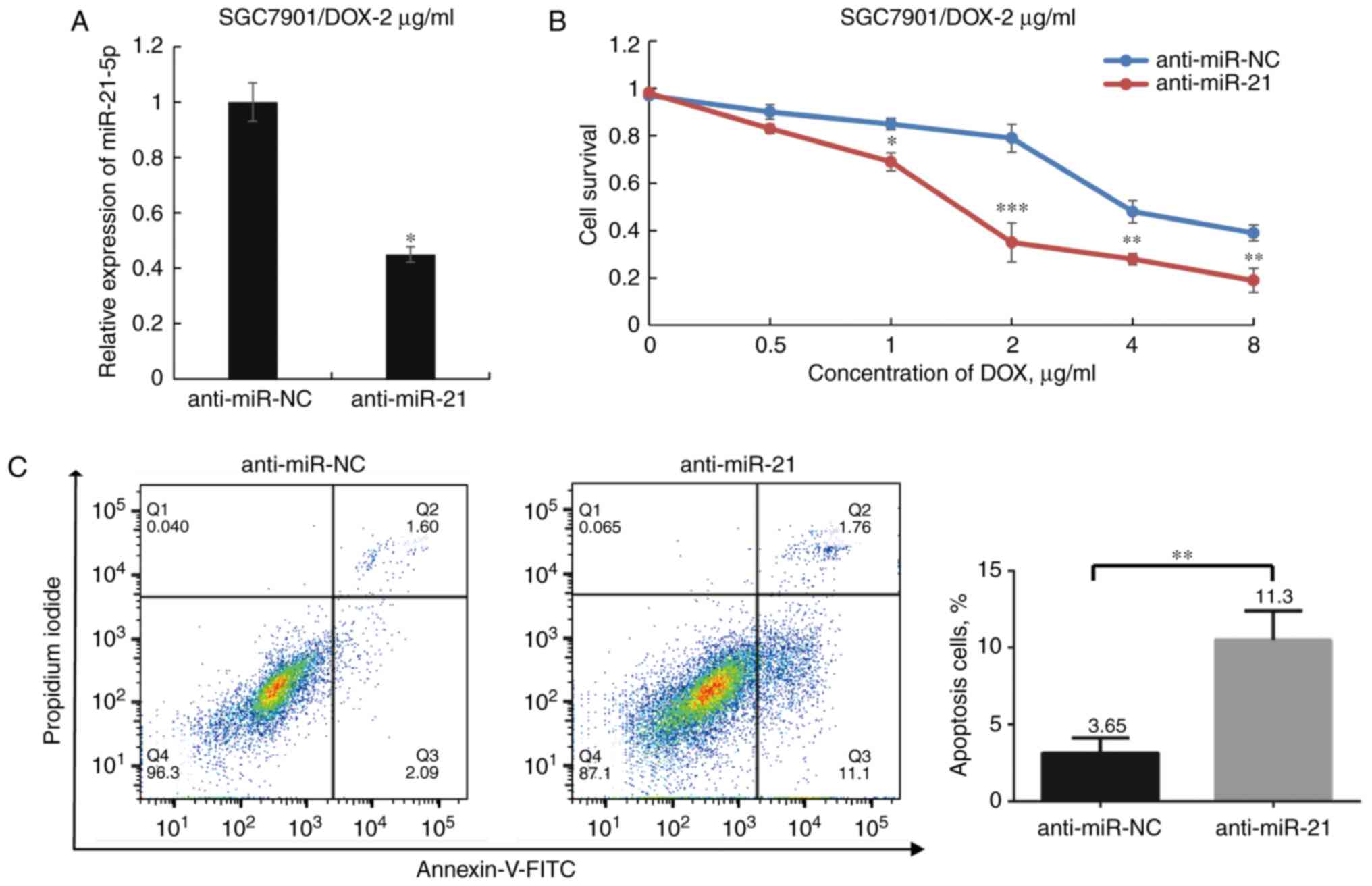 | Figure 3Effect of miR-21-5p antagomir on DOX
sensitivity of SGC7901/DOX-2 μg/ml cells. (A) Transfection
of anti-miR-21 decreased miR-21-5p expression levels in
SGC7901/DOX-2 μg/ml cells to 44.8% of those in the
anti-miR-NC group. (B) After transfection of SGC7901/DOX cells with
anti-miR-21 or anti-miR-NC for 24 h and treatment with various
doses of DOX for 48 h, the cell viability was assessed. (C) Flow
cytometric analysis determined the influence of miR-21-5p on the
apoptosis of SGC7901/DOX-2 μg/ml cells following treatment
with DOX (1 μg/ml) for 48 h. The proportions of dead cells
(Q1: Annexin V-FITC−/PI+), late apoptotic or
necrotic cells (Q2: Annexin V-FITC+/PI+),
early apoptotic cells (Q3: Annexin
V-FITC+/PI−) and live cells (Q4: Annexin
V-FITC−/PI−) are displayed.
*P<0.05, **P<0.01 and
***P<0.001 vs. anti-miR-NC. miR, microRNA; FITC,
fluorescein isothiocyanate; PI, propidium iodide; DOX, doxorubicin;
SGC7901/DOX, DOX-resistant SGC7901 cell line; NC, negative control;
Q, quadrant. |
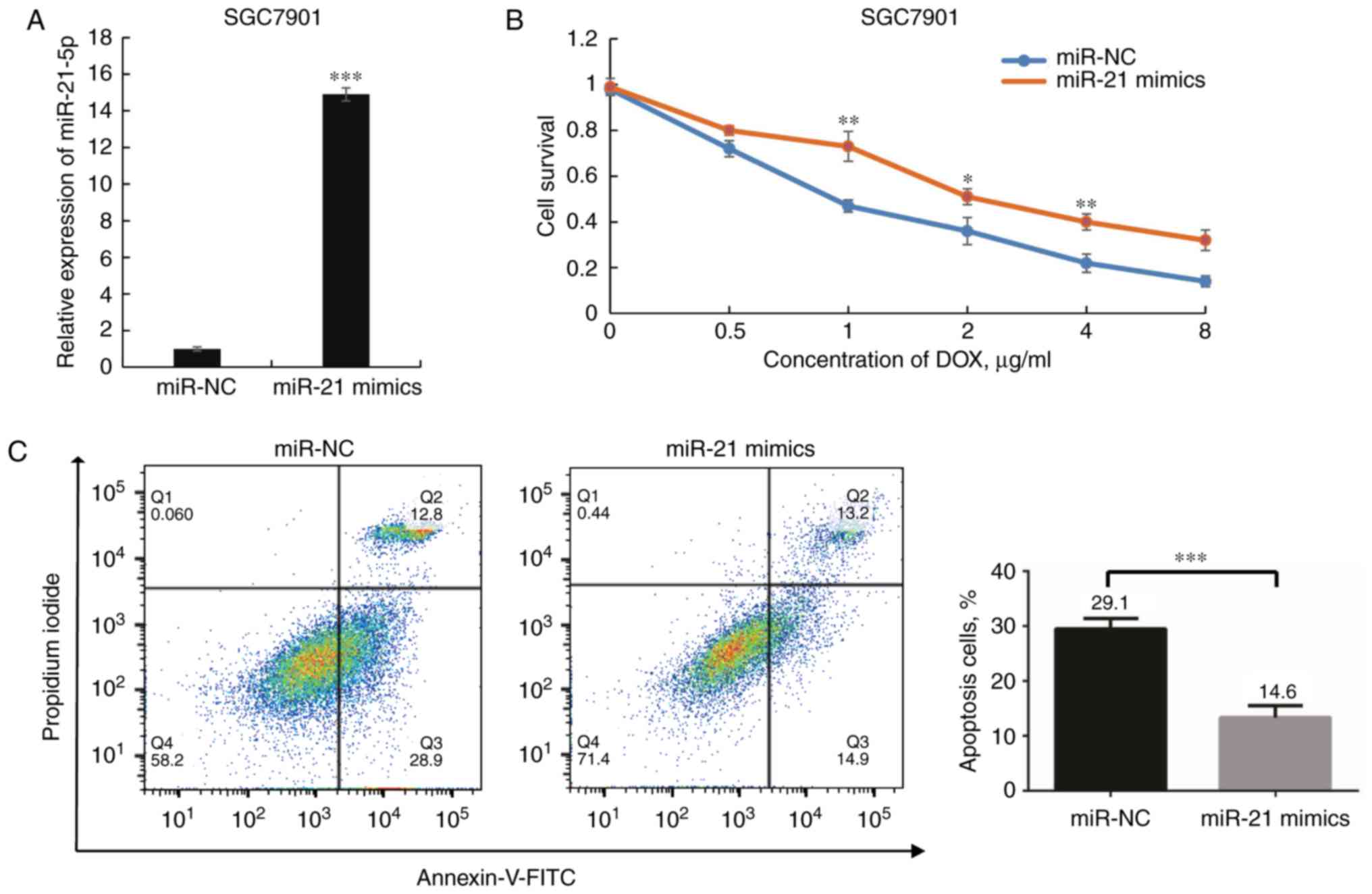
Upregulation of miR-21-5p renders SGC7901
cells resistant to DOX
Based on the above evidence, the present study
further investigated the effects of miR-21 in SGC7901 cells. The
miR-21-5p expression levels were significantly elevated following
transfection with miR-21-5p mimics (P<0.001; Fig. 4A). Next, the cell viability was
examined with the goal of determining whether overexpression of
miR-21-5p affects the sensitivity of SGC7901 cells to DOX. A
dose-dependent growth inhibition of SGC7901 cells in response to
DOX was observed, and cells transfected with miR-21 mimics
exhibited significantly lower DOX sensitivity than those
transfected with miR-NC (P<0.05; Fig. 4B). The IC50 of DOX in
the former group was higher than that in the latter group. In
addition, overexpression of miR-21-5p rendered SGC7901 cells more
resistant to DOX-induced apoptosis upon DOX treatment (P<0.05;
Fig. 4C). Taken together, these
results suggest that miR-21-5p has an important role in DOX
resistance of GC cells.
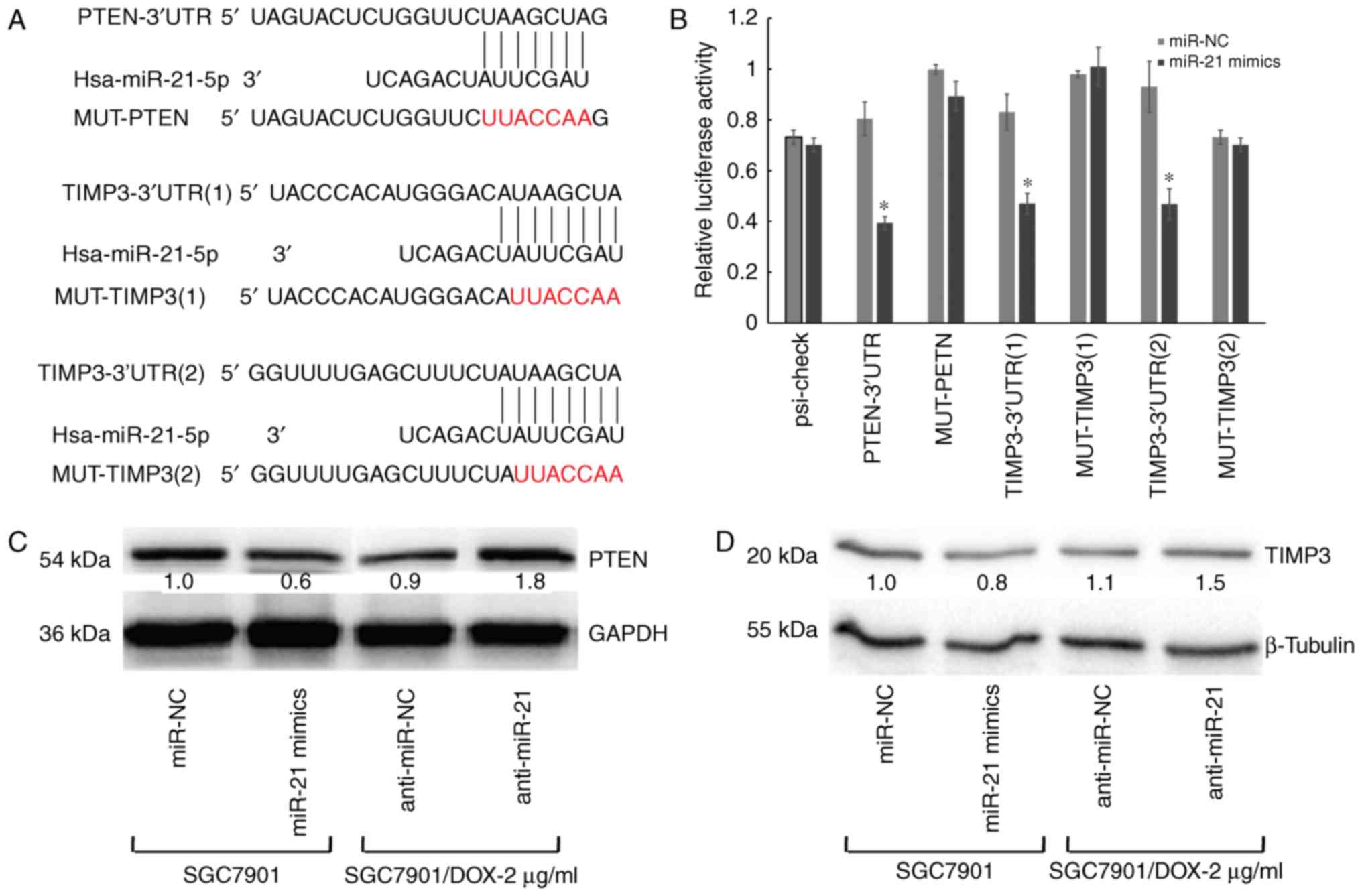
miR-21-5p mediates apoptosis by targeting
PTEN and TIMP3
The predicted targets of miR-21-5p are PTEN and
TIMP3, which were identified by using several miRNA target gene
prediction websites. The 3′UTR of PTEN and TIMP3 contains one and
two putative miR-21-5p-binding sites, respectively (Fig. 5A). To verify this, three plasmids
were generated by cloning the respective fragments from the 3′-UTR
of PTEN or TIMP3 into the downstream region of the Renilla
luciferase reporter gene. HEK-293T cells were then co-transfected
with one of the wild type or mutant reporter vectors and precursor
miR-21-5p or miR-NC. A significant reduction of luciferase activity
of the wild-type reporter vector was observed in the presence of
miR-21-5p. Conversely, partial mutation of the respective seed
regions blocked the ability of miR-21-5p to inhibit luciferase
activity (P<0.05; Fig. 5B). In
addition, PTEN protein was significantly downregulated in miR-21-5p
mimics-transfected cells, but was significantly increased in
inhibitor-transfected cells (Fig.
5C). Similar results were also observed for TIMP3 (Fig. 5D). Taken together, these results
indicated that miR-21-5p directly regulates PTEN and TIMP3 as
target mRNAs in GC cells.
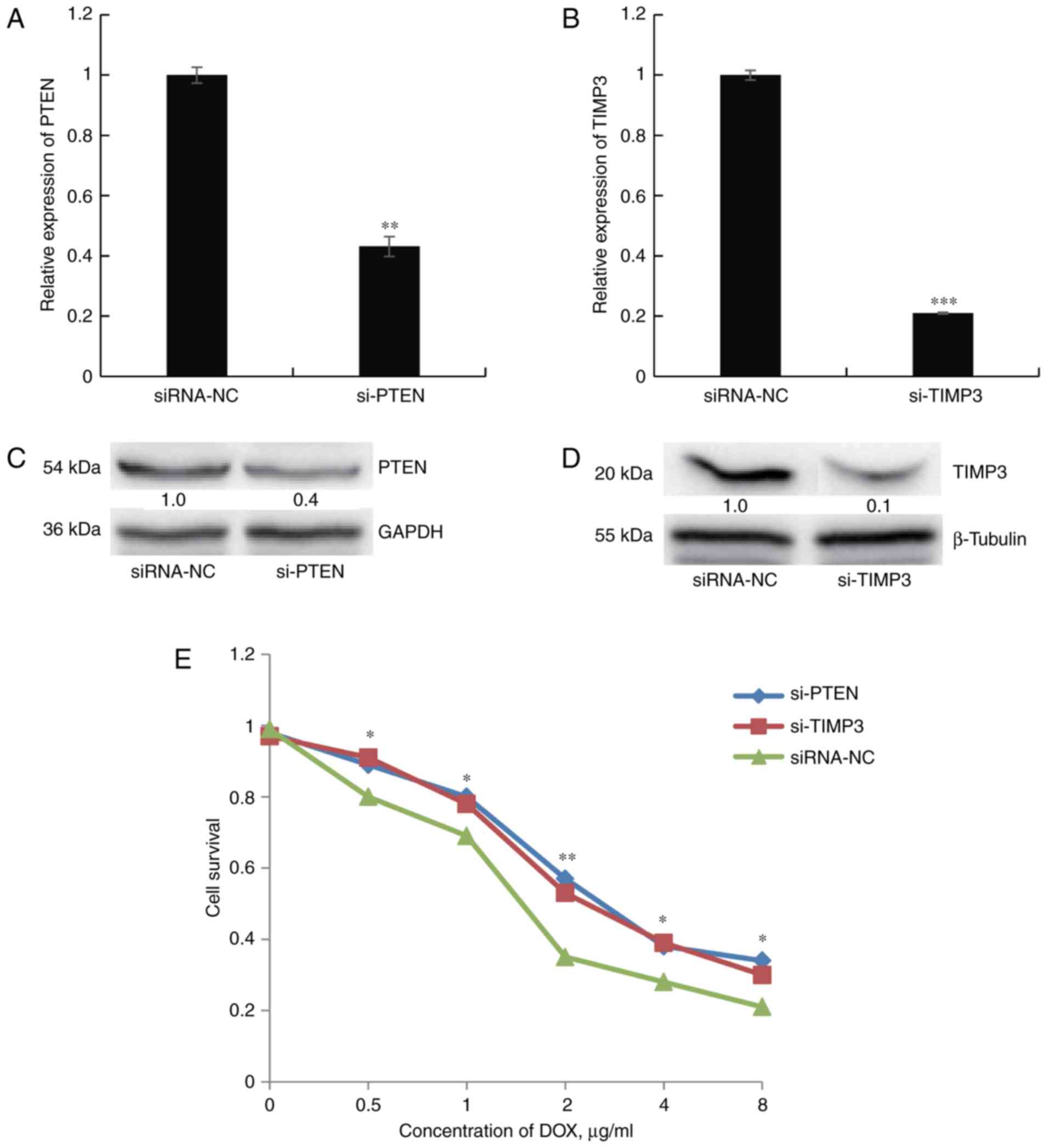
Knockdown of PTEN and TIMP3 enhances DOX
resistance of SGC7901 cells
Next, the present study sought to determine whether
direct knockdown of PTEN or TIMP3 confers cellular DOX resistance,
similar to the effect of miR-21-5p mimics. First, SGC7901 cells
were depleted of endogenous PTEN or TIMP3 by transfection of
specific siRNA. The transfection efficiency was confirmed through
RT-qPCR and western blot analysis (Fig. 6A–D). Silencing of PTEN or TIMP3
mRNA caused significant inhibition of the corresponding mRNA and
protein expression in SGC7901 cells. The cell survival assay
revealed that DOX resistance was significantly increased in
siRNA-PTEN- or siRNA-TIMP3-transfected cells compared with that of
cells transfected with control siRNA (Fig. 6E). All of these results mirrored
those obtained by transfection of miR-21-5p and further suggested
that the effects of miR-21-5p on DOX resistance in GC cells may be
mediated by targeting PTEN and TIMP3.
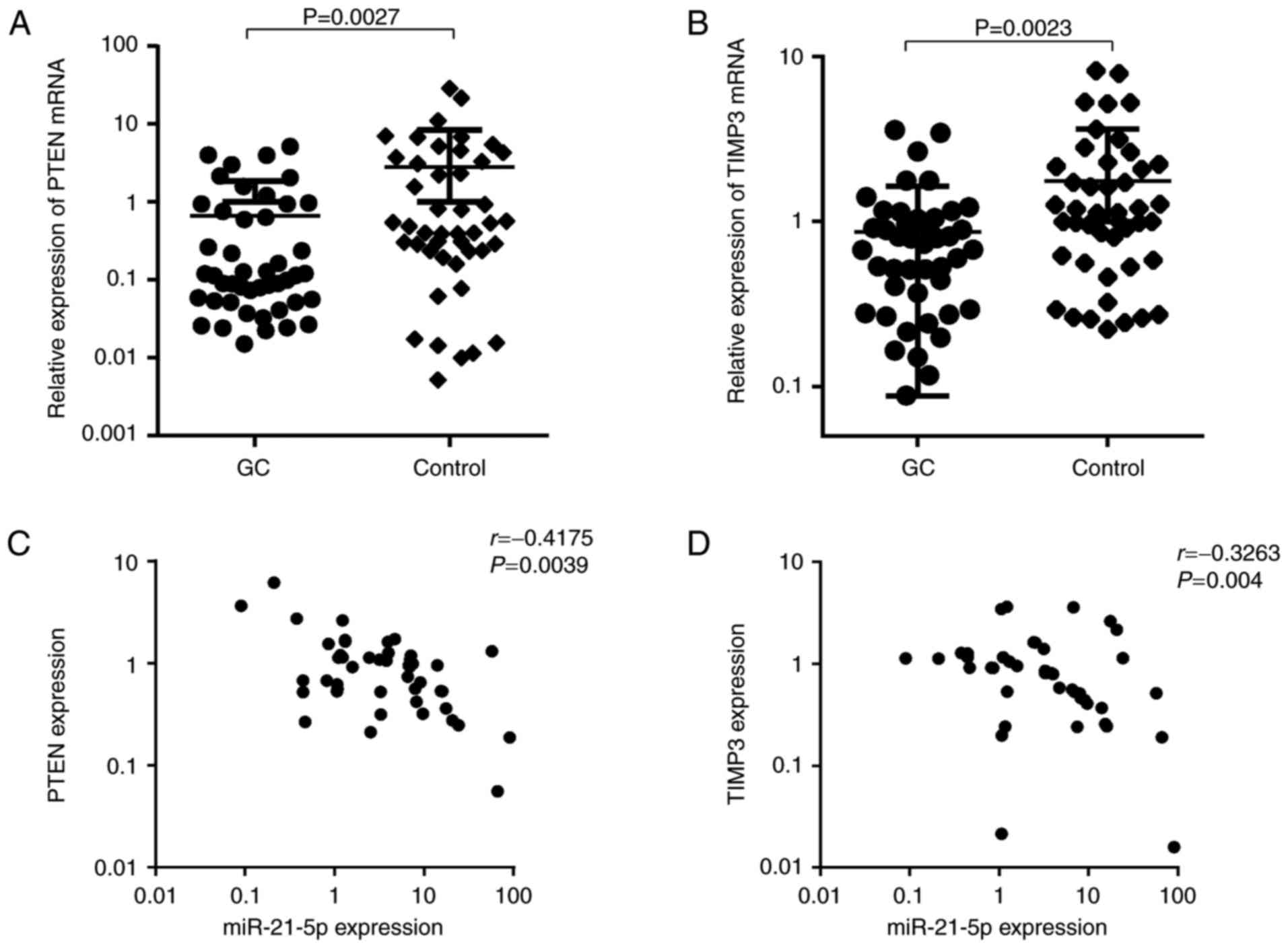
Clinical relevance of miR-21-5p and PTEN
or TIMP3 in GC patients
To assess the clinical relevance of miR-21-5p and
PTEN or TIMP3 in GC patients, PTEN and TIMP3 mRNA levels were first
detected in newly diagnosed GC patients, and the possible
correlation between miR-21-5p and the target genes was then
evaluated. As presented in Fig. 7A
and B, PTEN and TIMP3 were significantly downregulated in GC
patients (P<0.01). Furthermore, a significantly inverse
correlation between miR-21-5p expression and PTEN levels was
observed in this cohort (r=−0.4175, P=0.0039). In addition, a
statistically significant correlation between miR-21-5p levels and
TIMP3 expression was identified in the GC samples examined
(r=−0.3263, P=0.004). This result suggests that PTEN and TIMP3 are
also regulated by miR-21-5p in clinical samples, indicating that
these genes are bona fide targets of miR-21-5p in GC
cells.
Discussion
At present, the reversal of drug resistance largely
determines the efficacy of successful cancer treatment in the
clinic (35,36). One of the mechanisms accounting
for acquired drug resistance is failure to undergo apoptosis in
response to the drugs (37–39). The present study demonstrated that
miR-21-5p is upregulated in DOX-resistant GC cell lines and GC
patients, particularly in recurrent cases, which implies that
miR-21-5p is involved in DOX resistance. Knockdown of miR-21-5p
enhanced sensitivity and promoted DOX-induced apoptosis in
SGC7901/DOX cells. An in-depth examination of the inverse
correlation between PTEN or TIMP3 expression and miR-21-5p in
SGC7901 cells and in the sufficiently large patient cohort
indicated that these targets are involved in DOX resistance. The
results provide novel evidence regarding the implication of
miR-21-5p in DOX resistance of GC and suggest a significant
association of miR-21-5p with GC patient prognosis.
Previous studies have reported that miR-21-5p
participates in the tumorigenesis of several cancer types. Yan
et al (40) and Wu et
al (41) reported that the
dysregulated expression of miR-21 is correlated with lymph node
metastasis and poor prognosis in breast cancer and colorectal
cancer, respectively. It has been reported that overexpression of
miR-21 predicts poor survival of non-small-cell lung cancer
patients and inhibits apoptosis through the regulation of the
Ras/mitogen-associated protein kinase kinase/extracellular
signal-regulated protein kinase pathway (42). Furthermore, it has been reported
that miR-21-5p is upregulated in patients aged >40 years who
underwent gastrectomy for potentially curable GC (stage III) and
experienced a recurrence (43).
In addition, a meta-analysis study indicated that elevated miR-21
expression levels are correlated with tumor differentiation, lymph
node metastasis and TNM stage in digestive tract cancer (44), which is consistent with the
results of the present study.
The importance of miR-21-5p in the progression of
drug resistance is well documented. Gong et al (45) reported that ectopic expression of
miR-21 in HER2+ breast cancer cells conferred resistance
to trastuzumab and that miR-21-specific antisense oligonucleotides
reversed this resistance by inducing growth arrest through blocking
the G1/S cell cycle checkpoint. Tomimaru et al (46) reported that anti-miR-21 rendered
hepatocellular carcinoma cells sensitive to 5-fluorouracil by
targeting PTEN and programmed cell death protein (PDCD)4.
Consistent with these results, Yang et al (47) demonstrated that compared with the
parental SGC7901 cell line, miR-21 was upregulated in the cisplatin
(DPP)-resistant SGC7901/DDP cell line, and knockdown of miR-21
markedly increased the anti-proliferative effects and induced
apoptosis by DPP through targeting PTEN. A previous study reported
that miR-21 was upregulated in the paclitaxel-resistant cell line
SGC7901/paclitaxel and may modulate sensitivity to
paclitaxel-mediated P-glycoprotein expression (48). In the present study, SGC7901/DOX
cells did not display marked DOX resistance when miR-21-5p
expression was increased. This may be due to following aspects: The
elevated expression levels of miR-21-5p may not directly translate
to elevated DOX resistance, and miR-21-5p may influence the
expression of certain other genes and may be associated with the
resistance to other drugs. For instance, various other studies have
reported that miR-21-5p was associated with DPP resistance of
gastric cancer cells (47,49).
Furthermore, the increased resistance to DOX was not entirely due
to the elevation of miR-21-5p, and is probably regulated by
additional signaling pathways, including the inhibition of
P-glycoprotein expression and activity, or via AKT-mediated
upregulation of multidrug resistance-associated protein 1 (50,51). All of the above suggests that the
mechanism of drug resistance is highly complex, while it is
indicated that miR-21-5p may have an important role in the
multidrug resistance of cancer cells.
It is broadly accepted that certain miRNAs regulate
a large number of protein-coding genes during cell apoptosis. For
instance, PDCD, apoptotic protease-activating factor 1, Fas ligand
and B-cell lymphoma-2 (Bcl-2) have all been proven to be targets of
miR-21-5p that are associated with apoptotic signaling (46,52–54). Consistent with these results, the
present study demonstrated that miR-21-5p reduced apoptosis and
enhanced survival of GC cells in the presence of DOX through
inhibition of the expression of PTEN, as well as TIMP3, which is
known to induce apoptosis and act as a tumor suppressor gene.
PTEN is a tumor suppressor gene that suppresses the
proliferation, invasion and metastasis of tumor cells and induces
tumor cell apoptosis (55).
PTEN-mediated apoptosis in tumors is regulated by numerous
canonical signaling pathways, among which the phosphoinositide-3
kinase/AKT/mammalian target of rapamycin pathway is of high
interest. Activation of this pathway protects various cell types
from apoptosis that is induced by withdrawal of survival factors
(56,57), induces the expression of
anti-apoptotic Bcl-2, and phosphorylates and inactivates the
pro-apoptotic Bcl-2 family member Bcl-2-associated death promoter,
which induces the expression of genes that are critical for cell
death. The imbalance between Bcl-2 and Bcl-2-associated X protein
may trigger apoptosis (58).
TIMP3, a natural metalloproteinase inhibitor, is always silenced in
human cancers. Pertinent studies have identified TIMP3 to be a
critical regulator of cell death and survival. Two major pathways,
p38 and Notch, were activated in the livers of TIMP3−/−
mice (59). p38 negatively
regulates cell cycle progression, facilitating DNA repair by
inducing p21 or directly activating p53 (60,61), while Notch signaling, via p53 and
p21 activation, couples enhanced cell cycle arrest and senescence
with tumor cell differentiation to inhibit tumorigenesis (62). The present results indicated that
miR-21-5p regulates drug resistance and cell apoptosis in GC by
targeting the tumor suppressors PTEN and TIMP3, and then affect the
downstream signaling pathways. In future studies, specific
knockdown or inhibition of individual pathways may provide a deeper
understanding of these interactions.
The present study is limited by its lack of animal
experiments, which would have provided more powerful evidence for
implementation of the results in drug therapy (63,64). However, the present study employed
various experimental methods and combined clinical data for
analysis in the present study. Similar studies have been performed
previously, which used identical methods to verify the effect of a
gene or protein on tumor cell resistance to certain drugs (65,66). A future study by our group will
assess the function of miR-21-5p in a mouse model.
The present study underscored the value of miR-21-5p
in DOX resistance of GC cells. It is worth noting that the
increased resistance of GC cells was not consistent with changes in
miR-21-5p, which implies that a complex regulation network exists
in DOX-resistant GC cells.
In conclusion, miR-21-5p expression was upregulated
in DOX-resistant GC cells and patient tissue samples. Reduction of
miR-21-5p levels improved the sensitivity of GC cells to DOX-based
chemotherapy through directly upregulating PTEN and TIMP3,
suggesting that an appropriate combination of DOX with miR-21-5p
antagonism may represent a potential strategy for GC treatment.
Acknowledgments
This work was supported by the Projects of
International Cooperation and Achievements Management Application
of Nanchang City (grant no. KJHZ-YY-002).
Notes
[1] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Karimi P, Islami F, Anandasabapathy S,
Freedman ND and Kamangar F: Gastric cancer: Descriptive
epidemiology, risk factors, screening, and prevention. Cancer
Epidemiol Biomarkers Prev. 23:700–713. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Ali Z, Deng Y and Ma C: Progress of
research in gastric cancer. J Nanosci Nanotechnol. 12:8241–8248.
2012. View Article : Google Scholar
|
|
3
|
Kikuchi S, Kaibe N, Morimoto K, Fukui H,
Niwa H, Maeyama Y, Takemura M, Matsumoto M, Nakamori S, Miwa H, et
al: Overexpression of Ephrin A2 receptors in cancer stromal cells
is a prognostic factor for the relapse of gastric cancer. Gastric
Cancer. 18:485–494. 2015. View Article : Google Scholar
|
|
4
|
Garrido M, Fonseca PJ, Vieitez JM, Frunza
M and Lacave AJ: Challenges in first line chemotherapy and targeted
therapy in advanced gastric cancer. Expert Rev Anticancer Ther.
14:887–900. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Di Lauro L, Vici P, Belli F, Tomao S,
Fattoruso SI, Arena MG, Pizzuti L, Giannarelli D, Paoletti G, Barba
M, et al: Docetaxel, oxaliplatin, and capecitabine combination
chemotherapy for metastatic gastric cancer. Gastric Cancer.
17:718–724. 2014. View Article : Google Scholar
|
|
6
|
Ruffell B and Coussens LM: Macrophages and
therapeutic resistance in cancer. Cancer Cell. 27:462–472. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Zou HY, Friboulet L, Kodack DP, Engstrom
LD, Li Q, West M, Tang RW, Wang H, Tsaparikos K, Wang J, et al:
PF-06463922, an ALK/ROS1 inhibitor, overcomes resistance to first
and second generation ALK inhibitors in preclinical models. Cancer
Cell. 28:70–81. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Bartel DP: MicroRNAs: Genomics,
biogenesis, mechanism, and function. Cell. 116:281–297. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Adams CM, Hiebert SW and Eischen CM: Myc
induces miRNA-mediated apoptosis in response to HDAC inhibition in
hematologic malignancies. Cancer Res. 76:736–748. 2016. View Article : Google Scholar :
|
|
10
|
Palmbos PL, Wang L, Yang H, Wang Y,
Leflein J, Ahmet ML, Wilkinson JE, Kumar-Sinha C, Ney GM, Tomlins
SA, et al: ATDC/TRIM29 drives invasive bladder cancer formation
through miRNA-mediated and epigenetic mechanisms. Cancer Res.
75:5155–5166. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Zhang H, Pu J, Qi T, Qi M, Yang C, Li S,
Huang K, Zheng L and Tong Q: MicroRNA-145 inhibits the growth,
invasion, metastasis and angiogenesis of neuroblastoma cells
through targeting hypoxia-inducible factor 2 alpha. Oncogene.
33:387–397. 2014. View Article : Google Scholar
|
|
12
|
Henao-Mejia J, Williams A, Goff LA, Staron
M, Licona-Limón P, Kaech SM, Nakayama M, Rinn JL and Flavell RA:
The microRNA miR-181 is a critical cellular metabolic rheostat
essential for NKT cell ontogenesis and lymphocyte development and
homeostasis. Immunity. 38:984–997. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Huang E, Liu R and Chu Y: miRNA-15a/16: As
tumor suppressors and more. Future Oncol. 11:2351–2363. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Xie SY, Li YJ, Wang PY, Jiao F, Zhang S
and Zhang WJ: miRNA-regulated expression of oncogenes and tumor
suppressor genes in the cisplatin-inhibited growth of K562 cells.
Oncol Rep. 23:1693–1700. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Voorhoeve PM, le Sage C, Schrier M, Gillis
AJ, Stoop H, Nagel R, Liu YP, van Duijse J, Drost J, Griekspoor A,
et al: A genetic screen implicates miRNA-372 and miRNA-373 as
oncogenes in testicular germ cell tumors. Cell. 124:1169–1181.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Xue J, Chi Y, Chen Y, Huang S, Ye X, Niu
J, Wang W, Pfeffer LM, Shao ZM, Wu ZH and Wu J: MiRNA-621
sensitizes breast cancer to chemotherapy by suppressing FBXO11 and
enhancing p53 activity. Oncogene. 35:448–458. 2016. View Article : Google Scholar
|
|
17
|
Xu C, Xie S, Song C, Huang L and Jiang Z:
Lin28 mediates cancer chemotherapy resistance via Regulation of
miRNA signaling. Hepatogastroenterology. 61:1138–1141. 2014.
|
|
18
|
Massoner P, Thomm T, Mack B, Untergasser
G, Martowicz A, Bobowski K, Klocker H, Gires O and Puhr M: EpCAM is
overexpressed in local and metastatic prostate cancer, suppressed
by chemotherapy and modulated by MET-associated miRNA-200c/205. Br
J Cancer. 111:955–964. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Wang Y, Liu C, Luo M, Zhang Z, Gong J, Li
J, You L, Dong L, Su R, Lin H, et al: Chemotherapy-induced
miRNA-29c/Catenin-δ signaling suppresses metastasis in gastric
cancer. Cancer Res. 75:1332–1344. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Kumar S, Keerthana R, Pazhanimuthu A and
Perumal P: Overexpression of circulating miRNA-21 and miRNA-146a in
plasma samples of breast cancer patients. Indian J Biochem Biophys.
50:210–214. 2013.PubMed/NCBI
|
|
21
|
Zhang HL, Yang LF, Zhu Y, Yao XD, Zhang
SL, Dai B, Zhu YP, Shen YJ, Shi GH and Ye DW: Serum miRNA-21:
Elevated levels in patients with metastatic hormone-refractory
prostate cancer and potential predictive factor for the efficacy of
docetaxel-based chemotherapy. Prostate. 71:326–331. 2011.
View Article : Google Scholar
|
|
22
|
Wang XC, Wang W, Zhang ZB, Zhao J, Tan XG
and Luo JC: Overexpression of miRNA-21 promotes
radiation-resistance of non-small cell lung cancer. Radiat Oncol.
8:1462013. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Gao W, Lu X, Liu L, Xu J, Feng D and Shu
Y: MiRNA-21: A biomarker predictive for platinum-based adjuvant
chemotherapy response in patients with non-small cell lung cancer.
Cancer Biol Ther. 13:330–340. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Song J, Bai Z, Zhang J, Meng H, Cai J,
Deng W, Bi J, Ma X and Zhang Z: Serum microRNA-21 levels are
related to tumor size in gastric cancer patients but cannot predict
prognosis. Oncol Lett. 6:1733–1737. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Dillhoff M, Liu J, Frankel W, Croce C and
Bloomston M: MicroRNA-21 is overexpressed in pancreatic cancer and
a potential predictor of survival. J Gastrointest Surg.
12:2171–2176. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Mydlarz W, Uemura M, Ahn S, Hennessey P,
Chang S, Demokan S, Sun W, Shao C, Bishop J, Krosting J, et al:
Clusterin is a gene-specific target of microRNA-21 in head and neck
squamous cell carcinoma. Clin Cancer Res. 20:868–877. 2014.
View Article : Google Scholar
|
|
27
|
Fulci V, Chiaretti S, Goldoni M, Azzalin
G, Carucci N, Tavolaro S, Castellano L, Magrelli A, Citarella F,
Messina M, et al: Quantitative technologies establish a novel
microRNA profile of chronic lymphocytic leukemia. Blood.
109:4944–4951. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Paik WH, Kim HR, Park JK, Song BJ, Lee SH
and Hwang JH: Chemosensitivity induced by down-regulation of
microRNA-21 in gemcitabine-resistant pancreatic cancer cells by
indole-3-carbinol. Anticancer Res. 33:1473–1481. 2013.PubMed/NCBI
|
|
29
|
Wei X, Wang W, Wang L, Zhang Y, Zhang X,
Chen M, Wang F, Yu J, Ma Y and Sun G: MicroRNA-21 induces
5-fluorouracil resistance in human pancreatic cancer cells by
regulating PTEN and PDCD4. Cancer Med. 5:693–702. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Valeri N, Gasparini P, Braconi C, Paone A,
Lovat F, Fabbri M, Sumani KM, Alder H, Amadori D, Patel T, et al:
MicroRNA-21 induces resistance to 5-fluorouracil by down-regulating
human DNA MutS homolog 2 (hMSH2). Proc Natl Acad Sci USA.
107:21098–21103. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Davies GF, Juurlink BH and Harkness TA:
Troglitazone reverses the multiple drug resistance phenotype in
cancer cells. Drug Des Devel Ther. 3:79–88. 2009.PubMed/NCBI
|
|
32
|
Schmittgen TD and Livak KJ: Analyzing
real-time PCR data by the comparative C(T) method. Nat Protoc.
3:1101–1108. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Clément T, Salone V and Rederstorff M:
Dual luciferase gene reporter assays to study miRNA function.
Methods Mol Biol. 1296:187–198. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Li SC, Liao YL, Ho MR, Tsai KW, Lai CH and
Lin WC: miRNA arm selection and isomiR distribution in gastric
cancer. BMC Genomics 1. 3(Suppl 1): S132012. View Article : Google Scholar
|
|
35
|
Oddo D, Sennott EM, Barault L, Valtorta E,
Arena S, Cassingena A, Filiciotto G, Marzolla G, Elez E, van Geel
RM, et al: Molecular landscape of acquired resistance to targeted
therapy combinations in BRAF-mutant colorectal cancer. Cancer Res.
76:4504–4515. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Zhao Y, Khanal P, Savage P, She YM, Cyr TD
and Yang X: YAP-induced resistance of cancer cells to antitubulin
drugs is modulated by a Hippo-independent pathway. Cancer Res.
74:4493–4503. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Mueller T, Voigt W, Simon H, Fruehauf A,
Bulankin A, Grothey A and Schmoll HJ: Failure of activation of
caspase-9 induces a higher threshold for apoptosis and cisplatin
resistance in testicular cancer. Cancer Res. 63:513–521.
2003.PubMed/NCBI
|
|
38
|
Chauhan D, Tian Z, Nicholson B, Kumar KG,
Zhou B, Carrasco R, McDermott JL, Leach CA, Fulcinniti M, Kodrasov
MP, et al: A small molecule inhibitor of ubiquitin-specific
protease-7 induces apoptosis in multiple myeloma cells and
overcomes bortezomib resistance. Cancer Cell. 22:345–358. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Konopleva M, Contractor R, Tsao T, Samudio
I, Ruvolo PP, Kitada S, Deng X, Zhai D, Shi YX, Sneed T, et al:
Mechanisms of apoptosis sensitivity and resistance to the BH3
mimetic ABT-737 in acute myeloid leukemia. Cancer Cell. 10:375–388.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Yan LX, Huang XF, Shao Q, Huang MY, Deng
L, Wu QL, Zeng YX and Shao JY: MicroRNA miR-21 overexpression in
human breast cancer is associated with advanced clinical stage,
lymph node metastasis and patient poor prognosis. RNA.
14:2348–2360. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Wu CW, Ng SS, Dong YJ, Ng SC, Leung WW,
Lee CW, Wong YN, Chan FK, Yu J and Sung JJ: Detection of miR-92a
and miR-21 in stool samples as potential screening biomarkers for
colorectal cancer and polyps. Gut. 61:739–745. 2012. View Article : Google Scholar
|
|
42
|
Lee HW, Lee EH, Ha SY, Lee CH, Chang HK,
Chang S, Kwon KY, Hwang IS, Roh MS and Seo JW: Altered expression
of microRNA miR-21, miR-155, and let-7a and their roles in
pulmonary neuroendocrine tumors. Pathol Int. 62:583–591. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Park SK, Park YS, Ahn JY, Do EJ, Kim D,
Kim JE, Jung K, Byeon JS, Ye BD, Yang DH, et al: MiR 21-5p as a
predictor of recurrence in young gastric cancer patients. J
Gastroenterol Hepatol. 31:1429–1435. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Yin C, Zhou X, Dang Y, Yan J and Zhang G:
Potential role of circulating MiR-21 in the diagnosis and prognosis
of digestive system cancer: A systematic review and meta-analysis.
Medicine (Baltimore). 94:e21232015. View Article : Google Scholar
|
|
45
|
Gong C, Yao Y, Wang Y, Liu B, Wu W, Chen
J, Su F, Yao H and Song E: Up-regulation of miR-21 mediates
resistance to trastuzumab therapy for breast cancer. J Biol Chem.
286:19127–19137. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Tomimaru Y, Eguchi H, Nagano H, Wada H,
Tomokuni A, Kobayashi S, Marubashi S, Takeda Y, Tanemura M,
Umeshita K, et al: MicroRNA-21 induces resistance to the
anti-tumour effect of interferon-alpha/5-fluorouracil in
hepatocellular carcinoma cells. Br J Cancer. 103:1617–1626. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Yang SM, Huang C, Li XF, Yu MZ, He Y and
Li J: miR-21 confers cisplatin resistance in gastric cancer cells
by regulating PTEN. Toxicology. 306:162–168. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Jin B, Liu Y and Wang H: Antagonism of
miRNA-21 Sensitizes Human Gastric Cancer Cells to Paclitaxel. Cell
Biochem Biophys. 72:275–282. 2015. View Article : Google Scholar
|
|
49
|
Zheng P, Chen L, Yuan X, Luo Q, Liu Y, Xie
G, Ma Y and Shen L: Exosomal transfer of tumor-associated
macrophage-derived miR-21 confers cisplatin resistance in gastric
cancer cells. J Exp Clin Cancer Res. 36:532017. View Article : Google Scholar : PubMed/NCBI
|
|
50
|
Wang S, Wang A, Shao M, Lin L, Li P and
Wang Y: Schisandrin B reverses doxorubicin resistance through
inhibiting P-glycoprotein and promoting proteasome-mediated
degradation of survivin. Sci Rep. 7:84192017. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Cao Z, Liang N, Yang H and Li S: Visfatin
mediates doxorubicin resistance in human non-small-cell lung cancer
via Akt-mediated up-regulation of ABCC1. Cell Prolif. 50:2017.
View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Papagiannakopoulos T, Shapiro A and Kosik
KS: MicroRNA-21 targets a network of key tumor-suppressive pathways
in glioblastoma cells. Cancer Res. 68:8164–8172. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
53
|
Zhu S, Si ML, Wu H and Mo YY: MicroRNA-21
targets the tumor suppressor gene tropomyosin 1 (TPM1). J Biol
Chem. 282:14328–14336. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Sayed D, He M, Hong C, Gao S, Rane S, Yang
Z and Abdellatif M: MicroRNA-21 is a downstream effector of AKT
that mediates its antiapoptotic effects via suppression of Fas
ligand. J Biol Chem. 285:20281–20290. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Blumenthal GM and Dennis PA: Pten
hamartoma tumor syndromes. Eur J Hum Genet. 16:1289–1300. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Kennedy SG, Wagner AJ, Conzen SD, Jordán
J, Bellacosa A, Tsichlis PN and Hay N: The pi3-kinase/akt signaling
pathway delivers an anti-apoptotic signal. Genes Dev. 11:701–713.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Yea SS and Fruman DA: Achieving cancer
cell death with PI3K/mtor-targeted therapies. Ann N Y Acad Sci.
1280:15–18. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Westhoff MA, Faham N, Marx D, Nonnenmacher
L, Jennewein C, Enzenmuller S, Gonzalez P, Fulda S and Debatin KM:
Sequential dosing in chemo-sensitization: Targeting the
PI3K/akt/mtor pathway in neuroblastoma. PLoS One. 8:e831282013.
View Article : Google Scholar
|
|
59
|
Defamie V, Sanchez O, Murthy A and Khokha
R: TIMP3 controls cell fate to confer hepatocellular carcinoma
resistance. Oncogene. 34:4098–4108. 2015. View Article : Google Scholar
|
|
60
|
Han J and Sun P: The pathways to tumor
suppression via route p38. Trends Biochem Sci. 32:364–371. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Bulavin DV and Fornace AJ Jr: p38 MAP
kinase's emerging role as a tumor suppressor. Adv Cancer Res.
92:95–118. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Rangarajan A, Talora C, Okuyama R, Nicolas
M, Mammucari C, Oh H, Aster JC, Krishna S, Metzger D, Chambon P, et
al: Notch signaling is a direct determinant of keratinocyte growth
arrest and entry into differentiation. EMBO J. 20:3427–3436. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Shibata T, Watari K, Izumi H, Kawahara A,
Hattori S, Fukumitsu C, Murakami Y, Takahashi R, Toh U, Ito KI,
Ohdo S, et al: Breast cancer resistance to antiestrogens is
enhanced by increased ER degradation and ERBB2 expression. Cancer
Res. 77:545–556. 2017. View Article : Google Scholar
|
|
64
|
Schneider C, Oellerich T, Baldauf HM,
Schwarz SM, Thomas D, Flick R, Bohnenberger H, Kaderali L, Stegmann
L, Cremer A, et al: SAMHD1 is a biomarker for cytarabine response
and a therapeutic target in acute myeloid leukemia. Nat Med.
23:250–255. 2017. View Article : Google Scholar
|
|
65
|
Feng DD, Zhang H, Zhang P, Zheng YS, Zhang
XJ, Han BW, Luo XQ, Xu L, Zhou H, Qu LH and Chen YQ: Down-regulated
miR-331-5p and miR-27a are associated with chemotherapy resistance
and relapse in leukaemia. J Cell Mol Med. 15:2164–2175. 2011.
View Article : Google Scholar
|
|
66
|
Kimura A, Ogata K, Altan B, Yokobori T,
Ide M, Mochiki E, Toyomasu Y, Kogure N, Yanoma T, Suzuki M, et al:
Nuclear heat shock protein 110 expression is associated with poor
prognosis and chemotherapy resistance in gastric cancer.
Oncotarget. 7:18415–18423. 2016. View Article : Google Scholar : PubMed/NCBI
|