According to statistics published by the World
Health Organization, cardiovascular disease (CVD) is a leading
cause of mortality and morbidity worldwide (1). Between 1980 and 2014, the prevalence
of obesity worldwide doubled. Currently, nearly 40% of adults aged
≥18 years old, (~2 billion people) are overweight and ~13% (~600
million) are obese (2).
Furthermore, 41 million children aged <5 years old are
overweight or obese (3).
According to statistics compiled by the Center for Disease Control
and Prevention, ~610,000 people succumb to heart disease annually
in the US alone, representing 1 out of every 4 mortalities
(4). Various environmental and
behavioral factors may increase the risk of CVD, however, a
prevailing body of evidence suggests that contemporary lifestyles,
comprising sedentary lifestyles, low energy expenditure together
with overnutrition and the inadequate composition of nutrition are
primary reasons behind the increase in rates of obesity (5–8).
Obese individuals exhibit excessive adipose tissue and dyslipidemia
and it has been suggested that these conditions may cause many
chronic diseases, including metabolic syndrome, hypertension,
hypercholesterolemia and diabetes, all of which are major risk
factors for CVD (5–8). Excessive adiposity is accompanied by
the accumulation of activated macrophages, which release
proinflammatory cytokines, including tumor necrosis factor α,
interleukin-6 (IL-6) and IL-1 (9,10).
These pathological conditions increase endogenous formation of
blood cholesterol and its subsequent deposition in blood vessel
walls (11,12). Identifying ways of reducing
elevated low density lipoprotein cholesterol (LDL-C) levels is
therefore a key public health challenge and introducing novel
dietary supplements may be an important method of achieving this
(13).
A number of studies aiming to develop novel methods
of regulating cholesterol levels are currently in progress. It has
been proposed that products of natural origin, including
glucomannans, plant sterols, berberine or red yeast rice, which are
known as nutraceuticals, can be used either alone (14) or in combination with commonly used
statins, such as 3-hydroxy-3-methylglutaryl coenzyme A inhibitors.
Although statins represent an important method of regulating
cholesterol levels, they may induce significant side effects on
skeletal muscle, including myalgia and rhabdomyolysis (15,16), as well as on the homeostasis of
bowel microbiota, including dysbiosis with intestinal dysfunction
(17,18).
Prebiotics, such as dietary fiber, have been
evaluated due to their ability to modulate intestinal microbiota.
They may be able to improve the selection of protective bacteria
such as Lactobacillus and Enterococcus and therefore
help to prevent cancer (19–21). Studies have suggested that the
risk of colorectal cancer is increased by consumption of processed
and unprocessed meat, but is decreased suppressed by consumption of
fiber. Food composition may affect cancer risk due to its effects
on the metabolism of colon microbes. The gut microbiota ferment
complex dietary residues that are resistant to digestion by enteric
enzymes. This process provides energy for the microbiota but
culminates in the release of short-chain fatty acids (22).
However, it has also been demonstrated that they
help to regulate hyperlipidemia (23,24). There are two types of dietary
fiber: Soluble fiber, including pectin, fructo-oligosaccharides and
oat β-glucan; and insoluble fiber, including cellulose found in
whole wheat bread and brown rice and certain types of β-glucans,
found in yeast and mushrooms. These fibers, via their digestion or
fermentation by intestinal microbiota may affect cholesterol
metabolism by acting on bile acids and affecting lipid absorption
(25,26). They may also influence the local
microbial environment through bacterial selection by butyrate
production. Therefore, they may affect the colonic mucosa as well
as mucosal and systemic immunity, including mucosal repair in
chronic inflammation and the reduction of proinflammatory cytokines
(28–32).
Among different types of fiber, β-glucans have
attracted particular interest due to their well-documented
activities as modulators of immunity (33,34). The immunomodulatory activities
involve anti-infectious immunity, anti-cancer immunity and all
aspects of cellular and humoral defense reactions. In addition,
β-glucans are nutraceuticals able to decrease cholesterol levels
(35,36). Therefore, β-glucans may be the
best type of soluble fiber for dietary supplementation and to
improve general health. The aim of the current review is to assess
the most up-to-date information regarding various aspects of
dietary fibers and β-glucans in order to evaluate their potential
in regulating cholesterol levels.
Natural products that may prevent or treat various
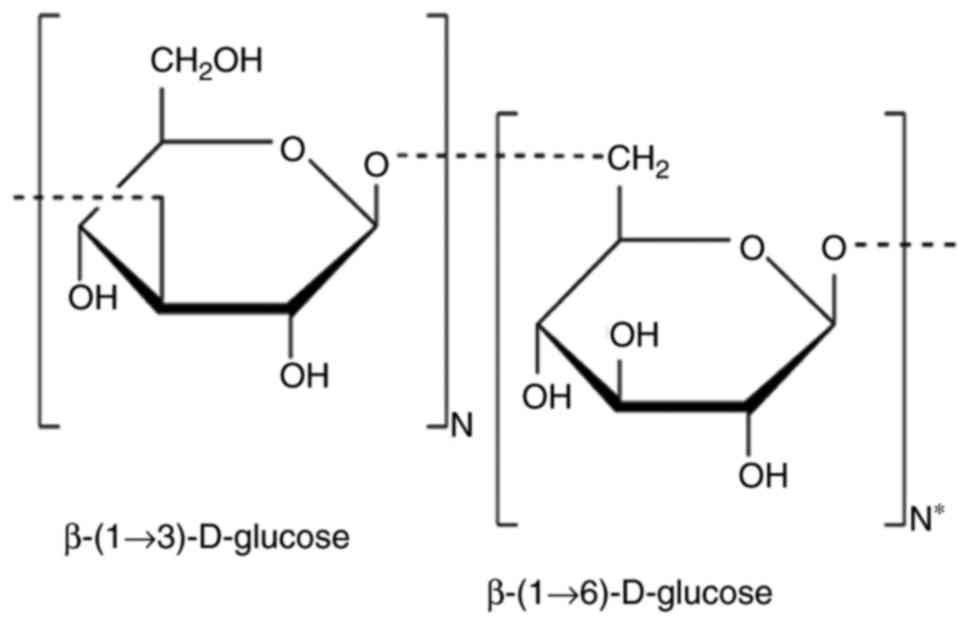
diseases have been identified, including β-glucans. Chemically,
β-glucans are heterogeneous non-starch polysaccharides, which form
the structural compounds of the cell wall of certain
microorganisms, including yeast and algae, and certain protists,
including mushrooms and grains, such as oats and wheat. β-glucans
may be insoluble or soluble. Insoluble β-glucans fibers consist of
β-(1,3/1,4)-D-linked glucose units, whereas soluble viscous
β-glucans fibers consist of β-(1,3)/1,6)-D-linked glucose (37). A schematic representation of the
basic molecular structure of β-glucan is presented in Fig. 1.
It has been demonstrated that β-glucans reduce
levels of a non-high-density lipoprotein-cholesterol (non-HDL-C)
fraction that contains LDL-C without affecting HDL-C or
triglyceride levels (35,38–40). β-glucans have been used to reduce
blood cholesterol levels since the 1960s (41). β-glucans are also used to enhance
the efficiency of the immune system (42,43).
Research conducted over the previous 40 years has
established the biological effects of β-glucan. Various studies
have indicated that β-glucans represent important substances to be
included, among other substances, within the biological response
modifiers, which are substances that can modulate immune activity
(44), as they exhibit strong
immunomodulatory properties (45). These immunomodulatory properties
primarily depend on the source they have been extracted from;
studies conducted in animals and humans and confirmed that the oral
administration of yeast β-glucans had a greater effect on the
immune system than grain-derived β-glucans (45–48). β-glucan is active in all species
from earthworms to humans, and is able to stimulate humoral and
cellular immunity (45),
metabolically regulate diabetes (49), stimulate wound healing (50), reduce psychophysical stress
(51), attenuate chronic fatigue
syndrome (52) and inhibit the
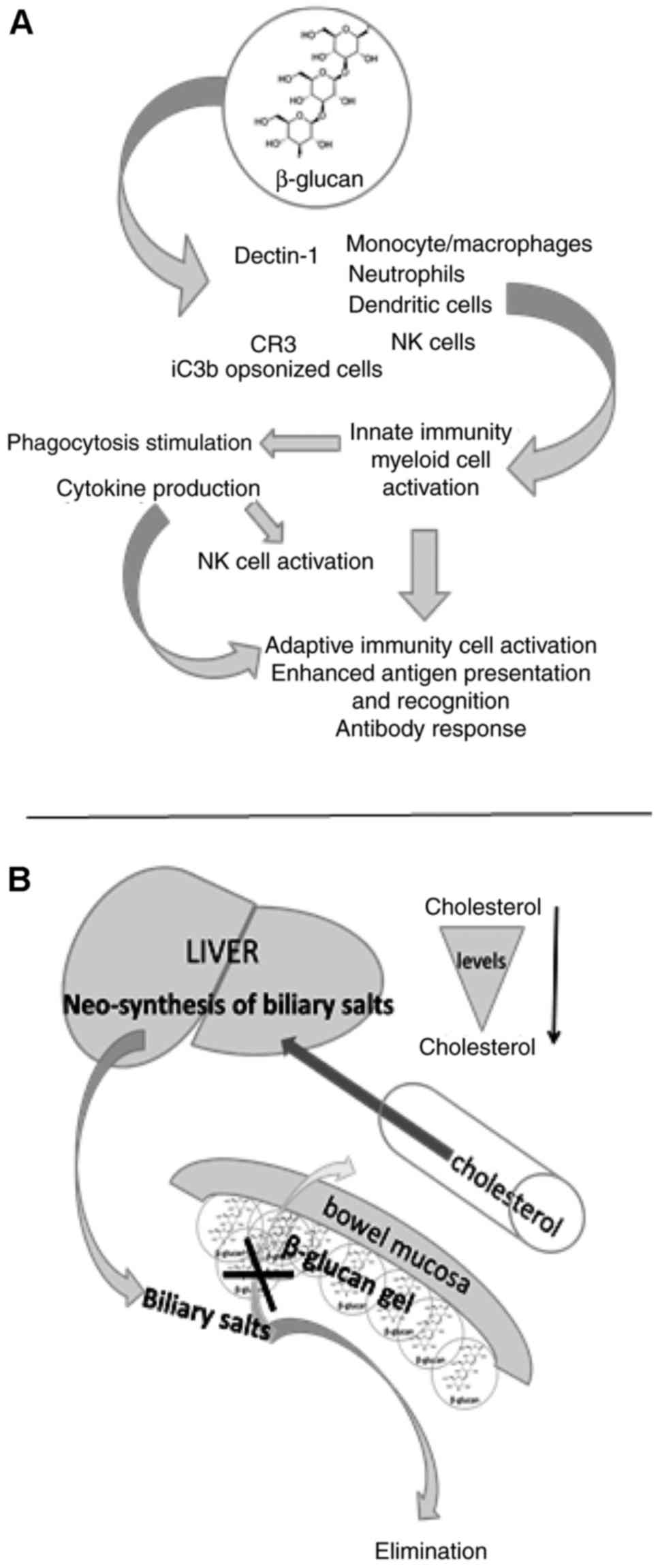
development of cancer (53,54). The effects of β-glucan on the
immune system are summarized in Fig.
2A. Clinical trials into the effects of β-glucan on the immune
system have now been conducted (55,56) and it has been demonstrated that
β-glucans not only affect the immune system but may also reduce
cholesterol levels. Different types of β-glucans, not only
β-glucans found in oats, are able to reduce cholesterol levels.
Initially, studies investigating the effects of
β-glucans assessed insoluble β-glucans; studies evaluating
β-glucan-receptor interactions were conducted later. Among the most
important receptors that interact with β-glucans are dectin-1,
complement receptor type 3 [CR3; also known as cluster of
differentiation (CD)11b/CD18] and langerin. Dectin-1 is a type II
transmembrane protein present in leukocytes, with the highest
levels of expression in neutrophils, macrophages and dendritic
cells (57). Langerin is composed
of two main parts: A carbohydrate-recognizing domain and a region
that can form trimmers and this receptor is most commonly expressed
in Langerhans cells (58). CR3 is
a receptor that forms part of the integrin β2 family. It is mostly
found on neutrophils, macrophages and natural killer cells, but is
also present on bone marrow progenitor cells (59). The special configuration assumed
by the CR3 receptor following interaction with complement iC3b on
opsonized target cells mediates cytotoxic activation (60). The importance of CR3 in β-glucan
binding and action was confirmed by the resistance of CR3-knockout
mice to β-glucan (61). More
detailed information regarding the molecular interactions of
β-glucans with their receptors has been documented in a recent
comprehensive review (62).
Experimental studies investigating the signaling
pathways activated following β-glucan-cell interaction indicated
that nuclear factor NF-κB was activated via inhibitor of NF-κB
kinase β activity and following alteration of the phosphorylation
and degradation of factor of k light polypeptide gene enhancer in
B-cells inhibitor a (IkBa) (63).
β-glucan may alter the response to polymicrobial sepsis via
modulation of IKK3 kinase activity with subsequent decreases in the
phosphorylation of IkBa and activation of NF-κB. In addition, the
involvement of tyrosine-protein kinase SYK (Syk) was confirmed on
the dectin-1 receptor. β-glucan binding is followed by the
phosphorylation of dectin-1 by proto-oncogene tyrosine-protein
kinase Src. As a result, Syk is activated and subsequently
activates the CARD9-Bc110-Malt1 complex (64). It was demonstrated that Syk
mediates the activation of downstream signaling; incusing the
extracellular-regulated kinase 1/2 and p38a mitogen-activated
protein kinase (MAPK) cascade; activating mitogen and stress
activated protein kinases 1 and 2 and subsequently phosphorylating
the cAMP response element binding protein. The involvement of the
extracellular-regulated kinase 1/2 and p38a mitogen-activated
protein kinase (MAPK) pathways was identified (65). Furthermore, β-glucan and its
associated signaling pathways have been evaluated previously
(66). However, current
understanding of the intracellular signaling that occurs following
β-glucan binding remains incomplete. There may either be too many
competing receptors able to bind β-glucan or different types of
β-glucans isolated from several different sources may have been
used in previous studies, complicating the results.
Cholesterol is a source of biologically active
substances important for the survival of organisms and serves a
pivotal role in the body; therefore, cholesterol should not be
unilaterally condemned. Together with other phospholipid
substances, cholesterol forms the basic structural components of
cell membranes and participates in their biochemical activity. To a
lesser extent, cholesterol is also located in membranes of
intracellular organelles and in lipoproteins found in the blood
plasma, where 70% of cholesterol is esterified with fatty acids to
form cholesteryl esters. The brain tissue contains ~25% of
cholesterol in the body (67).
Cholesterol is synthetized in the liver and is essential for the
formation of bile acids, steroid hormones and vitamin D (68). It then becomes involved in
regulating energy metabolism and overall energy expenditure; for
example, it promotes adaptive thermogenesis following exposure to
cold via its conversion to bile acids and by shaping the gut
microbiome (69). Primary bile
salts are produced in the liver via the oxidization of cholesterol
to produce cholic acid and chenodeoxycholic acid. These products
are then conjugated with taurine and glycine to produce taurocholic
and glycocholic acids, and once they are in the bile, they are
utilized in lipid metabolism and energy expenditure, together with
pancreatic lipase (70).
Exogenous cholesterol enters the body with food of
animal origin. It binds to LDL and enters the cells via receptor
endocytosis, which involves the binding of LDL-C to cell surface
receptors, which are subsequently engulfed (71). The enterohepatic circulation of
bile acids is physiologically significant. Bile acids are
efficiently reabsorbed from the intestine into the liver and only
5–20% of these bile acids are lost in the feces. If larger
quantities of bile acids are lost, a corresponding amount must be
synthesized de novo in the liver (72) and as a source of newly synthetized
bile acids, it lowers blood cholesterol levels. Therefore, it is
important to bear in mind that all processes leading to the
increased fecal excretion of bile acids reduce blood cholesterol
levels. The enterohepatic circulation of bile acids is decreased by
means of free calcium ions, primarily found in milk, which form
insoluble compounds with intestinal bile acids, known as 'soap
substances'. This leads to the potential lowering of circulating
cholesterol levels, resulting in the neo-synthesis of bile acids in
the liver (73). It has been
proposed that dietary fibers of various origins, including
β-glucans, bind to enteral bile acids and thereby promote their
excretion into feces. However, their link to cholesterol is based
on a different principle (Fig.
2B).
Cholesterol is a molecule essential for all animal
life; therefore every cell in the body is able to synthesize
cholesterol via a complex process starting with the mevalonate
pathway and ending with lanosterol conversion. Cholesterol is an
important substance that exhibits beneficial biological properties,
including as a constituent of cellular membranes and a base for
steroid hormones and biliary salt synthesis. There are more forms
of cholesterol that exist in the organism but the most important
molecules regarding health are LDL-C and HDL-C. Other types of
lipoproteins present in the blood include chylomicrons,
very-low-density lipoproteins (VLDL) and intermediate-density
lipoproteins (IDL) (74). The
cholesterol found in different lipoproteins is identical, although
some exists in its native free alcohol form, whereas others exist
as fatty acyl esters. Lipoprotein particles are organized by
complex apolipoproteins, which are bound by specific receptors
expressed on cell membranes (75). It is important to note that,
whereas cholesterol and particularly LDL are usually considered to
be the most harmful molecules, non-HDL-C has become a commonly used
marker to identify a blood lipid pattern associated with an
increased risk of heart disease (76). Non-HDL-C is inversely correlated
with HDL-C in children and adults (77,78). Indeed, it has been suggested that
non-HDL-C may replace LDL-C as the primary therapeutic target to
treat patients with hypercholesterolemia (79). The National Cholesterol Education
Program currently recommends that reducing non-HDL-C levels in
patients with hypercholesterolemia should be a secondary
lipid-lowering target (80).
Reducing non-HDL-C levels is becoming an increasingly important
target, as the incidence of visceral obesity, diabetes mellitus and
metabolic syndrome continues to increase. HDL levels are inversely
correlated with atherosclerotic disease; therefore efforts are
underway to increase HDL-C and major apolipoprotein of HDL (apoAI)
levels. Potential new treatments, including inhibiting the p38 MAPK
pathway or administrating apolipoprotein mimetics, were found to be
effective at lowering cholesterol in mice and humans (81,82).
Cholesterol can only harm organisms when its
concentrations are elevated in the blood, particularly when levels
of its oxidized LDL form (ox-LDL) are elevated (83). This may cause long-term chronic
inflammation in the artery wall, resulting in the increased storage
of cholesterol. HDL, which transports cholesterol to the liver, has
the opposite effect and reduces the atherogenic action of ox-LDL
(84). In humans (85) and some animals (86,87), atherosclerosis is a disease that
evolves following the induction of focal lesions, resulting in
atheroma plaque formation at the arterial level in different areas
of the body. It is an important cause of coronary vessel
obstruction and coronary artery disease, also known as ischemic
heart disease (88,89). Two reviews by Gistera and Hansson
(90), and by Jonsson and Backhed
(91) discuss the complexity of
atheroma lesion development, including its sequence of events and
underlying mechanisms of action, such as chronic inflammation and
macrophage activity fueled by increased cholesterol levels, as well
as the influence of the microbiome on atherogenesis.
None of the cholesterol lowering drugs currently
used are free of side effects. By contrast, it is generally
accepted that different types of dietary fiber, including
β-glucans, have positive effects on health without inducing
significant side effects (92).
The results of experimental studies in animals and clinical trials
in humans have demonstrated that dietary fiber supplements not only
improve general health but also reduce the risk of various chronic
diseases associated with lifestyle, including CVD, cancer and type
2 diabetes (93), which all have
a large impact on public health and society in general. Due to
their variety, accurately defining the different types of dietary
fibers has been challenging. This is exemplified by the 10-year
process that was required to obtain an international legal
definition for dietary fiber (https://www.fsis.usda.gov/wps/portal/fsis/topics/international-affairs/us-codex-alimentarius/recent-delegation-reports/delegate-report-31st-session-ccnfsdu/ct_index)
by the Codex Alimentarius Commission (94).
Previously, it was considered that soluble dietary
fiber lowered levels of lipids and cholesterol in the blood and
that insoluble dietary fiber only contributed to fecal bulking.
However, it has now been demonstrated that these effects depend on
the degree of viscosity and fermentation capability of fiber
(95). The definition of dietary
fiber has therefore been extended to include all types of
carbohydrates and other resistant substances, such as starches,
with similar properties. Soluble dietary fiber that resists
enzymatic digestion in the small intestine is not absorbed; it is
partially or fully fermented in the large intestine (38,96).
The hypocholesterolemic effects of dietary fiber are
still not fully understood. It has been attributed to the ability
of soluble dietary fiber to form viscous solutions that prolong
gastric emptying, inhibit the transport of triglycerides and
cholesterol across the intestine and reduce total LDL-lipoprotein
concentrations (97). The
consequences of the increased viscosity of the luminal contents
manifest via the amplification of the thickness of the water layer
and in the decrease of cholesterol uptake from the intestinal lumen
(98).
Dietary fiber is able to bind to bile acids,
monoglycerides, free fatty acids and cholesterol. Dietary fiber
also decreases absorption and increases the fecal excretion of
these chemical substances (99,100). The structure of insoluble
dietary fiber enables them to directly bind to bile acids and they
may lower blood cholesterol levels in this manner, whereas soluble
dietary fiber may increase the viscosity of the chyme, thus
reducing bile acid diffusion (101). Dietary fiber fractions from
mushrooms are also able to modulate cholesterol-related gene
expression (34). Furthermore, it
has been suggested that treatment with β-glucans prevents the
development of fatty liver (102).
Dietary fiber influences the expression of genes
that regulate lipid and carbohydrate metabolism. It has been
demonstrated that resistant starch, fructans, inulin and β-glucans
affect the phosphorylation of AMP-activated protein kinase (AMPK),
which is a key enzyme involved in energy exchange (103,104). Furthermore, it is well known
that higher concentrations of blood HDL protect against CVD. ApoAI
activates AMPK in the arterial endothelia and increases the
phosphorylation and activity of nitrogen oxide synthase (105).
Anaerobic gut bacteria in the cecum and large
intestine produce short chain fatty acids (SCFAs) as the end
products of the fermentation of non-digestible carbohydrates that
pass through the small intestine unaffected (106,107). These gut-generated SCFAs exert
multiple effects on energy metabolism and on immunity (108–110). SCFAs influence the immune system
via G-protein-coupled free fatty acid receptors (FFAs) 2 and 3.
FFA2 is expressed primarily in leukocytes and colonic L-cells and
also partially expressed in adipocytes, whereas FFA3 is primarily
expressed in adipocytes (111).
β-glucans may interact directly with immunocompetent cells, such as
mucosal macrophages and dendritic cells, which exhibit pattern
recognition receptors with carbohydrate-binding domains,
subsequently inducing a decrease in IL-12 and increase in IL-10
production by modulation of toll-like receptor signaling (110,112); this is consistent with an
anti-inflammatory phenotype. By contrast, the influence of dietary
fiber on metabolism and immunity may be indirect; for example, they
may act as a source of SCFAs (28,113). The uptake of cholesterol
together with fatty acids in the gut into the enterocytes may be
accomplished by passive transport and by means of transporter
proteins (114,115). Three key transcription factors,
known as sterol regulatory element binding proteins 1a, 1c and 2,
determine the expression of genes that regulate the synthesis of
cholesterol and fatty acids (116,117). Dietary fiber may downregulate
the expression of genes for these transporter proteins: Sterol
regulatory element binding transcription factor (SREBF)1,
which regulates lipid metabolism and SREBF2, which regulates
cholesterol synthesis (36,118,119). Therefore, among the potential
mechanisms involved in lowering LDL levels in the blood by dietary
fiber, the reduced adsorption of biliary salts from the bowel by
β-glucans, either by direct glucanbiliary salt complexes or the
formation of a viscous layer inhibiting absorption, are supported
by the majority of studies (40,41,119). Indeed, the neosynthesis of
biliary salts by the liver induces the utilization of the hepatic
free cholesterol. Lowering cholesterol levels induces the
upregulation of 7-α-hydroxylase, 3-hydroxy-3-methylglutaryl
coenzyme A reductase and LDL-C receptors to restore the original
conditions (120). This may
explain the lowering of LDL by β-glucans and other dietary fibers
reported in various studies, although some meta-analyses report
that β-glucans affect HDL and LDL cholesterol (35,118–121). At the same time, β-glucans may
downregulate the proinflammatory signals elicited by altered levels
of cholesterol.
To more effectively understand the mechanisms that
underline human CVD, suitable animal models are required. Although
there are currently no ideal animal models, the comparative
investigation of existing models may improve understanding of the
sequential onset of the pathological alterations; from the
initiation of the disease to the final stages of atherosclerotic
plaque development. It has been demonstrated that increasing
dietary fiber intake of diabetic animals may reduce the intestinal
absorption of nutrients, such as glucose (97,122). Kirby et al (123) indicated that serum cholesterol
concentration was reduced by 13% and plasma LDL-C levels were
reduced by 14%, whereas HDL-C concentrations remained unchanged in
hypercholesterolemic men on a diet supplemented with oat bran. The
hypocholesterolemic effects of β-glucans have been confirmed in
broiler chicks, which were fed a diet supplemented with barley
(124). Furthermore, feeding
diabetic rats a high fiber diet supplemented with barley
significantly improved plasma glucose concentrations and decreased
cholesterol levels compared with rats fed low and very low fiber
diets consisting of rice and cornstarch, respectively (125). Golden Syrian hamsters are a
suitable animal model to study the vascular changes that occur
during atherogenesis (126).
These hamsters have a number of advantages: As in humans, their
primary cholesterol carrier in the plasma is LDL, their lipoprotein
metabolism is similar to that of humans 84 and their LDL receptor
gene is similar to the LDL gene in humans. Lim et al
(127) applied β-glucans from
yeast-like fungus Aureobasidium pullulans, known as polycan,
to a hamster experimental animal model of hyperlipemia induced by a
high-fat diet. The authors concluded that polycan decreased the
atherosclerotic effects, hyperlipemia and hepatic damage induced by
a high-fat diet. Previous studies on a hamster model of
hypercholesterolemia using β-glucans isolated from oat and barley
also identified that they induce beneficial hypolipidemic and
anti-atherosclerotic effects (128,129). Another study found that β-glucan
treatment reduced the inflammation induced by a diet high in
cholesterol (130). In a rat
experimental model, β-glucans reduced total cholesterol,
triglyceride and malondialdehyde levels by 42% (131). Furthermore, using hamsters as a
model, Tong et al (132)
found that dietary β-glucans reduced the concentration of plasma
LDL-C by promoting the excretion of fecal lipids and regulating the
activities of 3′-hydroxy-3-methyl-glutaryl-coenzyme A reductase and
CEP7A1. In a study by the current authors, yeast-derived β-glucan
300 markedly lowered total cholesterol levels in mice with
experimentally-induced hypercholesterolemia. Supplementation with
three other β-glucans, Krestin, ImmunoFiber and Now glucan, induced
similar, albeit decreased effects (133). To the best of our knowledge,
this study was the first to directly compare the effects of several
different β-glucans. As the most effective glucan was
yeast-derived, it supports the claim that such results following
increased fiber intake may be explained by a subsequent decrease in
bile acid absorption (134). To
the best of our knowledge, no studies have yet investigated the
effects of β-glucans on cholesterol levels in apolipoprotein-E
knockout mice (135,136).
Long-term clinical studies investigating the effects
of soluble forms of β-glucans in men with hypercholesterolemia
demonstrated that they decrease blood cholesterol levels (137–139). In 1989, Newman et al
(140) demonstrated that certain
barley cultivars, which are rich in water-soluble fiber, have a
hypocholesterolemic effect in men. Several volunteer men were
randomly assigned to add wheat or barley bran diet supplements to
their normal diets for 28 days.
Those who consumed wheat supplements exhibited
significantly increased serum total cholesterol and LDL-C levels
compared with pretreatment. The cholesterol levels of subjects that
received barley supplements who had average cholesterol levels
pretreatment were not significantly altered; however, triglyceride
and LDL-C levels were reduced in subjects that had increased
cholesterol levels pretreatment. Studies published over the past 20
years have demonstrated that LDL-C levels are reduced following the
supplementation of diets with β-glucans in humans with increased
blood concentrations of triglycerides and cholesterol (119,120,141–151); however, it has not been
confirmed that these effects are caused by β-glucans (152–154). Ho et al (35) published a systematic review and
meta-analysis of 14 trials involving 615 healthy and
hypercholesterolemic participants, summarizing the evidence that
barley β-glucans decrease LDL-C but not HDL-C levels. Diets
supplemented with a median dose of 6.5 and 6.9 g/day β-glucans
decreased LDL-C and non-HDL-C levels by 7% compared with control
diets. Two previous meta-analyses have confirmed the
cholesterol-lowering effect of barley β-glucans. One reported a
reduction in LDL-C levels by 0.26 mmol.l−1 following a
median β-glucan dose of 7 g/day based on the results of seven
studies (155). A second
meta-analysis, published in 2010, demonstrated that LDL-C was
lowered by 0.27 mmol.l−1 following a median β-glucan
dose of 5 g/day (156).
It is generally accepted that barley β-glucans
reduce apolipoprotein B levels in patients with
hypercholesterolemia (157).
However, further studies are required to investigate the effects of
β-glucans in patients with different metabolic phenotypes, as well
as on the content of β-glucans in barley, which depends on the
genotype of the plant, and on the environmental conditions and
composition of the soil the plant grows in (158). Treatment with an average dose
osf 6.5 g/day barley β-glucan for ≥4 weeks may have beneficial
effects in individuals with and without elevated blood
concentrations of cholesterol (35,159). A further source of β-glucans
suitable for lowering blood cholesterol concentrations may include
different species of fungi. The underlying mechanisms of β-glucans
from mushrooms have not yet been explained. It has been repeatedly
confirmed that the consumption of fungi decreases blood cholesterol
levels in animals (160,161) and the increased intake of fungi
may reduce CVD risk. Clinical trials have also confirmed that the
consumption of soluble β-glucans reduces total cholesterol levels,
including LDL-C, without affecting HDL-C and triglyceride levels
(38,39,118). A previous study conducted in a
murine model (162) demonstrated
that the novel insoluble dietary fiber chitin-glucan, which is
extracted from the cell wall of fungi and combined with olive oil,
associated with the Mediterranean diet, which is known for its
beneficial effects on health (163), substantially reduces levels of
Ox-LDL suggesting that this combination may be used as a
therapeutic strategy to treat patients with CVD. Furthermore, in
patients with mild hypercholesterolemia it has been demonstrated
that high-molecular weight β-glucans decrease serum cholesterol
levels differently than low-molecular weight β-glucans (164). Beside differences in viscosity,
CYP7A1rs3808607 polymorphism also contributed to the different
effects of these two tested glucans (164).
Over the past several decades, it has been suggested
that increasing levels of fiber, including β-glucans, in the diet
leads to a reduction in cholesterol levels. However, the precise
mechanisms of this action remain unclear. Further research is
required to more specifically define the role of β-glucan in
modulating the immunological aspects of atherosclerosis. The
current review strengthens the hypotheses that of all forms of
dietary fiber, natural β-glucan molecules are the most promising to
use as a method of treating patients with dyslipidemia. It remains
unclear whether increasing β-glucan intake should be recommended to
patients with severe hypercholesterolemia, to be used in addition
to or even as an alternative to statins, as there have been very
few direct comparisons between the effects of using β-glucans and
statins on such patients. However, β-glucans may more effective as
they have a different mechanism of action than statins; while
statins block the action of the liver enzyme responsible for
producing cholesterol, β-glucans promote a more
physiologically-based rebalancing of cholesterol levels.
The authors wish to acknowledge the support of the
Institute of Microbiology (CZ; grant no. RVO 61388971) and
UniCredit Bank (CZ) and ARPA Foundation, Pisa (IT) donations.
The authors declare that they have no competing
interests.
|
1
|
World Health Organization: Cardiovascular
disease. 2016
|
|
2
|
World Health Organization: Obesity and
overweight fact sheet. 2017
|
|
3
|
World Health Organization: Obesity and
overweight fact sheet. 2016
|
|
4
|
CDC, National Center for Chronic Disease
Prevention and Health Promotion and Division for Heart Disease and
Stroke Prevention: Heart Disease Facts and Statistics. CDC;
|
|
5
|
Murray CJ and Lopez AD: Alternative
projections of mortality and disability by cause 1990–2020: Global
Burden of Disease Study. Lancet. 349:1498–1504. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Oreopoulos A, Padwal R, Kalantar-Zadeh K,
Fonarow GC, Norris CM and McAlister FA: Body mass index and
mortality in heart failure: A meta-analysis. Am Heart J. 156:13–22.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Clark JM and Brancati FL: The challenge of
obesity-related chronic diseases. J Gen Intern Med. 15:828–829.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Oreopoulos A, Ezekowitz JA, McAlister FA,
Kalantar-Zadeh K, Fonarow GC, Norris CM, Johnson JA and Padwal RS:
Association between direct measures of body composition and
prognostic factors in chronic heart failure. Mayo Clin Proc.
85:609–617. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bose KS, Gupta SK and Vyas P:
Adipocytokine levels in genetically high risk for type 2 diabetes
in the Indian population: A cross-sectional study. Exp Diabetes
Res. 2012:3865242012. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Olson NC, Callas PW, Hanley AJ, Festa A,
Haffner SM, Wagenknecht LE and Tracy RP: Circulating levels of
TNF-α are associated with impaired glucose tolerance, increased
insulin resistance, and ethnicity: The insulin resistance
atherosclerosis study. J Clin Endocrinol Metab. 97:1032–1040. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Libby P, Ridker PM and Maseri A:
Inflammation and atherosclerosis. Circulation. 105:1135–1143. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Emanuela F, Grazia M, Marco de R, Maria
Paola L, Giorgio F and Marco B: Inflammation as a link between
obesity and metabolic syndrome. J Nutr Metab. 2012:4763802012.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Harland JI: Food combinations for
cholesterol lowering. Nutr Res Rev. 25:249–266. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Johnston TP, Korolenko TA, Pirro M and
Sahebkar A: Preventing cardiovascular heart disease: Promising
nutraceutical and non-nutraceutical treatments for cholesterol
management. Pharmacol Res. 120:219–225. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Chatzizisis YS, Koskinas KC, Misirli G,
Vaklavas C, Hatzitolios A and Giannoglou GD: Risk factors and drug
interactions predisposing to statin-induced myopathy: Implications
for risk assessment, prevention and treatment. Drug Saf.
33:171–187. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Camerino GM, Musumeci O, Conte E, Musaraj
K, Fonzino A, Barca E, Marino M, Rodolico C, Tricarico D, Camerino
C, et al: Risk of myopathy in patients in therapy with statins:
Identification of biological markers in a pilot study. Front
Pharmacol. 8:5002017. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Caparros-Martin JA, Lareu RR, Ramsay JP,
Peplies J, Reen FJ, Headlam HA, Ward NC, Croft KD, Newsholme P,
Hughes JD, et al: Statin therapy causes gut dysbiosis in mice
through a PXR-dependent mechanism. Microbiome. 5:952017. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Verhaegh BP, de Vries F, Masclee AA,
Keshavarzian A, de Boer A, Souverein PC, Pierik MJ and Jonkers DM:
High risk of drug-induced microscopic colitis with concomitant use
of NSAIDs and proton pump inhibitors. Aliment Pharmacol Ther.
43:1004–1013. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
O'Keefe SJ: Diet, microorganisms and their
metabolites, and colon cancer. Nat Rev Gastroenterol Hepatol.
13:691–706. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Peters U, Sinha R, Chatterjee N, Subar AF,
Ziegler RG, Kulldorff M, Bresalier R, Weissfeld JL, Flood A,
Schatzkin A, et al: Dietary fibre and colorectal adenoma in a
colorectal cancer early detection programme. Lancet. 361:1491–1495.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Mao QQ, Lin YW, Chen H, Qin J, Zheng XY,
Xu X and Xie LP: Dietary fiber intake is inversely associated with
risk of pancreatic cancer: A meta-analysis. Asia Pac J Clin Nutr.
26:89–96. 2017.PubMed/NCBI
|
|
22
|
Ohira H, Tsutsui W and Fujioka Y: Are
short chain fatty acids in hut microbiota defensive players for
inflammation and atherosclerosis. J Atheroscler Thromb. 24:660–672.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Hernandez-Rodas MC, Valenzuela R and
Videla LA: Relevant aspects of nutritional and dietary
interventions in non-alcoholic fatty liver disease. Int J Mol Sci.
16:25168–25198. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Burton-Freeman B, Liyanage D, Rahman S and
Edirisinghe I: Ratios of soluble and insoluble dietary fibers on
satiety and energy intake in overweight pre- and postmenopausal
women. Nutr Healthy Aging. 4:157–168. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Adam CL, Thomson LM, Williams PA and Ross
AW: Soluble fermentable dietary fibre (Pectin) decreases caloric
intake, adiposity and lipidaemia in high-fat diet-induced obese
rats. PLoS One. 10:e01403922015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Torcello-Gómez A, Fernández Fraguas C,
Ridout MJ, Woodward NC, Wilde PJ and Foster TJ: Effect of
substituent pattern and molecular weight of cellulose ethers on
interactions with different bile salts. Food Funct. 6:730–739.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Meneses ME, Martinez-Carrera D, Torres N,
Sánchez-Tapia M, Aguilar-López M, Morales P, Sobal M, Bernabé T,
Escudero H, Granados-Portillo O and Tovar AR: Hypocholesterolemic
properties and prebiotic effects of Mexican Ganoderma lucidum in
C57BL/6 mice. PLoS One. 11:e01596312016. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hung TV and Suzuki T: Dietary fermentable
fiber reduces intestinal barrier defects and inflammation in
colitic mice. J Nutr. 146:1970–1979. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Zhong Y, Marungruang N, Fak F and Nyman M:
Effects of two whole-grain barley varieties on caecal SCFA, gut
microbiota and plasma inflammatory markers in rats consuming low-
and high-fat diets. Br J Nutr. 113:1558–1570. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Luo Y, Zhang L, Li H, Smidt H, Wright AG,
Zhang K, Ding X, Zeng Q, Bai S, Wang J, et al: Different types of
dietary fibers trigger specific alterations in composition and
predicted functions of colonic bacterial communities in BALB/c
mice. Front Microbiol. 8:9662017. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Winglee K and Fodor AA: Intrinsic
association between diet and the gut microbiome: Current evidence.
Nutr Diet Suppl. 7:69–76. 2015.PubMed/NCBI
|
|
32
|
Jakobsdottir G, Xu J, Molin G, Ahrné S and
Nyman M: High-fat diet reduces the formation of butyrate, but
increases succinate, inflammation, liver fat and cholesterol in
rats, while dietary fibre counteracts these effects. PLoS One.
8:e804762013. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Vannucci L, Krizan J, Sima P, Stakheev D,
Caja F, Rajsiglova L, Horak V and Saieh M: Immunostimulatory
properties and anti-tumor activities of glucans (Review). Int J
Oncol. 43:357–364. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Chanput W, Reitsma M, Kleinjans L, Mes JJ,
Savelkoul HF and Wichers HJ: β-Glucans are involved in
immune-modulation of THP-1 macrophages. Mol Nutr Food Res.
56:822–833. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Ho HV, Sievenpiper JL, Zurbau A, Blanco
Mejia S, Jovanovski E, Au-Yeung F, Jenkins AL and Vuksan V: The
effect of oat β-glucan on LDL-cholesterol, non-HDL-cholesterol and
apoB for CVD risk reduction: A systematic review and meta-analysis
of randomized-controlled trials. Br J Nutr. 116:1369–1382. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Caz V, Gil-Ramirez A, Largo C, Tabernero
M, Santamaría M, Martín-Hernández R, Marín FR, Reglero G and
Soler-Rivas C: Modulation of cholesterol-related gene expression by
dietary fiber fractions from edible mushrooms. J Agric Food Chem.
63:7371–7380. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Barsanti L, Passarelli V, Evangelista V,
Frassanito AM and Gualtieri P: Chemistry, physico-chemistry and
applications linked to biological activities of β-glucans. Nat Prod
Rep. 28:457–466. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Anderson JW, Baird P, Davis RH Jr, Ferreri
S, Knudtson M, Koraym A, Waters V and Williams CL: Health benefits
of dietary fiber. Nutr Rev. 67:188–205. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Van Horn L, McCoin M, Kris-Etherton PM,
Burke F, Carson JA, Champagne CM, Karmally W and Sikand G: The
evidence for dietary prevention and treatment of cardiovascular
disease. J Am Diet Assoc. 108:287–331. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Vetvicka V and Vetvickova J: Effects of
yeast-derived beta-glucans on blood cholesterol and macrophage
functionality. J Immunotoxicol. 6:30–35. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
de Groot A, Luyken R and Pikaar NA:
Cholesterol-lowering effect of rolled oats. Lancet. 2:303–304.
1963. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Czop JK: The role of beta-glucan receptors
on blood and tissue leukocytes in phagocytosis and metabolic
activation. Pathol Immunopathol Res. 5:286–296. 1986. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Estrada A, Yun CH, Van Kessel A, Li B,
Hauta S and Laarveld B: Immunomodulatory activities of oat
beta-glucan in vitro and in vivo. Microbiol Immunol. 41:991–998.
1997. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Torrence PF: Biological response
modifiers: New approaches to disease intervention. Academic Press;
Orlando: pp. 3971985
|
|
45
|
Novak M and Vetvicka V: Beta-glucans,
history, and the present: Immunomodulatory aspects and mechanisms
of action. J Immunotoxicol. 5:47–57. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
46
|
Vetvicka V and Vetvickova J: β1,3-Glucan:
Silver bullet or hot air. Open Glycoscience. 3:1–6. 2010.
|
|
47
|
Vetvicka V and Vetvickova J: Comparison of
immunological effects of commercially available β-glucans: Part
III. Int J Clin Pathol. 2:2006. View Article : Google Scholar
|
|
48
|
Würsch P and Pi-Sunyer FX: The role of
viscous soluble fiber in the metabolic control of diabetes. A
review with special emphasis on cereals rich in beta-glucan.
Diabetes Care. 20:1774–1780. 1997. View Article : Google Scholar : PubMed/NCBI
|
|
49
|
Mosikanon K, Arthan D, Kettawan A,
Tungtrongchitr R and Prangthip P: Yeast β-glucan modulates
inflammation and waist circumference in overweight and obese
subjects. J Diet Suppl. 14:173–185. 2017. View Article : Google Scholar
|
|
50
|
Browder W, Williams D, Lucore P, Pretus H,
Jones E and McNamee R: Effect of enhanced macrophage function on
early wound healing. Surgery. 104:224–230. 1988.PubMed/NCBI
|
|
51
|
Vetvicka V and Vetvickova J: Anti-stress
action of an orally-given combination of resveratrol, β-glucan, and
vitamin C. Molecules. 19:13724–13734. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Vetvicka V and Vetvickova J: β-glucan
attenuates chronic fatigue syndrome in murine model. J Nat Sci.
1:e1122015.
|
|
53
|
Sima P, Vannucci L and Vetvicka V: Glucans
and cancer: Historical perspective. Cancer Translational Med.
1:209–214. 2015. View Article : Google Scholar
|
|
54
|
Barbieri A, Quagliariello V, Del Vecchio
V, Falco M, Luciano A, Amruthraj NJ, Nasti G, Ottaiano A, Berretta
M, Iaffaioli RV and Arra C: Anticancer and anti-inflammatory
properties of gano-derma lucidum extract effects on melanoma and
triple-negative breast cancer treatment. Nutrients. 9:pii:E2102017.
View Article : Google Scholar
|
|
55
|
Richter J, Kral V, Svozil V, Rajnohova DL,
Pohorska JI and Vetvicka V: Effects of transfer point glucan #300
supplementation on children exposed to passive
smoking-placebo-driven double-blind clinical trials. J Nutr Health.
1:1052014.
|
|
56
|
Vetvicka V, Richter J, Svozil V, Rajnohová
Dobiášová L and Král V: Placebo-driven clinical trials of
yeast-derived β-(1-3) glucan in children with chronic respiratory
problems. Ann Transl Med. 1:262013.
|
|
57
|
Brown GD and Gordon S: Immune recognition.
A new receptor for beta-glucans. Nature. 413:36–37. 2001.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Stambach NS and Taylor ME:
Characterization of carbohydrate recognition by langerin, a C-type
lectin of Langerhans cells. Glycobiology. 13:401–410. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Allendorf DJ, Ostroff GR, BAran JT, et al:
Oral WGP beta glucan treatment accelerates myeloid recovery and
survival after irradiation wxposure. BTR 2003: Unified Science
& Technology for Reducing Biological Threats & Counterin
Terrorism Albuquesrque: University of New Mexico; pp. 104–113.
2003
|
|
60
|
Ross GD and Vĕtvicka V: CR3 (CD11b. CD18):
A phagocyte and NK cell membrane receptor with multiple ligand
specificities and functions. Cell Exp Immunol. 92:181–184. 1993.
View Article : Google Scholar
|
|
61
|
Xia Y, Vetvicka V, Yan J, Hanikyrová M,
Mayadas T and Ross GD: The beta-glucan-binding lectin site of mouse
CR3 (CD11b/CD18) and its function in generating a primed state of
the receptor that mediates cytotoxic activation in response to
iC3b-opsonized target cells. J Immunol. 162:2281–2290.
1999.PubMed/NCBI
|
|
62
|
Legentil L, Paris F, Ballet C, Trouvelot
S, Daire X, Vetvicka V and Ferrières V: Molecular interactions of
β-(1→>3)-glucans with their receptors. Molecules. 20:9745–9766.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Williams DL, Ha T, Li C, Laffan J,
Kalbfleisch J and Browder W: Inhibition of LPS-induced NFkappaB
activation by a glucan ligand involves down-regulation of IKKbeta
kinase activity and altered phosphorylation and degradation of
IkappaBalpha. Shock. 13:446–452. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Rogers NC, Slack EC, Edwards AD, Nolte MA,
Schulz O, Schweighoffer E, Williams DL, Gordon S, Tybulewicz VL,
Brown GD and Reis e Sousa C: Syk-dependent cytokine induction by
Dectin-1 reveals a novel pattern recognition pathway for C type
lectins. Immunity. 22:507–517. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Elcombe SE, Naqvi S, Van Den Bosch MW,
MacKenzie KF, Cianfanelli F, Brown GD and Arthur JS: Dectin-1
regulates IL-10 production via a MSK1/2 and CREB dependent pathway
and promotes the induction of regulatory macrophage markers. PLoS
One. 8:e600862013. View Article : Google Scholar : PubMed/NCBI
|
|
66
|
Větvička V: (Beta)-glucans as natural
biological response modifiers. Nova Science Publishers Inc; New
York: 2013
|
|
67
|
Orth M and Bellosta S: Cholesterol: Its
regulation and role in central nervous system disorders.
Cholesterol. 2012:2925982012. View Article : Google Scholar : PubMed/NCBI
|
|
68
|
Li T and Chiang JY: Regulation of bile
acid and cholesterol metabolism by PPARs. PPAR Res.
2009:5017392009. View Article : Google Scholar : PubMed/NCBI
|
|
69
|
Worthmann A, John C, Rühlemann MC, Baguhl
M, Heinsen FA, Schaltenberg N, Heine M, Schlein C, Evangelakos I,
Mineo C, et al: Cold-induced conversion of cholesterol to bile
acids in mice shapes the gut microbiome and promotes adaptive
thermogenesis. Nat Med. 23:839–849. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
70
|
Li T and Chiang JY: Bile acids as
metabolic regulators. Curr Opin Gastroenterol. 31:159–165. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
71
|
Brown MS and Goldstein JL: How LDL
receptors influence cholesterol and atherosclerosis. Sci Am.
251:58–66. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
72
|
Russell DW: The enzymes, regulation, and
genetics of bile acid synthesis. Annu Rev Biochem. 72:137–174.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
73
|
Kuipers F, Bloks VW and Groen AK: Beyond
intestinal soap-bile acids in metabolic control. Nat Rev
Endocrinol. 10:488–498. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
74
|
Gibbons GF, Wiggins D, Brown AM and
Hebbachi AM: Synthesis and function of hepatic
very-low-density-lipoprotein. Biochem Soc Trans. 32:59–64. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
75
|
Davidson WS and Hilliard GM: The spatial
organization of apolipoprotein A-I on the edge of discoidal high
density lipo-protein particles. J Biol Chem. 278:27199–27207. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
76
|
de Oliveira Alvim R, Mourao-Junior CA,
Magalhaes GL, de Oliveira CM, Krieger JE, Mill JG and Pereira AC:
Non-HDL cholesterol is a good predictor of the risk of incfeased
arterial stiffness in postmenopausal women in an urban Brazilian
population. Clinics (Sao Paulo). 72:106–110. 2017. View Article : Google Scholar
|
|
77
|
Gordon DJ, Probstfield JL, Garrison RJ,
Neaton JD, Castelli WP, Knoke JD, Jacobs DR Jr, Bangdiwala S and
Tyroler HA: High-density lipoprotein cholesterol and cardiovascular
disease. Four prospective American studies. Circulation. 79:8–15.
1989. View Article : Google Scholar : PubMed/NCBI
|
|
78
|
Srinivasan SR, Myers L and Berenson GS:
Distribution and correlates of non-high-density lipoprotein
cholesterol in children: The Bogalusa heart study. Pediatrics.
110:e292002. View Article : Google Scholar : PubMed/NCBI
|
|
79
|
Hoenig MR: Implications of the obesity
epidemic for lipid-lowering therapy: Non-HDL cholesterol should
replace LDL cholesterol as the primary therapeutic target. Vasc
Health Risk Manag. 4:143–156. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
80
|
The National Cholesterol Education
Program. https://www.nhlbi.nih.gov/files/docs/guidelines/atglance.pdf.
|
|
81
|
Masson D, Koseki M, Ishibashi M, Larson
CJ, Miller SG, King BD and Tall AR: Increased HDL cholesterol and
apoA-I in humans and mice treated with a novel SR-BI inhibitor.
Arterioscler Thromb Vasc Biol. 29:2054–2060. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
82
|
White CR, Garber DW and Anantharamaiah GM:
Anti-inflammatory and cholesterol-reducing properties of
apolipoprotein mimetics: A review. J Lipid Res. 55:2007–2021. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
83
|
Parathasarathy S, Raghavamenon A,
Garelnabi MO and Santanam N: Oxidized low-density lipoprotein.
Methods Mol Biol. 610:403–417. 2010. View Article : Google Scholar
|
|
84
|
Besler C, Lüscher TF and Landmesser U:
Molecular mechanisms of vascular effects of high-density
lipoprotein: Alternations in cardiovascular disease. EMBO Mol Med.
4:251–268. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
85
|
Stary HC: The sequence of cell and matrix
changes in athero-sclerotic lesions of coronary arteries in the
first forty years of life. Eur Heart J. 11(Suppl E): S3–S19. 1990.
View Article : Google Scholar
|
|
86
|
Sullivan MP, Cerda JJ, Robbins FL, Burgin
CW and Beatty RJ: The gerbil, hamster, and guinea pig as rodent
models for hyper-lipidemia. Lab Anim Sci. 43:575–578.
1993.PubMed/NCBI
|
|
87
|
Sima A, Stancu C, Constantinescu E,
Ologeanu L and Simionescu M: The hyperlipemic hamster-a model for
testing the anti-atherogenic effect of amlodipine. J Cell Mol Med.
5:153–162. 2001. View Article : Google Scholar
|
|
88
|
Ohmura H, Fukushima Y, Mizuno A, Niwa K,
Kobayashi Y, Ebina T, Kimura K, Ishibashi S and Daida H: Research
Committee on Primary Hyperlipidemia of the Ministry of Health and
Welfare of Japan: Estimated prevalence of heterozygous familial
hypercholesterolemia in patients with acute coronary syndrome. Int
Heart J. 58:88–94. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
89
|
Rosenson RS, Brewer HB Jr, Ansell BJ,
Barter P, Chapman MJ, Heinecke JW, Kontush A, Tall AR and Webb NR:
Dysfunctional HDL and atherosclerotic cardiovascular disease. Nat
Rev Cardiol. 13:48–60. 2016. View Article : Google Scholar
|
|
90
|
Gistera A and Hansson GK: The immunology
of atherosclerosis. Nat Rev Nephrol. 13:368–380. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
91
|
Jonsson AL and Bäckhed F: Role of gut
microbiota in atherosclerosis. Nat Rev Cardiol. 14:79–87. 2017.
View Article : Google Scholar
|
|
92
|
Babicek K, Cehová I, Simon RR, Harwood M
and Cox DJ: Toxicological assesment of a particulate yeast
(1,3/1,6)-beta-D-glucan in rats. Food Chem Toxicol. 45:1719–1730.
2007. View Article : Google Scholar
|
|
93
|
Brown L, Rosner B, Willett WW and Sacks
FM: Cholesterol-lowering effects of dietary fiber: A meta-analysis.
Am J Clin Nutr. 69:30–42. 1999.PubMed/NCBI
|
|
94
|
Phillips GO and Cui SW: An introduction:
Evolution and finalisation of the regulatory definition of dietary
fibre. Food Hydrocolloids. 25:139–143. 2011. View Article : Google Scholar
|
|
95
|
Slavin JL: Dietary fiber and body weight.
Nutrition. 21:4114182005. View Article : Google Scholar : PubMed/NCBI
|
|
96
|
Cheung PCK: Mini-review on edible
mushrooms as source of dietary fiber: Preparation and health
benefits. Food Sci Human Wellness. 2:162–166. 2013. View Article : Google Scholar
|
|
97
|
Jenkins DJ, Kendall CW, Axelsen M,
Augustin LS and Vuksan V: Viscous and nonviscous fibres,
nonabsorbable and low glycaemic index carbohydrates, blood lipids
and coronary heart disease. Curr Opin Lipidol. 11:49–56. 2000.
View Article : Google Scholar : PubMed/NCBI
|
|
98
|
Gee JM, Blackburn NA and Johnson IT: The
influence of guar gum on intestinal cholesterol transport in the
rat. Br J Nutr. 50:215–224. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
99
|
Chau CF and Huang YL: Effects of the
insoluble fiber derived from Passiflora edulis seed on plasma and
hepatic lipids and fecal output. Mol Nutr Food Res. 49:786–790.
2005. View Article : Google Scholar : PubMed/NCBI
|
|
100
|
Cho IJ, Lee C and Ha TY: Hypolipidemic
effect of soluble fiber isolated from seeds of Cassia tora Linn. In
rats fed a high-cholesterol diet. J Agric Food Chem. 55:1592–1596.
2007. View Article : Google Scholar : PubMed/NCBI
|
|
101
|
Zacherl C, Eisner P and Engel KH: In vitro
model to correlate viscosity and bile acid-binding capacity of
digested water-soluble and insoluble dietary fibres. Food Chem.
126:423–428. 2011. View Article : Google Scholar
|
|
102
|
Aoki S, Iwai A, Kawata K, Muramatsu D,
Uchiyama H, Okabe M, Ikesue M, Maeda N and Uede T: Oral
administration of the Aureobasidium pullulans-derived β-glucan
effectively prevents the development of high fat diet-induced fatty
liver in mice. Sci Rep. 5:104572015. View Article : Google Scholar
|
|
103
|
Steinberg GR and Kemp BE: AMPK in health
and disease. Physiol Rev. 89:1025–1078. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
104
|
Kaczmarczyk MM, Miller MJ and Freund GG:
The health benefits of dietary fiber: Beyond the usual suspects of
type 2 diabetes mellitus, cardiovascular disease and colon cancer.
Metabolism. 61:1058–1066. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
105
|
Drew BG, Fidge NH, Gallon-Beaumier G, Kemp
BE and Kingwell BA: High-density lipoprotein and apolipoprotein AI
increase endothelial NO synthase activity by protein association
and multisite phosphorylation. Proc Natl Acad Sci USA.
101:6999–7004. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
106
|
Roy CC, Kien CL, Bouthillier L and Levy E:
Short-chain fatty acids: Ready for prime time. Nutr Clin Pract.
21:351–366. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
107
|
den Besten G, van Eunen K, Groen AK,
Venema K, Reijngoud DJ and Bakker BM: The role of short-chain fatty
acids in the interplay between diet, gut microbiota, and host
energy metabolism. J Lipid Res. 54:2325–2340. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
108
|
Chan GC, Chan WK and Sze DM: The effects
of beta-glucan on human immune and cancer cells. J Hematol Oncol.
2:252009. View Article : Google Scholar : PubMed/NCBI
|
|
109
|
Al-Lahham SH, Peppelenbosch MP, Roelofsen
H, Vonk RJ and Venema K: Biological effects of propionic acid in
humans; metabolism, potential applications and underlying
mechanisms. Biochim Biophys Acta. 1801:1175–1183. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
110
|
Wismar R, Brix S, Frøkiaer H and Laerke
HN: Dietary fibers as immunoregulatory compounds in health and
disease. Ann NY Acad Sci. 1190:70–85. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
111
|
Vangaveti V, Shashidhar V, Jarrod G, Baune
BT and Kennedy RL: Free fatty acid receptors: Emerging targets for
treatment of diabetes and its complications. Ther Adv Endocrinol
Metab. 1:165–175. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
112
|
Warnberg J, Gomez-Martinez S, Romeo J,
Diaz LE and Marcos A: Nutrition, inflammation, and cognitive
function. Ann N Y Acad Sci. 1153:164–175. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
113
|
Cosola C, De Angelis M, Rocchetti MT,
Montemurno E, Maranzano V, Dalfino G, Manno C, Zito A, Gesualdo M,
Ciccone MM, et al: Beta-glucans supplementation associates with
reduction in P-Cresyl sulfate levels and improved endo-thelial
vascular reactivity in healthy individuals. PLoS One.
12:e01696352017. View Article : Google Scholar
|
|
114
|
Mansbach CM II and Gorelick F: Development
and physiological regulation of intestinal lipid absorption. II.
Dietary lipid absorption, complex lipid synthesis, and the
intracellular packaging and secretion of chylomicrons. Am J Physiol
Gastrointest Liver Physiol. 293:G645–G650. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
115
|
Black DD: Development and physiological
regulation of intestinal lipid absorption. I. Development of
intestinal lipid absorption: Cellular events in chylomicron
assembly and secretion. Am J Physiol Gastrointest Liver Physiol.
293:G519–G524. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
116
|
Brown MS and Goldstein JL: The SREBP
pathway: Regulation of cholesterol metabolism by proteolysis of a
membrane-bound transcription factor. Cell. 89:331–340. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
117
|
Sato R: Sterol metabolism and SREBP
activation. Arch Biochem Biophys. 501:177–181. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
118
|
Drozdowski LA, Reimer RA, Temelli F, Bell
RC, Vasanthan T and Thomson AB: Beta-glucan extracts inhibit the in
vitro intestinal uptake of long-chain fatty acids and cholesterol
and down-regulate genes involved in lipogenesis and lipid transport
in rats. J Nutr Biochem. 21:695–701. 2010. View Article : Google Scholar
|
|
119
|
Chen J and Huang XF: The effects of diets
enriched in beta-glucans on blood lipoprotein concentrations. J
Clin Lipidol. 3:154–158. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
120
|
Rondanelli M, Opizzi A, Monteferrario F,
Klersy C, Cazzola R and Cestaro B: Beta-glucan- or rice
bran-enriched foods: A comparative crossover clinical trial on
lipidic pattern in mildly hypercholesterolemic men. Eur J Clin
Nutr. 65:864–871. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
121
|
Wolever TM, Tosh SM, Gibbs AL,
Brand-Miller J, Duncan AM, Hart V, Lamarche B, Thomson BA, Duss R
and Wood PJ: Physicochemical properties of oat β-glucan influence
its ability to reduce serum LDL cholesterol in humans: A randomized
clinical trial. Am J Clin Nutr. 92:723–732. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
122
|
Johnson IT and Gee JM: Effect of
gel-forming gums on the intestinal unstirred layer and sugar
transport in vitro. Gut. 22:398–403. 1981. View Article : Google Scholar : PubMed/NCBI
|
|
123
|
Kirby RW, Anderson JW, Sieling B, Rees ED,
Chen WJ, Miller RE and Kay RM: Oat-bran intake selectively lowers
serum low-density lipoprotein cholesterol concentrations of
hypercholesterolemic men. Am J Clin Nutr. 34:824–829.
1981.PubMed/NCBI
|
|
124
|
Fadel JG, Newman RK, Newman CW and Barnes
AE: Hypocholesterolemic effects of beta-glucans in different barley
diets fed to broiler chicks. Nutr Rep Int. 35:1049–1058. 1987.
|
|
125
|
Li J, Kaneko T, Qin LQ, Wang J, Wang Y and
Sato A: Long-term effects of high dietary fiber intake on glucose
tolerance and lipid metabolism in GK rats: Comparison among barley,
rice, and cornstarch. Metabolism. 52:1206–1210. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
126
|
Sima A, Bulla A and Simionescu N:
Experimental obstructive coronary atherosclerosis in the
hyperlipidemic hamster. J Submicrosc Cytol Pathol. 22:1–16.
1990.PubMed/NCBI
|
|
127
|
Lim MK, Ku SK, Choi JS and Kim JW: Effect
of polycan, a β-glucan originating from Aureobasidium, on a
high-fat diet-induced hyperlipemic hamster model. Exp Ther Med.
9:1369–1378. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
128
|
Delaney B, Nicolosi RJ, Wilson TA, Carlson
T, Frazer S, Zheng GH, Hess R, Ostergren K, Haworth J and Knutson
N: Beta-glucan fractions from barley and oats are similarly
anti-atherogenic in hypercholesterolemic Syrian golden hamsters. J
Nutr. 133:468–475. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
129
|
Wilson TA, Nicolosi RJ, Delaney B,
Chadwell K, Moolchandani V, Kotyla T, Ponduru S, Zheng GH, Hess R,
Knutson N, et al: Reduced and high molecular weight barley
beta-glucans decrease plasma total and non-HDL-cholesterol in
hypercholesterolemic Syrian golden hamsters. J Nutr. 134:2617–2622.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
130
|
Wu YS, Ho SY, Nan FH and Chen SN:
Ganoderma lucidum beta 1,3/1,6 glucan as an immunomodulator in
inflammation induced by a high-cholesterol diet. BMC Complement
Altern Med. 16:5002016. View Article : Google Scholar : PubMed/NCBI
|
|
131
|
Kusmiati and Dhewantara FX:
Cholesterol-lowering effect of beta glucan extracted from
Saccharomyces cerevisiae in rats. Sci Pharm. 84:153–165. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
132
|
Tong LT, Zhong K, Liu L, Zhou X, Qiu J and
Zhou S: Effects of dietary hull-less barley β-glucan on the
cholesterol metabolism of hypercholesterolemic hamsters. Food Chem.
169:344–349. 2015. View Article : Google Scholar
|
|
133
|
Vetvicka V and Vetvickova J: Physiological
effects of different types of beta-glucan. Biomed Pap Med Fac Univ
Palacky Olomouc Czech Repub. 151:225–231. 2007. View Article : Google Scholar
|
|
134
|
Malkki Y: Oat fiber. Food Science and
Technology: Handbook of Dietary Fiber. Cho S and Dreher ML: M.
Dekker; New York: pp. 497–512. 2001
|
|
135
|
Veniant MM, Withycombe S and Young SG:
Lipoprotein size and atherosclerosis susceptibility in
Apoe−/− and Ldlr−/− mice. Arterioscler Thromb
Vasc Biol. 21:1567–1570. 2001. View Article : Google Scholar
|
|
136
|
Pendse AA, Arbones-Mainar JM, Johnson LA,
Altenburg MK and Maeda N: Apolipoprotein E knock-out and knock-in
mice: Atherosclerosis, metabolic syndrome, and beyond. J Lipid Res.
50(Suppl): S178–S182. 2009. View Article : Google Scholar :
|
|
137
|
Anderson JW, Story L, Sieling B, Chen WJ,
Petro MS and Story J: Hypocholesterolemic effects of oat-bran or
bean intake for hypercholesterolemic men. Am J Clin Nutr.
40:1146–1155. 1984. View Article : Google Scholar : PubMed/NCBI
|
|
138
|
Anderson JW and Gustafson NJ:
Hypocholesterolemic effects of oat and bean products. Am J Clin
Nutr. 48(Suppl 3): S749–S753. 1988. View Article : Google Scholar
|
|
139
|
Braaten JT, Wood PJ, Scott FW, Wolynetz
MS, Lowe MK, Bradley-White P and Collins MW: Oat beta-glucan
reduces blood cholesterol concentration in hypercholesterolemic
subjects. Eur J Clin Nutr. 48:465–474. 1994.PubMed/NCBI
|
|
140
|
Newman RK, Lewis SE, Newman CW, Boik RJ
and Ramage RT: Hypocholesterolemic effect of barley foods on
healthy men. Nutr Rep Int. 39:749–760. 1989.
|
|
141
|
McIntosh GH, Whyte J, McArthur R and
Nestel PJ: Barley and wheat foods: Influence on plasma cholesterol
concentrations in hypercholesterolemic men. Am J Clin Nutr.
53:1205–1209. 1991. View Article : Google Scholar : PubMed/NCBI
|
|
142
|
Lupton JR, Robinson MC and Morin JL:
Cholesterol-lowering effect of barley bran flour and oil. J Am Diet
Assoc. 94:65–70. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
143
|
Lia A, Hallmans G, Sandberg AS, Sundberg
B, Aman P and Anderson H: Oat beta-glucan increases bile excretion
and a fiber rich barley fracton increases cholesterol excretion in
ileostomy subjects. Am J Clin Nutr. 62:1245–1251. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
144
|
Bell S, Goldman VM, Bistrian BR, Arnold
AH, Ostroff G and Forse RA: Effect of beta-glucan from oats and
yeast on serum lipids. Crit Rev Food Sci Nutr. 39:189–202. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
145
|
Li J, Kaneko T, Qin LQ, Wang J and Wang Y:
Effects of barley intake on glucose tolerance, lipid metabolism,
and bowel function in women. Nutrition. 19:926–929. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
146
|
Behall KM, Scholfield DJ and Hallfrisch J:
Diets containing barley significantly reduce lipids in mildly
hypercholesterolemic men and women. Am J Clin Nutr. 80:1185–1193.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
147
|
Behall KM, Scholfield DJ and Hallfrisch J:
Lipids significantly reduced by diets containing barley in
moderately hypercholes-terolemic men. J Am Coll Nutr. 23:55–62.
2004. View Article : Google Scholar : PubMed/NCBI
|
|
148
|
Keenan JM, Goulson M, Shamliyan T, Knutson
N, Kolberg L and Curry L: The effects of concentrated barley
beta-glucan on blood lipids in a population of
hypercholesterolaemic men and women. Br J Nutr. 97:1162–1168. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
149
|
Shimizu C, Kihara M, Aoe S, Araki S, Ito
K, Hayashi K, Watari J, Sakata Y and Ikegami S: Effect of high
beta-glucan barley on serum cholesterol concentrations and visceral
fat area in Japanese men-a randomized, double-blinded,
placebo-controlled trial. Plant Foods Hum Nutr. 63:21–25. 2008.
View Article : Google Scholar
|
|
150
|
Sundberg B: Cholesterol lowering effects
of a barley fibre flake product. Agro Food Industry Hi-Tech.
19:14–17. 2008.
|
|
151
|
Zhu X, Sun X, Wang M, Zhang C, Cao Y, Mo
G, Liang J and Zhu S: Quantitative assessment of the effects of
beta-glucan consumption on serum lipid profile and glucose level in
hypercholesterolemic subjects. Nutr Metab Cardiovasc Dis.
25:714–723. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
152
|
Keogh GF, Cooper GJ, Mulvey TB, McArdle
BH, Coles GD, Monro JA and Poppitt SD: Randomized controlled
crossover study of the effect of a highly beta-glucan-enriched
barley on cardiovascular disease risk factors in mildly
hypercholesterol-emic men. Am J Clin Nutr. 78:711–718. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
153
|
Biörklund M, van Rees A, Mensink RP and
Onning G: Changes in serum lipids and postprandial glucose and
insulin concentrations after consumption of beverages with
beta-glucans from oats or barley: A randomised dose-controlled
trial. Eur J Clin Nutr 5. 9:1272–1281. 2005. View Article : Google Scholar
|
|
154
|
Ibrügger S, Kristensen M, Poulsen MW,
Mikkelsen MS, Ejsing J, Jespersen BM, Dragsted LO, Engelsen SB and
Bügel S: Extracted oat and barley β-glucans do not affect
cholesterol metabolism in young healthy adults. J Nutr.
143:1579–1585. 2013. View Article : Google Scholar
|
|
155
|
Talati R, Baker WL, Pabilonia MS, White CM
and Coleman CI: The effects of barley-derived soluble fiber on
serum lipids. Ann Fam Med. 7:157–163. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
156
|
AbuMweis SS, Jew S and Ames NP: β-glucan
from barley and its lipid-lowering capacity: A meta-analysis of
randomized, controlled trials. Eur J Clin Nutr. 64:1472–1480. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
157
|
Anderson TJ, Grégoire J, Hegele RA,
Couture P, Mancini GB, McPherson R, Francis GA, Poirier P, Lau DC,
Grover S, et al: 2012 update of the Canadian Cardiovascular Society
guidelines for the diagnosis and treatment of dyslipidemia for the
prevention of cardiovascular disease in the adult. Can J Cardiol.
29:151–167. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
158
|
Newman RK and Newman CW: Barley for food
and health: Science, technology, and products. Genetics and
Nutrient Composition. John Wiley & Sons; Hoboken, NJ: pp.
56–94. 2008
|
|
159
|
Ho HV, Sievenpiper JL, Zurbau A, Blanco
Mejia S, Jovanovski E, Au-Yeung F, Jenkins AL and Vuksan V: A
systematic review and meta-analysis of randomized controlled trials
of the effect of barley β-glucan on LDL-C, non-HDL-C and apoB for
cardiovascular disease risk reductioni-iv. Eur J Clin Nutr.
70:13402016. View Article : Google Scholar
|
|
160
|
Mori K, Kobayashi C, Tomita T, Inatomi S
and Ikeda M: Antiatherosclerotic effect of the edible mushrooms
Pleurotus eryngii (Eringi), Grifola frondosa (Maitake), and
Hypsizygus marmoreus (Bunashimeji) in apolipoprotein E-deficient
mice. Nutr Res. 28:335–342. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
161
|
Sun JE, Ao ZH, Lu ZM, Xu HY, Zhang XM, Dou
WF and Xu ZH: Antihyperglycemic and antilipidperoxidative effects
of dry matter of culture broth of Inonotus obliquus in submerged
culture on normal and alloxan-diabetes mice. J Ethnopharmacol.
118:7–13. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
162
|
Bays HE, Evans JL, Maki KC, Evans M,
Maquet V, Cooper R and Anderson JW: Chitin-glucan fiber effects on
oxidized low-density lipoprotein: A randomized controlled trial.
Eur J Clin Nutr. 67:2–7. 2013. View Article : Google Scholar :
|
|
163
|
Serra-Majem L, Roman B and Estruch R:
Scientific evidence of interventions using the Mediterranean diet:
A systematic review. Nutr Rev. 64(Suppl): S27–S47. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
164
|
Wang Y, Harding SV, Eck P, Thandapilly SJ,
Gamel TH, Abdel-Aal el-SM, Crow GH, Tosh SM, Jones PJ and Ames NP:
High-molecular-weight β-glucan decreases serum cholesterol
differentially based on the CYP7A1 rs3808607 polymorphism in mildly
hypercholesterolemic adults. J Nutr. 146:720–727. 2016. View Article : Google Scholar : PubMed/NCBI
|