Introduction
Breast cancer has the highest mortality rate of the
female cancers in developed and developing countries, and its
incidence is steadily increasing (1,2).
Genetic and epigenetic alterations in tumor-suppressor genes (TSGs)
and oncogenes serve crucial roles in the development of human
neoplasia (3). It has been
recognized that the methylation of TSG promoters is frequently
involved in numerous types of tumorigenesis, including breast
cancer (4). The aberrant promoter
methylation of certain TSGs has been identified in breast cancer by
the present research group and other researchers, where the TSGs
include BRCA1, p16/CDKN2A, DKK3, cyclin
D2, PLCD1, PCDH10 and UCHL1 genes
(5,6).
Homeobox proteins are essential transcriptional
regulators that function in various developmental processes,
including cell growth, proliferation, differentiation, cell-cell
communication and the apoptotic pathway during pattern formation in
embryogenesis (7). The abnormal
expression of homeobox genes may cause an abnormal phenotype and
cell growth (8). Msh homeobox 1
(MSX1) is a homeobox gene located in chromosomal region
4p16.1 (9). MSX1 interacts with
β-catenin to inhibit the cell proliferation mediated by WNT
signaling pathways (10).
MSX1 mutation is involved in the congenital lack of teeth
(tooth agenesis or hypodontia), limb deficiency, craniofacial bone
morphogenesis and cleft lip, but few studies have examined its role
in tumorigenesis (11–16). Previous studies suggest that the
deregulated expression of MSX1 is involved in several human
malignant neoplasms, including lung, gastric, ovarian and cervical
cancers, as well as acute lymphoblastic leukemia. However, to the
best of our knowledge, the expression pattern and biological
functions of MSX1 in breast cancer have not yet been investigated
(17–22).
In the present study, the expression and promoter
methylation of MSX1 in multiple breast cancer cell lines and
primary tumors was examined, and the associations between
MSX1 methylation and the clinicopathological features of
breast cancer patients were analyzed. The biological functions and
underlying mechanisms of MSX1 in breast cancer were also
investigated.
Materials and methods
Cell lines, tumor samples and normal
tissues
Nine breast cancer cell lines provided by Professor
Tao (Cancer Epigenetics Laboratory, Department of Clinical
Oncology, State Key Laboratory of Oncology in South China, Sir YK
Pao Center for Cancer and Li Ka Shing Institute of Health Sciences,
The Chinese University of Hong Kong and CUHK Shenzhen Research
Institute, Hong Kong, China) were used in this study: BT549, MCF-7,
MDA-MB-468, MDA-MB-231, SK-BR-3, T47D, YYC-B1, YCC-B3 and ZR-75-1.
All the cell lines were cultured in Dulbecco's modified Eagle's
medium (DMEM; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA,
USA) with 10% fetal bovine serum (Invitrogen; Thermo Fisher
Scientific, Inc.) and 100 U/ml penicillin/streptomycin at 37°C in
moist air containing 5% CO2.
Normal human adult tissue RNA samples were purchased
commercially (Stratagene; Agilent Technologies, Inc., Santa Clara,
CA, USA; EMD Millipore, Billerica, MA, USA; BioChain Institute,
Inc., Newark, CA, USA). Primary breast cancer tumor tissues,
matched adjacent non-malignant tissues and normal breast tissues
were obtained from patients who had undergone primary surgery at
the Department of Endocrinological and Breast Surgery, The First
Affiliated Hospital of Chongqing Medical University (Chongqing,
China), between January 2010 and March 2014. All samples were
confirmed by pathologist physicians. The clinical and pathological
data, including sex, age, tumor grade, tumor size, treatment and
follow-up data, were available for the majority of the breast
cancer samples. All patients signed informed written consent forms
for participation in the study at initial clinical investigation.
The present study was approved by the Ethics Committee of the First
Affiliated Hospital of Chongqing Medical University (approval no.
2010/2012-23). A multifunctional user-friendly online tool, Gene
Expression Based Outcome for Breast Cancer Online (GOBO; http://co.bmc.lu.se/gobo), was used to analyze the
expression level of MSX1 associated with molecular subtypes
of breast cancer, ER status and histological grade as previously
described (23).
Semiquantitative reverse
transcription-polymerase chain reaction (RT-PCR) analysis
Total RNA was isolated from tissues and cells using
TRI Reagent (Thermo Fisher Scientific, Inc.) according to the
manufacturer's protocol. Semi-quantitative RT-PCR was performed as
described previously, using GAPDH as the control (24). The primer sequences used for PCR
amplification are listed in Table
I. RT-PCR was performed with 32 cycles for MSX1 and 23 cycles
for GAPDH using Go-Taq Flexi DNA Polymerase (Promega Corporation,
Madison, WI, USA).
 | Table IList of primers used in this
study. |
Table I
List of primers used in this
study.
| PCR | | Sequence
(5′-3′) | Product size
(bp) | PCR cycles | Annealing
temperature |
|---|
| MSP | MSX1m3 |
GCGCCTCATCACATCAGCGC | 116 | 41 | 60°C |
| MSX1m4 |
GCGATTTCTGATGCTGGCGC | | | |
| MSX1u3 |
CAAGGCTAGTCATCATCAACCA | 121 | 41 | 58°C |
| MSX1u4 |
CGCCTAGGGCTCAGTCCACCATGT | | | |
| RT-PCR | MSX1-F |
CATTCGAATACCGGGGCCGACGA | 176 | 32 | 55°C |
| MSX1-R |
CGCCTAGGGCTCAGTCCACCATGT | | | |
| GAPDH-F |
CCTCAGTTGCCTAAACCA | 202 | 23 | 55°C |
| GAPDH-R |
CACTACCCTAAAGGTAACTA | | | |
| RT-qPCR | MSX1-F |
CTGCTCGTCTCGTTAATGTGG | 156 | 40 | 60°C |
| MSX1-R |
TGCGCAAACTTACCCGTCT | | | |
| β-actin-F |
GGACACCGTAGCGTGCTTTGA | 311 | 40 | 60°C |
| β-actin-R |
CTTCGCTAAACGCCACCTGCTA | | | |
5-Aza-2′-deoxycytidine (Aza) and
trichostatin A (TSA) treatment
The MDA-MB-468, MCF-7 and ZR-75-1 cell lines were
used for pharmacological demethylation. Briefly 1×106
cells were treated with 10 mmol/l Aza for 72 h and then with 100
nmol/l TSA (both from Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany) for 24 h at 37°C. The cells were then harvested for DNA
and RNA extraction, and further analysis.
DNA bisulfite treatment and
methylation-specific PCR (MSP)
Genomic DNA was extracted from the tumors, normal
tissues and cell pellets using the QIAamp DNA mini kit (Qiagen
GmbH, Hilden, Germany). DNA bisulfite treatment and MSP were
performed as previously described (25). The bisulfite-treated DNA was
amplified using MSP with a methylated-MSX1-specific or
unmethylated-MSX1-specific primer set. The primers used for
MSP and bisulfite sequencing are listed in Table I. The methylated and unmethylated
MSP primer sets target the same CpG sites and have been tested
previously to confirm that they do not amplify any genomic DNA
without bisulfite treatment, and are therefore specific. MSP was
performed for 40 cycles to amplify the unmethylated and methylated
gene using AmpliTaq Gold DNA Polymerase (Applied Biosystems; Thermo
Fisher Scientific, Inc.). The PCR products were analyzed on 2%
agarose gel.
RT-quantitative PCR (RT-qPCR)
Total RNA was purified from a panel of fresh, paired
primary breast tissues (20 tumors and the corresponding adjacent
tissues) using TRI Reagent as previously described (26). RT-qPCR with Maxima SYBR-Green/ROX
qPCR Master mix (MBI Fermentas; Thermo Fisher Scientific, Inc.) was
used to detect gene expression (Table
I) according to the manufacturer's protocol, using a HT7500
System (Applied Biosystems; Thermo Fisher Scientific, Inc.).
Melting-curve analysis and agarose gel electrophoresis of the PCR
products were performed to verify the specificity of the PCR and
the identity of the PCR products. Each experiment was performed in
triplicate and the relative expression levels of MSX1 in the breast
tissues were normalized to those of β-actin. Data were normalized
using the 2−ΔΔCq method (27).
Cell proliferation assay
MDA-MB-231 cells infected with LV-MSX1 or LV-empty
were trypsinized and resuspended. The cells were then cultured in
triplicate in 96-well plates at a density of 1×104
cells/well in a volume of 100 μl DMEM and allowed to grow
overnight. Cell viability was quantified with a Cell Counting kit-8
assay (CCK-8; Dojindo Molecular Technologies, Inc., Kumamoto,
Japan), in which only living cells are stained. After 24, 48 and 72
h, the medium was removed, and α-minimal essential medium (100
μl) containing 10 μl CCK-8 reagent was added to each
well of precultured cells. Following incubation of the cells for 2
h at 37°C, the optical density was determined at a wavelength of
450 nm with an automatic microplate reader. The results are the
means of three independent experiments conducted over several
days.
Colony formation assay
MDA-MB-231 and MDA-MB-468 cells infected by LV-MSX1
or LV-empty were plated at a density of 300 cells/well in 6-well
plates and cultured for 14 days in normal culture medium. The cells
were then washed twice with PBS, fixed with 10% buffered formalin,
dried and stained with 2% crystal violet. The surviving colonies
(≥50 cells/colony) were manually counted in four different fields
of vision and the mean values calculated. Each treatment was tested
in triplicate.
Wound healing assay
Cultured MDA-MB-231 cells infected with LV-MSX1 or
LV-empty were evenly seeded in 6-well plates and allowed to reach
100% confluence in DMEM. A straight wound was induced by dragging a
20-μl pipette tip through the confluent cell monolayer. The
cells were incubated and allowed to migrate in the medium. At 0,
12, 24 and 36 h after wounding, the plates were washed twice with
PBS to remove the dead cells, and images were captured in four
random fields at a magnification of ×100 (Leica DMI4000B; Leica
Microsystems, Ltd., Milton Keynes, UK). The rate of cell migration
was quantified according to the percentage of repaired wound area,
using Image Pro-Plus software (version 6.0; Media Cybernetics,
Inc., Rockville, MD, USA). Each experiment was performed in
triplicate.
Flow cytometric analysis of the cell
cycle
For cell-cycle analysis, MDA-MB-231 cells and
MDA-MB-468 cells infected with LV-MSX1 or LV-empty were seeded in
6-well plates (1×106 cells/well) and incubated overnight
at 37°C in 5% Co2. The cells were digested by trypsin at
48 h after infection and then centrifuged at 1,000 × g for 5 min at
room temperature. The cells were washed twice with PBS, fixed in
70% ethanol at 4°C for 2 h, and stained with propidium iodide
(Sigma-Aldrich; Merck KGaA) for 30 min at 37°C. The samples were
evaluated with the BD FACSCanto and the results were analyzed using
BD FACSDIVA software (version 4.1; BD Biosciences, San Jose, CA,
USA). All experiments were performed in triplicate, and one
representative figure is shown for each cell type.
Transwell cell migration and invasion
assays
Cell migration and invasion assays were carried out
using Transwell chambers (8 μm; Corning Incorporated,
Corning, NY, USA). The cells were plated at a density of
2.5×105 with RPMI-1640 in the upper well of each
Transwell chamber. For invasion assay, 100 μl of Matrigel
were added in the upper well first. The lower compartment was
filled with DMEM supplemented with 10% fetal bovine serum.
Following incubation for 24 h at 37°C with 5% CO2 in a
humidified atmosphere, the cells that had not migrated or invaded
were removed from the upper surface of the filter with gentle
swabbing. The cells on the lower surface of the filter were fixed
in 4% paraformaldehyde, stained with 0.1% crystal violet staining
solution for 15 min at room temperature, and counted by light
microscopy at ×100 magnification in 10 images taken in five random
fields. The averages were then calculated. The experiment was
performed in triplicate.
Western blot analysis
The infected cells were harvested and lysed in M-PER
Mammalian Protein Extraction Reagent (Pierce, Thermo Fisher
Scientific, Inc.) containing a protease inhibitor cocktail
(Sigma-Aldrich; Merck KGaA). The protein samples were incubated on
ice for 10 min, and then centrifuged at 10,000 × g for 5 min at 4°C
to remove the cell debris. The protein samples were quantified by
BCA Protein Assay kit (Thermo Fisher Scientific, Inc.), 20
μg protein were separated using 10% SDS-PAGE gel and then
electrophoretically transferred onto a polyvinylidene difluoride
membrane (Bio-Rad Laboratories, Inc., Hercules, CA, USA). The
membrane was blocked with 5% skimmed milk in Tris-buffered saline
with Tween-20 for 1 h at room temperature, and then incubated
overnight at 4°C with the following primary antibodies at the
manufacturers' recommended dilutions: antibodies directed against
β-catenin (#19807; Cell Signaling Technology, Inc., Danvers, MA,
USA) active β-catenin (#4270; Cell Signaling Technology, Inc.),
c-Myc (#1472-1; Epitomics; Abcam, Cambridge, UK), cleaved poly
(ADP-ribose) polymerase (cleaved PARP; #9541), cleaved caspase-3
(#9661) (both from Cell Signaling Technology, Inc.) and cyclin D1
(CCND1; #1677-1; Epitomics, Abcam). The membranes were washed and
then incubated with anti-mouse (#7076; Cell Signaling Technology,
Inc.) or anti-rabbit (#7074; Cell Signaling Technology, Inc.) IgG,
HRP-linked antibodies at a dilution of 1:3,000 at room temperature
for 1 h. The protein bands were visualized using a Fusion FX5
system (Vilber Lourmat, Eberhardzell, Germany). The results were
analyzed using ImageJ software (version 6.0; National Institutes of
Health, Bethesda, MD, USA). GAPDH (sc-47724; Santa Cruz
Biotechnology, Santa Cruz, CA, USA) was used as the endogenous
protein for normalization.
Statistical analysis
Statistical analyses were performed using the
Student's t-test, χ2 test and Fisher's exact test to
determine the significance of differences between groups. For all
tests, p<0.05 was considered to indicate a statistically
significant difference. Results are presented as the mean ±
standard deviation.
Results
Expression of MSX1 is reduced in breast
cancer tissues and cells
The expression of MSX1 was assessed in a
panel of normal human adult tissues, including breast tissues, and
breast cancer cell lines, using semi-quantitative RT-PCR (Fig. 1). MSX1 was clearly
expressed in all the normal human tissues, including normal breast
tissue, at varying levels (Fig.
1A), but was frequently silenced or downregulated in breast
cancer cell lines, with the exception of BT549 (Fig. 1C).
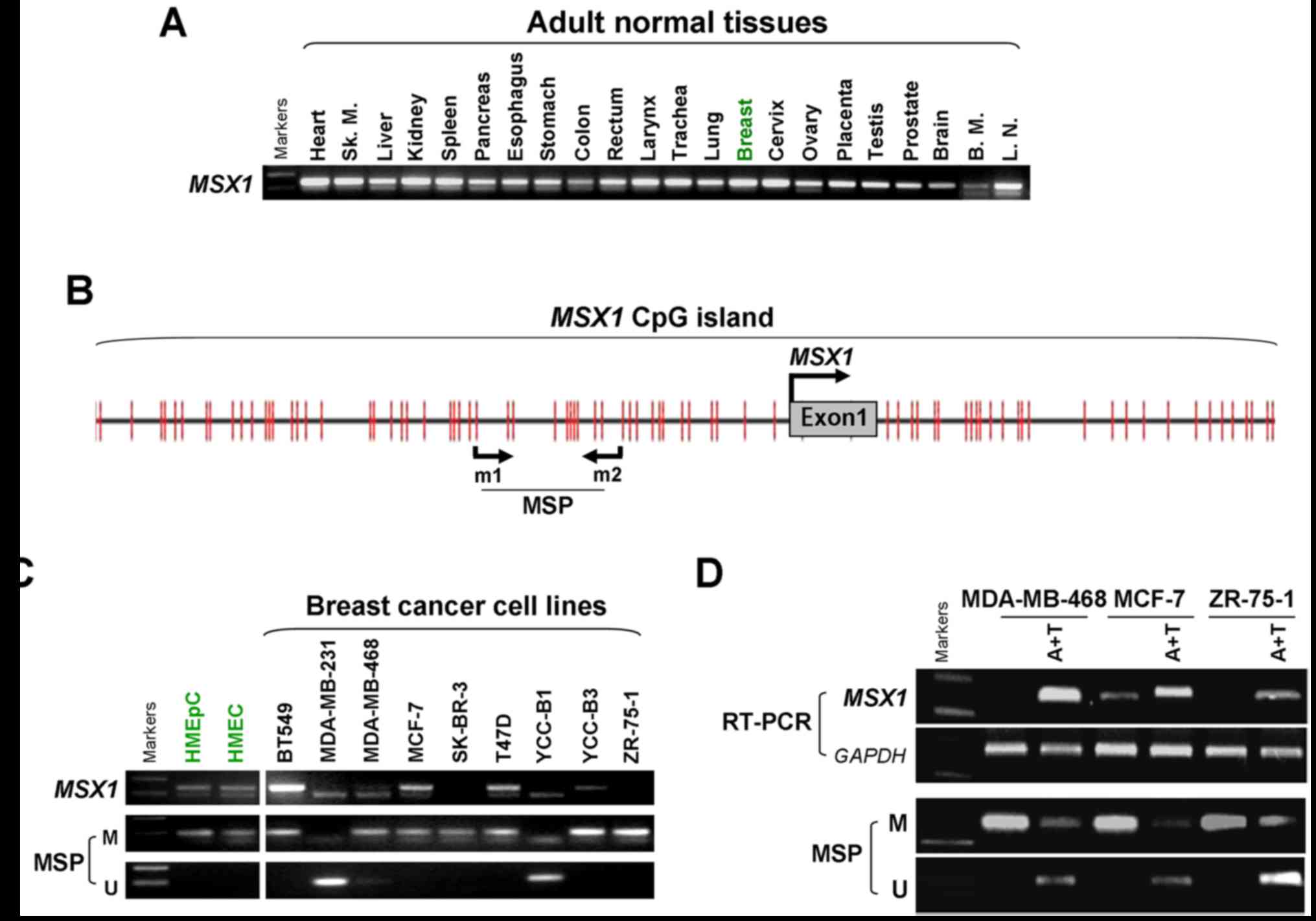 | Figure 1MSX1 is downregulated by
promoter methylation in breast cancer cell lines. (A) Analysis of
MSX1 expression in normal human adult tissues as measured by
semi-quantitative RT-PCR, using GAPDH as the control. Sk.m, Sk.
Muscle; B.M., Bone marrow; L.N., Lymph node. (B) Schematic
structure of the MSX1 promoter CpG island. The CpG sites are
indicated by the short vertical lines. The MSP sites analyzed are
shown. The transcription start site is indicated by a curved arrow.
(C) MSX1 is frequently silenced in breast cancer cell lines
by promoter methylation, demonstrated with RT-PCR and MSP. (D)
Pharmacological demethylation using A+T restored MSX1
expression in MSX1-methylated/silenced breast cancer cell
lines. MSX1, msh homeobox 1; RT-PCR, reverse
transcription-quantitative polymerase chain reaction; MSP,
methylation-specific PCR; M, methylated; U, unmethylated; A+T,
5-aza-2′-deoxycytidine plus trichostatin A. |
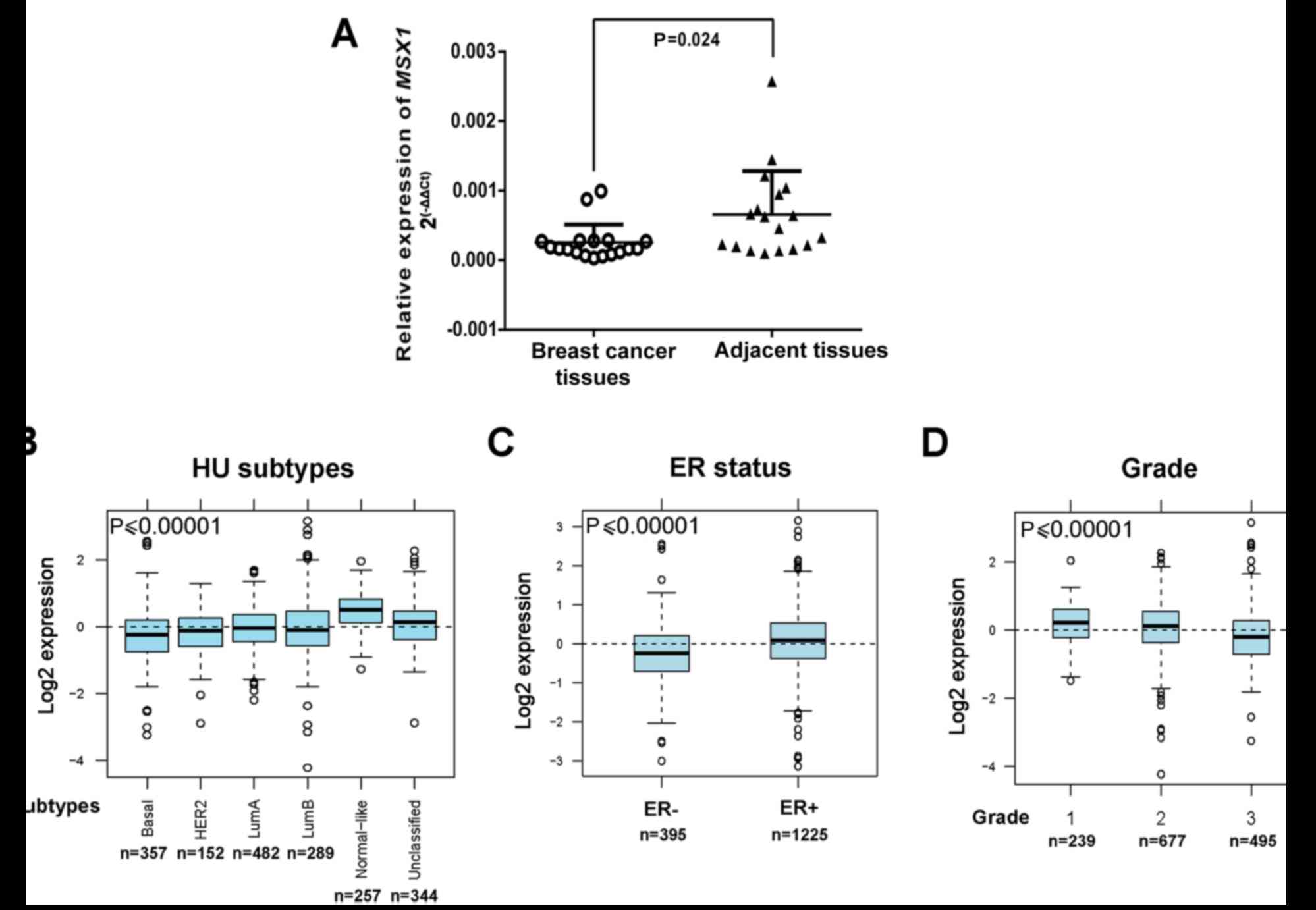
MSX1 expression was then examined at the mRNA
level in primary breast tumors. qPCR demonstrated that MSX1
mRNA was downregulated in the breast cancer tissues compared with
the normal breast tissues (p<0.05; Fig. 2A). The GOBO online tool was also
used to assess MSX1 expression in 1,881 breast cancers.
Gene-Set Analysis tumor demonstrated that the expression of
MSX1 differs among different subtypes of breast cancer
(28). The most aggressive
subtype, the basal-like subtype, exhibited the lowest MSX1
expression, whereas the least aggressive subtype, the normal-like
subtype, exhibited the highest expression of MSX1
(p<0.00001; Fig. 2B). The
expression of MSX1 was also reduced with a negative estrogen
receptor (ER) status and a higher histological grade (p<0.00001;
Fig. 2C and D). These results
indicate that MSX1 expression is frequently downregulated or
silenced in breast cancer, and is associated with the malignant
progression of breast cancer.
CpG methylation of the MSX1 promoter
contributes to its silencing or downregulation in breast
cancer
The involvement of promoter methylation in
MSX1 silencing in breast cancer was evaluated. Bioinformatic
analysis revealed a typical CpG island spanning the proximal
promoter and the exon 1 region of the MSX1 gene (Fig. 1B). To determine whether promoter
methylation leads to MSX1 silencing, MSX1 methylation
in breast tumor cell lines was analyzed using MSP. The results
demonstrated that MSX1 was methylated in seven breast cancer
cell lines (MDA-MB-231, MDA-MB-468, MCF-7, T47D, ZR75-1, SK-BR-3
and YCC-B3), consistent with its silencing, but not in another two
cell lines (BT549 and YCC-B1) in which MSX1 was unmethylated
(Fig. 1C).
Pharmacological demethylation was conducted to test
whether promoter methylation directly mediates the reduction of
MSX1 levels in breast cancer cells. Three cell lines
(MDA-MB-468, MCF-7 and ZR-75-1) lacking MSX1 expression were
treated with Aza and the histone deacetylase inhibitor TSA.
Following treatment, the expression of MSX1 in these cell
lines was significantly increased compared with that prior to
treatment, accompanied by decreased methylated alleles of
MSX1 (Fig. 1D). These
results indicate that promoter methylation is responsible for
MSX1 silencing in breast cancer cells.
Frequent MSX1 methylation and its
association with clinicopathological features in primary breast
tumors
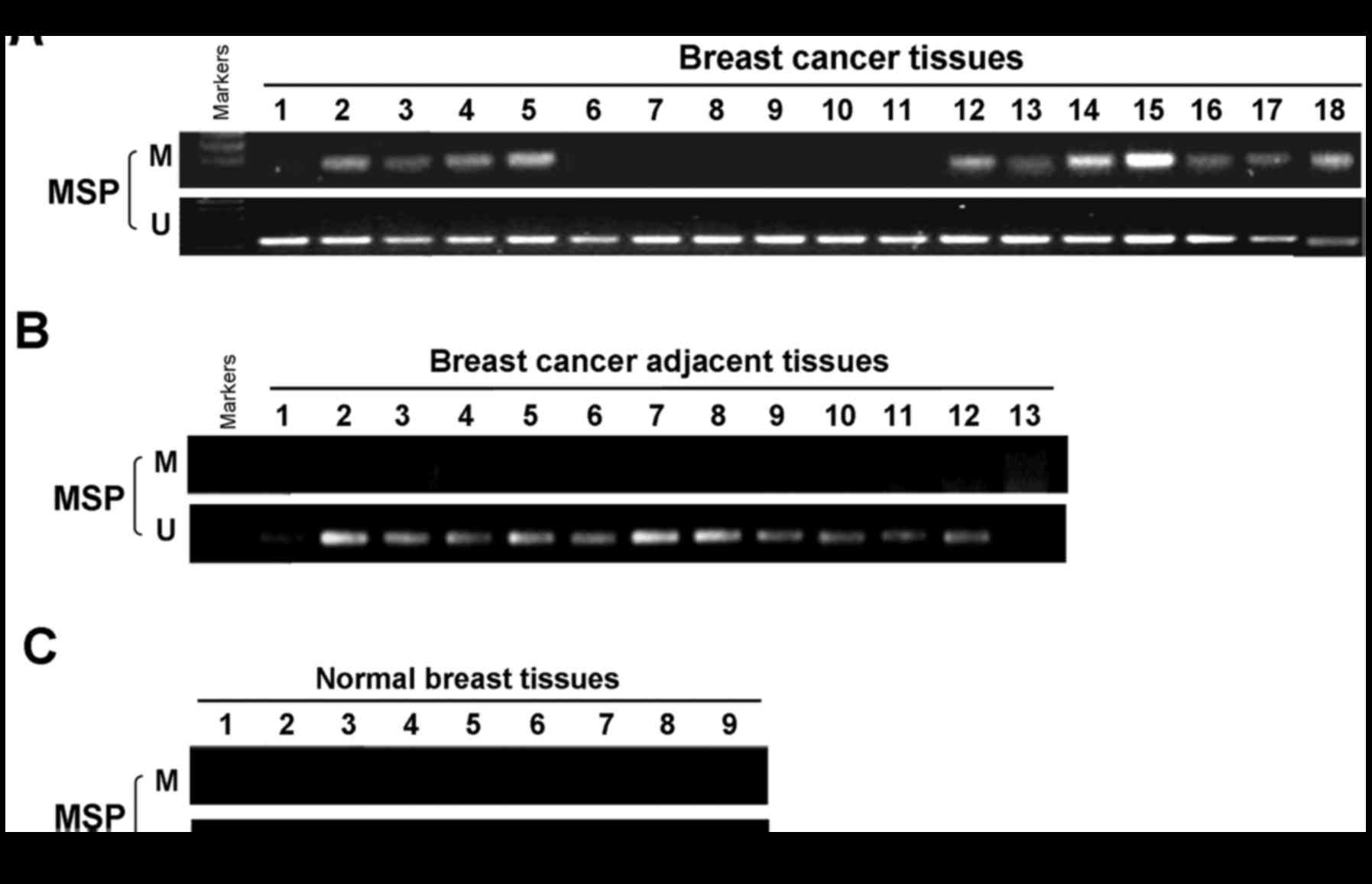
To ascertain whether MSX1 methylation occurs
in primary breast tumors, MSX1 methylation by MSP was examined in
99 primary breast tumor tissues, 8 surgical margin tissues and 13
normal breast tissues. Aberrant MSX1 methylation was
detected in 47 (47.5%) of the primary breast cancer tissues, but
not in the surgical margin tissues or normal breast tissues
(Fig. 3 and Table II). These results confirm the
tumor-specific methylation of the MSX1 promoter. The
association of MSX1 methylation with the clinicopathological
features of breast cancer patients was also analyzed, where the
clinicopathological features included age, tumor size, tumor grade,
lymph-node metastasis, ER status, progesterone receptor status,
human epidermal growth factor receptor 2 status and Ki67 status.
However, no significant difference of MSX1 methylation status
according to clinicopathological features of the patients was
detected (Table III).
| Table IIPromoter methylation status of
MSX1 in primary breast tumors. |
Table II
Promoter methylation status of
MSX1 in primary breast tumors.
| Tissue | Samples (n) | MSX1 promoter
| Methylation
frequency (%) |
|---|
| Methylated | Unmethylated |
|---|
| Breast cancer | 99 | 47 | 52 | 47/99 (47.5) |
| Breast cancer
surgical-margin | 8 | 0 | 8 | 0/8 (0) |
| Normal breast | 13 | 0 | 13 | 0/13 (0) |
| Table IIIClinicopathologic features of 99
breast cancer patients according to MSX1 methylation status. |
Table III
Clinicopathologic features of 99
breast cancer patients according to MSX1 methylation status.
| Clinicopathological
features | No. of
patients | MSX1 promoter
| P-value |
|---|
| Methylated | Unmethylated |
|---|
| Age (years) |
| ≥60 | 26 | 15 | 11 | 0.36 |
| <60 | 68 | 29 | 39 | |
| Unknown | 5 | 3 | 2 | |
| Tumor stage |
| I | 28 | 17 | 11 | 0.18 |
| II | 34 | 16 | 18 | |
| III | 28 | 9 | 19 | |
| Unknown | 9 | 5 | 4 | |
| Tumor size
(cm) |
| <2 | 23 | 13 | 10 | 0.53 |
| ≥2, ≤5 | 64 | 29 | 35 | |
| >5 | 7 | 2 | 5 | |
| Unknown | 5 | 3 | 2 | |
| Lymph node
metastasis |
| Positive | 47 | 20 | 27 | 0.35 |
| Negative | 45 | 22 | 23 | |
| Unknown | 7 | 5 | 2 | |
| Estrogen receptor
status |
| Positive | 53 | 24 | 29 | 0.85 |
| Negative | 33 | 17 | 16 | |
| Unknown | 13 | 6 | 7 | |
| Progesterone
receptor status |
| Positive | 42 | 18 | 24 | 0.73 |
| Negative | 43 | 22 | 21 | |
| Unknown | 14 | 7 | 7 | |
| HER2 status |
| Positive | 67 | 34 | 33 | 0.56 |
| Negative | 19 | 7 | 12 | |
| Unknown | 13 | 6 | 7 | |
| Ki67 status |
| <14% | 38 | 19 | 19 | 0.91 |
| >14% | 29 | 13 | 16 | |
| Unknown | 32 | 15 | 17 | |
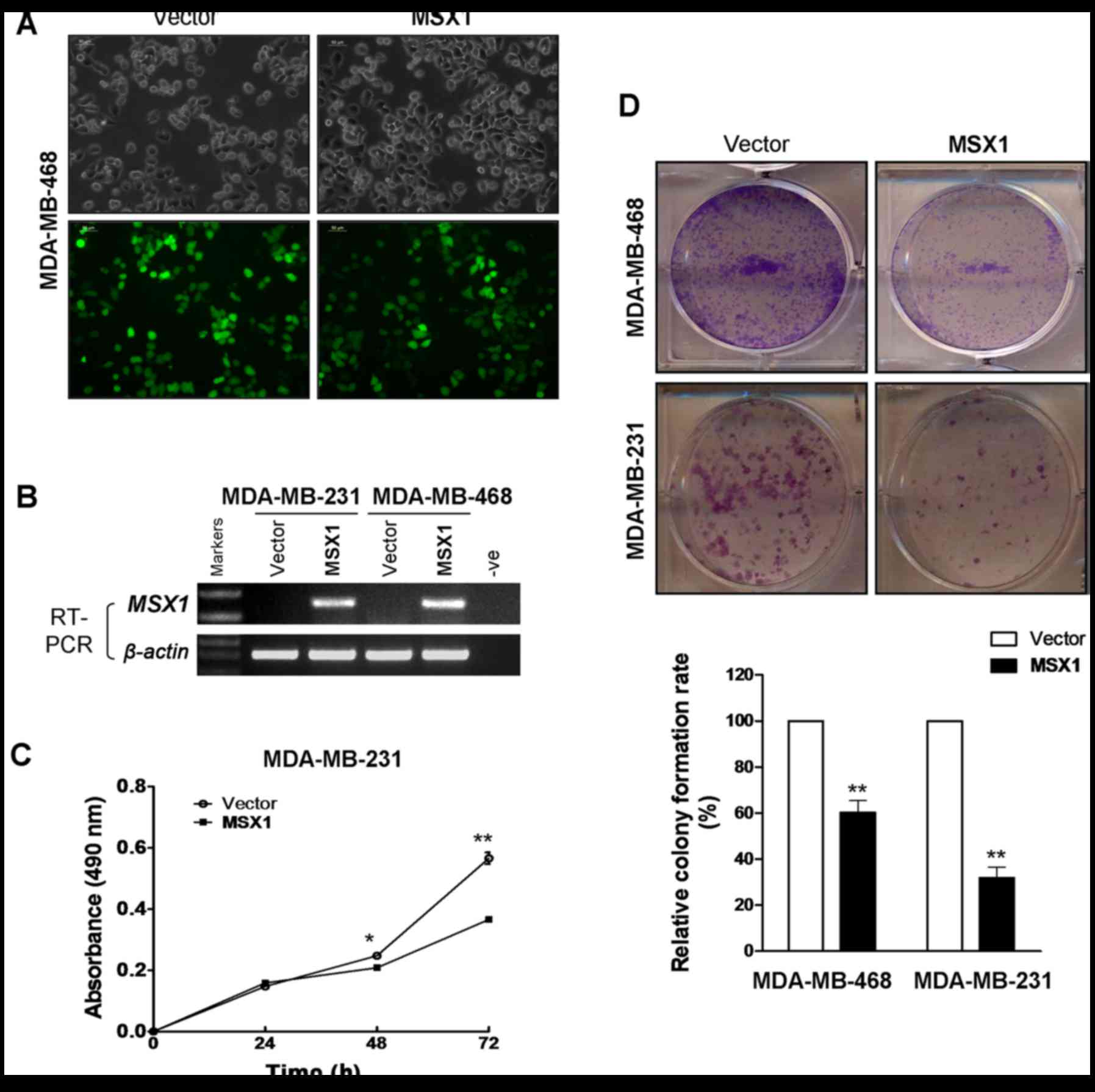
Ectopic expression of MSX1 suppresses
breast cancer cell growth
MDA-MB-231 and MDA-MB-468 cells, which are
MSX1-silenced, were infected with LV-MSX1 and LV-empty plasmids.
MDA-MB-231 and MDA-MB-468 cells stably overexpressing MSX1 were
successfully generated, which was confirmed by microscopy and
RT-PCR (Fig. 4A and B). CCK-8
cell proliferation and colony formation assays were performed to
clarify the function of MSX1 in the proliferation of breast cancer
cells. Cell viability was significantly reduced at 48 and 72 h
after the infection of the MDA-MB-231 cells with MSX1 (p<0.05
and p<0.01, respectively; Fig.
4C). The colony formation assay demonstrated that MSX1-infected
MDA-MB-231 and MDA-MB-468 cell colonies were reduced by 40 and 65%
compared with those of the respective control cells (p<0.01;
Fig. 4D). These results suggest
that MSX1 inhibits the proliferation of breast cancer cells.
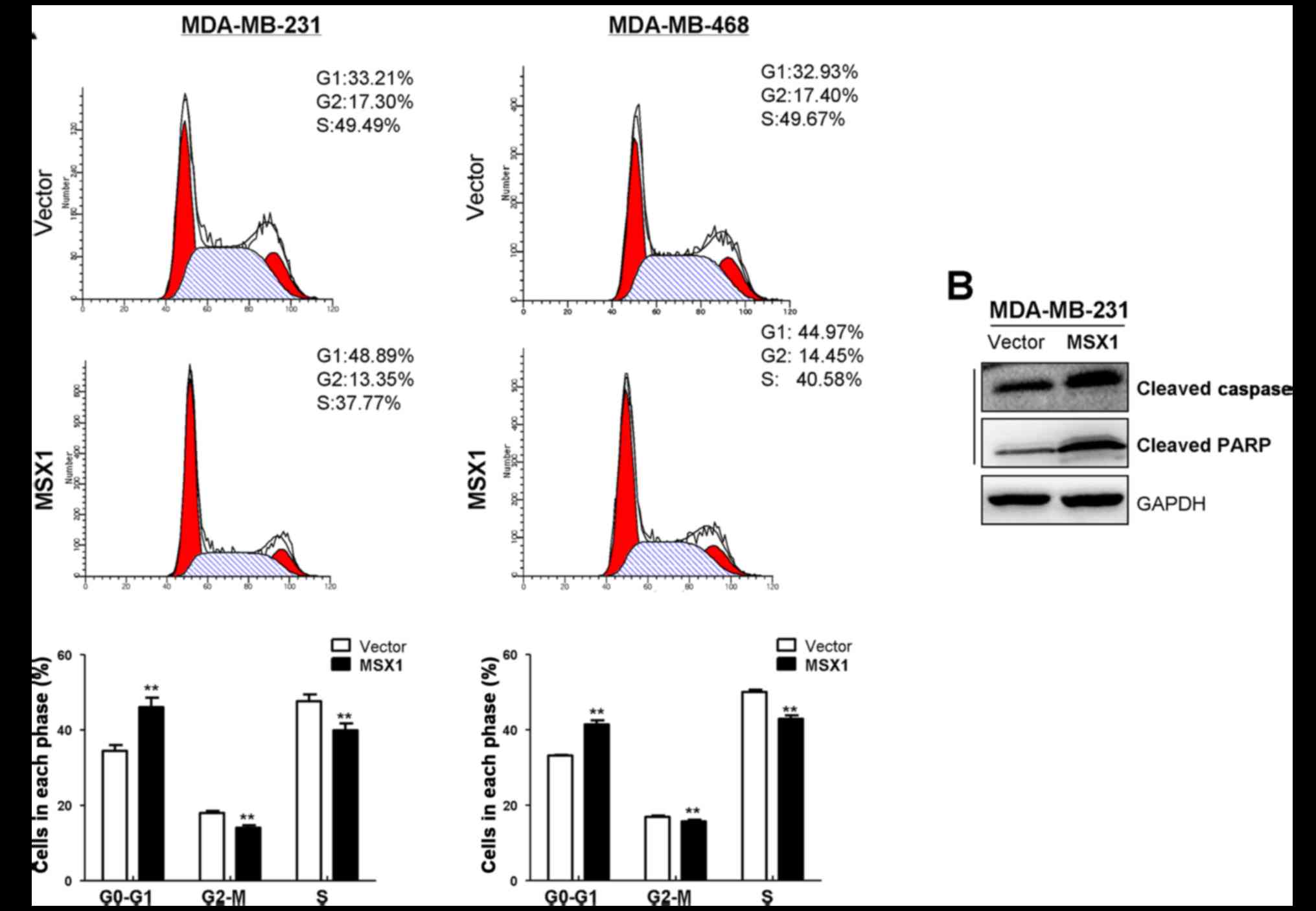
MSX1 induces the G1/S cell-cycle arrest
and apoptosis of breast cancer cells
To investigate the mechanism underlying the
growth-inhibitory effect of MSX1 on breast cancer cells, the cell
cycle status of breast cancer cells was examined using flow
cytometry. The number of cells arrested in the G1/S phase increased
significantly in the MSX1-expressing MDA-MB-231 and MDA-MB-468
cells compared with the respective control cells, which was
accompanied by a significant reduction in S-phase cells (p<0.01;
Fig. 5A). Western blot analysis
revealed upregulated expression levels of cleaved caspase-3 and
cleaved PARP in the MSX1-expressing MDA-MB-231 cells compared with
the control cells infected with empty vector. These results
indicate that MSX1 causes G1/S cell cycle arrest and apoptosis in
breast cancer cells.
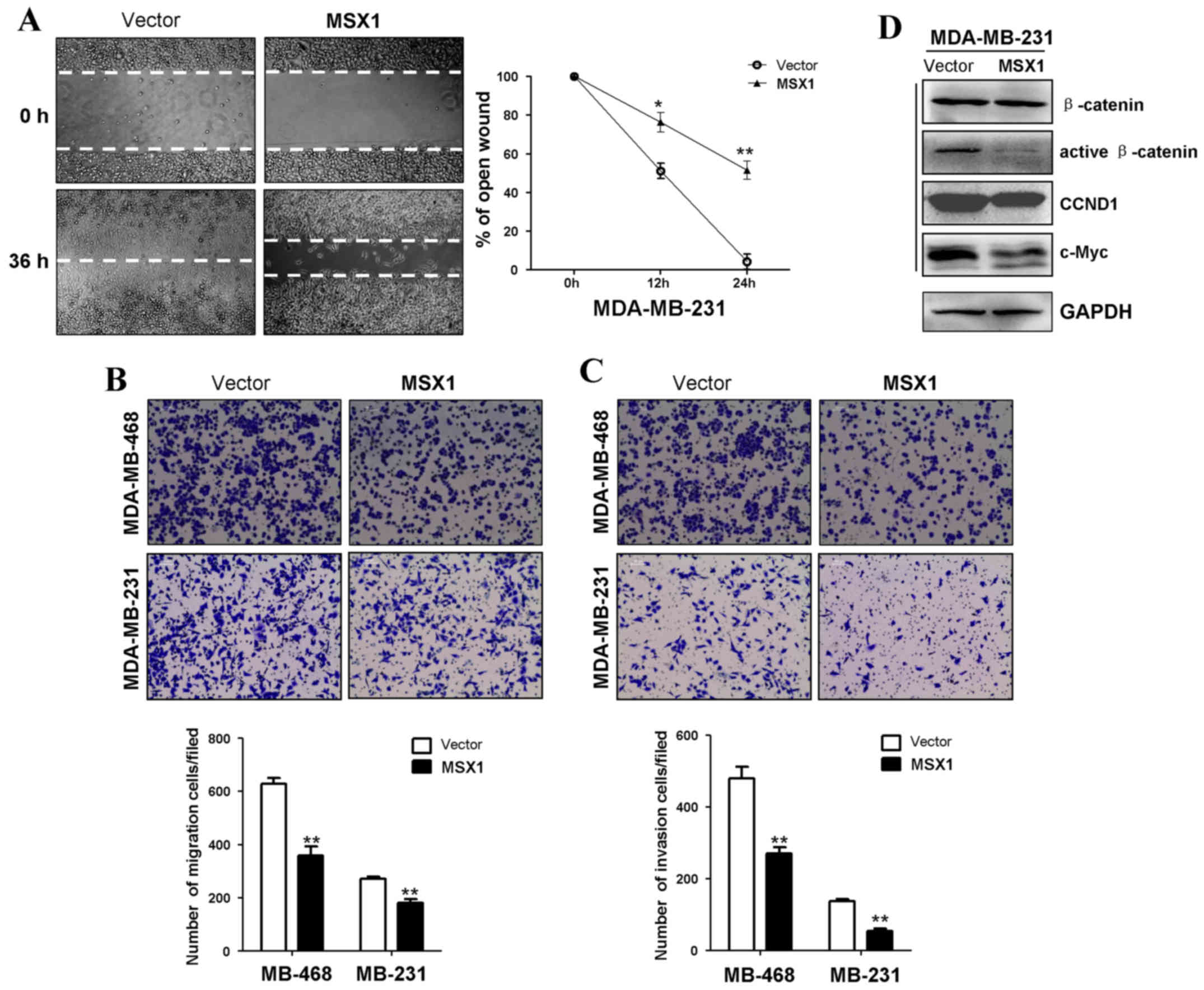
MSX1 inhibits breast tumor cell migration
and invasion
Wound-healing and Transwell assays were performed to
assess the effects of MSX1 expression on the migration and invasion
of breast tumor cells. In the scratch wound-healing assay, the
percentage wound closure of a confluent monolayer of
MSX1-expressing MDA-MB-231 cells was significantly lower compared
with that of control cells at 12 and 14 h post-wounding (p<0.05
and p<0.01, respectively; Fig.
6A). In the Transwell assays, the numbers of migrating and
invading cells were significantly reduced in the
MSX1-overexpressing cells compared with the control cells,
suggesting that MSX1 inhibits breast cancer cell migration and
invasion (p<0.01; Fig. 6B and
C).
MSX1 reduces the expression of β-catenin
and its downstream target genes
As the β-catenin signaling pathway is critical in
the regulation of cell proliferation, whether MSX1 affects this
pathway in breast cancer cells was investigated. The expression of
active β-catenin was examined using western blotting. The
expression levels of active β-catenin and its downstream targets
c-Myc and CCND1 were reduced in the MSX1-expressing MDA-MB-231
cells compared with the control cells (Fig. 6D). These results indicate that
MSX1 reduces the levels of active β-catenin and its downstream
target genes, and thus may antagonize the WNT/β-catenin signaling
pathway.
Discussion
In the present study, it was demonstrated that
MSX1 is expressed in normal breast tissues, but frequently
methylated and silenced in breast cancer cell lines and primary
tumor tissues. MSX1 was methylated in 77.8% of the breast
cancer cell lines and 47.5% of the primary breast tumors that were
tested, but not in surgical margin tissues or normal breast
tissues. Pharmacological demethylation restored the expression of
MSX1. The present study also demonstrated that MSX1 inhibits
breast cancer cell proliferation, migration and invasion by
inducing G1/S cell-cycle arrest and apoptosis, through the
downregulation of β-catenin activity. These findings indicate that
MSX1 acts as a functional tumor suppressor in breast cancer.
The epigenetic inactivation of TSGs, including
promoter methylation, histone modification and RNA interference,
serves an important role in tumor initiation and progression
(30). To the best of our
knowledge, the present study is the first to demonstrate that
MSX1 is widely expressed in normal adult tissues, including
mammary tissue, but frequently downregulated or totally silenced in
breast cancer cell lines and primary tumors, and that this is
frequently accompanied by its methylation. However, no MSX1
primer methylation was detected in certain breast cancer cell lines
(MDA-MB-231 and YCC-B1) in which MSX1 was silenced,
indicating that other epigenetic alterations, such as histone
modification, may be an alternative mechanism contributing to
MSX1 silencing, in addition to promoter CpG methylation.
The WNT signaling pathway is an evolutionarily
conserved signaling pathway that is important in the regulation of
numerous fundamental cellular processes, including cell
proliferation, migration, stemness and tumorigenesis, with a wide
range of biological activities (31). The WNT/β-catenin signaling pathway
has frequently been implicated in the development of various
cancers, and serves an important role in tumor initiation and
progression (7,11,32). MSX1 has been shown to induce the
expression of the WNT-pathway-antagonistic genes Dickkopf 1–3, and
secreted frizzled-related protein 1, thus deregulating the WNT
signaling pathway in neuroblastoma cells (7). In the present study, the ectopic
expression of MSX1 was observed to reduce the expression of active
β-catenin and its downstream target genes c-Myc and CCND1 in breast
tumor cells, and induce G1/S cell cycle arrest and apoptosis, thus
inhibiting tumor cell proliferation and metastasis, which is
consistent with a previous study of glioblastoma (33). MSX1 also interacts with p53 and
inhibits tumor growth by inducing apoptosis (18). Indeed, the present study indicated
that MSX1 induced the apoptosis of breast cancer cells by
upregulating cleaved caspase 3 and cleaved PARP.
The association of the MSX1 gene with breast
cancer and its characteristics has previously been reported. The
MSX1 gene was observed to be associated with an increased risk of
breast cancer in a Polish population and may be considered as an
early marker of the disease (11). In the present study, the
tumor-specific methylation of MSX1 was observed in breast
cancer, although no significant correlation between its methylation
status and the clinicopathological features of primary breast
cancer tumors, including clinical stage, lymph-node metastasis and
ER status was detected, which requires further confirmation in a
study with a larger sample size. Future studies are also required
to investigate whether circulating methylated MSX1 in the
serum or in combination with other methylated TSGs can be used to
detect early breast cancer.
In conclusion, the present study provides the first
evidence that MSX1 is frequently downregulated or silenced
in breast cancer by promoter CpG methylation. MSX1 acts as a
functional TSG in breast carcinogenesis through the inhibition of
active β-catenin. The methylation of MSX1 could be used as
an epigenetic biomarker for the early detection and diagnosis of
breast cancer.
Acknowledgements
The authors thank Professor Qian Tao for his
guidance for the project and comments on the manuscript, and
Professor Kathleen Kelly (National Cancer Institute, National
Institutes of Health, Bethesda, MD, USA) for all the breast cell
lines used in this paper.
Notes
[1]
Funding
The present study was supported by the National
Natural Science Foundation of China (grant nos. 81302307, 81572769
and 81372898) and special research funds from The Chinese
University of Hong Kong.
[2] Availability
of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
[3] Authors'
contributions
YiY and JF detected the cell function, CL and LL
performed the DNA bisulfite treatment and MSP examination, YuY and
WP finished the western blot experiment and wrote the paper, GR
designed the experiment and finished the figures. All authors read
and approved the final manuscript.
[4] Ethics
approval and consent to participate
The study was approved by the Ethics Committee of
the First Affiliated Hospital of Chongqing Medical University
(approval no. 2010/2012-23). All patients signed informed written
consent forms for participation in the study at initial clinical
investigation.
[5] Consent for
publication
Not applicable.
[6] Competing
interests
References
|
1
|
Torre LA, Bray F, Siegel RL, Ferlay J,
Lortet-Tieulent J and Jemal A: Global cancer statistics, 2012. CA
Cancer J Clin. 65:87–108. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Jemal A, Bray F, Center MM, Ferlay J, Ward
E and Forman D: Global cancer statistics. CA Cancer J Clin.
61:69–90. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Taby R and Issa JP: Cancer epigenetics. CA
Cancer J Clin. 60:376–392. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Esteller M: Epigenetics in cancer. N Engl
J Med. 358:1148–1159. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Hoque MO, Prencipe M, Poeta ML, Barbano R,
Valori VM, Copetti M, Gallo AP, Brait M, Maiello E, Apicella A, et
al: Changes in CpG islands promoter methylation patterns during
ductal breast carcinoma progression. Cancer Epidemiol Biomarkers
Prev. 18:2694–2700. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Xiang TX, Yuan Y, Li LL, Wang ZH, Dan LY,
Chen Y, Ren GS and Tao Q: Aberrant promoter CpG methylation and its
translational applications in breast cancer. Chin J Cancer.
32:12–20. 2013. View Article : Google Scholar :
|
|
7
|
Revet I, Huizenga G, Koster J, Volckmann
R, van Sluis P, Versteeg R and Geerts D: MSX1 induces the Wnt
pathway antagonist genes DKK1, DKK2, DKK3, and SFRP1 in
neuroblastoma cells, but does not block Wnt3 and Wnt5A signalling
to DVL3. Cancer Lett. 289:195–207. 2010. View Article : Google Scholar
|
|
8
|
Shah N and Sukumar S: The Hox genes and
their roles in oncogenesis. Nat Rev Cancer. 10:361–371. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Bhatlekar S, Fields JZ and Boman BM: HOX
genes and their role in the development of human cancers. J Mol Med
(Berl). 92:811–823. 2014. View Article : Google Scholar
|
|
10
|
McGinnis W and Krumlauf R: Homeobox genes
and axial patterning. Cell. 68:283–302. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Sliwinski T, Synowiec E, Czarny P, Gomulak
P, Forma E, Morawiec Z, Morawiec J, Dziki L, Wasylecka M and
Blasiak J: The c.469+46_56del mutation in the homeobox MSX1 gene -
a novel risk factor in breast cancer? Cancer Epidemiol. 34:652–655.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Saadi I, Das P, Zhao M, Raj L, Ruspita I,
Xia Y, Papaioannou VE and Bei M: Msx1 and Tbx2 antagonistically
regulate Bmp4 expression during the bud-to-cap stage transition in
tooth development. Development. 140:2697–2702. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Nassif A, Senussi I, Meary F, Loiodice S,
Hotton D, Robert B, Bensidhoum M, Berdal A and Babajko S: Msx1 role
in craniofacial bone morphogenesis. Bone. 66:96–104. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Bendall AJ and Abate-Shen C: Roles for Msx
and Dlx homeoproteins in vertebrate development. Gene. 247:17–31.
2000. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Nakatomi M, Wang XP, Key D, Lund JJ,
Turbe-Doan A, Kist R, Aw A, Chen Y, Maas RL and Peters H: Genetic
interactions between Pax9 and Msx1 regulate lip development and
several stages of tooth morphogenesis. Dev Biol. 340:438–449. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Ceyhan D, Kirzioglu Z and Calapoglu NS:
Mutations in the MSX1 gene in Turkish children with non-syndromic
tooth agenesis and other dental anomalies. Indian J Dent.
5:172–182. 2014. View Article : Google Scholar :
|
|
17
|
Dunwell TL, Hesson LB, Pavlova T,
Zabarovska V, Kashuba V, Catchpoole D, Chiaramonte R, Brini AT,
Griffiths M, Maher ER, et al: Epigenetic analysis of childhood
acute lymphoblastic leukemia. Epigenetics. 4:185–193. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Park K, Kim K, Rho SB, Choi K, Kim D, Oh
SH, Park J, Lee SH and Lee JH: Homeobox Msx1 interacts with p53
tumor suppressor and inhibits tumor growth by inducing apoptosis.
Cancer Res. 65:749–757. 2005.PubMed/NCBI
|
|
19
|
Yamashita S, Tsujino Y, Moriguchi K,
Tatematsu M and Ushijima T: Chemical genomic screening for
methylation-silenced genes in gastric cancer cell lines using
5-aza-2′-deoxycytidine treatment and oligonucleotide microarray.
Cancer Sci. 97:64–71. 2006. View Article : Google Scholar
|
|
20
|
Bonds J, Pollan-White S, Xiang L, Mues G
and D' Souza R: Is there a link between ovarian cancer and tooth
agenesis? Eur J Med Genet. 57:235–239. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Baylin SB and Ohm JE: Epigenetic gene
silencing in cancer - a mechanism for early oncogenic pathway
addiction? Nat Rev Cancer. 6:107–116. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Park J, Park K, Kim S and Lee JH: Msx1
gene overexpression induces G1 phase cell arrest in human ovarian
cancer cell line OVCAR3. Biochem Biophys Res Commun. 281:1234–1240.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Ringner M, Fredlund E, Hakkinen J, Borg A
and Staaf J: GOBO: gene expression-based outcome for breast cancer
online. PLoS One. 6:e179112011. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Xiang T, Li L, Yin X, Yuan C, Tan C, Su X,
Xiong L, Putti TC, Oberst M, Kelly K, et al: The ubiquitin
peptidase UCHL1 induces G0/G1 cell cycle arrest and apoptosis
through stabilizing p53 and is frequently silenced in breast
cancer. PLoS One. 7:e297832012. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Klaus A and Birchmeier W: Wnt signalling
and its impact on development and cancer. Nat Rev Cancer.
8:387–398. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Giusti AF, O' Neill FJ, Yamasu K, Foltz KR
and Jaffe LA: Function of a sea urchin egg Src family kinase in
initiating Ca2+ release at fertilization. Dev Biol.
256:367–378. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) Method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
28
|
Chesi M and Bergsagel PL: Epigenetics and
microRNAs combine to modulate the MDM2/p53 axis in myeloma. Cancer
Cell. 18:299–300. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Clevers H: Wnt/beta-catenin signaling in
development and disease. Cell. 127:469–480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Veeman MT, Axelrod JD and Moon RT: A
second canon. Functions and mechanisms of beta-catenin-independent
Wnt signaling. Dev Cell. 5:367–377. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Seidensticker MJ and Behrens J:
Biochemical interactions in the wnt pathway. Biochim Biophys Acta.
1495:168–182. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Moon RT, Kohn AD, De Ferrari GV and Kaykas
A: WNT and beta-catenin signalling: Diseases and therapies. Nat Rev
Genet. 5:691–701. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Tao H, Guo L, Chen L, Qiao G, Meng X, Xu B
and Ye W: MSX1 inhibits cell migration and invasion through
regulating the Wnt/β-catenin pathway in glioblastoma. Tumour Biol.
37:1097–1104. 2016. View Article : Google Scholar
|
|
34
|
Hu Z, Fan C, Oh DS, Marron JS, He X,
Qaqish BF, Livasy C, Carey LA, Reynolds E, Dressler L, et al: The
molecular portraits of breast tumors are conserved across
microarray platforms. BMC Genomics. 7:962006. View Article : Google Scholar : PubMed/NCBI
|