Introduction
Podocytes, also known as visceral epithelial cells,
are terminally differentiated cells of the kidney glomerulus, which
have a key role in glomerular development and maintenance (1). The cytoskeletal dynamics and
structural plasticity of podocytes, as well as the function of
received and transmitted signals, are important for glomerular
filtration and thus for renal function. Furthermore, podocyte
injury participates in the occurrence and development of human and
experimental glomerular diseases (2), among which, membranous nephropathy
(MN) is recognized as an organ-specific autoimmune disease, which
is well known to be associated with podocyte injury (3). The subepithelial immune complexes in
MN in renal tissue immunofluorescence have been reported to contain
circulating autoantibodies, intrinsic podocyte antigens (4) and complement components. Serum
autoantibodies to phospholipase-A2-receptor have been suggested not
only to be crucial in the advancement of MN, but also to act as a
biomarker for diagnosis, disease activity and treatment efficiency
(5).
Antibodies, also known as immunoglobulins (Igs), are
expressed by B lymphocytes; however, accumulating evidence has
indicated that Igs, including IgG, IgA and IgM, are produced in
non-B lineage cells, such as epithelial cancer cells (6–9)
and leukemic cells (10–12). Furthermore, several types of
normal cells, including germ cells (13,14), neurons (15,16), endothelial cells (17) and skin epidermal cells (18), can also express Igs. Although
non-B cell-derived Ig and traditional Ig have some similar
characteristics, the former possesses unique characteristics,
particularly with regards to structure and function. In contrast to
the diversity of the Ig variable region, non-B cell-derived Ig
tends to exhibit specific or restricted usage of certain sequences
(11,19). It has a unique glycosylation
profile, through which the RP215 monoclonal antibody (mAb) can
recognize non-B cell-derived Ig (20,21). According to the results of
previous studies, these Igs are essential for cell vitality and
proliferation (6,22–24), and are also associated with the
migration, invasiveness and metastasis of cancer cells, thus
suggesting that non-B Igs participate in tumorigenesis and
development (25,26). Furthermore, the regulatory
mechanisms of gene expression have been explored in several studies
(27–29).
The present study aimed to explore the expression of
IgG in podocytes, and demonstrated that human podocytes may express
and secrete IgG. In addition, not only was transcription of the
constant region detected, V(D)J rearrangement of the variable
region was demonstrated, and the expression levels of certain
proteins were verified. Furthermore, the results suggested that
podocyte-derived IgG may participate in the maintenance of cell
morphology and growth.
Materials and methods
Podocyte culture
The conditionally immortalized human podocyte cell
line was previously established, as reported in a previous study
(30), and was donated by
Professor Moin A. Saleem (Children's Renal Unit and Academic Renal
Unit, University of Bristol, Bristol, UK). The study was approved
by the ethics committee of Peking University Third Hospital
(approval document issued on February 17th, 2016; under no. 053).
The cells were cultured under growth-permissive conditions in
RPMI-1640 medium (Gibco; Thermo Fisher Scientific, Inc., Waltham,
MA, USA) supplemented with 1% insulin-transferrin-selenium-A
supplement (ITS; Gibco; Thermo Fisher Scientific, Inc.), 100 U/ml
penicillin, 0.1 mg/ml streptomycin (Gibco; Thermo Fisher
Scientific, Inc.) and 10% fetal bovine serum (FBS; Australian
origin; Biological Industries USA, Cromwell, CT, USA) at 33°C.
Podocytes were then incubated at 37°C without ITS for 10–14 days
for differentiation; these cells were used for subsequent
experiments. Protein and mRNA samples were collected from podocytes
after 48 h stimulation with or without Staphylococcus aureus
(SAC; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at 37°C.
Immunofluorescence
The podocytes were cultured on cover-slips, which
were fixed in 4% paraformaldehyde for 30 min at room temperature.
Subsequently, 0.2% Triton X-100 was used to permeabilize the cells
for 15 min at room temperature. The slides were washed in PBS and
blocked with 5% bovine serum albumin (BSA; Invitrogen; Thermo
Fisher Scientific, Inc.) at room temperature for 30 min, after
which they were incubated with primary antibodies at 4°C overnight.
The antibodies used were as follows: Rabbit anti-human IgG heavy
chain (Igγ) (ab109489; 1:150), anti-human Igκ (ab134929; 1:250),
anti-human Igλ (ab124719; 1:250) (all from Abcam, Cambridge, MA,
USA), mouse anti-human Igγ4 (GI-0910; 1:200; Beijing Xiya Golden
Bridge Biotechnology Co., Ltd., Beijing, China), rabbit anti-human
F-actin (bs-1571R; 1:500; Beijing Bioss Biotechnology Co., Ltd.,
Beijing, China) and RP215 mAb (dilution, 1:200; donated by
Professor Xiaoyan Qiu, Peking University, Beijing, China), which
specifically identified a carbohydrate-associated epitope on non-B
cell-derived Igγ. PBS was used as a negative control. After washing
in PBS, the slides were incubated with fluorescein
isothiocyanate-labeled goat anti-rabbit (ZF-0311; 1:200) or goat
anti-mouse IgG antibodies (ZF-0312; 1:200) (both from Beijing
Zhongshan Golden Bridge Biotechnology Co., Ltd., Beijing, China) at
room temperature for 1 h. Nuclei were stained with DAPI (C0060; 15
µg/ml; Beijing SUOLAIBAO Biotechnology Co., Ltd., Beijing,
China). Images were captured under a Leica DFC300 FX fluorescence
microscope (Leica Microsystems GmbH, Wetzlar, Germany).
Protein extraction and western
blotting
Podocytes were lysed in TSD lysis buffer (1% SDS, 50
mM pH 7.5 Tris-HCL, 50 mM DTT) containing protease inhibitors
(Applygen Technologies Inc., Beijing, China). Total protein
concentrations were measured using a bicinchoninic acid kit
(Applygen Technologies Inc.). Following centrifugation at 12,000 ×
g for 10 min at 4°C, the lysate supernatants were prepared for
western blotting. Human serum (obtained from one of the authors,
female, 26 years old) enriched with Igs was used as a positive
control. The cell supernatant was purified by affinity
chromatography using protein G sepharose; the target protein was
purified according to the manufacturer's protocol (R8300; Beijing
SUOLAIBAO Biotechnology Co., Ltd.). The protein samples then
underwent SDS-PAGE, western blot analysis and mass spectrometry,
which was conducted by Beijing Protein Innovation (Beijing,
China).
Western blotting was carried out according to
standard procedures. Briefly, lysate proteins were separated by 10%
SDS-PAGE and were transferred onto a nitrocellulose membrane.
Subsequently, the membrane was blocked in 5% BSA (Amresco, LLC,
Solon, OH, USA) and was incubated with primary antibodies at 4°C
overnight, including rabbit anti-human Igγ (ab109489; 1:1,000),
rabbit anti-human Igγ1 (ab108969; 1:1,000), rabbit anti-human Igγ2
(ab134050; 1:10,000), rabbit anti-human Igγ3 (ab109761; 1:5,000),
rabbit anti-human Igγ4 (ab109493; 1:5,000) and anti-Igλ (ab124719;
1:20,000), rabbit anti-human β-actin (ab8227; 1:2,000), rabbit
anti-human GAPDH (ab181602; 1:10,000) (all from Abcam), mouse
anti-human Igκ (GTX21050; GeneTex, Inc., Irvine, CA, USA) and RP215
mAb. The membrane was then incubated with goat anti-rabbit (cat.
no. 926-32211) or anti-mouse (cat. no. 926-32210) IgG-IRDyeTM680CW
secondary antibodies (1:10,000; LI-COR Biosciences, Lincoln, NE,
USA) at room temperature for 1 h. The results were analyzed using
an Odyssey Infrared imager and Odyssey V3.0 software (LI-COR
Biosciences).
Isolation of peripheral blood mononuclear
cells (PBMCs)
Mononuclear cells were prepared from the peripheral
blood of a healthy person (author of the present study as mentioned
above). PBMCs were isolated using Ficoll-Paque PLUS (GE Healthcare,
Chicago, IL, USA) by density-gradient centrifugation. The cells
from the interface of the solution were considered PBMCs, which
were collected and washed in PBS. The isolated PBMCs were
immediately utilized.
Reverse transcription-polymerase chain
reaction (RT-PCR)
Total RNA was extracted from podocytes and PBMCs
according to a standard procedure using TRIzol reagent (Invitrogen;
Thermo Fisher Scientific, Inc.). RNA concentration was estimated
using a NanoDrop spectrophotometer (NanoDrop Technologies; Thermo
Fisher Scientific, Inc., Wilmington, DE, USA). Subsequently, RT was
performed with oligo(dT) primer using GoScript™ Reverse
Transcriptase (Promega Corporation, Madison, WI, USA) according to
the manufacturer's protocol. Transcripts of the Igγ, Igκ, Igλ and
Iγ-Cγ constant regions (31) were
amplified by PCR using Taq PCR MasterMix (Beijing Biomed
Biotechnology Co., Ltd., Beijing, China). Transcripts of the Igγ
and Igκ variable regions were amplified by semi-nested PCR with Taq
PCR MasterMix (Biomed). The PCR products were separated by
electrophoresis on a 1.5% agarose gel at a constant voltage of 120
V for 40–50 min. The thermocycling conditions were as mentioned in
previous studies (6,18).
To detect the mRNA expression levels of
recombination activating gene (RAG)1 and RAG2, a nested RT-PCR
assay was used (32). Total RNA
was treated with RQ1 RNase-free DNase (Promega Corporation) to
eliminate the contamination of genomic DNA. The cDNA template was
replaced with treated RNA as a negative control. For amplification
of activation-induced cytidine deaminase (AID), the primer
sequences and protocol were used as previously described (33). Details of the primer sequences
used in the present study are listed in Table I. The reliability of PCR products
was analyzed by DNA sequence analysis, which was performed by
Invitrogen Trading (Shanghai) Co., Ltd. (Shanghai, China).
 | Table ISequences of polymerase chain
reaction primers used in the present study. |
Table I
Sequences of polymerase chain
reaction primers used in the present study.
| Gene name | Primer | Primer sequence
(5′-3′) | Product length
(bp) |
|---|
| Ig Vγ | External sense |
VH1:GAGGTGCAGCTCGAGGAGTCTGGG | |
|
VH2:CAGGTGCAGCTCGAGCAGTCTGGG | |
|
VH3:CAGGTACAGCTCGAGCAGTCAGG | |
|
VH4:CAGGTGCAGCTGCTCGAGTCGGG | 280–300 |
| External
antisense |
CH1:ACACCGTCACCGGTTCGG | |
| Internal
antisense |
FR2:TGGRTCCGVCAGSCYCCNGG | |
| Internal
antisense |
JH:AACTGCAGAGGAGACGGTGACC | |
| Ig Vκ | External sense |
GACATCGAGCTCACCCAGTCTCC | 340–360 |
| External
antisense |
CGGGAAGATGAAGACAGATGGTGC | |
| Internal sense |
GAAATTGAGCTCACGCAGTCTCCA | |
| Internal
antisense |
TGGTGCAGCCACAGTTCGTT | |
| Ig Cγ | Sense |
AGGACTCTACTCCCTCAGCAG | 566 |
| Antisense |
TCAGGCTGACCTGGTTCTTG | |
| Ig Cκ | Sense |
TGAGCAAAGCAGACTACGAGA | 231 |
| Antisense |
GGGGTGAGGTGAAAGATGAG | |
| Ig Cλ | Sense |
GGGACCAAGCTCACCGTCCTAG | 316 |
| Antisense |
TCTTCTCCACGGTGCTCCCTTC | |
| Ig Cγ1 | Sense |
GGGCTTCCAAGCCAACAGGGCAGGACA | 603 |
| Antisense |
GTTTTGTCACAAGATTTGGGCTC | |
| Ig Cγ2 | Sense |
GGGCTTCCAAGCCAACAGGGCAGGACA | 597 |
| Antisense |
GTGGGCACTCGACACAACATTTGCG | |
| Ig Cγ3 | Sense |
AGGTGGGCAGGCTTCAGGCACCGAT | 670 |
| Antisense |
TTGTGTCACCAAGTGGGGTTTTGAGC | |
| Ig Cγ4 | Sense |
GATGGCGTGGAGGTGCATAA | 212 |
| Antisense |
GTGTACACCTGTGGCTCTCG | |
| AID | Sense |
GAGGCAAGAAGACACTCTG | 647 |
| Antisense |
GTGACATTCCTGGAAGTTGC | |
| RAG1 | External sense |
TGGATCTTTACCTGAAGATG | 327 |
| External
antisense |
CTTGGCTTTCCAGAGAGTCC | |
| Internal sense |
CACAGCGTTTTGCTGAGCTC | |
| Internal
antisense |
AGCTTGCCTCAGGGTTCATG | |
| RAG2 | External sense |
TGGAAGCAACATGGGAAATG | 193 |
| External
antisense |
CATCATCTTCATTATAGGTGTC | |
| Internal sense |
TTCTTGGCATACCAGGAGAC | |
| Internal
antisense |
CTATTTGCTTCTGCACTG | |
| CD19 | Sense |
AAGGGGCCTAAGTCATTGCT | 347 |
| Antisense |
CACGTTCCCGTACTGGTTCT | |
| β-actin | Sense |
AGAGCTATGAGCTGCCTGAC | 121 |
| Antisense |
AATTGAATGTAGTTTCATGGATG | |
Analysis of rearrangement gene
PCR products of the Igγ and Igκ variable regions
obtained from both PBMCs and podocytes were respectively cloned
into a pGEM-T Easy Vector System I (A1360; Promega Corporation),
which was transfected into Competent Escherichia coli TOP10
cells (Tiangen Biotech Co., Ltd., Beijing, China). The transfection
experiments were performed according to the manufacturer's
protocols. Briefly, ligation reactions were set up using
transfection reagents in pGEM-T Easy Vector System I and the
transformations were performed using the ligation reactions.
Subsequently, clones were formed and amplified through the
proliferation of bacteria. Following DNA sequencing with an ABI
3730XL Genetic Analyzer (Applied Biosystems; Thermo Fisher
Scientific, Inc.), which was performed by Invitrogen Trading
(Shanghai) Co., Ltd., the variable sequences were compared with
germline gene segments using Basic Local Alignment Search Tool in
the National Center for Biotechnology Information (NCBI) database
(https://blast.ncbi.nlm.nih.gov/Blast.cgi).
Small interfering (si)RNA
transfection
Synthetic siRNA targeting the human Igγ chain
constant region and control siRNAs were purchased from Shanghai
GenePharma Co., Ltd. (Shanghai, China). The siRNA sequences were as
follows, siRNA-1, 5′-GCA AGG AGU ACA AGU GCA ATT-3′, siRNA-2,
5′-CCG GAG AAC AAC UAC AAG ATT-3′, siRNA-3, 5′-CAC AAC CAC UAC ACA
CAG ATT-3′, siRNA-positive control, 5′-UGA CCU CAA CUA CAU GGU
U-3′, siRNA-negative control, 5′-UUC UCC GAA CGU GUC ACG UTT-3′.
Lipofectamine 3000 transfection reagent (Invitrogen; Thermo Fisher
Scientific, Inc.) was used to transfect the siRNAs into the
podocyte cell line. Following transfection, which was conducted
according to the manufacturer's protocol, the knockdown efficiency
was detected by western blotting. Approximately 3×105
cells/well were cultured in complete medium with a final siRNA
concentration of 50 nM. The cells were transfected at 37°C for 48 h
and subsequently collected for further experiments.
Cell viability assay
Cell viability was measured by Cell Counting kit-8
(CCK-8; Dojindo Molecular Technologies, Inc., Kumamoto, Japan).
Briefly, cells from each siRNA group were seeded into 3 wells of a
96-well plate for 48 h at 37°C (1,000–2,000 cells per well).
Subsequently, the medium was replaced with 100 µl fresh
culture medium containing 8 µl CCK-8 reagent. After
incubation for 2 h at 37°C, a spectrophotometric microplate reader
(BioTek Instruments, Inc., Winooski, VT, USA) was used to measure
the absorbance at 450 nm; cell viability was indirectly
reflected.
Cell adhesion assay
To analyze the adhesive ability of the cells, a
CCK-8 assay (Dojindo Molecular Technologies, Inc.) was conducted
following siRNA transfection for 48 h. Briefly, 1×104
cells resuspended in 100 µl media were seeded in each well
of a 96-well plate and were incubated at 37°C for 1 h. Each group
(3 wells per group) including siRNA-1, siRNA-2, siRNA-3, siRNA-NC,
mock and blank, was divided into 3 groups: washed, unwashed and
blank group. In the washed group, the cells were washed gently with
PBS three times and 100 µl fresh media with 8 µl
CCK-8 reagent was added to the wells. In the unwashed group, the
cells were added with 100 µl fresh media with 8 µl
CCK-8 reagent. In the blank group, CCK-8 was directly added into
the wells without cells. After incubation for 2 h at 37°C, the
adhered cells were determined by detecting the absorbance at 450 nm
using a spectrophotometric microplate reader (BioTek Instruments,
Inc.). Cell adhesion was determined using the following
formula:
Cell adhesion rate=ODwashed−ODblankODunwashed−ODblank
Statistical analysis
Data are expressed as the means ± standard deviation
and were analyzed by SPSS 20.0 software (IBM Corp., Armonk, NY,
USA). All experiments were repeated 3 times. To determine
significant differences between two groups, Student's t-test was
used. The significant differences between more than two groups were
analyzed using one-way analysis of variance and a least significant
difference multiple comparison test. P<0.05 was considered to
indicate a statistically significant difference.
Results
IgG heavy and light chains were stained
in human podocytes by immunofluorescence staining
To determine the expression of IgG in podocytes, the
present study conducted an immunofluorescence analysis using
commercially available anti-human Igγ, Igγ1-4, Igκ and Igλ light
chain antibodies; and RP215 mAb, which recognizes non-B
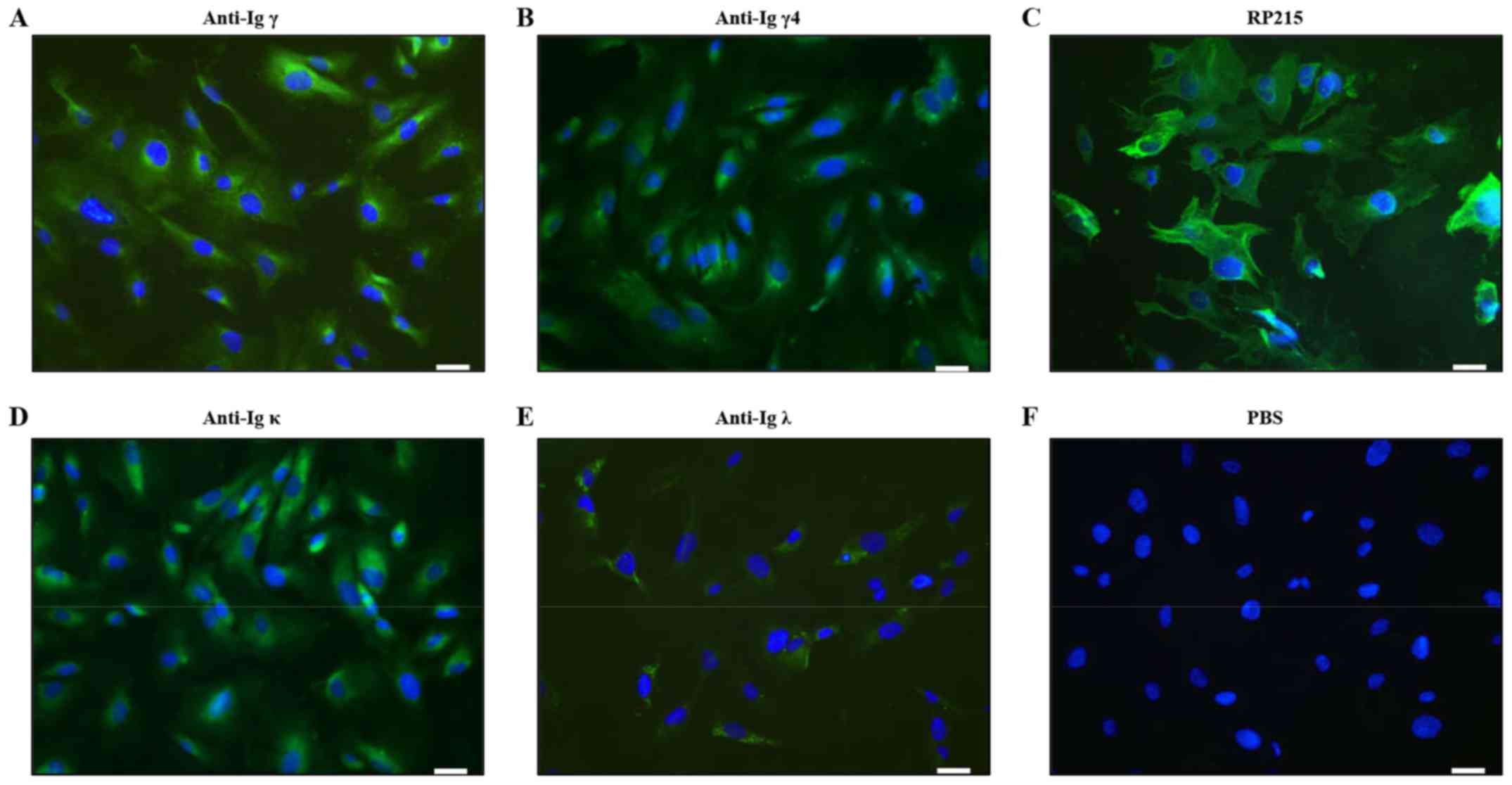
cell-derived Igγ (34) (Fig. 1). Positive staining of Igγ, γ4, κ
and λ was detected in the cytoplasm of podocytes (Fig. 1A, B, D and E). However, Igγ2 and
Igγ3 staining was not found by immunofluorescence staining.
Stronger positive staining of RP215 was predominantly localized in
the cytoplasm and on the cell membrane of podocytes (Fig. 1C).
Confirmation of the expression of IgG in
podocytes by western blotting and mass spectrometry
To confirm the expression of corresponding proteins,
IgG heavy and light chains were detected in podocyte cell lysates
and culture supernatants by western blotting. Human serum that
contains a substantial amount of Igs was used as a positive
control. In order to eliminate the interference of FBS in the
media, FBS in electrophoresis was blotted with anti-human Igγ, κ
and λ antibodies and negatively stained. The commercial anti-Igγ
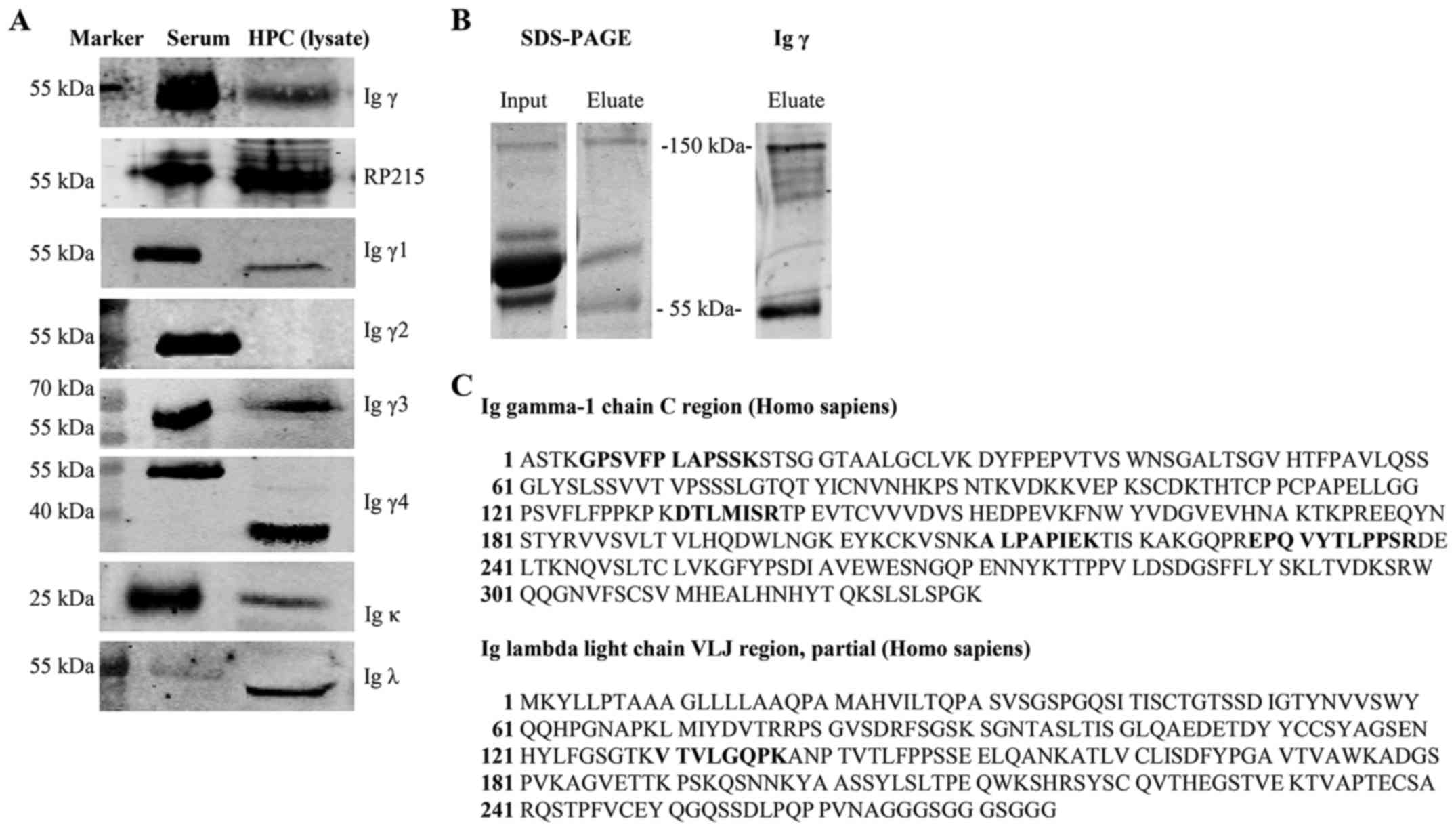
antibody and RP215 mAb detected Igγ at 55 kDa. In addition,
subclasses of Igγ, including Igγ1 (52 kDa), Igγ3 (58 kDa), Igγ4 (36
kDa), were detected and the bands obtained were consistent with the
predicted molecular weight, or the detected bands in the antibody
instructions. However, Igγ2 expression was not found after several
experiments. Furthermore, Igκ (25 kDa) and Igλ (50 kDa) expression
was observed in the cell lysate (Fig.
2A).
To further verify the secretion of IgG by podocytes,
IgG was extracted from the culture supernatant using protein G
beads, which bind to human IgG with high specificity. The protein
in the eluate was recognized at 150 kDa and 55 kDa, following
incubation with an anti-human IgG antibody. These weights
correspond with the weight of the intact tetramer at 150 kDa and
that of the single γ chain at 55 kDa (Fig. 2B). Mass spectrometry demonstrated
that the 150 kDa band contained segments of the Igγ1 chain constant
region and Igλ chain VLJ region, according to the NCBI database
(Fig. 2C), and the Igγ1 chain
constant region could be found at 55 kDa.
Transcription of IgG heavy and light
chain constant regions and related enzymes in podocytes
The expression levels of IgG genes and associated
enzymes were explored in podocytes by RT-PCR. The B lymphocyte
marker CD19 was used to exclude the possible contamination of B
lymphocytes. CD19 expression was positive in the PBMC control
group, but not in the podocyte group. The mRNA expression levels of
Igγ, κ and λ constant regions, and the Igγ subclasses, including
Igγ1, γ3 and γ4 constant regions were detected (Fig. 3A), whereas Igγ2 was not; these
results were similar to those of western blotting.
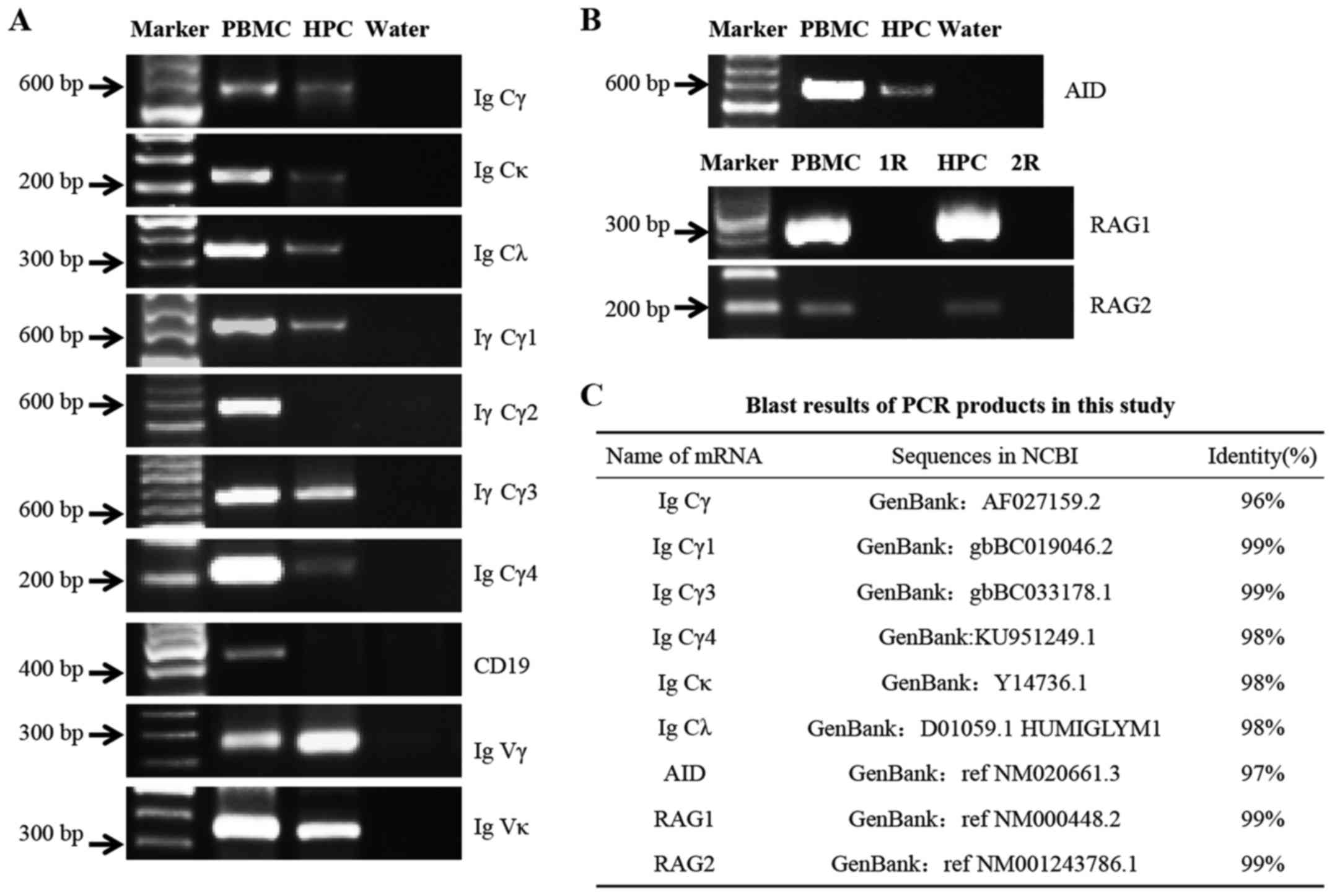 | Figure 3Detection of the mRNA expression
levels of IgG and related enzymes, including RAG1, RAG2 and AID in
podocytes. (A) Transcripts of Igγ, γ1, γ3, γ4, κ and λ C regions
were detected by RT-PCR. CD19 was detected as a marker of B
lymphocytes. The mRNA of Igγ and κ V regions was amplified by
nested RT-PCR. (B) mRNA expression levels of AID, RAG1 and RAG2.
PBMCs were used as a positive control; water instead of cDNA was
used as a blank control; R (cDNA template replaced by DNase-treated
RNA) was used as negative control. (C) DNA sequencing results. The
sequences of the PCR products were aligned with the mRNA sequences
available in the NCBI database. HPC, human podocytes; AID,
activation-induced cytidine deaminase; C, constant; Ig,
immunoglobulin; Igγ, IgG heavy chain; PBMCs, peripheral blood
mononuclear cells; RAG. recombination activating gene; RT-PCR,
reverse transcription-polymerase chain reaction; V, variable. |
Notably, the mRNA expression levels of AID, which is
the essential element for somatic hypermutation and class switch
recombination (CSR) in B lymphocytes, and of RAG1 and RAG2, which
are necessary for V(D)J rearrangement, were detected in human
podocytes (Fig. 3B), thus
indicating that class switching of Igγ and gene rearrangement of
the variable region may occur in podocytes.
After sequencing, the sequences of PCR products were
aligned with the mRNA sequences available in the NCBI database;
homologies between 96 and 99% were demonstrated (Fig. 3C).
V(D)J rearrangement of the IgG variable
region and rearrangement patterns
V(D)J rearrangements take place at the genomic level
prior to the expression of corresponding proteins in B lymphocytes.
By using semi-nested RT-PCR and DNA sequence analysis, the present
study identified gene recombination sequences of IgG heavy and
light chains in podocytes (Fig.
3A). T-A cloning and sequencing demon strated that the Igγ and
Igκ genes from clones had typical V(D)J rearrangements, with the
V-D, D-J or V-J junctions (data not shown). All sequences obtained
were productive and there were no stop codons in each region.
Notably, in contrast to the diverse gene
rearrangements in B lymphocytes, the present study detected
conservative V(D)J patterns in podocyte-derived Ig, with
VH3-23/D3-22/JH4 (12/22), VH3-23/D3-16/JH4 (8/22) and
VH3-23/D4-17/JH6 (2/22) as the predominant rearrangements in the γ
chain variable region (Table
II), and Vκ1-39/Jκ1 (10/10) in the κ chain variable region
(Table III).
| Table IIIGHV/IGHD/IGHJ rearrangement patterns
of the Ig heavy chain variable region in podocytes and PBMCs. |
Table II
IGHV/IGHD/IGHJ rearrangement patterns
of the Ig heavy chain variable region in podocytes and PBMCs.
| Name of cells | No. of clones | IGHV/IGHD/IGHJ
usage | Identity with
germlines (%) |
|---|
| Podocytes (NC) | 12 |
IGHV3-23*01/IGHD3-22*01/IGHJ4*02 | 88.8–90.6 |
| 8 |
IGHV3-23*01/IGHD3-16*02/IGHJ4*02 | 85.9–87.0 |
| 2 |
IGHV3-23*01/IGHD4-17*01/IGHJ6*03 | 89.0–91.6 |
| Podocytes
(SAC) | 6 |
IGHV3-23*01/IGHD1-7*01/IGHJ6*03 | 91.6–94.8 |
| 6 |
IGHV3-23*01/IGHD2-8*01/IGHJ6*02 | 84.6–86.3 |
| 10 |
IGHV3-30-3*03/IGHD3-16*02/
IGHJ6*02 | 88.0–90.1 |
| 1 |
IGHV3-30-3*01/IGHD2-15*01/
IGHJ4*02 | 96.9 |
| PBMCs
(control) | 1 |
IGHV5-51*01/IGHD3-22*01/IGHJ3*02 | 96.9 |
| 1 |
IGHV3-7*03/IGHD1-26*01/IGHJ4*02 | 92.3 |
| 1 |
IGHV3-11*05/IGHD1-26*01/IGHJ4*02 | 95.4 |
| 1 |
IGHV6-1*02/IGHD5-24*01/IGHJ4*02 | 93.9 |
| 1 |
IGHV3-33*03/IGHD5-24*01/IGHJ6*03 | 96.9 |
| 1 |
IGHV3-7*01/IGHD3-16*02/IGHJ4*02 | 91.7 |
| Table IIIIGKV/IGKJ rearrangement patterns of
the Igκ variable region in podocytes and PBMCs. |
Table III
IGKV/IGKJ rearrangement patterns of
the Igκ variable region in podocytes and PBMCs.
| Name of cells | No. of clones | IGKV/IGKJ
usage | Identity with
germlines (%) |
|---|
| Podocytes (NC) | 10 |
IGKV1-39*01/IGKJ1*01 | 92.9–93.6 |
| Podocytes
(SAC) | 3 |
IGKV1-39*01/IGKJ1*01 | 90.1–90.5 |
| 8 |
IGKV1-12*01/IGKJ4*01 | 90.1–95.3 |
| PBMC (control) | 1 |
IGKV1-9*01/IGKJ1*01 | 91.5 |
| 1 |
IGKV1-12*01/IGKJ4*02 | 83.5 |
| 1 |
IGKV1-39*01/IGKJ2*02 | 95.3 |
| 1 |
IGKV1-39*01/IGKJ5*01 | 88.0 |
| 1 |
IGKV2-29*02/IGKJ2*01 | 94.7 |
| 2 |
IGKV4-1*01/IGKJ2*01 | 98.0 |
| 2 |
IGKV4-1*01/IGKJ3*01 | 96.4 |
| 1 |
IGKV4-1*01/IGKJ4*01 | 92.7 |
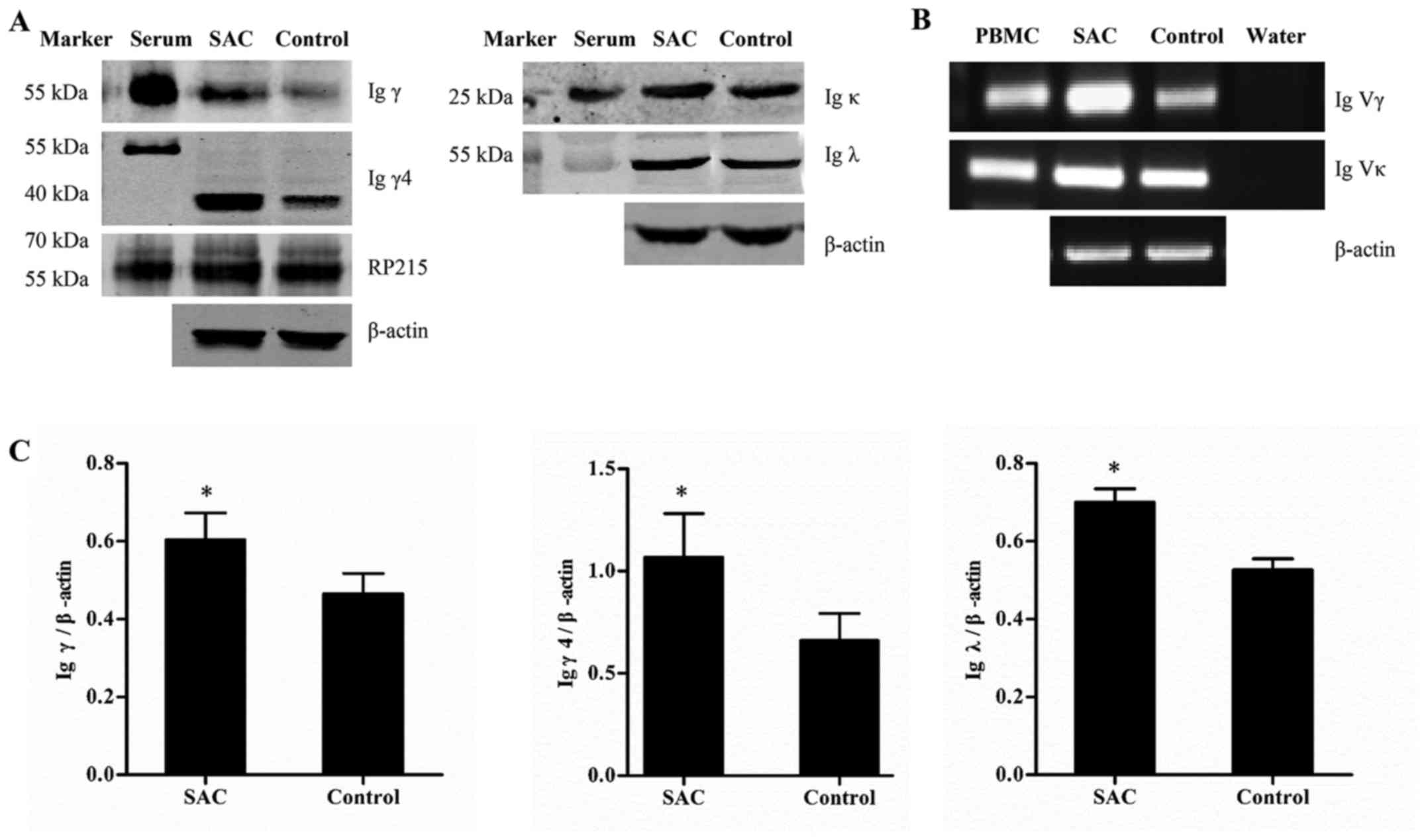
SAC upregulates the expression of IgG and
alters the conservative variable regions in podocytes
A total of 48 h after stimulation with SAC, Igγ, γ4
and λ were significantly upregulated (P<0.05), and the mRNA
expression levels of the Igγ and Ig κ variable regions were
increased, thus indicating that SAC upregulated the expression of
IgG in podocytes (Fig. 4).
Furthermore, the conservative rearrangements in the variable
regions were altered following SAC incubation. In the SAC group,
VH3-23/D1-7/JH6, VH3-23/D2-8/JH6 and VH3-30/ D3-16/JH6
rearrangements were found in the γ chain variable region (Table II). Vκ1-39/Jκ1 and Vκ1-12/Jκ4
rearrangements were found in the κ chain variable region (Table III)..
Downregulation of IgG following siRNA
transfection, and its role in cell viability and adhesion
The present study used three siRNAs (siRNA 1-3)
against the Igγ chain constant region. A total of 48 h
post-transfection, western blot analysis was used to detect Igγ
expression. The protein expression levels of Igγ were significantly
reduced in podocytes by siRNA-1 (Fig.
5A), thus indicating that siRNA-1 was effective.
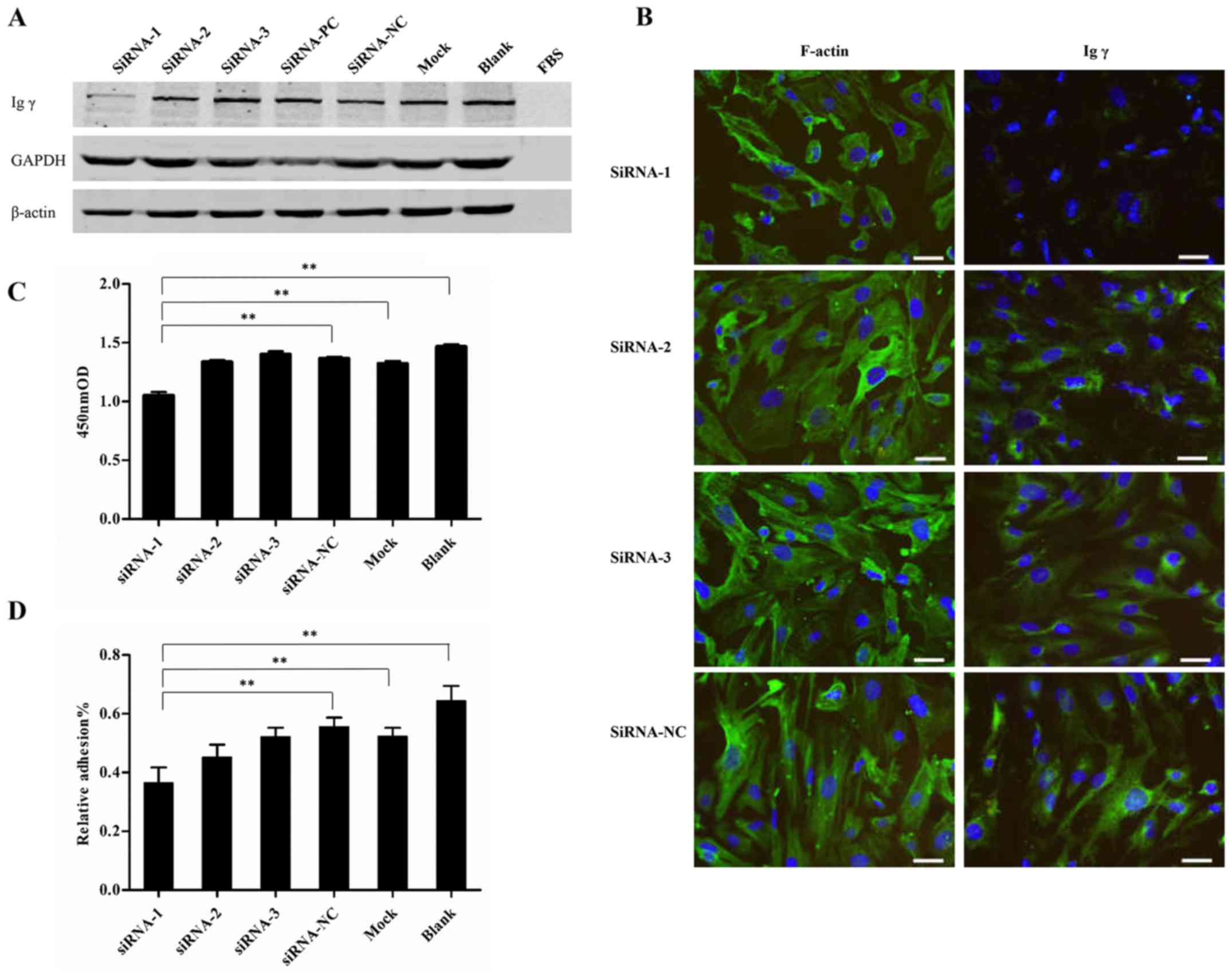 | Figure 5IgG expression 48 h post-transfection
with siRNA. (A) Western blot analysis confirmed successful
knockdown of IgG in podocytes by siRNAs. GAPDH was used as a
positive control. (B) Immunofluorescence assay of F-actin and Igγ
staining. F-actin and IgG were immunostained (green), and nuclei
were stained with DAPI (blue).(C) CCK-8 analysis of cell viability.
(D) CCK-8 analysis of the adhesive ability of podocytes. siRNA-1,
siRNA-2 and siRNA-3, siRNAs targeted the Igγ constant region;
siRNA-PC, siRNA targeted GAPDH; siRNA-NC, scrambled siRNA; mock,
cells treated with only Lipofectamine 3000; blank, untreated cells;
FBS, culture medium without cells. Scale bar, 50 µm. Data
are presented as the means ± standard deviation, n=3. **P<0.01.
CCK-8, Cell Counting kit-8; FBS, fetal bovine serum; Ig,
immunoglobulin; Igγ, IgG heavy chain; NC, negative control; OD,
optical density; PC, positive control. |
To further confirm the involvement of
podocyte-derived IgG in podocyte function, the alterations in cell
morphology, viability and adhesion were detected in each group.
Immunofluorescence assay using F-actin staining demonstrated that
cell volume was reduced and morphology was altered in response to
IgG expression knockdown in the siRNA-1 group (Fig. 5B). Furthermore, in the siRNA-1
group, viability was reduced compared with in the other groups,
including siRNA-NC, Mock and Blank (1.05±0.05 vs. 1.37±0.02,
1.32±0.04 and 1.47 ±0.03, respectively) (P<0.01). In addition,
adhesive capability was significantly reduced in the siRNA-1 group
compared with in the other groups, including siRNA-NC, Mock and
Blank (0.37±0.05 vs. 0.56±0.03, 0.52±0.03 and 0.65±0.05%) (Fig. 5C and D) (P<0.01), as determined
by CCK-8 assay.
Discussion
The present study detected the expression levels of
IgG heavy and light chains at the protein and mRNA levels in
podocytes. The results demonstrated that the expression was
associated with cell viability and adhesion.
Firstly, the present study used immunofluorescence
to determine the expression and distribution of Igγ, γ1-4 heavy
chains, and κ and λ light chains in podocytes. Positive staining of
Igγ, γ4, κ and λ chains was detected in the cytoplasm of podocytes,
thus indicating that podocytes may express IgG. Shao et al
(35) reported that IgM was
expressed and presented in kidney tubules in µMT mice, which
have hardly any IgM in circulation due to the disruption of the
µ heavy chain, thus lacking mature B cells. Compared with
commercial antibodies, the RP215 antibody recognizes a specific
carbohydrate-associated epitope on non-B cell-derived Igγ (34); in the present study, RP215 mAb
detected IgG more clearly both on the cell membranes and in the
cytoplasm, suggesting that podocyte-derived IgG was more likely to
be non-B cell derived as the findings of Liao et al
(26). The positive staining of
Igγ4 is consistent with the fact that IgG4 is the predominant
subtype of immune complexes in MN, which is the most common cause
of adult nephrotic syndrome (3).
Western blotting confirmed that the Igγ protein band
was recognized by a commercial anti-Igγ antibody at 55 kDa, thus
indicating that podocyte-derived Ig may have a similar molecular
size to that in circulation. Similar to the immunofluorescence
findings, RP215 mAb binding suggested that podocyte-derived IgG was
more similar to non-B cell-derived IgG and contained a unique
glycosylated epitope (25). In
addition, the present study detected subclasses of Igγ, with Igγ1
and γ3 located at 52 and 58 kDa, respectively, which were near the
molecular weight of Igγ in the serum. Igγ4 was located at 36 kDa,
which was consistent with the predicted molecular weight in the
antibody instructions. Furthermore, Igκ in the podocyte lysate had
the same weight as in the serum at 25 kDa, whereas Igλ was detected
at 50 kDa, just two times the weight of a single λ chain, which may
be attributable to modification or polymerization. To determine if
podocytes secreted IgG, protein G sepharose and affinity
chromatography were conducted; IgG with a molecular weight of 150
kDa, as an intact tetramer, and 55 kDa, as a single heavy chain,
were obtained in the supernatant. Mass spectrometry indicated that
the 150 kDa band contained segments of Igγ1 constant region and λ
chain, whereas the 55 kDa band contained peptides from the Igγ1
chain constant region, further suggesting that podocytes may
synthesize and secrete full length IgG.
Ig gene recombination and transcription are
prerequisites for protein production. In the present study, we
first eliminated B lymphocyte contamination by detecting the CD19
expression and then obtained the positive amplification of the mRNA
of Igγ, γ1, γ3, γ4 subclasses, κ and λ constant regions. After DNA
sequencing, the PCR products conformed to the Ig mRNA sequences
contained in the NCBI database, indicating that transcription of
the IgG gene occurs in podocytes. During the process of B
lymphocyte development, the variable region of the IgG heavy and
light chains undergoes rearrangement, thus generating antibody
diversity (36). Furthermore,
podocytes have the ability to amplify functional transcripts of Igγ
and κ light chain variable regions. The present study compared the
podocyte-derived V(D)J rearrangements of Ig transcripts with
conventional V(D)J rearrangements in B cells. Although they
displayed some common characteristics, including different
junctions between V-D, D-J or V-J segments, numerous gene
rearrangement differences existed between podocytes and B
lymphocytes. Firstly, podocyte-derived Igγ and κ chains exhibited
restricted rearrangement patterns compared with B cells, which
exhibit Ig diversity; this result is in accordance with the
findings of other studies (12,18). Secondly, more common V segments
were detected in podocyte as the VH3 gene family was present in all
clones with VH3-23 positive in 34 clones (75.6%) and VH3-30 in 11
clones (24.4%), which was similar to the reports of Glanville et
al (37). Finally, some
stimulating factors, including SAC may alter the selection trends
of rearrangement patterns in podocytes.
Gene rearrangement is initiated by RAG1 and RAG2
specifically binding to the recombination signal sequence.
Subsequently, this compound cleaves DNA and induces the
recombination of V, D and J segments (38). The present study detected RAG1 and
RAG2 mRNA expression in human podocytes by nested RT-PCR, as in
other studies (6,9). When the body is stimulated by
antigens, somatic hypermutation and CSR induce the generation of
additional gene alterations. The present study demonstrated that
transcription of AID, which is an important mediator of CSR and
somatic hypermutation, exists in podocytes. These results indicated
that V(D)J rearrangement and CSR may take place in podocytes, with
a similar mechanism to in B lymphocytes.
Previous studies have suggested that non-B
cell-derived Igs are associated with cell growth and proliferation
(6,23,24). In the kidney, cytoskeletal
dynamics determine the correct morphology of podocytes, and the
slit diaphragm complex, actin cytoskeleton and cell adhesion
molecules form an intricate network to stabilize filtration barrier
function (39,40). Conversely, foot process fusion is
a pathobiological manifestation of glomerular diseases, including
MN. To investigate the association between IgG expression and
cellular functions, siRNA targeting the Igγ constant region was
transfected into podocytes. In response to IgG knockdown, cell
viability and adhesion were reduced, and alterations were detected
in the actin cytoskeleton, thus indicating that IgG expression was
associated with cellular bioactivities, including survival,
proliferation and adhesion (6).
In conclusion, the present study demonstrated that
human podocytes exhibit the capacity to produce IgG, and IgG
expression is significantly upregulated when podocytes are
stimulated by SAC. In addition, downregulation of IgG may have
negative consequences for cell survival, viability and adhesion.
The present study is the first, to the best of our knowledge, to
indicate that IgG may be expressed in podocytes and provides
additional evidence to suggest that Igs may be produced in cells
other than B lymphocytes. The potential role of podocyte-derived
IgG in the pathogenesis of podocytic diseases, such as membranous
nephropathy (MN), focal segmental glomerular sclerosis (FSGS), and
its possible clinical application, require further
investigation.
Acknowledgments
The authors would like to thank the Department of
Immunology, Peking University, for supporting their work.
Abbreviations:
|
RAG
|
recombination activating gene
|
|
AID
|
activation-induced cytidine
deaminase
|
|
SAC
|
Staphylococcus aureus
|
|
MN
|
membranous nephropathy
|
Notes
[1]
Funding
This study was supported by grants from the National
Natural Science Foundation of China (91642109, 91229102 and
81272237).
[2] Authors'
contributions
As the corresponding authors, YW and XQ planned and
supervized the study. ZJ participated in the research design,
performed most experiments and wrote the manuscript. HD, JM and YG,
were partly involved in immunohistochemistry assay, western blot
analysis and writing of the manuscript. YL, RW, LA and ZG provided
suggestions and technical supports. All authors read and approved
the final manuscript.
[3] Availability
of data and materials
The data and materials described in the manuscript
will be freely available to any scientist wishing to use them for
noncommercial purposes.
[4] Ethics
approval and consent to participate
The study was approved by the ethics committee of
Peking University Third Hospital (approval document issued on
February 17th, 2016; under no. 053). Human serum and PBMC, used as
positive controls in both Western blot and RT-PCR tests in the
study, were obtained from the blood of first author ZJ. Her results
of blood pressure, blood routine, urine routine, liver function,
kidney function, serum biochemical are normal. She knew the
research well and was exempted from written informed consent.
[5] Consent for
publication
Not applicable.
[6] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Reiser J and Altintas MM: Podocytes.
F1000Research F1000. Faculty Rev. 5:1142016.
|
|
2
|
Wiggins RC: The spectrum of
podocytopathies: A unifying view of glomerular diseases. Kidney
Int. 71:1205–1214. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Beck LH Jr and Salant DJ: Membranous
nephropathy: From models to man. J Clin Invest. 124:2307–2314.
2014. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Beck LH Jr, Bonegio RGB, Lambeau G, Beck
DM, Powell DW, Cummins TD, Klein JB and Salant DJ: M-type
phospholipase A2 receptor as target antigen in idiopathic
membranous nephropathy. N Engl J Med. 361:11–21. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Schlumberger W, Hornig N, Lange S, Probst
C, Komorowski L, Fechner K, Dähnrich C and Stöcker W: Differential
diagnosis of membranous nephropathy with autoantibodies to
phospholipase A2 receptor 1. Autoimmun Rev. 13:108–113. 2014.
View Article : Google Scholar
|
|
6
|
Qiu X, Zhu X, Zhang L, Mao Y, Zhang J, Hao
P, Li G, Lv P, Li Z, Sun X, et al: Human epithelial cancers secrete
immunoglobulin g with unidentified specificity to promote growth
and survival of tumor cells. Cancer Res. 63:6488–6495.
2003.PubMed/NCBI
|
|
7
|
Li M, Feng DY, Ren W, Zheng L, Zheng H,
Tang M and Cao Y: Expression of immunoglobulin kappa light chain
constant region in abnormal human cervical epithelial cells. Int J
Biochem Cell Biol. 36:2250–2257. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Babbage G, Ottensmeier CH, Blaydes J,
Stevenson FK and Sahota SS: Immunoglobulin heavy chain locus events
and expression of activation-induced cytidine deaminase in
epithelial breast cancer cell lines. Cancer Res. 66:3996–4000.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen Z and Gu J: Immunoglobulin G
expression in carcinomas and cancer cell lines. FASEB J.
21:2931–2938. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Qiu X, Sun X, He Z, Huang J, Hu F, Chen L,
Lin P, You MJ, Medeiros LJ and Yin CC: Immunoglobulin gamma heavy
chain gene with somatic hypermutation is frequently expressed in
acute myeloid leukemia. Leukemia. 27:92–99. 2013. View Article : Google Scholar
|
|
11
|
Huang J, Sun X, Gong X, He Z, Chen L, Qiu
X and Yin CC: Rearrangement and expression of the immunoglobulin
µ-chain gene in human myeloid cells. Cell Mol Immunol. 11:94–104.
2014. View Article : Google Scholar
|
|
12
|
Wang C, Xia M, Sun X, He Z, Hu F, Chen L,
Bueso-Ramos CE, Qiu X and Yin CC: IGK with conserved IGKV/IGKJ
repertoire is expressed in acute myeloid leukemia and promotes
leukemic cell migration. Oncotarget. 6:39062–39072. 2015.PubMed/NCBI
|
|
13
|
Huang J, Zhang L, Ma T, Zhang P and Qiu X:
Expression of immunoglobulin gene with classical V-(D)-J
rearrangement in mouse testis and epididymis. J Histochem Cytochem.
57:339–349. 2009. View Article : Google Scholar :
|
|
14
|
Yan M, Zhang X, Pu Q, Huang T, Xie Q, Wang
Y, Li J, Wang Y, Gu H, Huang T, et al: Immunoglobulin g expression
in human sperm and possible functional significance. Sci Rep.
6:201662016. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Huang J, Sun X, Mao Y, Zhu X, Zhang P,
Zhang L, Du J and Qiu X: Expression of immunoglobulin gene with
classical V-(D)-J rearrangement in mouse brain neurons. Int J
Biochem Cell Biol. 40:1604–1615. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Niu N, Zhang J, Guo Y, Zhao Y, Korteweg C
and Gu J: Expression and distribution of immunoglobulin G and its
receptors in the human nervous system. Int J Biochem Cell Biol.
43:556–563. 2011. View Article : Google Scholar
|
|
17
|
Zhao Y, Liu Y, Chen Z, Korteweg C and Gu
J: Immunoglobulin g (IgG) expression in human umbilical cord
endothelial cells. J Histochem Cytochem. 59:474–488. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Jiang D, Ge J, Liao Q, Ma J, Liu Y, Huang
J, Wang C, Xu W, Zheng J, Shao W, et al: IgG and IgA with potential
microbial-binding activity are expressed by normal human skin
epidermal cells. Int J Mol Sci. 16:2574–2590. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zheng J, Huang J, Mao Y, Liu S, Sun X, Zhu
X, Ma T, Zhang L, Ji J, Zhang Y, et al: Immunoglobulin gene
transcripts have distinct VHDJH recombination characteristics in
human epithelial cancer cells. J Biol Chem. 284:13610–13619. 2009.
View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Lee G and Ge B: Cancer cell expressions of
immunoglobulin heavy chains with unique carbohydrate-associated
biomarker. Cancer Biomark. 5:177–188. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Lee G and Ge B: Inhibition of in vitro
tumor cell growth by RP215 monoclonal antibody and antibodies
raised against its anti-idiotype antibodies. Cancer Immunol
Immunother. 59:1347–1356. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wen YJ, Mancino A, Pashov A, Whitehead T,
Stanley J and Kieber-Emmons T: Antigen binding of human IgG Fabs
mediate ERK-associated proliferation of human breast cancer cells.
DNA Cell Biol. 24:73–84. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Wang J, Lin D, Peng H, Huang Y, Huang J
and Gu J: Cancer-derived immunoglobulin G promotes tumor cell
growth and proliferation through inducing production of reactive
oxygen species. Cell Death Dis. 4:e9452013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li M, Zheng H, Duan Z, Liu H, Hu D, Bode
A, Dong Z and Cao Y: Promotion of cell proliferation and inhibition
of ADCC by cancerous immunoglobulin expressed in cancer cell lines.
Cell Mol Immunol. 9:54–61. 2012. View Article : Google Scholar
|
|
25
|
Liu Y, Liu D, Wang C, Liao Q, Huang J,
Jiang D, Shao W, Yin CC, Zhang Y, Lee G, et al: Binding of the
monoclonal antibody RP215 to immunoglobulin G in metastatic lung
adenocarcinomas is correlated with poor prognosis. Histopathology.
67:645–653. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Liao Q, Liu W, Liu Y, Wang F, Wang C,
Zhang J, Chu M, Jiang D, Xiao L, Shao W, et al: Aberrant high
expression of immunoglobulin G in epithelial stem/progenitor-like
cells contributes to tumor initiation and metastasis. Oncotarget.
6:40081–40094. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Liu H, Zheng H, Duan Z, Hu D, Li M, Liu S,
Li Z, Deng X, Wang Z, Tang M, et al: LMP1-augmented kappa intron
enhancer activity contributes to upregulation expression of Ig
kappa light chain via NF-kappaB and AP-1 pathways in nasopharyngeal
carcinoma cells. Mol Cancer. 8:922009. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhu X, Wu L, Zhang L, Hao P, Zhang S,
Huang J, Zheng J, Liu Y, Li W, Zhang Y, et al: Distinct regulatory
mechanism of immunoglobulin gene transcription in epithelial cancer
cells. Cell Mol Immunol. 7:279–286. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wu L, Liu Y, Zhu X, Zhang L, Chen J, Zhang
H, Hao P, Zhang S, Huang J, Zheng J, et al: The immunoglobulin
heavy chain VH6-1 promoter regulates Ig transcription in non-B
cells. Cancer Cell Int. 14:1142014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Saleem MA, O'Hare MJ, Reiser J, Coward RJ,
Inward CD, Farren T, Xing CY, Ni L, Mathieson PW and Mundel P: A
conditionally immortalized human podocyte cell line demonstrating
nephrin and podocin expression. J Am Soc Nephrol. 13:630–638.
2002.PubMed/NCBI
|
|
31
|
Cerutti A, Zan H, Schaffer A, Bergsagel L,
Harindranath N, Max EE and Casali P: CD40 ligand and appropriate
cytokines induce switching to IgG, IgA, and IgE and coordinated
germinal center and plasmacytoid phenotypic differentiation in a
human monoclonal IgM+IgD+ B cell line. J
Immunol. 160:2145–2157. 1998.PubMed/NCBI
|
|
32
|
Lynch S, Kelleher D, McManus R and
O'Farrelly C: RAG1 and RAG2 expression in human intestinal
epithelium: Evidence of extrathymic T cell differentiation. Eur J
Immunol. 25:1143–1147. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Revy P, Muto T, Levy Y, Geissmann F,
Plebani A, Sanal O, Catalan N, Forveille M, Dufourcq-Labelouse R,
Gennery A, et al: Activation-induced cytidine deaminase (AID)
deficiency causes the autosomal recessive form of the Hyper-IgM
syndrome (HIGM2). Cell. 102:565–575. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lee G, Huang C, Chow S and Chien C:
Carbohydrate-associated epitope-based anti-cancer drugs and
vaccines. Adv Biosci Biotechnol. 4:18–23. 2013. View Article : Google Scholar
|
|
35
|
Shao W, Zhang C, Liu E, Zhang L, Ma J, Zhu
Z, Gong X, Qin Z and Qiu X: Identification of liver epithelial
cell-derived Ig expression in µ chain-deficient mice. Sci Rep.
6:236692016. View Article : Google Scholar
|
|
36
|
Tonegawa S: Somatic generation of antibody
diversity. Nature. 302:575–581. 1983. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Glanville J, Zhai W, Berka J, Telman D,
Huerta G, Mehta GR, Ni I, Mei L, Sundar PD, Day GM, et al: Precise
determination of the diversity of a combinatorial antibody library
gives insight into the human immunoglobulin repertoire. Proc Natl
Acad Sci USA. 106:20216–20221. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Bassing CH, Swat W and Alt FW: The
mechanism and regulation of chromosomal V(D)J recombination. Cell.
109(Suppl 1): S45–S55. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Welsh GI and Saleem MA: The podocyte
cytoskeleton--key to a functioning glomerulus in health and
disease. Nat Rev Nephrol. 8:14–21. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nagata M: Podocyte injury and its
consequences. Kidney Int. 89:1221–1230. 2016. View Article : Google Scholar : PubMed/NCBI
|