Introduction
The skin is a primary of protective organ system and
guards against destructive stimuli, including solar ultraviolet
(UV) radiation. With ozone layer depletion through
halogen-containing compound emissions caused by human activities,
large amounts of UV radiation are able to reach to the earth, which
increases the level of UV radiation. Once absorbed by human skin,
severe skin problems, including cutaneous malignancies then arise
(1). After sun exposure, specific
pathophysiological insults, including massive elastosis, collagen
degeneration, pigmentation, sallowness and deep wrinkling, occur in
response to UV (2,3). Solar UV radiation on the surface of
the earth consists of 90-99% UVA and 1-10% UVB rays (4), which are classified as wavelength
ranges of 315-400 and 280-315 nm, respectively. While UVB radiation
constitutes 1-10% of the UV energy that reaches the surface of the
earth, it is 1,000-10,000-fold more carcinogenic than UVA
radiation, which makes up 90-99% of UV energy (5).
Excessive exposure to UV radiation predisposes
humans to unhealthy conditions, including sunburn, inflammation,
oxidative stress, DNA damage, solar erythema, premature aging,
suppression of the immune system and skin cancers (6-9).
Of note, exposure to UVB radiation induces excessive generation of
reactive oxygen species (ROS), including hydrogen peroxide,
superoxide anions, hydroxyl radicals and singlet oxygen (10), which may oxidize and damage
cellular lipids, proteins and DNA, leading to pathophysiological
changes and even skin aging (11,12). Accumulative studies have indicated
that the direct absorption of UVB photons, which underlies
UVB-induced oxidative injury, leads to the formation of oxidative
products (13,14). In addition, UVB influences the
expression of anti-oxidant enzymes, activates the anti-oxidant
defense system and ultimately interferes with the ability of the
skin to protect itself against ROS and reactive nitrogen species
(15).
Nuclear factor erythroid-2-related factor 2 (Nrf2)
is a crucial anti-oxidant signaling molecule in mammalian cells. Of
note, the Nrf2/antioxidant response element (ARE) signaling pathway
has a vital protective role in UV radiation-induced skin cell
injury, which results from the induction of DNA damage or apoptosis
(16-18). After exposure to UV, Nrf2
dissociates from Kelch-like ECH-associated protein 1 (Keap1) to
translocate into the nucleus and combine with AREs. The Nrf2/ARE
signaling pathway is thereby activated, which further activates
downstream cytoprotective genes and upregulates anti-oxidant
enzymes, including heme oxygenase-1 (HO-1), to counteract
UVB-mediated cellular injury and eventually maintain cell survival
(19). Zhong and Li (20) revealed that SKPs have a protective
role in resisting UVB-induced dermal fibroblast cell damage and
aging via upregulation of Nrf2. Furthermore, the
phosphatidylinositol 3-kinase (PI3K)/Akt pathway may also improve
cell survival and is required for Nrf2 activation (21). Activation of the Nrf2 signaling
pathway requires the involvement of PI3K phosphorylation and
downstream Akt phosphorylation (22-25). Activation of Nrf2/ARE via PI3K
signaling has a protective effect on skin cells against UV-induced
DNA damage (26).
Numerous studies have verified that hydrogen exerts
protective anti-oxidant and anti-inflammatory effects, and
effectively scavenges ROS to promote a protective system in cells
and organs. It was also revealed that hydrogen protects cells and
tissues from oxidative injury by activating Nrf2/HO-1 (27) and the Akt signaling pathway
(28,29). In addition, hydrogen was also
reported to protect the skin from psoriasis (30), dermatitis induced by local
radiation (31) and oxidative
damage induced by high glucose or mannitol (32). Previous studies by our group have
reported that hydrogen alleviates UVB-induced inflammation
(33,34) and oxidative damage (35).
Although hydrogen mitigates inflammation and
oxidative damage in skin-associated diseases, the detailed
mechanisms of the effects of hydrogen in UVB-radiated HaCaT cells
has remained elusive. The present study first assessed the effects
of hydrogen on oxidative damage in HaCaT cells irradiated with UVB
and subsequently aimed to examine the mechanistic involvement of
the regulatory effects of the PI3K/Akt/Nrf2 signaling pathway.
Materials and methods
Cell culture and UVB radiation
The human keratinocyte cell line HaCaT was purchased
from the American Type Culture Collection (Manassas, VA, USA) and
cultured in Dulbecco's modified of Eagle's medium (DMEM) containing
10% heat-inactivated fetal bovine serum (FBS; Thermo Fisher
Scientific, Inc., Waltham, MA, USA), 100 U/ml penicillin and 100
μg/ml streptomycin in a humidified atmosphere containing 5%
CO2 at 37°C. HaCaT cells were seeded 1×106
cells/ml in 6-well plates and allowed to adhere for 24 h. Cells
were exposed to UVB with an energy spectrum of UVB radiation (1, 10
or 50 mJ/cm2) with a thin layer of PBS using a UVB lamp
with a peak emission at 301 nm (280-320 nm; Sigma-Aldrich; Merck
KGaA, Darmstadt, Germany). After UVB irradiation for 30 sec, the
cells were washed with warm PBS and incubated with serum-free DMEM
for 24 h. LY294002 (30 μM; Sigma-Aldrich; Merck KGaA) was
added to the medium 2 h prior UVB irradiation. Cells in the control
group underwent the same schedule for medium changes without UVB
irradiation.
Hydrogen treatment
A previously described method was used with certain
modifications (36). In brief,
hydrogen was dissolved in DMEM supplemented with 10% FBS for 4 h
under high pressure (0.4 MPa) to reach a supersaturated level. The
saturated hydrogen-rich medium was stored with no air in the vessel
at 4°C under atmospheric pressure in an aluminum bag. Hydrogen-rich
medium was freshly prepared every week to ensure that a
concentration of 0.6 mmol/l hydrogen was maintained. A needle-type
H2 sensor (Unisense A/S, Aarhus, Denmark) was used to
measure the hydrogen concentration of the media according to the
method published by Chen et al (27).
MTT assay
Cells were seeded into a 96-well plate at a density
of 1,000 cells/well. Different treatments were completed as per the
experimental schedule. The cells were washed with PBS, the medium
was removed from each well, and the cells were incubated with 200
μl of MTT (0.5 mg/ml) for 4 h at 37°C. The formazan crystals
that had formed were dissolved with 150 μl dimethylsulfoxide
after the medium was removed. The plates were incubated on an
agitator for 15 min at room temperature. The absorbance of each
well was measured at 490 nm using a microplate reader (Bio-Rad
Laboratories, Inc., Hercules, CA, USA). The experiment was repeated
five times.
Lactate dehydrogenase (LDH)
detection
Cells were seeded at a density of 1×105
cells/well in a 12-well plate and incubated at 37°C with 5%
CO2 for 24 h. Different treatments were completed as per
the experimental schedule. Subsequently, the supernatant from each
well was collected and the cytolytic activity was measured with an
LDH cytotoxicity detection kit (Nanjing Jiancheng, Nanjing, China).
The LDH levels were expressed as percentages of the LDH levels
observed in the control cultures.
Intracellular ROS detection
Intracellular ROS levels were measured with a
dichlorodihydrofluorescein diacetate (DCFH-DA) assay. HaCaT cells,
at a density of 1×104 cells/well, were seeded in a
96-well plate, cultured for 6 h and treated according to the
indicated experimental schedules for 24 h. A total of 25 μM
DCFH-DA was added to the medium, followed by incubation at 37°C for
30 min in the dark. The fluorescence of the
2′,7′-dichlorofluorescein product was detected with a PerkinElmer
LS-5B fluorescence spectrometer (PerkinElmer, Waltham, MA, USA).
The amounts of ROS were expressed as percentages of the ROS
observed in the control cultures. All experiments for the standards
and samples were performed in duplicate.
8-iso-prostaglandin F2α (8-iso PGF2α) and
malondialdehyde (MDA) detection
The culture media were collected and centrifuged at
1,000 × g for 10 min at 4°C, and the supernatants were collected
for detection. To estimate oxidative stress, 8-iso-prostaglandin
F2α (8-iso PGF2α) was measured with a kit (cat. no. 316350; Cayman
Chemical Company, Ann Arbor, MI, USA) using a microplate reader (CA
94089; Molecular Devices, Sunnyvale, CA, USA) according to the
manufacturer's recommendations. For MDA detection, supernatants
were processed using the kit (cat. no. 10009202-1; Cayman Chemical
Company) as per the manufacturer's protocols. All experiments for
the standards and samples were performed in duplicate.
Western blot analysis
After the incubations were completed, the cells were
lysed with lysis buffer on ice for 10 min in
radioimmunoprecipitation assay lysis buffer, and the protein was
thereby extracted. The lysates were then collected and the protein
concentration was evaluated via a Bicinchoninic Acid protein assay
kit (Thermo Fisher Scientific, Inc.). The concentration of all
lysates was adjusted to be the same according to the results of the
BCA assay. Proteins (30 μg per lane) were separated by 12%
SDS-PAGE and transferred onto polyvinylidene fluoride membranes
(Immobilon®-P Transfer Membrane; EMD Millipore,
Billerica, MA, USA). Nuclear extracts for Nrf2 immunoblotting were
prepared with the NE-PER nuclear kit (Pierce; Thermo Fisher
Scientific, Inc.). The membranes were blocked with 5% non-fat milk
in Tris-HCL-based buffered saline with 0.1% Tween-20
(Sigma-Aldrich; Merck KGaA) at pH 7.4 for 1 h and incubated with
the following primary antibodies overnight at 4°C: β-actin
(1:2,000; cat. no. ab8226), Nrf2 (1:1,000; cat. no. ab62352), HO-1
(1:1,000; cat. no. ab13243), PI3K (1:1,000; cat. no. ab182651),
phosphorylated (p)-PI3K (1:500 dilution; cat. no. ab182651), Akt
(1:500 dilution; cat. no. ab18785) and p-Akt (1:500; cat. no.
ab38449) (Abcam, Cambridge, MA, USA). The membranes were then
incubated with corresponding horseradish peroxidase-conjugated
secondary antibodies (anti-mouse, sc-2005; 1:5,000; Santa Cruz
Biotechnology, Inc., Dallas, TX, USA and anti-rabbit, sc-2357,
1:5,000; Santa Cruz Biotechnology, Inc.). Protein bands were
detected with a ChemiDoc™ (Bio-Rad Laboratories, Inc., Hercules,
CA, USA). β-actin expression was used as the reference. The protein
expression level was quantified with Quantity One version 4.5.2
software (Bio-Rad Laboratories, Inc.)
Statistical analysis
Values are expressed as the mean ± standard error of
the mean. Multigroup comparisons were evaluated using one-way
analysis of variance followed by Tukey's post hoc test using
GraphPad Prism version 5 software (GraphPad Inc., La Jolla, CA,
USA). P<0.05 was considered to indicate a statistically
significant difference.
Results
Effects of different doses of UVB
radiation on LDH release and cell viability
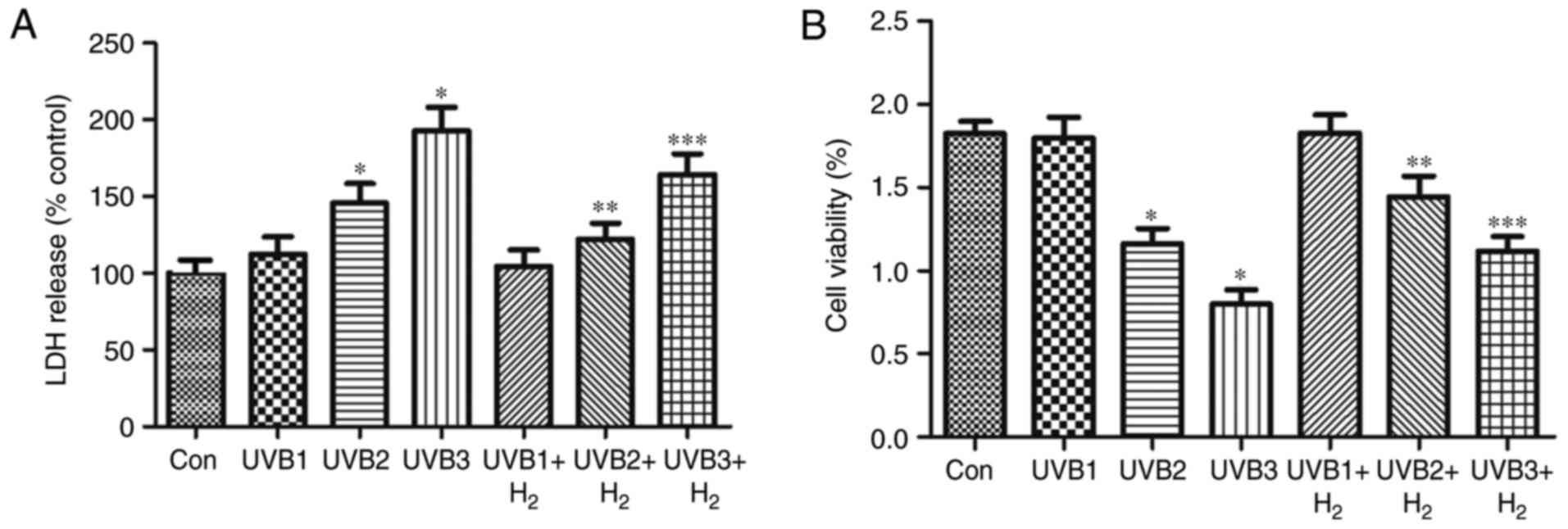
UVB radiation disrupted the cells, decreased cell
viability and increased LDH release. The LDH leakage assay is based
on the measurement of LDH activity in the extracellular medium. In
the present study, various doses of UVB radiation were used to
imitate the effects of UVB on human keratinocytes. Irradiation of
HaCaT cells with 1, 10 and 50 mJ/cm2 UVB had different
effects on LDH release and inhibited cell viability, but only 10
and 50 mJ/cm2 UVB had significant effects compared with
the control group (P<0.05; Fig. 1A
and B). Furthermore, it was investigated whether hydrogen
exerts a protective effect on HaCaT cells irradiated by UVB.
Hydrogen treatment reduced the LDH release of cells after exposure
to 1, 10 and 50 mJ/cm2 UVB, which was significant at 10
and 50 mJ/cm2 UVB (P<0.05; Fig. 1A). The results of the cell
viability assay were similar to those of the LDH assay, namely that
10 and 50 mJ/cm2 UVB significantly reduced the cell
viability compared with that in the control group, while hydrogen
significantly promoted the cell viability after exposure to 10 and
50 mJ/cm2 UVB (P<0.05; Fig. 1B).
Hydrogen treatment alleviates ROS, 8-iso
PGF2α and MDA release in UVB-induced HaCaT cells
UV radiation is associated with the formation of
ROS, leading to skin aging and photocarcinogenesis. To determine
whether hydrogen alleviated UVB-mediated ROS accumulation and
oxidative products, ROS, 8-iso PGF2α and MDA release levels caused
by the augmentation of the endogenous anti-oxidant capacity was
measured in UVB-induced HaCaT cells. The UVB-induced intracellular
oxidative stress level was measured with a redox-sensitive dye,
DCFH-DA. The results indicated that UVB radiation stimulated ROS
production in a dose-dependent manner (Fig. 2A). Of note, hydrogen treatment
obviously decreased the levels of ROS in UVB-irradiated cells
(P<0.05; Fig. 2A). Similarly,
8-iso PGF2α and MDA were also induced by UVB radiation in a
dose-dependent manner compared with the control group; in addition,
hydrogen significantly reduced the excessive 8-iso PGF2α and MDA
release after UV irradiation (P<0.05; Fig. 2B and C). These results indicated
that hydrogen alleviated ROS and their oxidative products induced
by UVB irradiation.
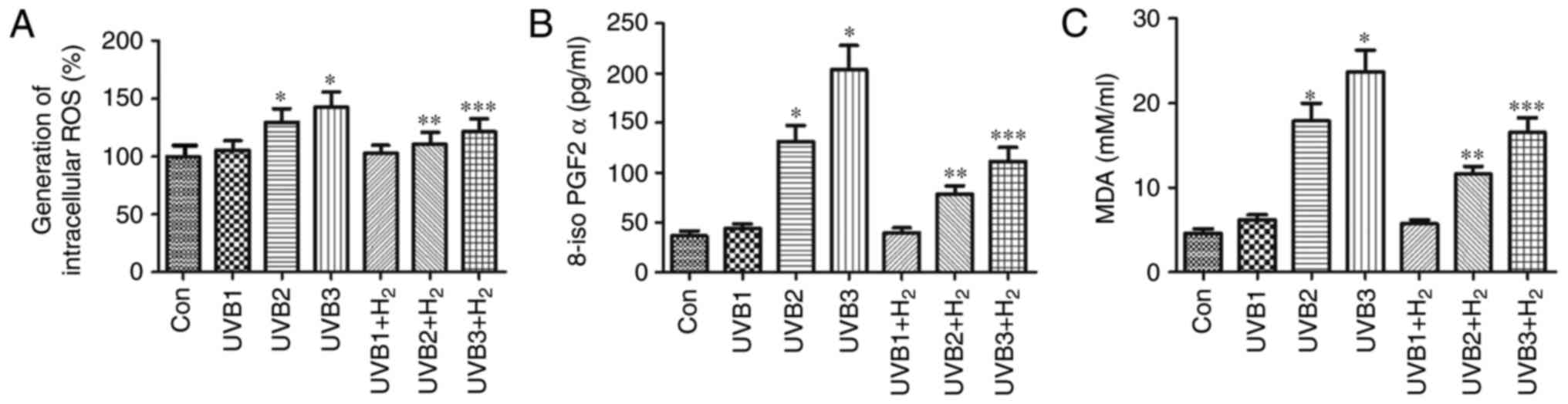 | Figure 2Effect of hydrogen on (A) ROS, (B)
8-iso PGF2α and (C) MDA induced by various doses of UVB radiation.
HaCaT cells were irradiated with UVB at 1, 10 and 50
mJ/cm2 for 30 sec. In groups UVB1+H2,
UVB2+H2 and UVB3+H2, normal medium was
replaced with hydrogen-rich medium, and cells were further cultured
for 24 h. Supernatant was collected for detection of ROS and 8-iso
PGF2α and MDA. Values are expressed as the mean ± standard error of
the mean (n=6). *P<0.05 compared with Con group;
**P<0.05 compared with UVB2 group;
***P<0.05 compared with UVB3 group. UVB1/2/3, UVB at
1, 10 and 50 mJ/cm2. UVB, ultraviolet B; Con, control;
ROS, reactive oxygen species; MDA, malondialdehyde; 8-iso PGF2α,
8-iso-prostaglandin F2α. |
Hydrogen increases the protein levels of
p-PI3K, p-Akt and Nrf2 in UVB-irradiated HaCaT cells
Hydrogen reduced ROS and oxidation product levels in
UVB-irradiated HaCaT cells. The Nrf2 pathway is an endogenous
anti-oxidant system that protects cells from oxidative injury. The
present study assessed whether hydrogen was able to activate the
Nrf2 pathway to reduce ROS and oxidative products. The expression
levels of Nrf2 and its downstream protein HO-1 were determined to
confirm the role of Nrf2 in hydrogen-mediated protection (Fig. 3). UVB radiation increased Nrf2
expression in the nucleus in a concentration-dependent manner,
while cytoplasmic Nrf2 expression was reduced (P<0.05; Fig. 3A, C and D). In addition, the
changes in HO-1 protein expression in the cytoplasm were similar
with Nrf2 expression in the nucleus after UVB-radiation (P<0.05;
Fig. 3A and B). Hydrogen
treatment clearly increased nuclear Nrf2 expression and HO-1
expression after UVB irradiation (P<0.05; Fig. 3A-C).
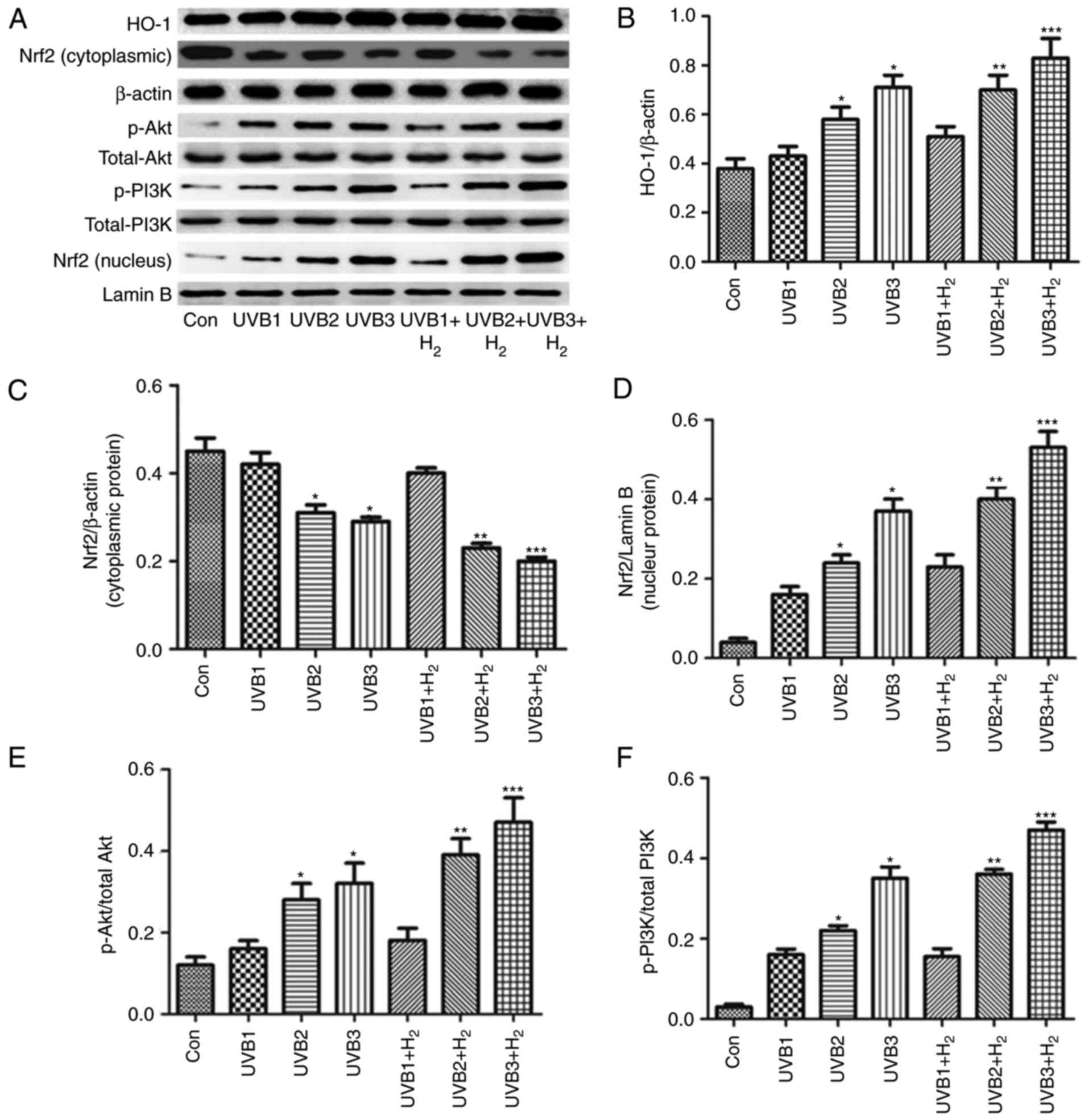 | Figure 3Nrf2, HO-1, Akt and PI3K expression
after hydrogen and UVB treatment of HaCaT cells. HaCaT cells were
irradiated with UVB at 1, 10 and 50 mJ/cm2 for 30 sec.
In groups UVB1+H2, UVB2+H2 and
UVB3+H2, normal medium was replaced with hydrogen-rich
medium, and cells were further cultured for 24 h. Cells were
collected and subjected to western blot analysis. (A)
Representative western blot image. (B-F) Quantified expression
levels of (B) HO-1, (C) cytosolic Nrf2, (D) nuclear Nrf2, (E)
p/total Akt and (F) p/total PI3K. Values are expressed as the mean
± standard error of the mean (n=6). *P<0.05 compared
with Con group; **P<0.05 compared with UVB2 group;
***P<0.05 compared with UVB3 group. UVB, ultraviolet
B; Con, control; p-PI3K, phosphorylated phosphoinositide-3 kinase;
HO-1, heme oxygenase 1; Nrf2, nuclear factor erythroid-2-related
factor 2. |
PI3K is the upstream factor of the Nrf2 signaling
pathway (37). To determine the
effect of PI3K on the hydrogen-activated Nrf2 pathway in
UVB-irradiated HaCaT cells, the protein levels of PI3K and its
downstream effector, Akt, were measured by western blot analysis
(Fig. 3). It was indicated that
the p-Akt and p-PI3K levels were increased by UVB radiation
compared with those in the control group (P<0.05; Fig. 3A, E and F). Hydrogen further
increased the p-Akt and p-PI3K levels in UVB-irradiated cells
(P<0.05; Fig. 3A, E and F).
These results indicated that hydrogen increased the expression of
PI3K/Akt and Nrf2/HO-1 pathway modulators.
Hydrogen exerts its protective effects
against UV-induced cell damage through the PI3K/Akt pathway
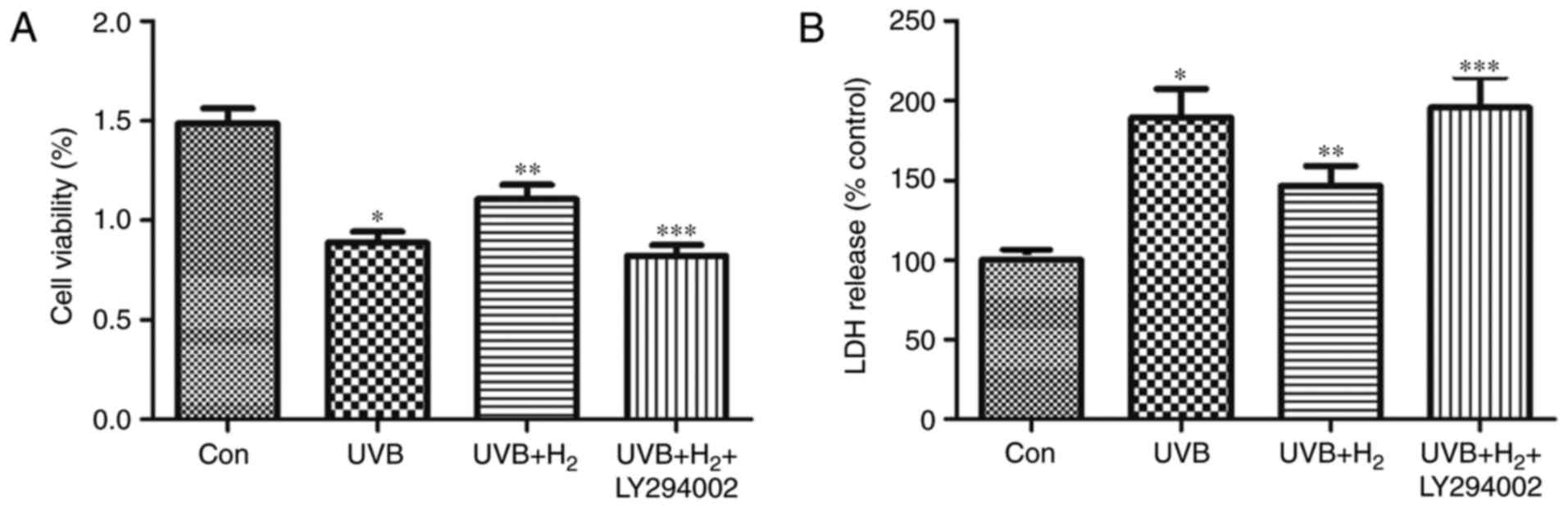
To confirm that the effect of hydrogen on LDH
release and cell viability was mediated through the PI3K/Akt
pathway, cells were treated with a PI3K-specific inhibitor,
LY294002, and its impact on the effects of hydrogen on LDH release
and viability of irradiated cells was measured. Compared with the
UVB group, the cell the viability increased and LDH release
decreased in the UVB+H2 group; however, following PI3K
inhibition, it appeared that the protective roles exhibited by
hydrogen on cell viability and damage were reduced (P<0.05;
Fig. 4A and B). These results
suggested that the protective effect of hydrogen was mediated by
the activation of the PI3K/Akt pathway.
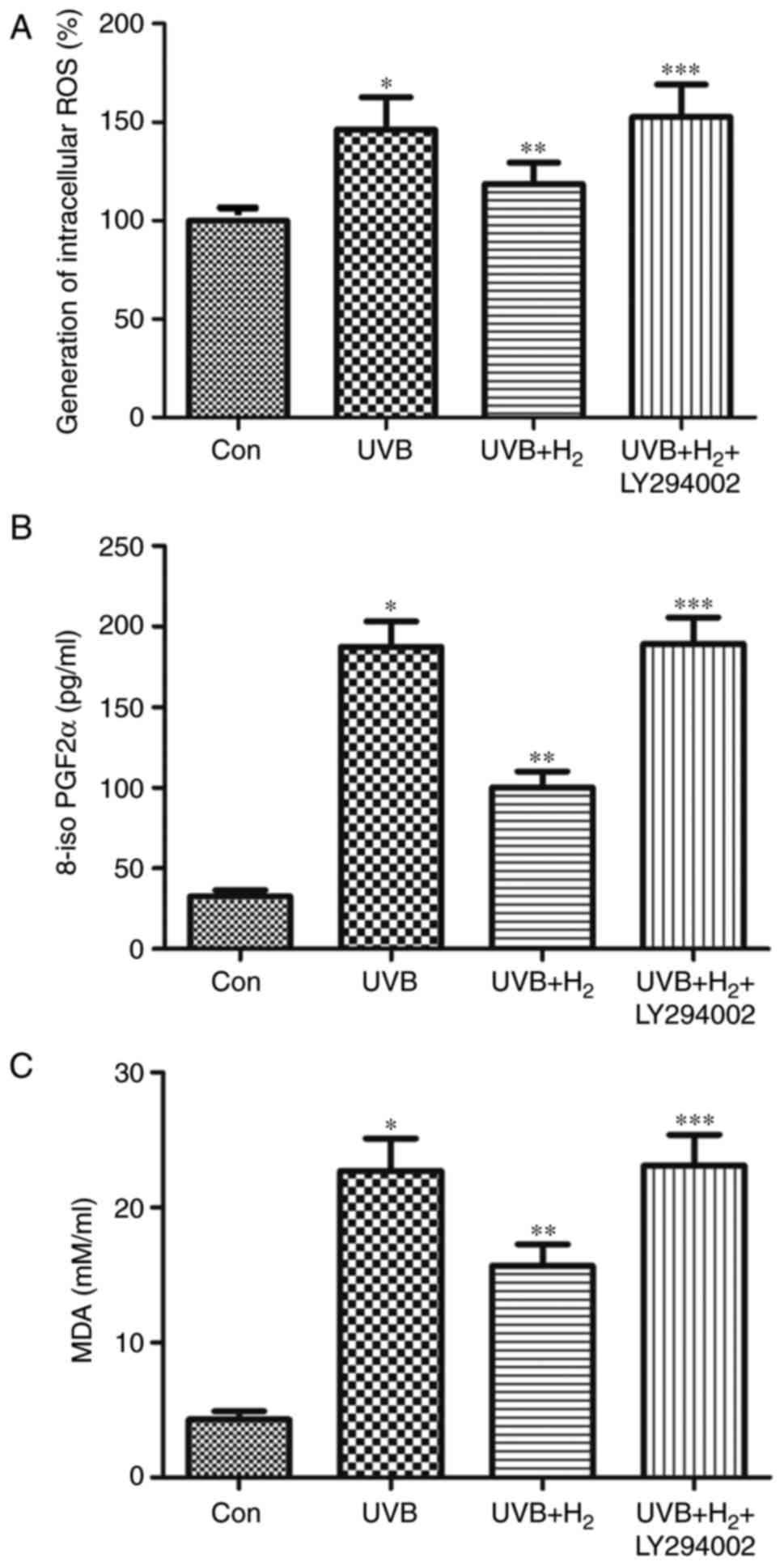
Hydrogen exerts its anti-oxidative
effects via the PI3K/Akt pathway in UVB-irradiated HaCaT cells
The present study further investigated whether the
effects of hydrogen to decrease ROS and its oxidative products
occurred via PI3K/Akt. Compared with the UVB group, ROS, 8-iso
PGF2α and MDA release were reduced in the UVB+H2 group,
but LY294002 abolished the anti-oxidant effect of hydrogen
(P<0.05; Fig. 5A-C). These
results indicated that hydrogen exerted its anti-oxidative effects
in UVB-irradiated HaCaT cells through the activation of the
PI3K/Akt pathway.
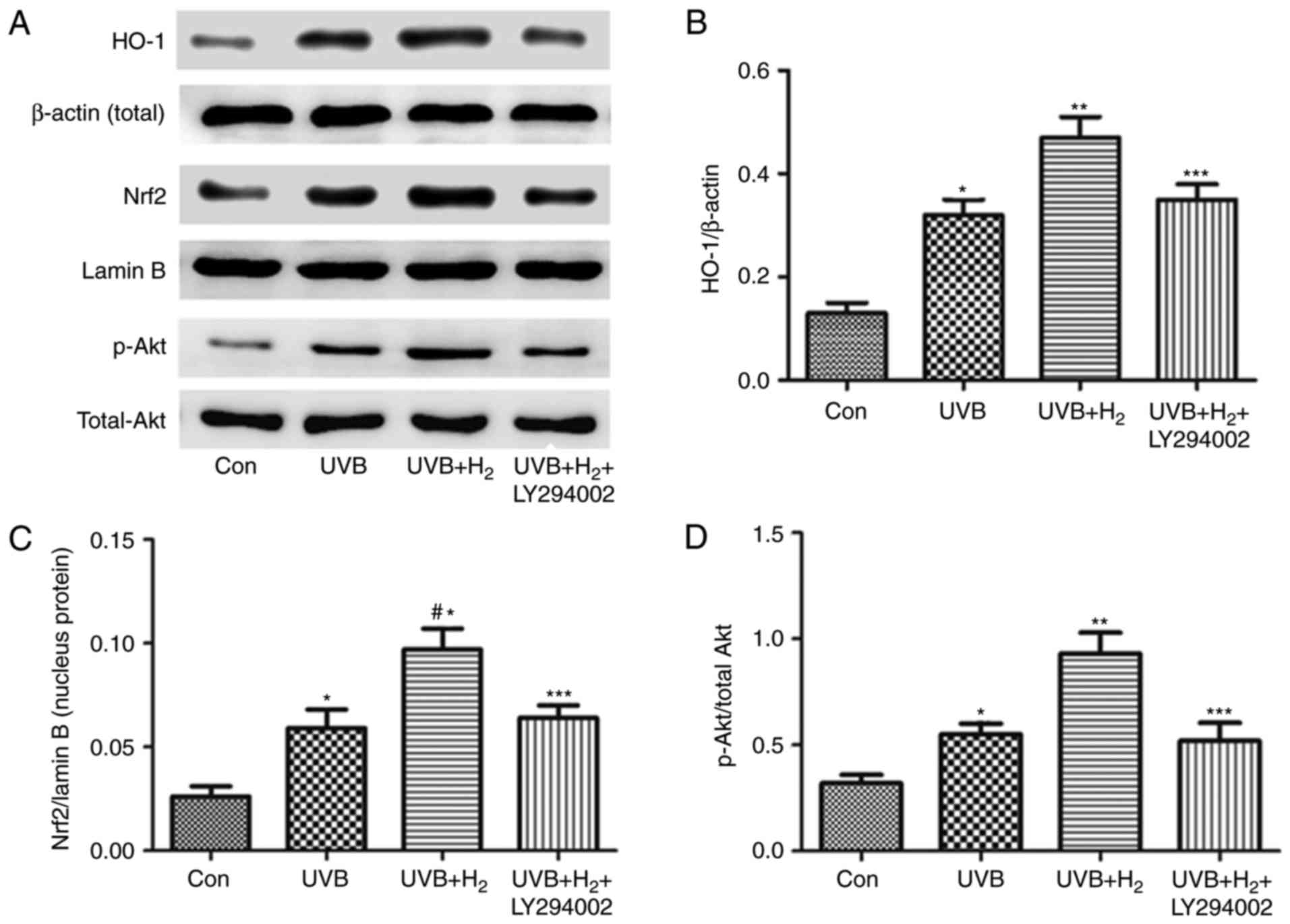
Hydrogen activates Nrf2 expression
through the PI3K/Akt pathway
To explore whether hydrogen stimulates the increase
in Nrf2 protein via PI3K/Akt, the expression of the upstream
proteins PI3K and Akt had been assessed (Fig. 3); furthermore, LY294002 was
applied to assess the involvement of the PI3K/Akt pathway in the
effects of hydrogen on the Nrf2 pathway. The results indicated that
LY294002 inhibited Nrf2 translocation into the nucleus and reduced
HO-1 and p-Akt levels in the UVB+H2+LY294003 group
compared with those in the UVB+H2 group (P<0.05;
Fig. 6A–D).
Discussion
As a potent environmental risk factors, exposure to
UV irradiation is involved in the pathogenesis of several skin
diseases (38). Of note, UVB
primarily acts on the epidermal basal cell layer, whereas UVB may
directly cause pathological changes of the skin. The present study
focused on the effects of hydrogen, a newly identified gaseous
signaling molecule, on HaCaT cells treated with UVB radiation. The
results indicated that UVB radiation increased cell toxicity,
reduced HaCaT cell proliferation, exacerbated oxidative stress
injury by increasing the levels of ROS, 8-iso PGF2α and MDA, and
upregulated the protein levels of Nrf2, HO-1, p-PI3K and p-Akt.
However, hydrogen reduced the UVB radiation-induced cytotoxicity,
inhibition of cell proliferation, and release of ROS, 8-iso PGF2α
and MDA in HaCaT cells, and further increased the levels of Nrf2,
HO-1, p-PI3K and p-Akt, which were all upregulated via the PI3K-Akt
signaling pathway.
The depletion of the ozone layer markedly aggravates
the intensity of UVB radiation on the surface of the earth.
Although UVB has certain beneficial functions, including its
involvement in vitamin D synthesis, excessive exposure to UVB
radiation causes various undesirable effects, including premature
aging, solar erythema, cell damage and even skin cancer (39). Excessive generation of ROS from
exposure of the skin to UVB may not be compensated by the
anti-oxidant defense capacity of the skin, which leads to oxidative
stress and oxidative photodamage to the skin (40,41). Imbalances between the production
and scavenging of ROS may stimulate the secretion of 8-iso PGF from
HaCaT cells, which is a specific index of lipid peroxidation
(42). As an end product of
peroxidation, MDA damages cell membranes and indirectly indicates
the extent of oxidative stress (43). Therefore, these biomarkers were
assessed in the present study to reflect oxidative stress in HaCaT
cells, and the results indicated that UVB radiation induced
oxidative stress injury in HaCaT cells with increased levels of
8-iso PGF and MDA.
However, hydrogen, an anti-oxidant gas, was reported
to inhibit oxidative processes associated with numerous diseases.
For instance, in 2007, Ohsawa et al (44) indicated that hydrogen reduced the
oxidative stress damage induced by focal ischemia and reperfusion
of the brain by selectively reducing hydroxyl radicals and
peroxynitrite (another cytotoxic ROS) in vitro. Also,
hydrogen attenuated acute kidney injury induced by pacreatitis
through inhibition of ROS in patients (45). In the present study, to verify
whether hydrogen was able to protect HaCaT cells from UVB
radiation-induced oxidative stress injury, cells were treated with
hydrogen-saturated medium, which was revealed to obviously
ameliorate cell toxicity and inhibition of cell proliferation, and
reduce the production of ROS, 8-iso PGF and MDA induced by UVB
radiation. These results indicated that hydrogen has a protective
effect on HaCaT cells exposed to UVB radiation.
Although ROS were initially described as a
disruptive factor, it is now known to be involved in regulating
cell signaling, immune functions and gene transcription (46). ROS are associated with signaling
that may activate transcription factor Nrf2 (47). In addition, as a master regulator
of anti-oxidant enzymes and cellular stress resistance, Nrf2 is
associated with UV-induced skin cell damage (20,48-51). After cell damage through exposure
to UVB, Nrf2 translocates from the cytoplasm into the nucleus to
combine with AREs, activate downstream cytoprotective genes and
upregulate the antioxidant enzyme HO-1 to counteract UVB-mediated
cellular injury (19). In
addition, the present study indicated that 1-50 mJ/cm2
UVB stimulated HaCaT cells to produce excessive amounts of ROS.
Subsequently, Nrf2 was presumably separated from Keap1 and released
into the cytoplasm, after which Nrf2 translocated into the nucleus
and activated the expression of the downstream HO-1 gene, resulting
in the protection of HaCaT cells from the harms induced by
excessive UVB radiation. These results were consistent with those
of a previous study (52). Of
note, the therapeutic effects of hydrogen in alleviating oxidative
stress in numerous diseases, including dermatitis,
neurodegenerative diseases and ischemia-reperfusion, are mediated
via several important signaling pathways (53). The protective effect of hydrogen
has been associated with the Nrf2 signaling pathway in different
diseases models (54-56). A previous study by our group
indicated that molecular hydrogen protected mice from polymicrobial
sepsis via the Nrf2/HO-1 signaling pathway (27). The present study indicated that
hydrogen stimulated UVB radiation-induced Nrf2 translocation and
HO-1 expression to further protect cells against oxidative stress
injury.
Furthermore, Nguyen et al (57) demonstrated that the activation of
Nrf2-mediated HO-1 production via the PI3K/Akt and/or protein
kinase C pathways has a critical role in the protective effect of
caffeoylserotonin against oxidative stress-induced keratinocyte
cell death. Of note, PI3K/Akt signaling is important for boosting
cell proliferation and migration in various diseases and cancer
types (58-61). In addition, ROS not only activated
the Nrf2 pathway directly to amplify its downstream signaling, but
also positively regulated PI3K activation and expression of its
target genes, which was activated through autophosphorylation
(62). Once PI3K is
phosphorylated, it activates Akt, which is a key downstream
molecule of PI3K. The PI3K/Akt signaling pathway is associated with
the Nrf2 pathway in human keratinocytes (57,63,64). The PI3K/Akt-mediated Nrf2/HO-1
signaling pathway was also reported to have a critical role in the
7,8-dihydroxyflavone-mediated protection of human keratinocytes
from oxidative stress-induced cell damage (63). In addition, Li et al
(65) demonstrated that hydrogen
ameliorated liver ischemia/reperfusion injury, which featured
marked oxidative stress injury, through inhibition of the PI3K/Akt
signaling pathway. Collectively, these aboveme ntioned studies
prompted us to hypothesize that PI3K/Akt signaling is involved in
the protective effects of hydrogen on HaCaT cells that were exposed
to excessive UVB radiation. An inhibitor of PI3K was then employed
to confirm this notion. The results of the present study indicated
that UVB radiation stimulated the phosphorylation of PI3K and Akt
in HaCaT cells, while the hydrogen treatment obviously further
increased the p-PI3K and p-Akt levels after UVB irradiation.
Furthermore, the specific PI3K inhibitor LY294002 partially
reversed the effect of hydrogen in attenuating the cytotoxicity,
inhibition of cell proliferation and production of ROS, 8-iso PGF2α
and MDA stimulated by UVB. The results of the present study
indicated that hydrogen may serve a role in amplifying the
protective response to UVB-induced damage, which may be inhibited
by LY294002. Hydrogen may have amplified the effects of irradiation
on the expression PI3K in response to radiation; however, it may be
difficult to distinguish between the effects of hydrogen and the
cellular response to radiation. When PI3K was inhibited, Nrf2
translocation to eliminate ROS was suppressed, so that ROS
generation accumulated to decrease the cellular function and
proliferation. Thus, a negative feedback loop was then formed.
These results supported the conclusion that hydrogen exerted
protective effects against UVB-induced cytotoxicity and reduction
of proliferation of HaCaT cells by quenching excessive ROS, leading
to the inhibition of the production of 8-iso PGF2α and MDA, as well
as by promotion of nuclear translocation of Nrf2 and HO-1
expression through the PI3K-Akt signaling pathway.
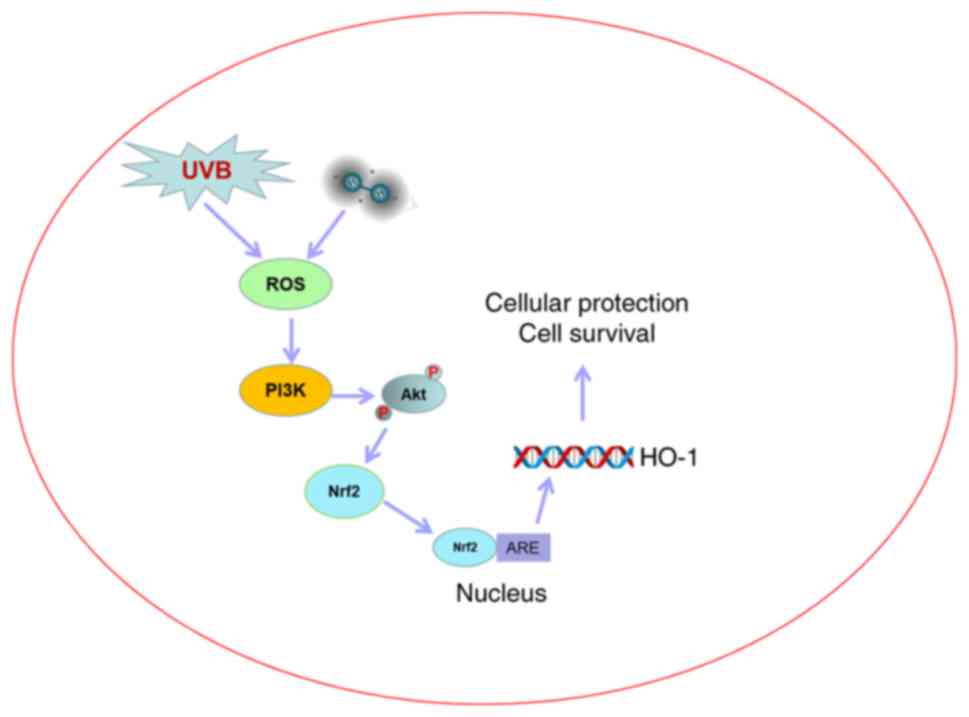
In conclusion, the present study provided evidence
that UVB radiation has cytotoxic effects and inhibits cell
proliferation, and stimulates ROS production and the generation of
their oxidation products, and that PIK3/Akt and Nrf2/HO-1
contribute to the oxidative stress response of HaCaT cells induced
by UVB radiation. Furthermore, hydrogen exerts a protective effect
by increasing Nrf2/HO-1 levels via the PI3K-Akt signaling pathway
in UVB-irradiated HaCaT cells (Fig.
7). Although PI3K/Akt was demonstrated to be involved in the
protective effect of hydrogen in UVB-induced oxidative stress
injury, further investigations of the specific mechanisms are
required for future applications of hydrogen in the clinic.
Acknowledgments
Not applicable.
Notes
[1]
Funding
The present study was supported by the grants from
the National Natural Science Foundation of China (grant no.
81601667).
[2] Availability
of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
[3] Authors'
contributions
ZBX and ZZS designed the research. ZBX, ZZS and MXY
conducted the experimental protocols in the present study. CHG
acquired analytical reagents and tools. Data analysis was performed
by CHG. ZBX and ZZS prepared the manuscript.
[4] Ethics
approval and consent to participate
Not applicable.
[5] Consent for
publication
Not applicable.
[6] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Sharma SD and Katiyar SK: Dietary grape
seed proanthocyanidins inhibit UVB-induced cyclooxygenase-2
expression and other inflammatory mediators in UVB-exposed skin and
skin tumors of SKH-1 hairless mice. Pharm Res. 27:1092–1102. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Helfrich YR, Sachs DL and Voorhees JJ:
Overview of skin aging and photoaging. Dermatol Nurs. 20:177–183;
quiz 184. 2008.PubMed/NCBI
|
|
3
|
Bernstein EF, Chen YQ, Tamai K, Shepley
KJ, Resnik KS, Zhang H, Tuan R, Mauviel A and Uitto J: Enhanced
elastin and fibrillin gene expression in chronically photodamaged
skin. J Invest Dermatol. 103:182–186. 1994. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Avila Acevedo JG, Espinosa González AM, De
Maria y Campos DM, Benitez Flores Jdel C, Hernández Delgado T,
Flores Maya S, Campos Contreras J, Muñoz López JL and García Bores
AM: Photoprotection of Buddleja cordata extract against UVB-induced
skin damage in SKH-1 hairless mice. BMC Complement Altern Med.
14:2812014. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
de Gruijl FR: Photocarcinogenesis: UVA vs
UVB. Methods Enzymol. 319:359–366. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Katiyar SK: Silymarin and skin cancer
prevention: Anti- inflammatory, antioxidant and immunomodulatory
effects (Review). Int J Oncol. 26:169–176. 2005.
|
|
7
|
Sime S and Reeve VE: Protection from
inflammation, immunosuppression and carcinogenesis induced by UV
radiation in mice by topical Pycnogenol. Photochem Photobiol.
79:193–198. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Narayanan DL, Saladi RN and Fox JL:
Ultraviolet radiation and skin cancer. Int J Dermatol. 49:978–986.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lee SJ and Park JW: Enhancement of UVB
radiation-mediated apoptosis by knockdown of cytosolic
NADP+-dependent isocitrate dehydrogenase in HaCaT cells. BMB Rep.
47:209–214. 2014. View Article : Google Scholar :
|
|
10
|
Lee KO, Kim SN and Kim YC: Anti-wrinkle
effects of water extracts of teas in hairless mouse. Toxicol Res.
30:283–289. 2014. View Article : Google Scholar
|
|
11
|
Palmer DM and Kitchin JS: Oxidative
damage, skin aging, antioxidants and a novel antioxidant rating
system. J Drugs Dermatol. 9:11–15. 2010.PubMed/NCBI
|
|
12
|
Sklar LR, Almutawa F, Lim HW and Hamzavi
I: Effects of ultraviolet radiation, visible light, and infrared
radiation on erythema and pigmentation: A review. Photochem
Photobiol Sci. 12:54–64. 2013. View Article : Google Scholar
|
|
13
|
Vayalil PK, Mittal A, Hara Y, Elmets CA
and Katiyar SK: Green tea polyphenols prevent ultraviolet
light-induced oxidative damage and matrix metalloproteinases
expression in mouse skin. J Invest Dermatol. 122:1480–1487. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Verschooten L, Claerhout S, Van Laethem A,
Agostinis P and Garmyn M: New strategies of photoprotection.
Photochem Photobiol. 82:1016–1023. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Piao MJ, Kang KA, Kim KC, Chae S, Kim GO,
Shin T, Kim HS and Hyun JW: Diphlorethohydroxycarmalol attenuated
cell damage against UVB radiation via enhancing antioxidant effects
and absorbing UVB ray in human HaCaT keratinocytes. Environ Toxicol
Pharmacol. 36:680–688. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Tao S, Justiniano R, Zhang DD and Wondrak
GT: The Nrf2-inducers tanshinone I and dihydrotanshinone protect
human skin cells and reconstructed human skin against solar
simulated UV. Redox Biol. 1:532–541. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Schäfer M, Dütsch S, auf dem Keller U,
Navid F, Schwarz A, Johnson DA, Johnson JA and Werner S: Nrf2
establishes a glutathione-mediated gradient of UVB cytoprotection
in the epidermis. Genes Dev. 24:1045–1058. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Saw CL, Huang MT, Liu Y, Khor TO, Conney
AH and Kong AN: Impact of Nrf2 on UVB-induced skin
inflammation/photo-protection and photoprotective effect of
sulforaphane. Mol Carcinog. 50:479–486. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Kannan S and Jaiswal AK: Low and high dose
UVB regulation of transcription factor NF-E2-related factor 2.
Cancer Res. 66:8421–8429. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Zhong J and Li L: Skin-derived precursors
against UVB-induced apoptosis via Bcl-2 and Nrf2 upregulation.
Biomed Res Int. 2016.6894743:2016.
|
|
21
|
Wang L, Chen Y, Sternberg P and Cai J:
Essential roles of the PI3 kinase/Akt pathway in regulating
Nrf2-dependent antioxidant functions in the RPE. Invest Ophthalmol
Vis Sci. 49:1671–1678. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Lee YJ, Jeong HY, Kim YB, Lee YJ, Won SY,
Shim JH, Cho MK, Nam HS and Lee SH: Reactive oxygen species and
PI3K/Akt signaling play key roles in the induction of Nrf2-driven
heme oxygenase-1 expression in sulforaphane-treated human
mesothelioma MSTO-211H cells. Food Chem Toxicol. 50:116–123. 2012.
View Article : Google Scholar
|
|
23
|
Xu Y, Duan C, Kuang Z, Hao Y, Jeffries JL
and Lau GW: Pseudomonas aeruginosa pyocyanin activates
NRF2-ARE-mediated transcriptional response via the
ROS-EGFR-PI3K-AKT/MEK-ERK MAP kinase signaling in pulmonary
epithelial cells. PLoS One. 8:e725282013. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Li KR, Yang SQ, Gong YQ, Yang H, Li XM,
Zhao YX, Yao J, Jiang Q and Cao C: 3H-1,2-dithiole-3-thione
protects retinal pigment epithelium cells against Ultra-violet
radiation via activation of Akt-mTORC1-dependent Nrf2-HO-1
signaling. Sci Rep. 6:255252016. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Zhang H, Liu YY, Jiang Q, Li KR, Zhao YX,
Cao C and Yao J: Salvianolic acid A protects RPE cells against
oxidative stress through activation of Nrf2/HO-1 signaling. Free
Radic Biol Med. 69:219–228. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Rodriguez KJ, Wong HK, Oddos T, Southall
M, Frei B and Kaur S: A purified feverfew extract protects from
oxidative damage by inducing DNA repair in skin cells via a
PI3-kinase-dependent Nrf2/ARE pathway. J Dermatol Sci. 72:304–310.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Chen H, Xie K, Han H, Li Y, Liu L, Yang T
and Yu Y: Molecular hydrogen protects mice against polymicrobial
sepsis by ameliorating endothelial dysfunction via an Nrf2/HO-1
signaling pathway. Int Immunopharmacol. 28:643–654. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Li DZ, Zhang QX, Dong XX, Li HD and Ma X:
Treatment with hydrogen molecules prevents RANKL-induced osteoclast
differentiation associated with inhibition of ROS formation and
inactivation of MAPK, AKT and NF-kappa B pathways in murine
RAW264.7 cells. J Bone Miner Metab. 32:494–504. 2014. View Article : Google Scholar
|
|
29
|
Hong Y, Shao A, Wang J, Chen S, Wu H,
McBride DW, Wu Q, Sun X and Zhang J: Neuroprotective effect of
hydrogen-rich saline against neurologic damage and apoptosis in
early brain injury following subarachnoid hemorrhage: Possible role
of the Akt/GSK3beta signaling pathway. PLoS One. 9:e962122014.
View Article : Google Scholar
|
|
30
|
Yu P, Wang Z, Sun X, Chen X, Zeng S, Chen
L and Li S: Hydrogen-rich medium protects human skin fibroblasts
from high glucose or mannitol induced oxidative damage. Biochem
Biophys Res Commun. 409:350–355. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mei K, Zhao S, Qian L, Li B, Ni J and Cai
J: Hydrogen protects rats from dermatitis caused by local
radiation. J Dermatolog Treat. 25:182–188. 2014. View Article : Google Scholar
|
|
32
|
Ishibashi T, Ichikawa M, Sato B, Shibata
S, Hara Y, Naritomi Y, Okazaki K, Nakashima Y, Iwamoto Y, Koyanagi
S, et al: Improvement of psoriasis-associated arthritis and skin
lesions by treatment with molecular hydrogen: A report of three
cases. Mol Med Rep. 12:2757–2764. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Meng CY: Effect of hydrogen-rich saline on
allergic contact dermatitis in mice. Chin J Dermatol. 46:275–278.
2013.
|
|
34
|
Zhang BX: Hydrogen-rich liquid
down-regulates the expressions of inflammatory facmrs by
ultraviolet B-induced human HaCaT keratinocytes through the
autophagy pathway. Chin J Dermatol. 49:123–127. 2016.In
Chinese.
|
|
35
|
Xing WB, Fu GJ, Ye WJ, Qin LY, Chen HG,
Meng XY and Meng CY: Effects of hydrogen on ultraviolet B-induced
oxidative damage to skin fibroblasts. Chin J Dermatol. 46:424–426.
2013.
|
|
36
|
Chen HG, Xie KL, Han HZ, Wang WN, Liu DQ,
Wang GL and Yu YH: Heme oxygenase-1 mediates the anti-inflammatory
effect of molecular hydrogen in LPS-stimulated RAW 264.7
macrophages. Int J Surg. 11:1060–1066. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Tongqiang L, Shaopeng L, Xiaofang Y, Nana
S, Xialian X, Jiachang H, Ting Z and Xiaoqiang D: Salvianolic acid
B prevents iodinated contrast media-induced acute renal injury in
rats via the PI3K/Akt/Nrf2 pathway. Oxid Med Cell Longev.
2016:70794872016. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Song JL and Gao Y: Protective effects of
lindera coreana on UVB-induced oxidative stress in human HaCaT
keratinocytes. Iran J Pharm Res. 13:1369–1378. 2014.
|
|
39
|
Drouin R and Therrien JP: UVB-induced
cyclobutane pyrimidine dimer frequency correlates with skin cancer
mutational hotspots in p53. Photochem Photobiol. 66:719–726. 1997.
View Article : Google Scholar
|
|
40
|
Podhaisky HP, Riemschneider S and Wohlrab
W: UV light and oxidative damage of the skin. Pharmazie. 57:30–33.
2002.PubMed/NCBI
|
|
41
|
Sander CS, Chang H, Salzmann S, Müller CS,
Ekanayake-Mudiyanselage S, Elsner P and Thiele JJ: Photoaging is
associated with protein oxidation in human skin in vivo. J Invest
Dermatol. 118:618–625. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Beauchamp MC, Letendre E and Renier G:
Macrophage lipoprotein lipase expression is increased in patients
with heterozygous familial hypercholesterolemia. J Lipid Res.
43:215–222. 2002.PubMed/NCBI
|
|
43
|
Bai K, Xu W, Zhang J, Kou T, Niu Y, Wan X,
Zhang L, Wang C and Wang T: Assessment of free radical scavenging
activity of dimethylglycine sodium salt and its role in providing
protection against lipopolysaccharide-induced oxidative stress in
mice. PLoS One. 11:e1553932016. View Article : Google Scholar
|
|
44
|
Ohsawa I, Ishikawa M, Takahashi K,
Watanabe M, Nishimaki K, Yamagata K, Katsura K, Katayama Y, Asoh S
and Ohta S: Hydrogen acts as a therapeutic antioxidant by
selectively reducing cytotoxic oxygen radicals. Nat Med.
13:688–694. 2007. View
Article : Google Scholar : PubMed/NCBI
|
|
45
|
Shi Q, Liao KS, Zhao KL, Wang WX, Zuo T,
Deng WH, Chen C, Yu J, Guo WY, He XB, et al: Hydrogen-rich saline
attenuates acute renal injury in sodium taurocholate-induced severe
acute pancreatitis by inhibiting ROS and NF-kB pathway. Mediators
Inflamm. 2015:6850432015. View Article : Google Scholar
|
|
46
|
Done AJ and Traustadóttir T: Nrf2 mediates
redox adaptations to exercise. Redox Biol. 10:191–199. 2016.
View Article : Google Scholar : PubMed/NCBI
|
|
47
|
Shih PH, Yeh CT and Yen GC: Anthocyanins
induce the activation of phase II enzymes through the antioxidant
response element pathway against oxidative stress-induced
apoptosis. J Agric Food Chem. 55:9427–9435. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
48
|
Uchi H, Yasumatsu M, Morino-Koga S, Mitoma
C and Furue M: Inhibition of aryl hydrocarbon receptor signaling
and induction of NRF2-mediated antioxidant activity by
cinnamaldehyde in human keratinocytes. J Dermatol Sci. 85:36–43.
2017. View Article : Google Scholar
|
|
49
|
Sun Z, Park SY, Hwang E, Zhang M, Seo SA,
Lin P and Yi TH: Thymus vulgaris alleviates UVB irradiation induced
skin damage via inhibition of MAPK/AP-1 and activation of Nrf2-ARE
antioxidant system. J Cell Mol Med. 21:336–348. 2017. View Article : Google Scholar
|
|
50
|
Shan Y, Wei Z, Tao L, Wang S, Zhang F,
Shen C, Wu H, Liu Z, Zhu P, Wang A, et al: Prophylaxis of Diallyl
disulfide on skin carcinogenic model via p21-dependent Nrf2
stabilization. Sci Rep. 6:356762016. View Article : Google Scholar : PubMed/NCBI
|
|
51
|
Ji C, Huang JW, Xu QY, Zhang J, Lin MT, Tu
Y, He L, Bi ZG and Cheng B: Gremlin inhibits UV-induced skin cell
damages via activating VEGFR2-Nrf2 signaling. Oncotarget.
7:84748–84757. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
52
|
Yuan XY, Pang XW, Zhang GQ and Guo JY:
Salidroside's protection against UVB-mediated oxidative damage and
apoptosis is associated with the upregulation of Nrf2 expression.
Photomed Laser Surg. 35:49–56. 2017. View Article : Google Scholar
|
|
53
|
Xie K, Liu L, Yu Y and Wang G: Hydrogen
gas presents a promising therapeutic strategy for sepsis. Biomed
Res Int. 2014:8076352014. View Article : Google Scholar : PubMed/NCBI
|
|
54
|
Diao M, Zhang S, Wu L, Huan L, Huang F,
Cui Y and Lin Z: Hydrogen gas inhalation attenuates seawater
instillation-induced acute lung injury via the Nrf2 pathway in
rabbits. Inflammation. 39:2029–2039. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
55
|
Tamaki N, Orihuela-Campos RC, Fukui M and
Ito HO: Hydrogen-rich water intake accelerates oral palatal wound
healing via activation of the Nrf2/antioxidant defense pathways in
a rat model. Oxid Med Cell Longev. 2016:56790402016. View Article : Google Scholar : PubMed/NCBI
|
|
56
|
Yu J, Zhang W, Zhang R, Jiang G, Tang H,
Ruan X, Ren P and Lu B: Molecular hydrogen attenuates
hypoxia/reoxygenation injury of intrahepatic cholangiocytes by
activating Nrf2 expression. Toxicol Lett. 238:11–19. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
57
|
Nguyen CN, Kim HE and Lee SG:
Caffeoylserotonin protects human keratinocyte HaCaT cells against
H2O2-induced oxidative stress and apoptosis
through upregulation of HO-1 expression via activation of the
PI3K/Akt/Nrf2 pathway. Phytother Res. 27:1810–1818. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
58
|
Hermida MA, Dinesh Kumar J and Leslie NR:
GSK3 and its interactions with the PI3K/AKT/mTOR signalling
network. Adv Biol Regul. 65:5–15. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
59
|
Follo MY, Manzoli L, Poli A, McCubrey JA
and Cocco L: PLC and PI3K/Akt/mTOR signalling in disease and
cancer. Adv Biol Regul. 57:10–16. 2015. View Article : Google Scholar
|
|
60
|
Jhanwar-Uniyal M, Amin AG, Cooper JB, Das
K, Schmidt MH and Murali R: Discrete signaling mechanisms of mTORC1
and mTORC2: Connected yet apart in cellular and molecular aspects.
Adv Biol Regul. 64:39–48. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
61
|
Ricciardi MR, Mirabilii S, Licchetta R,
Piedimonte M and Tafuri A: Targeting the Akt, GSK-3, Bcl-2 axis in
acute myeloid leukemia. Adv Biol Regul. 65:36–58. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
62
|
Jayasooriya RG, Lee KT, Choi YH, Moon SK,
Kim WJ and Kim GY: Antagonistic effects of acetylshikonin on
LPS-induced NO and PGE2 production in BV2 microglial
cells via inhibition of ROS/PI3K/Akt-mediated NF-κB signaling and
activation of Nrf2-dependent HO-1. In Vitro Cell Dev Biol Anim.
51:975–986. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
63
|
Ryu MJ, Kang KA, Piao MJ, Kim KC, Zheng J,
Yao CW, Cha JW, Chung HS, Kim SC, Jung E, et al:
7,8-Dihydroxyflavone protects human keratinocytes against oxidative
stress-induced cell damage via the ERK and PI3K/Akt-mediated
Nrf2/HO-1 signaling pathways. Int J Mol Med. 33:964–970. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
64
|
Seo SH and Jeong GS: Fisetin inhibits
TNF-α-induced inflammatory action and hydrogen peroxide-induced
oxidative damage in human keratinocyte HaCaT cells through
PI3K/AKT/Nrf-2-mediated heme oxygenase-1 expression. Int
Immunopharmacol. 29:246–253. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
65
|
Li H, Chen O, Ye Z, Zhang R, Hu H, Zhang
N, Huang J, Liu W and Sun X: Inhalation of high concentrations of
hydrogen ameliorates liver ischemia/reperfusion injury through A2A
receptor mediated PI3K-Akt pathway. Biochem Pharmacol. 130:83–92.
2017. View Article : Google Scholar : PubMed/NCBI
|