Introduction
Spinal cord injury (SCI) is recognized as a
catastrophic threat to human health, with a gradually increasing
morbidity rate worldwide (1,2).
It is known that hemi-sectioned SCI (hSCI) can lead to spastic
paralysis on the injured side, as well as flaccid paralysis on the
other side, and its injury degree lies between that of contusion
injury and transection injury. hSCI results in marked motor
impairments that harm the daily quality of life (3), however, stem cell therapy could
offer an effective treatment solution (4-7).
It is known that stem cells are a type of
multi-potent cell with the ability to replicate (self-renewal).
Under certain conditions, they can differentiate into various
functional cells (8,9). Neural stem cells (NSCs), an
important type of stem cell, possess the potential for giving rise
to cells that are identical to themselves (self-renewal) (10) and producing neurons, astrocytes
and oligodendrocytes (pluripotency) (11). NSC transplantation may shed new
light on the practical study of central nervous system injury
(4,12), and NSCs could serve as the ideal
source of transplanted cells. Moreover, NSCs are able to restore
defective nerve tissues by self-proliferation and differentiation
(13), and secrete a variety of
neurotrophic factors (NTFs), including ciliary NTF (CNTF),
transforming growth factor-β1 (TGF-β1), glial cell line-derived NTF
(GDNF), nerve growth factor (NGF), brain-derived NTF (BDNF) and
insulin-like growth factor (IGF) (14). As grafted cells, the expression of
these NTFs from NSCs may support the survival and proliferation of
host neural cells by improving the microenvironment, promoting
expression of regeneration-related genes and accelerating local
spinal cord restoration (15-17). However, the extensive evidence
concerning the recovery of motor function following NSC
transplantation after SCI has mainly been reported in rodents
(18,19). Primates are so scarce and
valuable, while rodents including rats and mice are greatly
different from primates. Therefore, tree shrews are the best choice
for the source of NSCs. Crab-eating monkeys have such disadvantages
as a high cost, lower availability and difficulty in breeding.
However, for human embryos, ethical issues exist (20-23). Thus, a novel animal model for NSC
transplantation is required. With the successful isolation of NSCs
in the adult rodent, the study of NSCs in tree shrews (TS), a
non-human primate, is becoming a focus, as they present with more
advanced development than rats, have a lower economic cost and are
a more convenient resource than monkeys, and the closest relative
to primates (24,25).
Furthermore, TSs have successfully been turned into
an animal model for the study of hepatitis B virus infection
(26-28), myopia (29), depression (30) and bacterial infection (31). However, TSs in the hSCI model and
TS NSC transplantation into the hSC have not previously been
studied, and the molecular factors of TS NSC grafts remain to be
decided. Therefore, in the present study, a TS hSCI model was
established to investigate the effect of TS NSC transplantation on
the motor function improvement in the acute and chronic stages, and
the possible mechanisms involved in associated features such as
proliferation, differentiation and NTF secretion were examined.
Materials and methods
Animals and ethics statement
The present study used 3 female TSs who were 20 days
pregnant and 50 healthy adult female TSs, weighing 120±20 g, 6
months old, which were obtained from the Experimental Animal Center
of Kunming Medical University (Kunming, Yunnan, China) and were
housed in the Laboratory Animal Center of Sichuan University
(Chengdu, Sichuan, China). Animal experimental protocols were
approved and performed according to the guidelines of the
Institutional Medical Experimental Animal Care Committee of Sichuan
University, West China Hospital (Sichuan, China). Guidelines for
Laboratory Animal Care and Safety from the Unites States National
Institutes of Health (Bethesda, MD, USA) were also followed.
Animals were raised in separated cages in a room with a temperature
of 20±5°C, 40-60% humidity and using a 12:12-h light/dark cycle,
with free access to pellet chow and water. The breeding cages were
plastic, equipped with stainless steel cup with lid cover and a
plastic water jar. Following hSCI experiments, body temperatures
were maintained, and the bladders of the TSs were manually massaged
three times daily to enhance their function.
Isolation of NSCs
The pregnant TSs were euthanized with a mixture of
70% CO2 and 30% O2. Next, the embryonic TSs
were harvested under sterile conditions and immersed in an
incubator filled with D-Hank's solution (Thermo Fisher Scientific,
Inc., Waltham, MA, USA). Under an anatomical microscope, the
hippocampi of the embryonic TSs were harvested and placed in a
centrifuge tube for later use. To isolate the NSCs, the hippocampi
were sectioned into small pieces and digested with 0.25% trypsin
(1:250; Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) for 20 min,
and then Dulbecco's modified Eagle's medium (DMEM)/F12 (1:1; Gibco;
Thermo Fisher Scientific, Inc.) containing serum was added to stop
the digestion. Following centrifugation at 560 × g (4°C) for 5 min,
the supernatant was discarded, and 100 ml DMEM/F12 medium (1:1) was
added, with addition of 2 ml 1% B27 (both Gibco; Thermo Fisher
Scientific, Inc.), 2 mmol/l glutamine, 2 μg basic fibroblast
growth factor (basic FGF; PeproTech, Inc., Rocky Hill, NJ, USA), 1%
N2 (Gibco; Thermo Fisher Scientific, Inc.), 10,000 U/l penicillin
and 10 mg/l streptomycin. Next, the cell suspension was harvested
and the cells were inoculated onto culture plates or bottles at a
density of 5×105/ml, then kept in an incubator
containing 5% CO2 at 37°C. The culture medium was half
replaced every other day.
NSC passaging
At 7 days post-culture, when the cells had grown to
near confluency, they were passaged two to four times. Briefly,
following centrifugation at 560 × g (4°C) for 5 min in a 15-ml
centrifuge tube, the supernatant was discarded. The cell suspension
was re-suspended into DMEM/F12 containing FGF (20 ng/ml) and
epidermal growth factor (20 ng/ml). The cellular density was
adjusted to 1.5-2.5×106/ml and inoculated into the
culture bottles (25 ml in volume). Subsequent to lightly swaying
the culture bottle for well distribution, the cultured NSCs were
incubated in an 37°C incubator.
Cultured NSCs at passage two (P2) were collected for
later use: A small quantity of NSCs was used for cell
identification, some of them were used to identify their
differentiation ability, some were used to detect their secretory
functions and a large number were marked by Hoechst 33342 for
transplantation in the hSC.
Morphological observation
During the primary and secondary culture of cells
derived from the hippocampus of the TSs, an inverted phase-contrast
microscope (Leica Microsystems GmbH, Wetzlar, Germany) was employed
to observe and record the morphology and growth situation,
including size, diameter and shape of NSCs at day 1-7 of P0, P1 and
P2, respectively. The formation of neurospheres was also
observed.
Experimental groups
A total of 50 female TSs were randomly divided into
the sham group, the acute hSCI with medium (MEM) (acute control)
group, the acute hSCI with NSC implantation (acute) group, the
chronic hSCI with NSC implantation (chronic) group and the chronic
hSCI with MEM (chronic) group, with 10 TSs in each group (Table I). TSs in the acute or acute
control groups underwent hSCI and were treated with NSC suspension
or MEM immediately after injury. TSs in the chronic or chronic
control groups were subjected to hSCI and received the NSC
suspension or MEM at day 9 post-injury. TSs in the sham group
underwent neither hSCI nor transplant injections (Table I).
 | Table IAnimals and grouping (n=50). |
Table I
Animals and grouping (n=50).
| Groups | n | BBB | RT-qPCR | IF |
|---|
| Sham | 10 | 10 | 5 | 5 |
| Acute hSCI with
NSCs | 10 | 10 | 5 | 5 |
| Acute hSCI with
MEM | 10 | 10 | 5 | 5 |
| Chro hSCI with
NSCs | 10 | 10 | 5 | 5 |
| Chro hSCI with
MEM | 10 | 10 | 5 | 5 |
hSCI model
All TSs were anesthetized intraperitoneally with by
2% sodium pentobarbital (30 mg/kg) and placed in the prone
position. A midline skin incision was made at the thoracic area
(T8-T12) and then the paravertebral muscles and supraspinal
ligaments were separated. TSs then underwent a T10 laminectomy and
received an hSCI. The surgical wounds were closed with 3-0 silk
sutures, and the TSs were injected with normal saline (0.9%; 5 ml)
and received cefotaxime sodium (2.5%; 0.5 ml) for 7 days. The
bladders of the TSs were manually compressed three times a day
until the recovery of the micturition reflex.
NSC labeling and transplantation
Hoechst 33342, a nuclear dye was used for NSC
transplantation labeling. To label implanted cells, 3 mg/ml Hoechst
(Sigma-Aldrich; Merck KGaA) was added to the medium at 72 h prior
to transplantation. For tracing cells after transplantation, NSCs
labeled with Hoechst 33342 could be observed under a fluorescence
microscope (Leica Microsystems GmbH). For transplantation, cells
were isolated by treatment with 0.25% trypsin (Sigma-Aldrich; Merck
KGaA) and 0.5 mm EDTA (Sigma-Aldrich; Merck KGaA) at room
temperature for 10 min. The digestion was stopped by adding 1 ml
fetal bovine serum. The cells were then washed five times with
phosphate-buffered saline (PBS). Nucleated NSCs were counted using
a cytometer to guarantee sufficient NSC numbers for
transplantation. In total, ~6 μl cell suspension
(1×107/ml) or equivalent normal MEM was slowly
transplanted into the spinal cord around the injury site, and at
day 9 post-injury, respectively. Following transplantation, the T10
spinous process was tightened with 1-0 silk suture to prevent
kyphosis and to obtain contact between the graft and spinal cord
stumps. Muscle and skin were sutured layer to layer, and the TSs
were placed in warm cages overnight. Food and water were provided
ad libitum. Manual bladder expression was performed twice a
day until recovery of the bladder reflex.
Behavior evaluation
To compare the effect of NSC transplantation on the
injured spinal cord, locomotor functional recovery was examined in
the TSs at days 1-16 following hSCI, as determined by the
Basso-Beattie-Bresnahan motor score (BBB score), which was graded
from 0 points (absence of any hind limb movement) to 21 points
(normal mobility) (32). Subjects
were acclimated to the open enclosure (99 cm in diameter, 23 cm
deep) for 3 days prior to detection for 5 min/day, as TSs often
remain motionless when introduced to a new apparatus. Each subject
was then observed for 4 min. All measurements were conducted in a
double-blind manner. The mean score was determined from 3
individual researchers.
Immunohistochemistry
Immunohistochemical staining was performed to
confirm the cultured NSCs and their differentiation ability, as
well as to detect the expression of the trophic factors in
vitro. Meanwhile, the survival, migration and differentiation
of NSCs in vivo after transplantation were also
evaluated.
For in vitro detection, the cell suspension
of P2 was moved to the 6-well plates and dropped onto sterile cover
slips. The 6-well plates containing the NSCs were incubated at 37°C
and the purified NSCs were fixed with 4% paraformaldehyde for 20
min. For in vivo detection, at 16 days post-injury, the TSs
were anesthetized with 2% pentobarbital sodium 30 mg/kg
[intraperitoneal (i.p.)] and underwent transcardiac perfusion with
heparinized 0.9% saline, followed by 4% formaldehyde in 0.1 ml
ice-cold phosphate buffer (pH 7.4; Shanghai Haoran Biotechnology
Co., Ltd., Shanghai, China). Next, the samples collected from 20-mm
long spinal cord segments containing the injury and injection
sites, were post-fixed for 5 h at 4°C. Once the spinal cords from
the different groups had been embedded in the same paraffin,
respectively, the blocks were cut into 4-μm sections for
later use. After routine de-paraf-finization and rehydration,
immunohistochemistry analysis was performed on the sections of
spinal cord tissue.
Rinsed with 0.01 M PBS, the glass slides of the
cultured NSCs and spinal cord sections were incubated with 5% goat
serum for 30 min at 37°C to quench non-specific binding. Next, they
were incubated overnight at 4°C with primary antibodies of nestin
(a marker of NSCs), neuron-specific nuclear protein (NeuN; a marker
of neurons), glial fibrillary acidic protein (GFAP; marker of
astrocytes) and trophic factors (CNTF, TGF-β1, GDNF, NGF, BDNF and
IGF) (Table II). As for the
control group, the primary antibody was substituted with 0.01 M
PBS. Thereafter, the glass slides and tissue sections were washed
with 0.01 M PBS three times, each for 2 min. The slides and
sections were then incubated with secondary antibodies (Table II) at 37°C for 30 min. Sections
were observed under an immunofluorescence microscope (Leica
Microsystems GmbH). Furthermore, 4′,6-diamidino-2-phenylindole
(DAPI) was used to counterstain the nuclei. Finally, in order to
determine the positive number stained in vitro, cells from
five fields in each well were collected, each detection in
vitro was prepared for 6 plates (6-pore plate) of cells and
each pore was put into one sterile cover slip.
| Table IIAntibody list. |
Table II
Antibody list.
| Primary
antibody | Company | Species | Concentration | Secondary
antibody | Company | Species | Concentration | Cat. nos. |
|---|
| IGF | ZSGB-BIOa | Rabbit | 1:50 | Cy3 | Jacksonb | Goat
anti-rabbit | 1:200 | 111-165-003 |
| BDNF | ZSGB-BIO | Rabbit | 1:50 | Cy3 | Jackson | Goat
anti-rabbit | 1:200 | 111-165-003 |
| NGF | ZSGB-BIO | Rabbit | 1:50 | Cy3 | Jackson | Goat
anti-rabbit | 1:200 | 111-165-003 |
| GDNF | ZSGB-BIO | Rabbit | 1:100 | Cy3 | Jackson | Goat
anti-rabbit | 1:200 | 111-165-003 |
| TGF-β1 | Abcamc | Rabbit | 1:100 | Cy3 | Jackson | Goat
anti-rabbit | 1:200 | 111-165-003 |
| CNTF | Biossd | Rabbit | 1:50 | Cy3 | Jackson | Goat
anti-rabbit | 1:200 | 111-165-003 |
| GFAP | ZSGB-BIO | Rabbit | 1:50 | Cy3 | Jackson | Goat
anti-rabbit | 1:200 | 111-165-003 |
| NeuNe | Bioss | Rabbit | 1:100 | Cy3 | Jackson | Goat
anti-mouse | 1:200 | 111-165-003 |
| NeuNf | Bioss | Rabbit | 1:100 | 488 | Invitrogeng | Goat
anti-rabbit | 1:100 | A-11034 |
| Nestin | ZSGB-BIO | Rabbit | 1:50 | Cy3 | Jackson | Goat
anti-rabbit | 1:200 | 111-165-003 |
For detection in vivo, five fields for each
section and five sections for each detection were selected
randomly. All the detections were evaluated by 3 investigators
blinded to the experimental information using Image-Pro Plus 6.0
software (Media Cybernetics, Silver Spring, MD, USA), and then the
mean rates of positive cells for each detection were
calculated.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
At 16 days post-injury, the spinal cord caudal to
the injury site (20-mm long containing the injury and injection
sites) was harvested after 2% pentobarbital sodium (30 mg/kg; i.p.)
anesthesia and placed into a 1.5-ml Eppendorf tube without RNase,
then stored at −80°C. Afterwards, the mRNA of the NTFs, including
NGF, CNTF, BDNF, GDNF, TGF-β1 and IGF, were detected by RT-qPCR.
Briefly, total RNA from the spinal cord tissues was isolated using
TRIzol reagent (SuperfecTRI™) according to the manufacturer's
protocol (Invitrogen; Thermo Fisher Scientific, Inc.), and reverse
transcribed to cDNA with the RevertAid™ First Strand cDNA synthesis
kit (Thermo Fisher Scientific, Inc.). Single-stranded cDNAs were
synthesized by incubating template RNA (2.5 μg) with
oligo(dT)18 primer (1 μl) and nuclease-free water
(to 12 μl) at 65°C for 5 min in a volume of 12 μl,
followed by mixing RevertAid M-MuLV Reverse Transcriptase (200
μ/μl, 1 μl) with 5X reaction buffer (4 μl),
RiboLock RNase Inhibitor (20 μ/μl, 1 μl) and
10 mM dNTP Mix (2 μl) with incubation for 60 min at 42°C in
a final volume of 20 μl. The reaction was terminated by
heating to 70°C for 5 min. PCR was performed using s T100™ Thermal
Cycler (Bio-Rad Laboratories, Inc., Hercules, CA, USA) to validate
the level of NTFs; 5 μl of 5-fold diluted template cDNA was
added in each system, with a final volume of 25 μl.
Subsequently, RT-qPCR of cDNA was performed using the forward and
reverse primer sequences as shown in Table III. PCR amplification was
performed as follows: i) Initial denaturation (1 cycle, 95°C for 3
min); ii) denaturation (40 cycles, 95°C for 15 sec); and iii)
amplification (40 cycles, 53°C for 30 sec and 60°C for 40 sec). The
PCR products were verified by 1% agarose gel electrophoresis,
visualized by GoldView (Wolsen) staining (Xi'an Wolsen
Bio-Technology Co., Ltd, Xi'an, China). The gels were scanned using
an AlphaImager gel documentation system (Bio-Rad Laboratories,
Inc.), and bands were quantified using ImageJ software (NIH,
Bethesda, MD, USA).
| Table IIIInformation of primer sequences. |
Table III
Information of primer sequences.
| Gene | Upstream | Downstream | Annealing
temperature, °C |
|---|
| CNTF |
5′-AGGAAGATTCGTTCAGACCT-3′ |
5′-GTTCTCTTGGAGTCGCTCTG-3′ | 53 |
| TGF-β1 |
5′-GGCAGCTGTACATTGACTT-3′ |
5′-AGGGCAAGGACCTTGCTGT-3′ | 53 |
| GDNF | 5′-
TCTGCCTGGTGTTGCTCC-3′ |
5′-CCTCTGCGACCTTTCCCT-3′ | 52 |
| NGF |
5′-GAAGCCCACTGGACTAAACT-3′ |
5′-ACAGTGATGTTGCGGGTCTG-3′ | 54 |
| BDNF |
5′-GGTGTCGTAAAGTTCCACCA-3′ |
5′-GCCAAGTTGCCTTGTCCGT-3′ | 54 |
| IGF |
5′-GATACACATCATGTCGTCTT-3′ |
5′-GCCTGTGGGCTTGTTGAAGT-3′ | 50 |
| β-actin |
5′-GAAGATCAAGATCATTGCTCCT-3′ |
5′-TACTCCTGCTTGCTGATCCA-3′ | 52 |
Statistical analysis
SPSS software (version 17.0; SPSS Inc., Chicago, IL,
USA) was used to process data. The experimental results are
expressed as the mean ± standard deviation, and were analyzed by
Student's t-test with a two-tailed distribution. For multiple group
comparisons, analysis of variance with Tukey's post hoc test was
applied. P<0.05 was considered to indicate a statistically
significant difference.
Results
Morphology of the TS NSCs
At day 1 in primary culture, the cells were small
and round, 5-10 μm in diameter, with no cell processes under
a microscope. The distribution appeared scattered in cell
suspension, but with excellent refraction. At day 3 in primary
culture, small cell spheres arose in the liquid and grew in a
clustered and floating manner. At day 5 in primary culture,
spherical colonies formed by dozens of cells could be observed.
With the development of time, the number and the diameter of the
spherical colonies markedly increased. Following continuous
passaging, at P1 and P2, greater numbers of NSCs were obtained,
whose growth characteristics were similar to that of the original
generation (Fig. 1A).
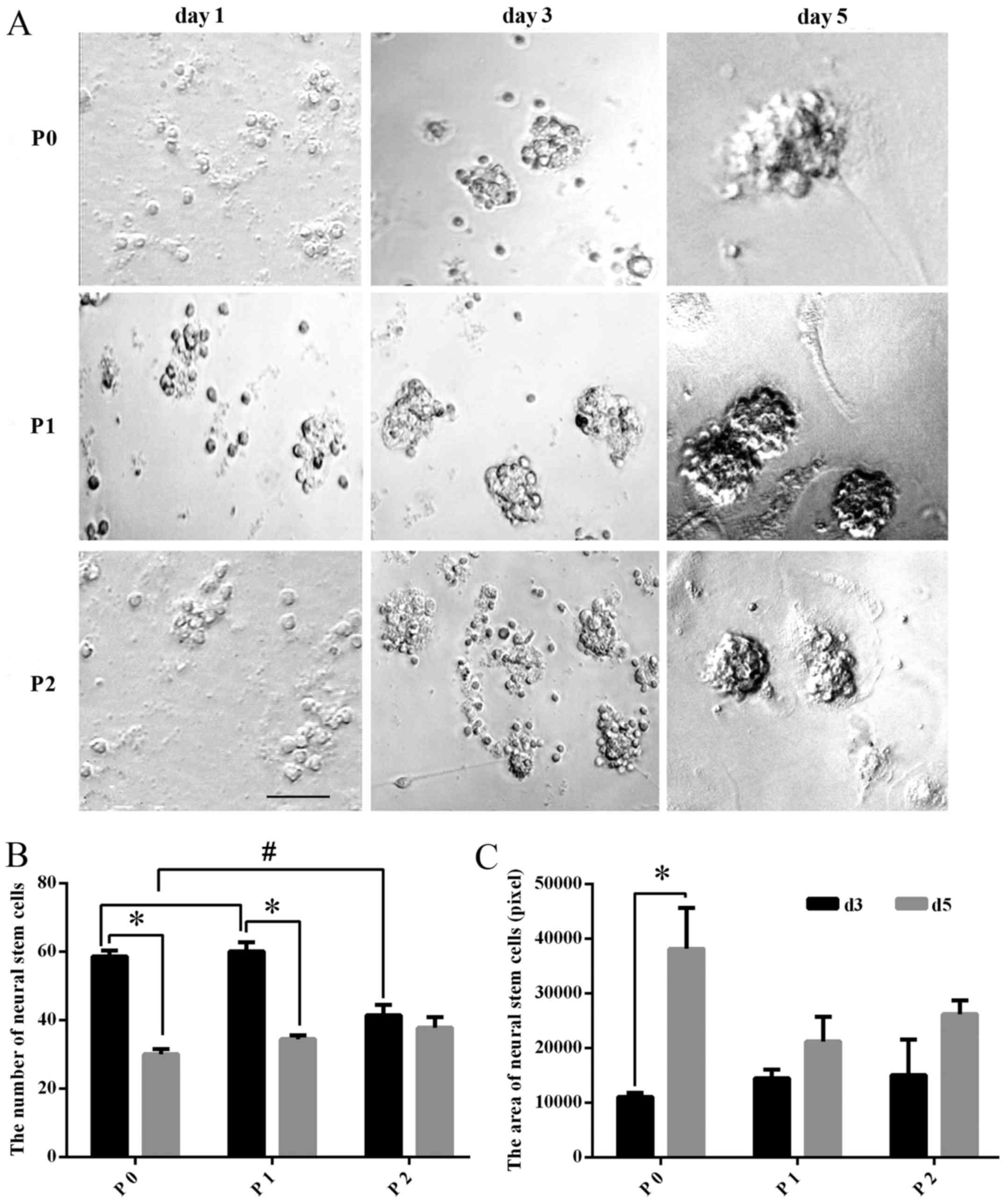 | Figure 1Morphology of TS NSCs cultured in
vitro. (A) TS NSCs were observed under a microscope without dye
processing on days 1, 3 and 5 of primary culture (P0), and
subculture (P1 and P2). Scale bar, 100 μm. (B and C) The
number of NSCs in the different culture stages (P0, P1 and P2) was
compared at different culture times (days 3 and 5). Data are
presented as the mean ± SD (n=5). The area of NSCs in the different
culture stages (P0, P1 and P2) was compared at different times
(days 3 and 5). Data are presented as the mean ± SD (n=6). TS, tree
shrew; NSC, neural stem cell; P, passage; d, day; SD, standard
deviation. |
Proliferation of NSCs cultured in
vitro
The number of NSCs decreased significantly in the P0
and P1 groups (P<0.01), while in P2, the difference in the
number of NSC exhibited no statistical significance (P>0.05)
between days 3 and 5. Among all groups, the number of the cultured
NSCs at day 3 in P2 was significantly fewer than that in P0 or P1
(P<0.01), while for the cultured NSCs at day 5, there was no
significant difference among P0, P1 and P2 (P>0.05) (Fig. 1B). Meanwhile, the size of the
cultured NSCs was also calculated, and the comparison between days
3 and 5 of P0, P1 and P2 showed that the mean size of the NSCs
(neurospheres) at day 5 was significantly larger than that at day 3
in P0, whereas the size changes between days3 and 5 in P1 and P2
exhibited no statistical significance (P>0.05). In addition,
comparison among P0, P1 and P2 groups revealed no significance
(P>0.05) (Fig. 1C).
Identification and differentiation of TS
NSCs
To identify the cultured TS NSCs and evaluate the
differentiation phenotype of the grafted cells in vitro,
immunohistochemical staining of Nestin, NeuN and GFAP was performed
at P2 after culture. In close approximation of our expectations,
the rate of Nestin-positive expression was 99.58%, which confirmed
the purity of the cultured NSCs. Almost all NSCs were NeuN-positive
(97.23%), which implied that the majority of the cultured TS NSCs
differentiated into neurons. The GFAP-positive rate was 46.99% in
NSCs, which explained why a small quantity of NSCs differentiated
into astrocytes (Fig. 2).
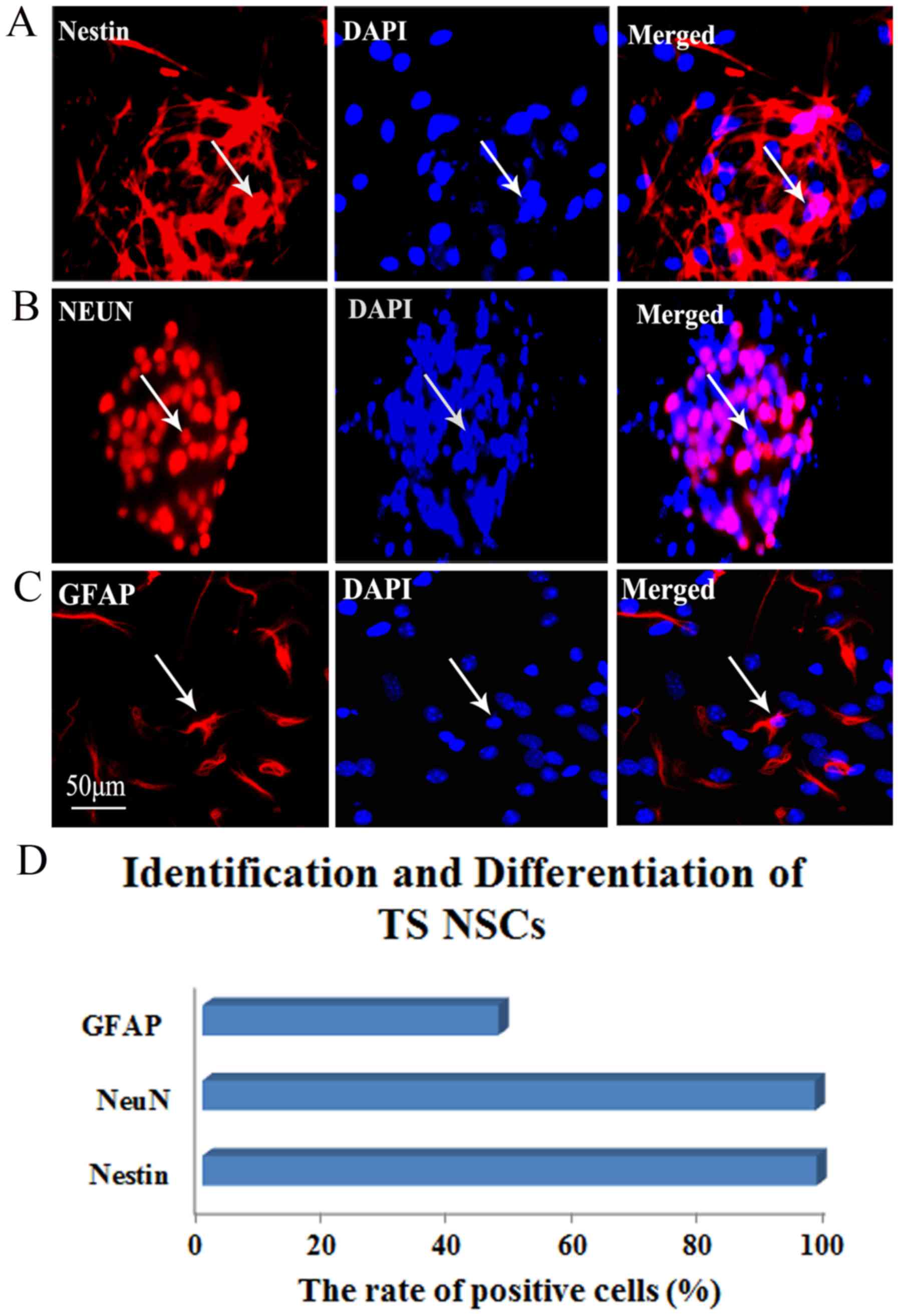 | Figure 2Identification and differentiation of
the cultured TS NSCs. (A) Immunofluorescent staining of Nestin
(red, left) for identifying the TS NSCs, DAPI with blue staining
was shown in middle and the merge picture showed Nestin positive
rate was 99.58% (right). (B and C) Immunofluorescent staining of
NeuN and GFAP for confirmation of the differentiation of the
cultured NSCs into neurons and astrocytes (red, left). DAPI stained
the nuclei with blue florescence (middle). Merged images show the
NeuN- and GFAP-positive cells, from which expression rates of 97.23
and 46.99%, respectively, were obtained (right). White arrows
represent the positive cells. Scale bar, 50 μm. Cells from
five fields in each well were collected, and each detection in
vitro was prepared for 6 plates (6-pore plate) of cells. (D)
Representative bar graph for the rate of positive cells. TS, tree
shrew; NSC, neural stem cell; DAPI, 4′,6-diamidino-2-phenylindole;
GFAP, glial fibrillary acidic protein; NeuN, neuron-specific
nuclear protein. |
Expression of NTFs in the cultured TS
NSCs
Using the double-labeled immunofluorescence staining
of BDNF CNTF, GDNF, IGF, TGF-β1 and NGF with DAPI, the expression
of these NTFs expressed in the cultured TS NSCs were detected at P2
following culture. The results showed that the cultured TS NSCs
exhibited positive immunoreactivity for the aforementioned NTFs.
Moreover, quantitative analysis showed that the positive rates of
CNTF, GDNF and IGF were 100%, and that the positive rate of TGF-β1
was 96.67%. NGF had a positive rate of 89.96%. Furthermore, BDNF
was observed with a positive rate of 50%. This outcome indicated
that NSCs can secrete CNTF, GDNF, IGF, TGF-β1, NGF and a small
amount of BDNF (Fig. 3).
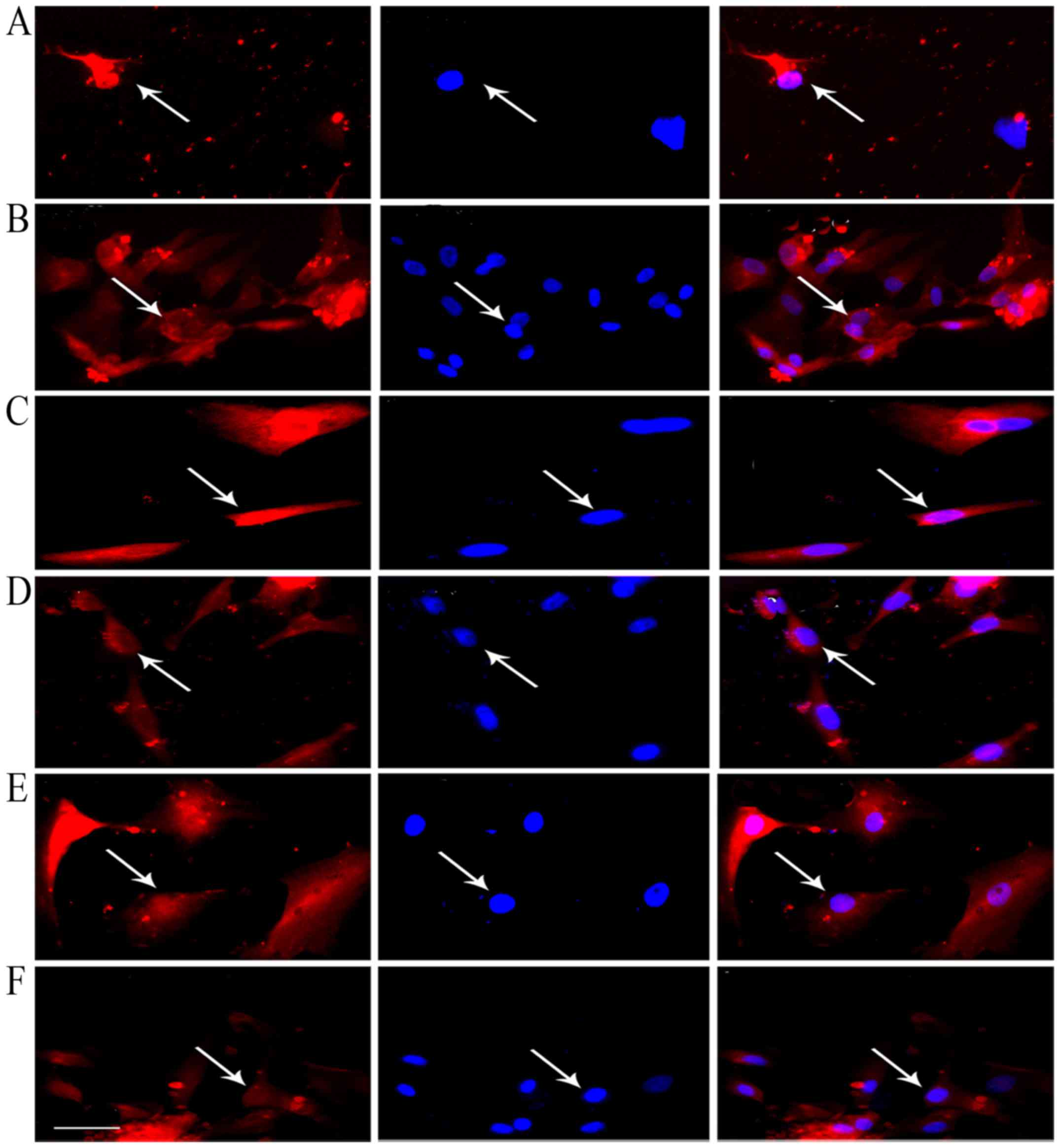 | Figure 3Expression of NTFs in the NSCs of
tree shrews. Immunofluorescent staining of CNTF, TGF-β1, GDNF, NGF,
BDNF and IGF is shown in the left of (A-F) (red), and cell nuclei
were redyed by 4′,6-diamidino-2-phenylindole (DAPI) (blue) in
vitro (A-F, middle). The merged images show positive CNTF (B;
right), GDNF (C; right) and IGF (D; right) expression, with a
determined mean rate of 100%, BDNF (A, right) was 50%, NGF was
89.96% (E, right) and TGF-β1 was 96.67% (F, right). Scale bar, 50
μm. White arrows represented the positive cells. Cells from
five fields in each well were collected, each detection in
vitro was prepared for six plates (six-pore plate) of cells.
NTF, neurotrophic factors; NSCs, neural stem cells; CNTF, ciliary
NTF; TGF-β1, transforming growth factor-β1; GDNF, glial cell
line-derived NTF; NGF, nerve growth factor; BDNF, brain-derived
NTF; IGF, insulin-like growth factor. |
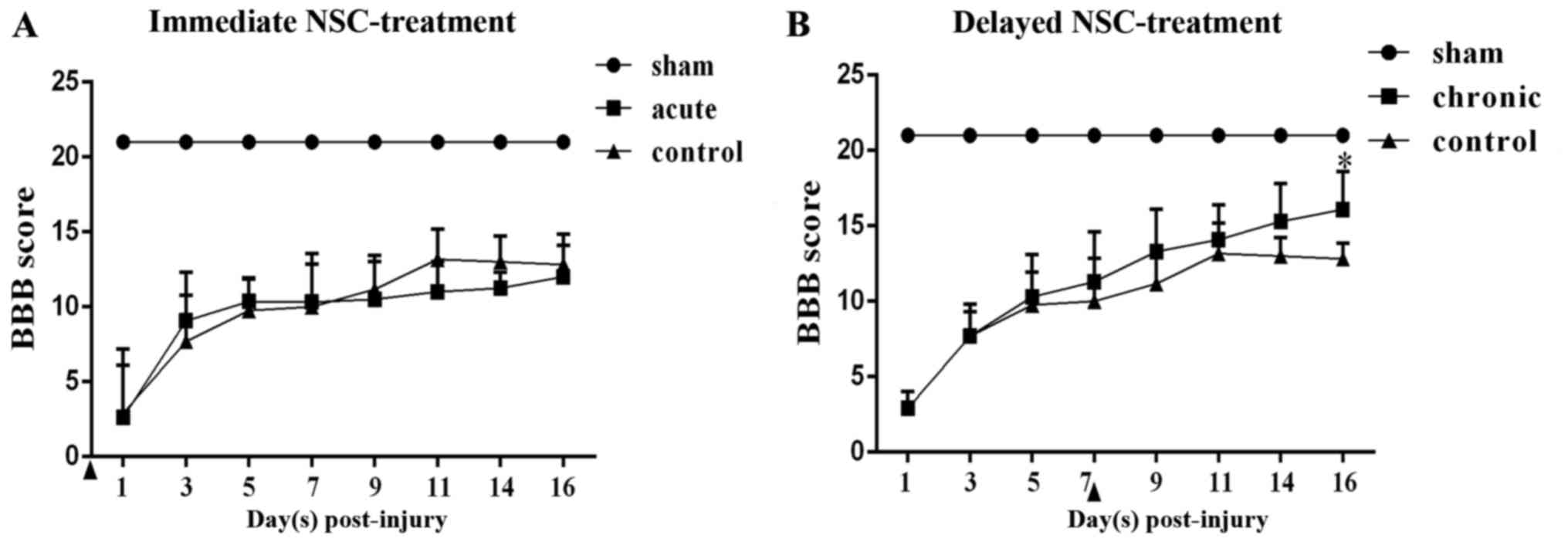
BBB score
The changes of hind-limb function were assessed
using the BBB scores from days 1 to 16 post-hSCI. hSCI resulted in
spastic paralysis on the injury side of the TSs, as well as flaccid
paralysis on the other side, with a BBB score of 3 at day 1
post-injury in the four groups. Over time, the locomotor function
could be partially restored. At day 7 post-injury, the hind-limb
locomotor functions had recovered spontaneously to an approximate
BBB score of 10. Moreover, in the chronic group, significantly
greater functional recovery was observed compared with that in the
chronic control group at day 16 post-injury (P<0.05). By
contrast, the acute group did not show any functional recovery
compared with the acute control group (Fig. 4).
Survival, migration and differentiation
of the transplanted NSCs in vivo
At day 16 post-injury in the chronic group, a large
number of surviving NSCs with blue nuclei staining labeled by
Hoechst 33342 could be found in the spinal cord of the
NSC-transplanted TSs (near the needle passage), whereas there were
no Hoechst-positive cells in the control group; the results
indicated that NSCs could survive and migrate in the spinal cord
around the injection site (Fig.
5A). In order to detect the differentiation of NSCs in the host
spinal cord, the immunohistochemical staining of specific markers
was performed to recognize neurons and astrocytes, as performed
in vitro. The implanted NSCs with blue nuclei staining were
found in the host spinal cord, confirming the survival of the NSCs
(Fig. 5B and C). Simultaneously,
positive staining of neuron marker NeuN confirmed the
differentiation of NSCs into neurons (Fig. 5B). In addition, a few NSCs
exhibited GFAP-positive staining, a marker of astrocytes,
demonstrating the differentiation of the transplanted NSCs into
astrocytes (Fig. 5C). The merged
images revealed that in the chronic group, certain grafted cells
differentiated into NeuN-positive cells (33.8±0.6%), followed by
GFAP-positive astrocytes (20.3±5.1%) (Fig. 5B-D).
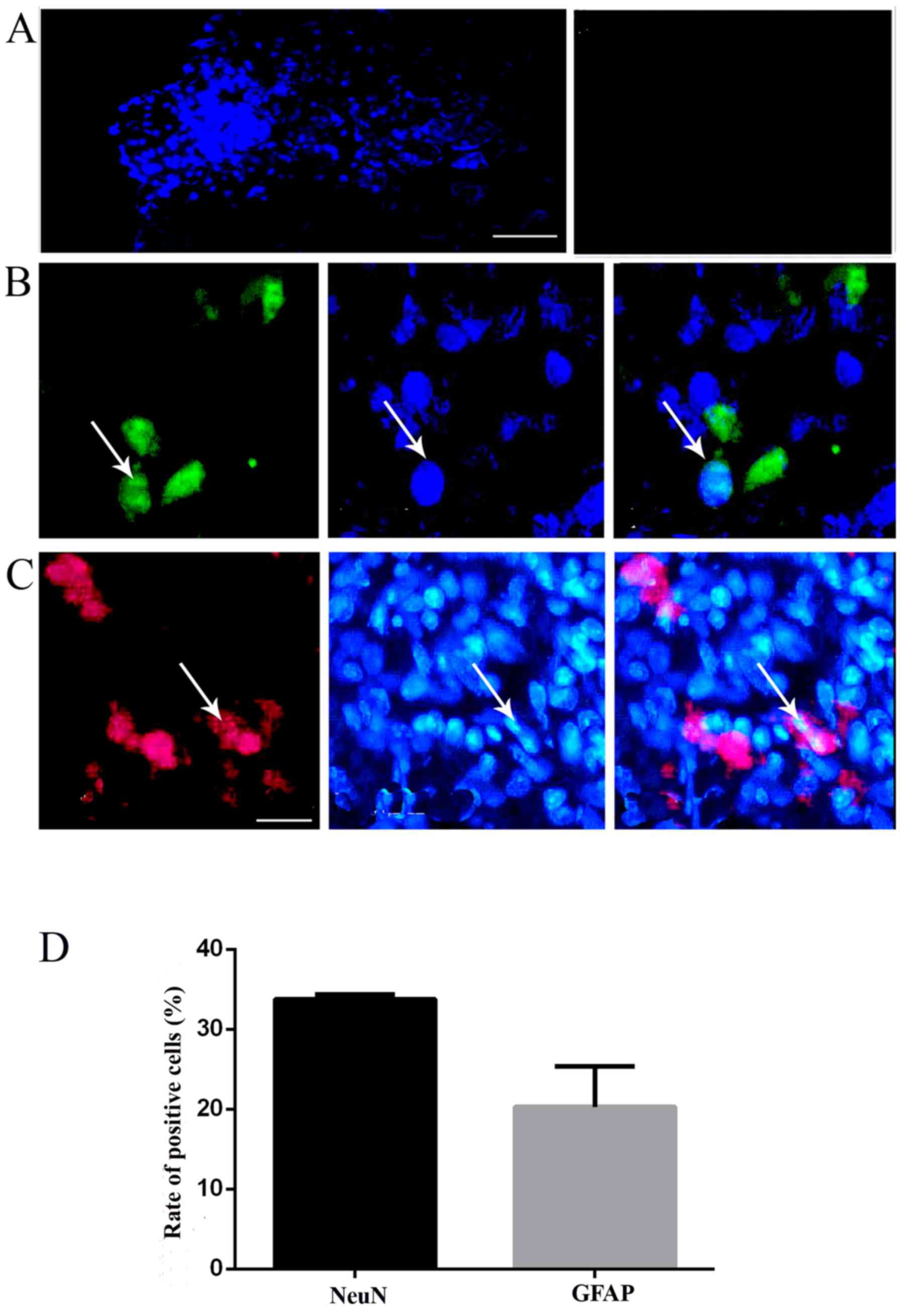 | Figure. 5Transplanted TS NSCs survived and
differentiated into neurons and glia-like cells in vivo. (A)
NSCs labeled with Hoechst (blue fluorescence, left) survived and
migrated at day 16 post-injury in vivo. The control
exhibited no positive reactivity (right). (B) Hoechst+
grafted NSCs (blue florescence, middle) differentiated into
NeuN+ neurons (green florescence, left), and
GFAP+ astrocytes (red, left) in the chronic groups, with
merged images shown in the right panels. Scale bar, (A) 100
μm and (B and C) 50 μm. White arrows indicate the
positive cells. The images were captured 1 cm below the lesion. (D)
Representative bar graph for the rate of NeuN- and GFAP-positive
cells. NSC, neural stem cell; TS, tree shrew; NeuN, neuron-specific
nuclear protein; GFAP, glial fibrillary acidic protein. |
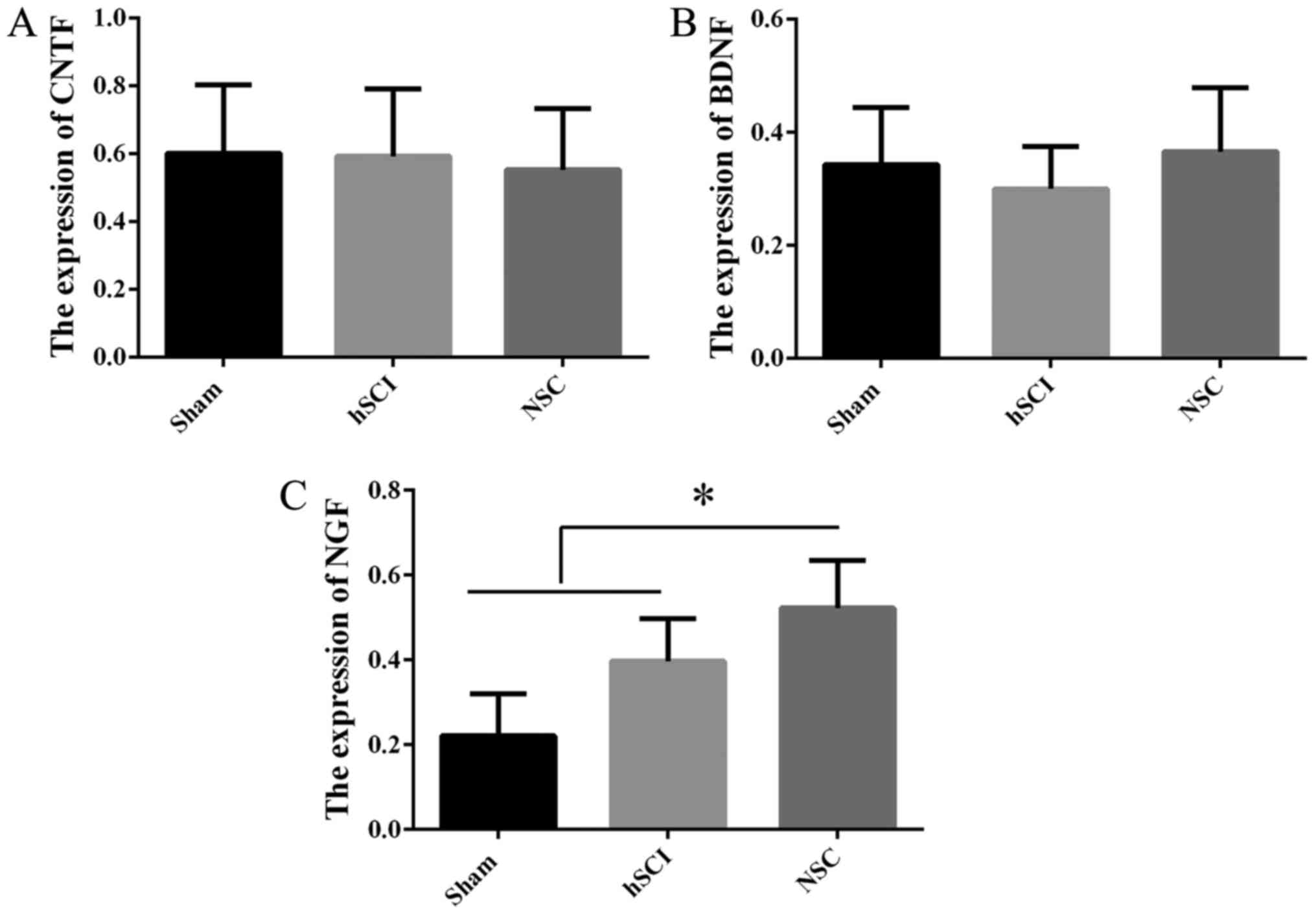
Expression of NTFs in vivo following NSC
transplantation
In order to detect the mechanism of the functional
recovery at day 16 post-injury in the chronic group, RT-qPCR was
employed to confirm NTF gene expression in vivo among the
sham, hSCI and NSCs transplantation groups. Compared with the mRNA
of NSCs from the sham group, NGF gene expression was increased
markedly in the chronic group (P<0.05), while there was no
significant difference for CNTF, BDNF (Fig. 6), IGF, TGF-β1 and GDNF (data not
shown) among the above three groups.
Discussion
In the present study, it was demonstrated in
vitro and in vivo that the NSCs from TSs possessed the
potential for self-renewal, pluripotency into neurons and
astrocytes, and the production of different types of NTFs,
including CNTF, TGF-β1, GDNF, NGF, BDNF and IGF. Following
transplantation in the chronic phase of hSCI, the BBB score
revealed that the locomotor function in the hind-limbs was
improved, but that there was no statistical significance in the
acute-phase transplantation, when compared with the control group.
To the best of our knowledge, these findings, for the first time,
indicate that the transplantation of NSCs from TSs is available for
neurological function improvement following hSCI, but only in the
acute phase, and that the expression of multiple NTFs linked with
the upregulation of NGF is probably involved in the underlying
mechanism. In the experiments assessing the proliferation of NSCs
from TSs, it was confirmed that the NSCs possessed the capacity to
proliferate in vitro and in vivo. In vitro,
the number of NSCs in P0 and P1 reached the growth peak in culture
on day 3. The area of neurosphere formation kept increasing as well
throughout the culture, which innovatively illustrated that NSCs
from TSs can successfully proliferate in vitro. These
results suggested that NSCs from TSs could be considered as an
available cell source for the treatment of disease. In vivo,
via local injections of NSCs, transplanted NSCs from TSs were
verified to survive and proliferate in the hSC of other TSs, which
is useful for understanding the functional repair occurring
following hSCI in non-human primates.
The majority of NSC experimental models have been
focused on rodents and few studies have involved the use of
primates or humans due to the associated ethical issues and the
lack of availability (33-37).
Compared with rats, TSs exhibit biological characteristics and
gross anatomy that are more similar to those of humans (24,38). Furthermore, TSs have a lower
economic cost and are a more convenient resource than other
animals. Therefore, TSs have more advantages as translational
research on NSCs than rodents.
The present study verified that NSCs from TSs could
differentiate into neurons and astrocytes in vivo and in
vitro, which may replace the damaged nerve cells to restore the
structure of the injured spinal cord following transplantation.
Over the past decades, NSCs have been reported to exhibit
multi-directional differentiation to neurons and astrocytes
(13,21,36,39). However, the differentiation
ability of NSCs in non-human primates, such as TSs, was previously
unknown. To the best of our knowledge, the present study was the
first to demonstrate the differentiation characteristics of NSCs
from TSs. The present experiments therefore provided crucial
evidence that NSCs can differentiate into neurons and glial cells,
which assist in reconstructing neural injury.
In the present study, the BBB score exhibited no
significant difference in terms of varying observation points in
the acute phase group compared with those in the acute control
group; this indicated that NSC implantation in the chronic phase
could contribute to the recovery of nervous function in TSs, but
not in the acute phase. A number of studies found that the
degenerative degree of the spinal cord tissue near the spinal cord
transection or contusion was reduced significantly following NSC
transplantation in the chronic phase (40,41), but associated mechanisms involved
in the NTFs were not mentioned, let alone the non-human primate
tree shrew model. Therefore, the present study obtained novel
findings that NSC transplantation into TSs in the chronic phase
could effectively improve nervous function, which may be linked to
the secretion of NTFs in the chronic group.
The results of IHC and RT-qPCR showed that NSCs
could secrete CNTF, TGF-β1, GDNF, NGF, BDNF and IGF in
vitro, while NGF gene expression increased markedly in
vivo. The upregulation of NGF may contribute to neuronal
growth, development and functional integrity, therefore improving
the BBB scores in the hSCI TSs with NSC transplantation. According
to previous studies, there are two main hypotheses for the
effectively promotion of neural functional recovery by NSCs: The
alternative theory and the nutrition theory. Previous studies have
demonstrated that NSC transplantation can differentiate into
neurons and glial cells to repair the neuronal necrosis, but little
evidence shows that grafted NSCs can integrate into the neural
networks of the host. Therefore, whether grafted NSCs can integrate
into host neural network has become important for research, as this
may help to further interpret the role of NSCs transplantation in
the restoration of nerve function (14-17,42-44). The present results showed that the
upregulated NGF expression expressed by NSCs directly or caused by
NSCs indirectly could promote survival and proliferation, and
ultimately improve motor function following hSCI. These results
will aid in understanding the molecular mechanisms for stem cell
therapy in diseases of non-human primates, which may ultimately
become available to future patients in the clinic.
Acknowledgments
This study was supported by a grant from the Key
Grant of Natural Science Foundation from Yunnan Province
(2014FA009) and the National Key Technology Research and
Development Program of the Ministry of Science and Technology of
China (no. 2014BAI01B10) and by the Program Innovative Research
Team In Science and Technology in Yunnan Province.
Notes
[1] Competing
interests
The authors declare that they have no competing
interests.
References
|
1
|
Jain NB, Ayers GD, Peterson EN, Harris MB,
Morse L, O'Connor KC and Garshick E: Traumatic spinal cord injury
in the United States, 1993-2012. JAMA. 313:2236–2243. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Löfvenmark I, Norrbrink C, Nilsson-Wikmar
L, Hultling C, Chakandinakira S and Hasselberg M: Traumatic spinal
cord injury in Botswana: Characteristics, aetiology and mortality.
Spinal Cord. 53:150–154. 2015. View Article : Google Scholar
|
|
3
|
Rognoni C, Fizzotti G, Pistarini C and
Quaglini S: Quality of life of patients with spinal cord injury in
Italy: Preliminary evaluation. Stud Health Technol Inform.
205:935–939. 2014.PubMed/NCBI
|
|
4
|
Dooley D, Vidal P and Hendrix S:
Immunopharmacological intervention for successful neural stem cell
therapy: New perspectives in CNS neurogenesis and repair. Pharmacol
Ther. 141:21–31. 2014. View Article : Google Scholar
|
|
5
|
Whittemore SR: Neuronal replacement
strategies for spinal cord injury. J Neurotrauma. 16:667–673. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Park KI, Liu S, Flax JD, Nissim S, Stieg
PE and Snyder EY: Transplantation of neural progenitor and stem
cells: Developmental insights may suggest new therapies for spinal
cord and other CNS dysfunction. J Neurotrauma. 16:675–687. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Neirinckx V, Agirman G, Coste C, Marquet
A, Dion V, Rogister B, Franzen R and Wislet S: Adult bone marrow
mesenchymal and neural crest stem cells are chemoattractive and
accelerate motor recovery in a mouse model of spinal cord injury.
Stem Cell Res Ther. 6:2112015. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Barnabé-Heider F and Frisén J: Stem cells
for spinal cord repair. Cell Stem Cell. 3:16–24. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Enzmann GU, Benton RL, Talbott JF, Cao Q
and Whittemore SR: Functional considerations of stem cell
transplantation therapy for spinal cord repair. J Neurotrauma.
23:479–495. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Gross CG: Neurogenesis in the adult brain:
death of a dogma. Nat Rev Neurosci. 1:67–73. 2000. View Article : Google Scholar
|
|
11
|
Doetsch F: The glial identity of neural
stem cells. Nat Neurosci. 6:1127–1134. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Gage FH: Mammalian neural stem cells.
Science. 287:1433–1438. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Bonaguidi MA, Wheeler MA, Shapiro JS,
Stadel RP, Sun GJ, Ming GL and Song H: In vivo clonal analysis
reveals self-renewing and multipotent adult neural stem cell
characteristics. Cell. 145:1142–1155. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Lu P, Jones LL, Snyder EY and Tuszynski
MH: Neural stem cells constitutively secrete neurotrophic factors
and promote extensive host axonal growth after spinal cord injury.
Exp Neurol. 181:115–129. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Blesch A and Tuszynski MH: GDNF gene
delivery to injured adult CNS motor neurons promotes axonal growth,
expression of the trophic neuropeptide CGRP, and cellular
protection. J Comp Neurol. 436:399–410. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Grill R, Murai K, Blesch A, Gage FH and
Tuszynski MH: Cellular delivery of neurotrophin-3 promotes
corticospinal axonal growth and partial functional recovery after
spinal cord injury. J Neurosci. 17:5560–5572. 1997.PubMed/NCBI
|
|
17
|
McTigue DM, Horner PJ, Stokes BT and Gage
FH: Neurotrophin-3 and brain-derived neurotrophic factor induce
oligodendrocyte proliferation and myelination of regenerating axons
in the contused adult rat spinal cord. J Neurosci. 18:5354–5365.
1998.PubMed/NCBI
|
|
18
|
Karimi-Abdolrezaee S and Eftekharpour E:
Stem cells and spinal cord injury repair. Adv Exp Med Biol.
760:53–73. 2012. View Article : Google Scholar
|
|
19
|
Wang D and Zhang J: Effects of hypothermia
combined with neural stem cell transplantation on recovery of
neurological function in rats with spinal cord injury. Mol Med Rep.
11:1759–1767. 2015. View Article : Google Scholar
|
|
20
|
Hu YR, Chen G, Wei Y, Wan H, Li JH, Chang
WJ, Zhang XL, Liu R and Sun ZZ: In vitro culture, induction and
differentiation of neural stem cells from rat embryo. J Clin
Rehabilitative Tissue Eng Res. 13:3651–3655. 2009.
|
|
21
|
Hu YY, Duan XH, Chen M and Wang D:
Proliferation and differentiation of neural stem cells from newborn
mouse hippocampi in vitro. Chin J Tissue Eng Res. 17:79–85.
2013.
|
|
22
|
Yan SX, Liu XM, Xiang P, Wang P and Wang
D: Culture and differentiation of cynomolgus monkey neural stem
cells. J Clin Rehabilitative Tissue Eng Res. 13:8821–8824.
2009.
|
|
23
|
Razavi S, Razavi MR, Ahmadi N and Kazemi
M: Estrogen treatment enhances neurogenic differentiation of human
adipose derived stem cells in vitro. Iran J Basic Med Sci.
18:799–804. 2015.PubMed/NCBI
|
|
24
|
Janecka JE, Miller W, Pringle TH, Wiens F,
Zitzmann A, Helgen KM, Springer MS and Murphy WJ: Molecular and
genomic data identify the closest living relative of primates.
Science. 318:792–794. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Fan Y, Huang ZY, Cao CC, Chen CS, Chen YX,
Fan DD, He J, Hou HL, Hu L, Hu XT, et al: Genome of the Chinese
tree shrew. Nat Commun. 4:14262013. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Ruan P, Yang C, Su J, Cao J, Ou C, Luo C,
Tang Y, Wang Q, Yang F, Shi J, et al: Histopathological changes in
the liver of tree shrew (Tupaia belangeri chinensis) persistently
infected with hepatitis B virus. Virol J. 10:3332013. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Wang Q, Yang C, Su JJ, Cao J, Ou C, Yang
F, Zhang JJ, Shi JL, Wang DP, Wang XJ, et al: Factors influencing
long-term hepatitis B virus infection of the tree shrew (Tupaia
belangeri chinensis) as an in vivo model of chronic hepatitis B.
Zhonghua Gan Zang Bing Za Zhi. 20:654–658. 2012.PubMed/NCBI
|
|
28
|
Xu G, Gao Z, He W, Ma Y, Feng X, Cai T, Lu
F, Liu L and Li W: microRNA expression in hepatitis B virus
infected primary treeshrew hepatocytes and the independence of
intracellular miR-122 level for de novo HBV infection in culture.
Virology. 448:247–254. 2014. View Article : Google Scholar
|
|
29
|
Grytz R and Siegwart JT Jr: Changing
material properties of the tree shrew sclera during minus lens
compensation and recovery. Invest Ophthalmol Vis Sci. 56:2065–2078.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Wang J, Zhou QX, Tian M, Yang YX and Xu L:
Tree shrew models: A chronic social defeat model of depression and
a one-trial captive conditioning model of learning and memory.
Dongwuxue Yanjiu. 32:24–30. 2011.PubMed/NCBI
|
|
31
|
Li SA, Lee WH and Zhang Y: Two bacterial
infection models in tree shrew for evaluating the efficacy of
antimicrobial agents. Dongwuxue Yanjiu. 33:1–6. 2012.PubMed/NCBI
|
|
32
|
Basso DM, Beattie MS and Bresnahan JC: A
sensitive and reliable locomotor rating scale for open field
testing in rats. J Neurotrauma. 12:1–21. 1995. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Coumans JV, Lin TT, Dai HN, MacArthur L,
McAtee M, Nash C and Bregman BS: Axonal regeneration and functional
recovery after complete spinal cord transection in rats by delayed
treatment with transplants and neurotrophins. J Neurosci.
21:9334–9344. 2001.PubMed/NCBI
|
|
34
|
Wu S, Suzuki Y, Kitada M, Kitaura M,
Kataoka K, Takahashi J, Ide C and Nishimura Y: Migration,
integration, and differentiation of hippocampus-derived neurosphere
cells after transplantation into injured rat spinal cord. Neurosci
Lett. 312:173–176. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Yi BR, Kim SU and Choi KC: Development and
application of neural stem cells for treating various human
neurological diseases in animal models. Lab Anim Res. 29:131–137.
2013. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Reynolds BA and Weiss S: Generation of
neurons and astrocytes from isolated cells of the adult mammalian
central nervous system. Science. 255:1707–1710. 1992. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Vroemen M, Aigner L, Winkler J and Weidner
N: Adult neural progenitor cell grafts survive after acute spinal
cord injury and integrate along axonal pathways. Eur J Neurosci.
18:743–751. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Fan Y, Yu D and Yao YG: Tree shrew
database (TreeshrewDB): A genomic knowledge base for the Chinese
tree shrew. Sci Rep. 4:71452014. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Dromard C, Guillon H, Rigau V, Ripoll C,
Sabourin JC, Perrin FE, Scamps F, Bozza S, Sabatier P, Lonjon N, et
al: Adult human spinal cord harbors neural precursor cells that
generate neurons and glial cells in vitro. J Neurosci Res.
86:1916–1926. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Nishimura S, Yasuda A, Iwai H, Takano M,
Kobayashi Y, Nori S, Tsuji O, Fujiyoshi K, Ebise H, Toyama Y, et
al: Time-dependent changes in the microenvironment of injured
spinal cord affects the therapeutic potential of neural stem cell
transplantation for spinal cord injury. Mol Brain. 6:32013.
View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Tsuji O, Miura K, Fujiyoshi K, Momoshima
S, Nakamura M and Okano H: Cell therapy for spinal cord injury by
neural stem/progenitor cells derived from iPS/ES cells.
Neurotherapeutics. 8:668–676. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Kamei N, Tanaka N, Oishi Y, Hamasaki T,
Nakanishi K, Sakai N and Ochi M: BDNF, NT-3, and NGF released from
transplanted neural progenitor cells promote corticospinal axon
growth in organotypic cocultures. Spine. 32:1272–1278. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Weishaupt N, Blesch A and Fouad K: BDNF:
The career of a multifaceted neurotrophin in spinal cord injury.
Exp Neurol. 238:254–264. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
44
|
Maisonpierre PC, Belluscio L, Friedman B,
Alderson RF, Wiegand SJ, Furth ME, Lindsay RM and Yancopoulos GD:
NT-3, BDNF, and NGF in the developing rat nervous system: Parallel
as well as reciprocal patterns of expression. Neuron. 5:501–509.
1990. View Article : Google Scholar : PubMed/NCBI
|