Introduction
A major priority for public health authorities is to
manage the effects of osteoporosis. Nearly a third of women over
the age of 50 are affected by the disease, around 50% of whom have
a combined lifetime risk of forearm, vertebral or hip fracture,
approximately the same risk that is related to cardiovascular
disease (1). In Europe, the
estimated total direct cost of osteoporosis was €31.7 billion in
2000 and likely to have nearly doubled by 2050 due to demographic
changes in the population (2). An
osteoporotic fracture will by experienced by ~50% of women and 22%
of men over the age of 50 in their lifetime (3). The weakness of bone associated with
osteoporosis may increase the incidence of fractures in the elderly
aged 50 and above. The incidence of osteoporosis is increasing in
the aging society (4).
There is an awareness that there is an interaction
between certain 'lifestyle-related diseases', such as chronic
kidney disease, hypertension or type 2 diabetes with osteoporosis
(5). Furthermore, it has been
shown that certain lifestyle-related diseases (e.g.,
atherosclerosis and type 2 diabetes) increase fracture risk while
being unrelated to changes in bone mineral density (BMD) (6). It is believed that increased
oxidative stress contributes to the association between bone
metabolism and lifestyle-related diseases, by promoting the
fragility of bone in patients through accumulation of advanced
glycation products in bone through various mechanisms, such as
abnormalities in collagen cross-link formation in bone and
decreased osteoblast differentiation resulting in increased death
of osteoblasts or osteocytes (6).
The main feature of osteoporosis is low BMD which is
a highly heritable trait with heritability of 0.5-0.8 (7). Osteoporotic fracture (OF) also has
moderate heritability ranging from about 0.5 to 0.7 and thus acts
as an end-point clinical outcome of osteoporosis (8). We now know that genome-wide
association studies (GWASs) and their meta-analyses have identified
over 20 genes/loci related to risk of osteoporosis and more than 60
related to variations in BMD. In addition, a novel rare nonsense
mutation within the gene leucine-rich repeat-containing
G-protein-coupled receptor 4 (LGR4) strongly related to low BMD and
OF has been identified in a recently published study on
whole-genome sequencing.
MicroRNAs (miRNAs), a class of non-coding RNAs 19-25
nucleotides in length, control the expression of genes at the
post-transcriptional level (9).
The first miRNA to be discovered, lin-4, was found in
Caenorhabditis elegans in 1993, and since then a number of
miRNAs have been identified in different organisms (10). The latest miRBase release (v20,
June 2013) reported that 24,521 miRNA loci had been discovered from
206 species producing 30,424 mature miRNA products. Numerous miRNAs
regulate different pathophysiological events such as organogenesis,
apoptosis, tumorigenesis, proliferation, organ development and
hematopoietic function (11).
A recent in vitro study found that
osteoclastogenesis in human circulating mononuclear cells was
inhibited by miR-146a (12).
Furthermore, another in vitro and in vivo study
showed that osteoclastogenesis in human circulating mononuclear
cells was promoted by miR-148a (13). A further study found higher levels
of miR-133a in circulating monocytes in vivo in
postmenopausal women with low BMD than in those with high BMD, and
thus miR-133a can be identified as a potential biomarker related to
postmenopausal osteoporosis (14). Recently, an analysis of miRNAs
from total bone tissue comparing osteoporotic vs. non-osteoporotic
bone was conducted (15,16). Seeliger et al and
Garmilla-Ezquerra et al identified six miRNAs that were
upregulated in osteoporotic fracture patients: miR-21, miR-23a,
miR-24, miR-25, miR-100 and miR-125b (15,16). LGR4 has been shown to be
associated with the development of osteoporotic facture, and the
differential expression of miR-137 has been identified in the
disease (16-18). LGR4 was also found to be a
potential target of miR-137 in an online miRNA database. In this
study, we investigated the relationship between miR-137 and LGR4
and explored their roles in the pathogenesis of fracture in
patients with osteoporosis.
Patients and methods
Sample collection
A total of 30 osteoporosis patients with OF (n=16)
or without OF (n=14), and 18 patients without osteoporosis at
Department of Orthopedics, The People's Hospital of Huangdai
(Qingdao, China) were included. Patients with metabolic bone
disease, who had undergone therapies such as hormone replacement
therapy, calcitonin, bisphosphonates and fluoride were excluded
from the study. Information of the participants, such as sex, age
at diagnosis, meal preferences (meat vs. vegetables), physical
activity, and previous disease history were collected prior to
study. The Institution's Ethics and Research Committees approved
this study. The patients signed informed consent for participation
in the study. The study was conducted according to the Declaration
of Helsinki. All specimens were obtained after surgical
resection.
RNA isolation and real-time PCR
TRIzol (Invitrogen, San Diego, CA, USA) reagent was
used to extract the total RNA from cultured cells and tissue
samples in accordance with manufacturer's instructions. NanoDrop
ND-2000c (Thermo Fisher Scientific, Inc., Wilmington, DE, USA) was
used to determine the concentrations and purities of RNA extracted.
Spot Check Nucleic Acid Quantitation kit (Sigma-Aldrich, St. Louis,
MO, USA) and gel electrophoresis were used to test the integrity of
RNA following standard protocol. TaqMan MicroRNA reverse
transcription kit (Applied Biosystems, Foster City, CA, USA) was
used to reverse transcribe the RNA to DNA (cDNA) with carefully
designed primer purchased from Applied Biosystems in accordance
with the protocol by supplier. TaqMan hsa-miR-137 amplification kit
(Applied Biosystems) was used to amplify the cDNA in accordance
with manufacturer's instructions. Roche LightCycler 480-II
real-time thermal cycler (Roche, Basel, Switzerland) was used to
perform the quantitative RT-PCR analysis of miR-137 expression
according to the manufacturer's recommendation, and the real-time
PCR were performed using microRNA specific probes and TaqMan
Universal Master Mix (Applied Biosystems) according to the
manufacturer's instructions. RNU43 was used as the internal control
to normalize the expression of LGR4 mRNA. ΔΔCt method was used to
analyze the expression of LGR4 mRNA and miR-137. All experiments
were run 3 times.
Cell culture and transfection
RPMI-1640 medium (Hyclone, Logan, UT, USA)
containing 10% fetal bovine serum (FBS), antibiotics, 1%
streptomycin-penicillin and L-glutamine, was used to incubate U-2
and MC3T3 cells in a humidified atmosphere 5% CO2/95%
air at 37°C at a final concentration of 1×106 cell/ml
for 72 h according to the manufacturer's instructions. After the
confluence reached 80%, Lipofectamine 2000 (Invitrogen Life
Technologies, Carlsbad, CA, USA) was used to transfect 20 pmol of
miR-137 mimics or its inhibitors with U-2 or MC3T3 cells according
to the manufacturer's description. The test was carried out in
triplicate.
ALP activity determination
ALP activity determination kit (Abcam, Boston, MA,
USA) was used to measure the ALP activity in the bone tissue
samples.
Luciferase assay
The fragment 3′ untranslated region (3′UTR) of LGR4
amplified by PCR was inserted into the multiple restrictive sites
of psiCHECK-2 vector (Promega, Madison, WI, USA), and site-directed
mutagenesis kit was used to introduce the mutation to replace the
identified binding sites in the 3′UTR of the LGR4.
Lipofectamine 2000 (Invitrogen Life Technologies)
was used to transiently transfect the U-2 cells with pRL
Renilla as the control vector (Promega) and the
promoter/luciferase reporter gene in accordance with the
manufacturer's instructions, and the pRL Renilla as the
control vector (Promega) was used to correct transfection
efficiency. The cells were collected 48 h after transfection, and
phosphate-buffered saline (PBS) was used to wash the cells, and
then the lysis reagent (Promega) was used to lyze the cells in
accordance with the manufacturer's instruction. Luciferase assay
reagent (Promega) was used to detect the firefly luciferase
activity, Stop & Glo reagent (Promega) was used to detect the
Renilla luciferase activity based on the standard by
supplier. Three independent experiments were carried out.
Western blot analysis
RIPA buffer (Invitrogen Life Technologies) was used
to extract the whole protein from U-2 cells in accordance with
manufacturer's instruction. XCell SureLock Mini-Cell
electrophoresis system (Invitrogen Life Technologies) with 12%
SDS-PAGE was used to separate the total protein, and then
transferred to the polyvinylidene fluoride membranes (Immobilon-P
membrane; Millipore, Billerica, MA, USA). Ponceau S staining was
used to verify the equal loading of proteins. Tris-buffered
saline-Tween-20 (TBS-T) with 5% dried milk (Abcam, Cambridge, UK)
was used to block the membranes, and the anti-LGR4 and anti-ALP
antibodies at a dilution of 1:500 and anti-β-actin at a dilution of
1:5,000 (all from Santa Cruz Biotechnology, Inc., Santa Cruz, CA,
USA) were used to incubate the membrane at 4°C overnight in
accordance with the manufacturer's recommendation, and then the
membrane was washed, the secondary antibody anti-rabbit IgG with a
1:10,000 dilution (Molecular Probes, Eugene, OR, USA) was used to
detect the primary antibody for another 1 h. LI-COR Odyssey Blots
scanner (LI-COR, Lincoln, NE, USA) was used to visualize the
corresponding fluorescence signal in accordance with the
manufacturer's instructions. Anti-α-tubulin (DM1A; Abcam) at a
dilution of 1:1,000 was used to assess the equal protein loading.
Odyssey 2.1 software (LI-COR) was used to calculate the
quantification of protein. The band intensities of LGR4 and
α-tubulin (αTUB) were corrected by subtracting the respective
background intensities. All experiments were performed 3 times.
Immunohistochemistry
PBS with 4% paraformaldehyde was used to fix the
bone tissue samples, and xylenes were used to deparaffinize the
sections, which were next dehydrated by a series of graded ethanol.
Subsequently, hematoxylin and eosin (H&E) was used to stain the
section. The histological score of each slide was evaluated.
Statistical analysis
All data are shown as mean ± standard deviation
(SD). One-way analysis of variance (ANOVA) and Student's t-test
were used to compare between the groups. SPSS version 17.0 (SPSS
Inc., Chicago, IL, USA) was used to analyze the statistical
procedures.
Results
miR-137 is a potential biomarker for the
development of fractures
Real-time PCR was used to find potential biomarkers
for the development of a fracture. We recruited 48 participants for
our study, 18 osteoporosis patients with fracture, 16 without
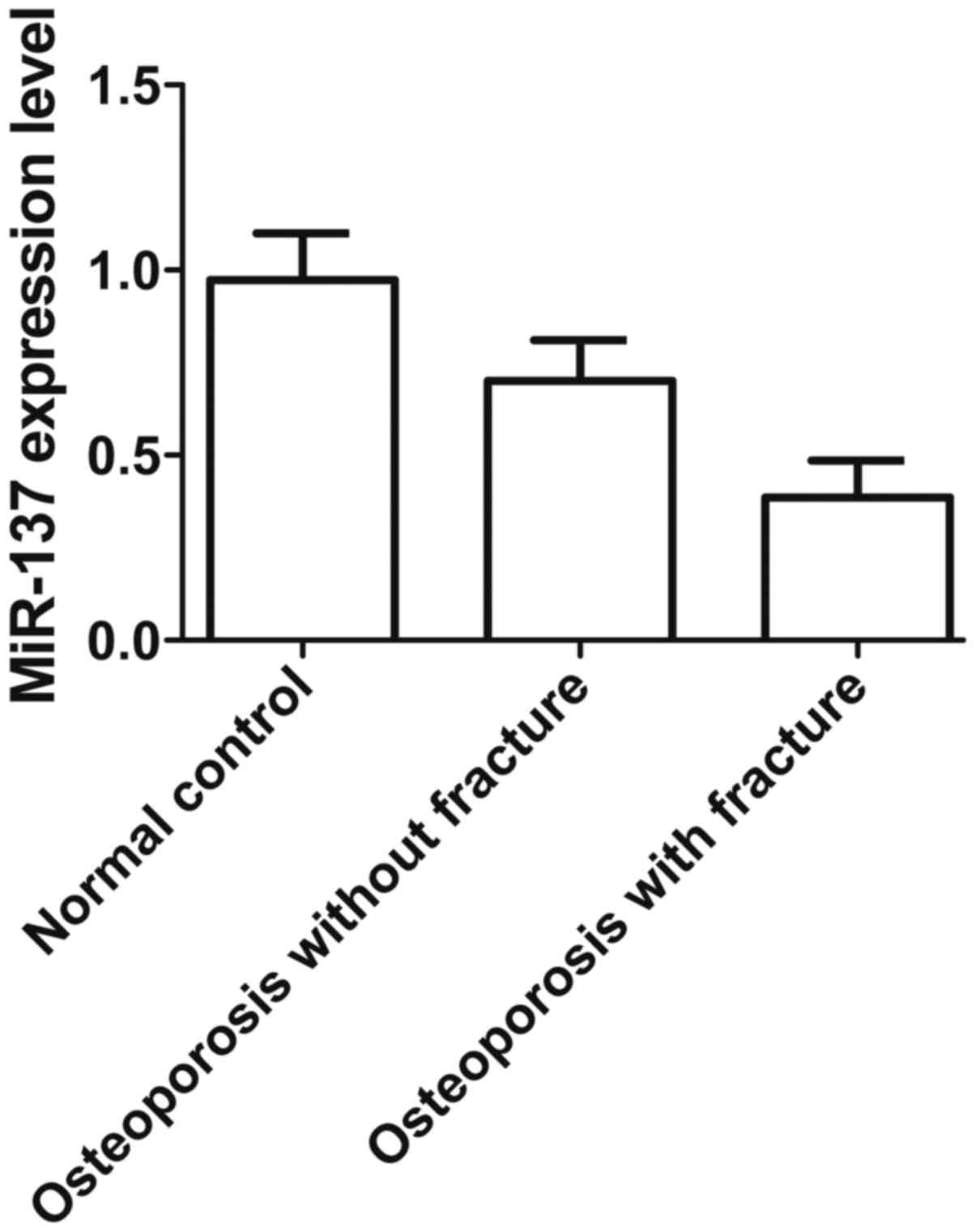
fracture and 14 in the normal group. As shown in Fig. 1, the level of miR-137 expression
was much lower in the group of osteoporosis patients with fracture
compared with those without fracture or the normal group, while the
level of miR-137 expression was significantly higher in the normal
group than in osteoporosis patients without fracture, indicating
that downregulation of miR-137 expression contributes to the
formation of osteoporosis and fracture.
LGR4 is predicted to be a target of
miR-137
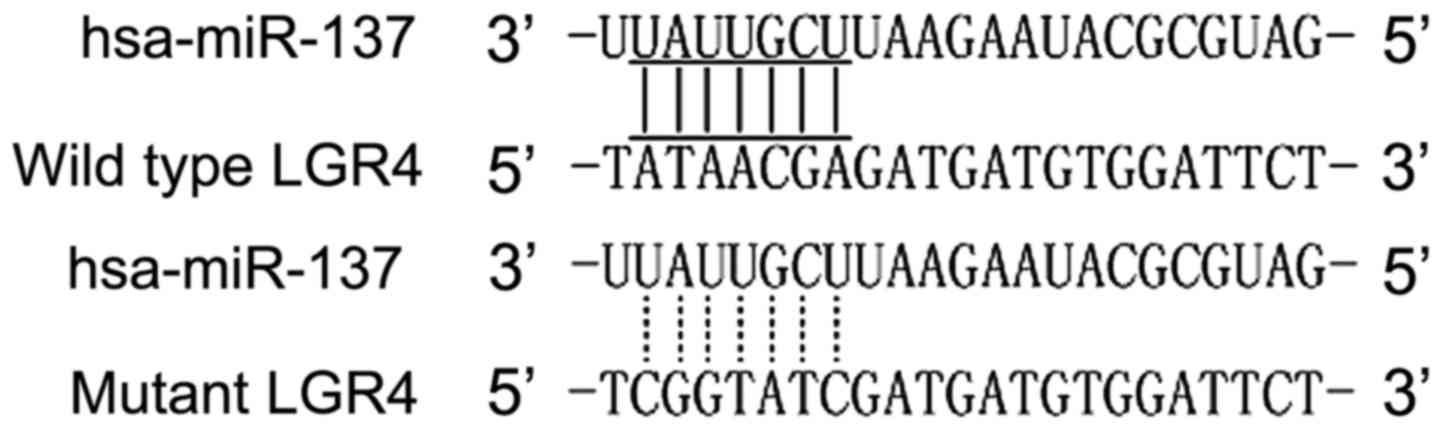
miR-137 was found to be involved in the pathogenesis
of various human diseases, such as osteoporosis and fracture. Based
on computational analysis using the online target predicting tool
TargetScan (www.targetscan.org), we identified
that miR-137 was able to bind to the 3′UTR of LGR4, indicating this
gene may be a potential molecular target for miR-137, as shown in
Fig. 2.
LGR4 was confirmed to be a direct target
of miR-137
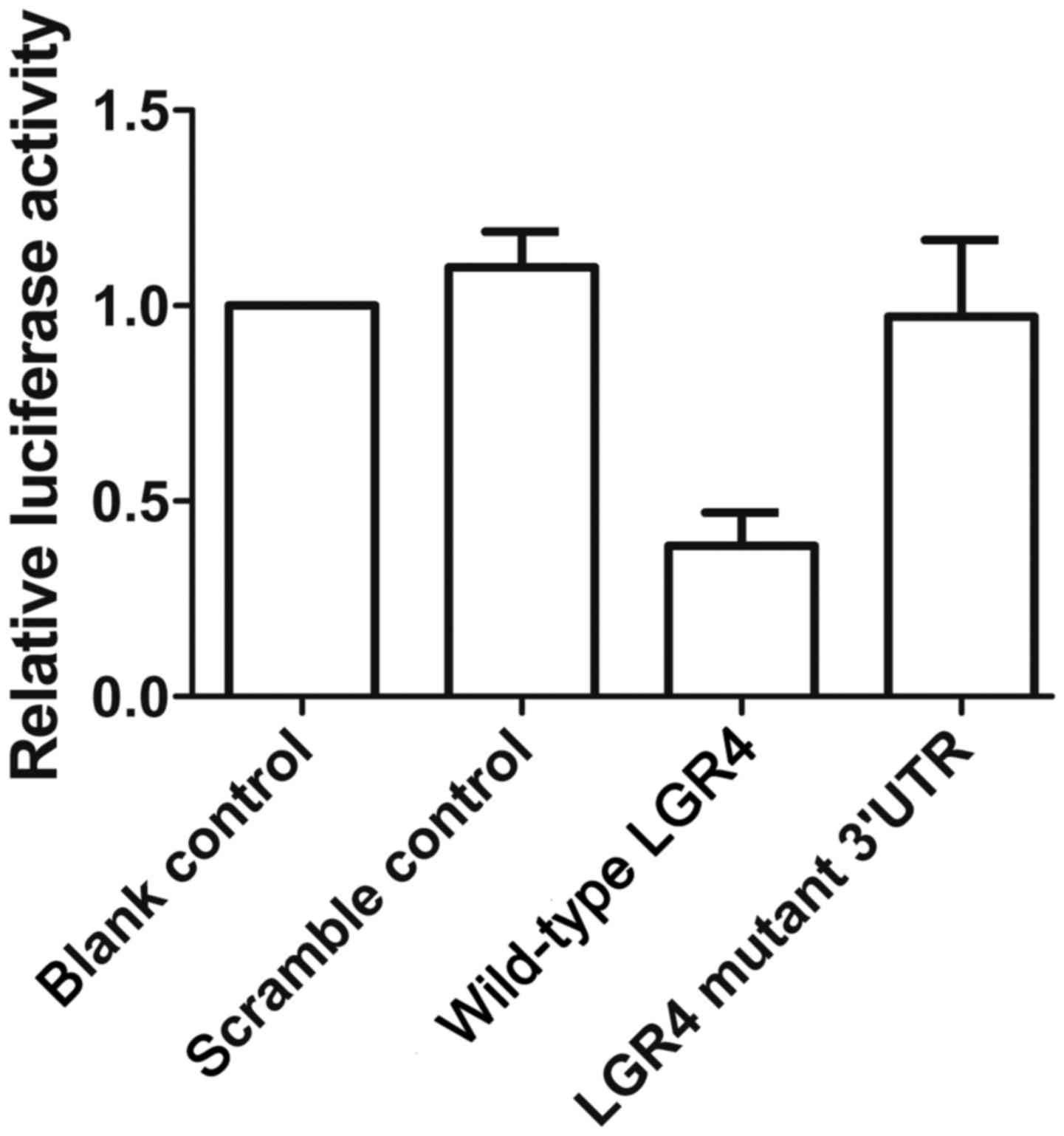
To verify that miR-137 and LGR4 interact with each
other, we performed a luciferase assay to confirm that LGR4 is a
direct shared target of miR-137. As shown in Fig. 3, the luciferase activity of cells
transfected with wild-type LGR4 was significantly downregulated
compared with cells transfected with mutant-type LGR4 and scramble
control, while the luciferase activity in the cells transfected
with mutant 3′UTR of LGR4 was clearly upregulated compared with the
scramble control, confirming that LGR4 is a direct target of
miR-137.
Interaction between miR-137 and LGR4
mRNA
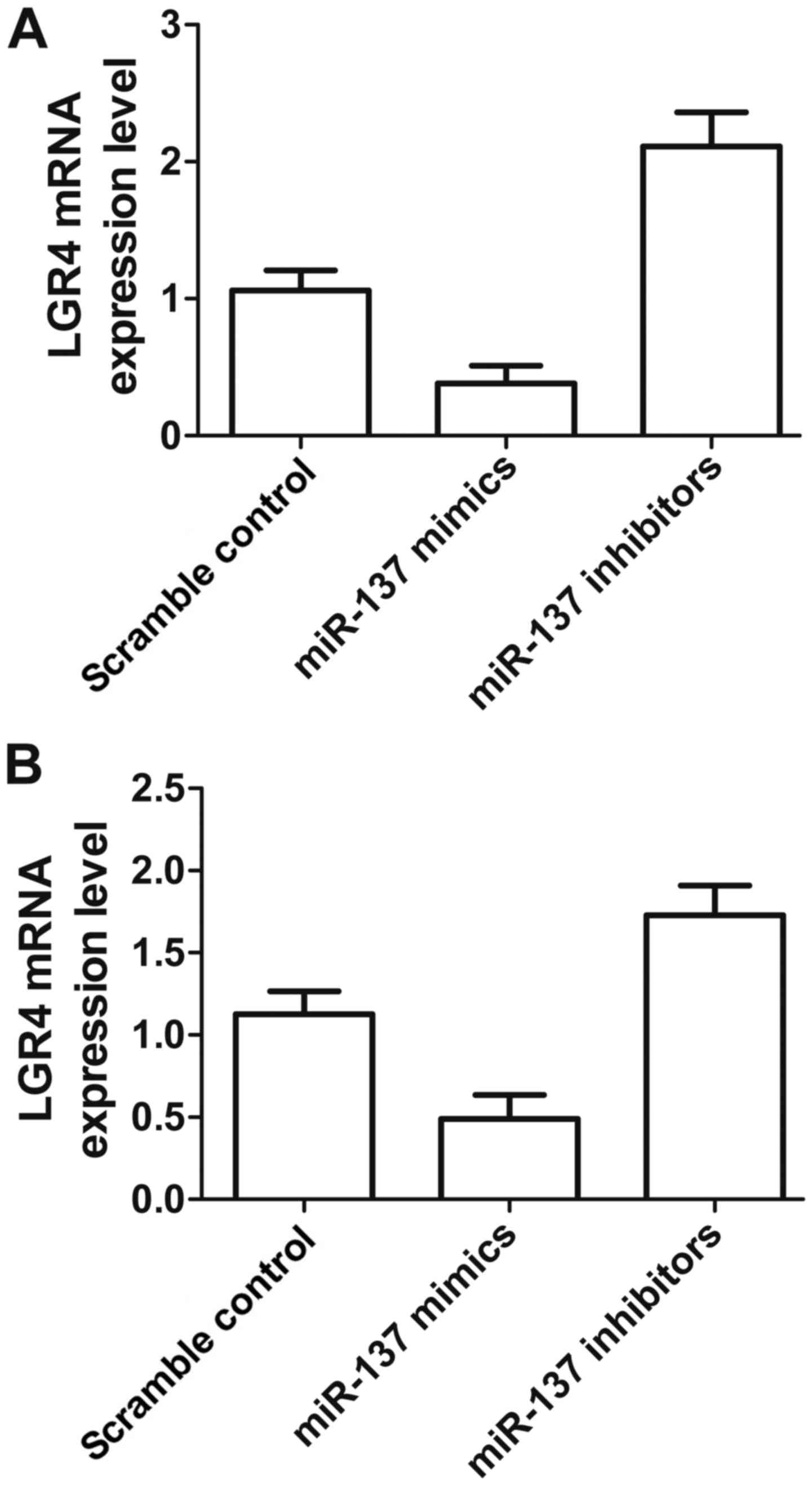
To further confirm the regulatory relationship
between miR-137 and LGR4 mRNA in different cell lines, we
transfected miR-137 mimic and its inhibitor into cultured U-2 and
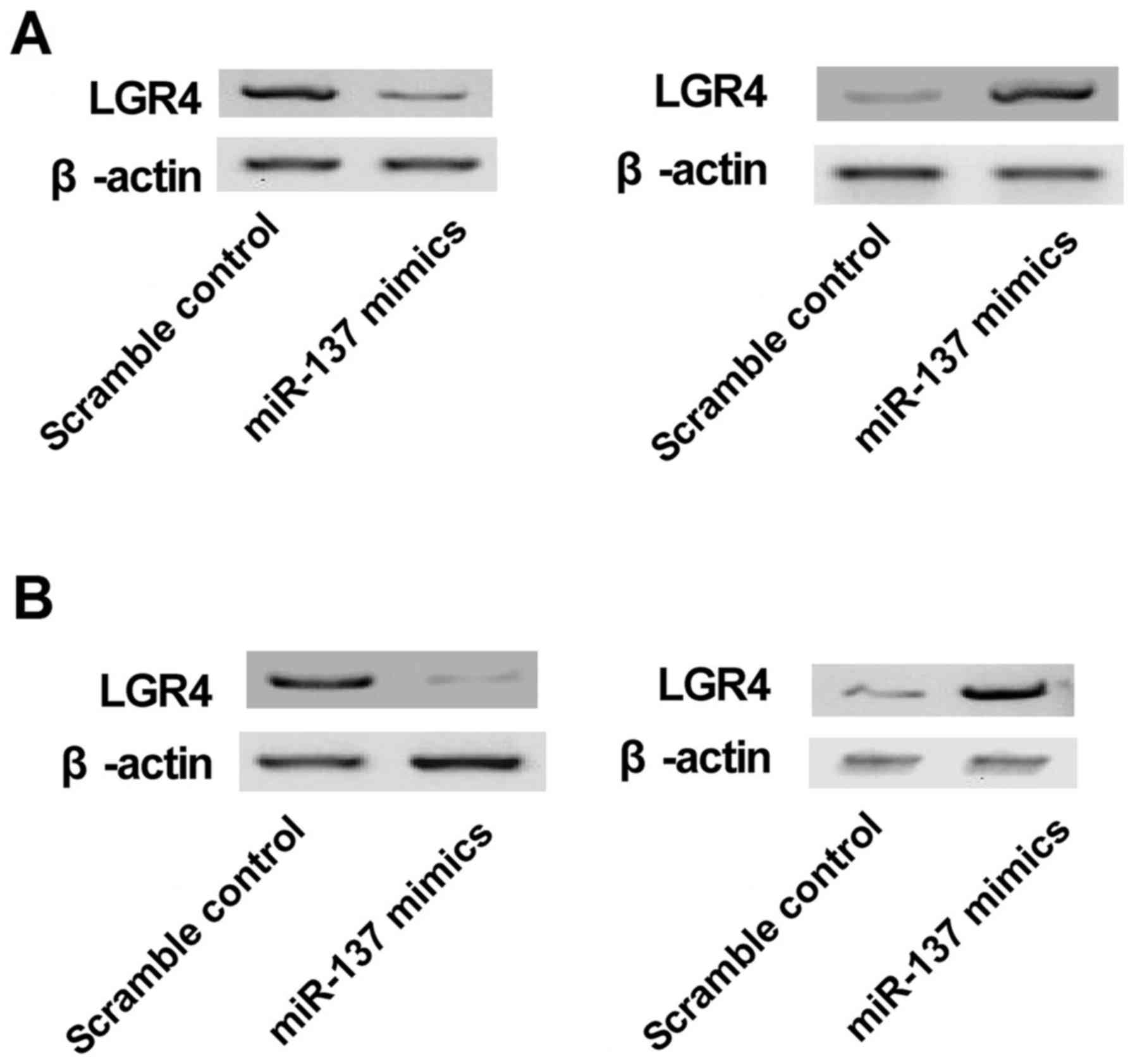
MC3T3 cells. The results of qRT-PCR analysis (Fig. 4A) revealed significantly enhanced
LGR4 mRNA expression in U-2 cells transfected with miR-137
inhibitor and reduced LGR4 mRNA expression in the miR-137 mimic
treatment group compared with the scramble control. The expression
of LGR4 mRNA in MC3T3 cells transfected with miR-137 mimic was also
clearly downregulated compared to that of the scramble control, and
transfection with miR-137 inhibitor was also much higher than that
of the scramble control, establishing the downregulatory effect of
miR-137 on the LGR4 gene. The inhibitory effect of miR-137 was
stronger in U-2 cells.
Interaction between miR-137 and LGR4
protein
To further confirm the regulatory relationship
between miR-137 and LGR4 protein in different cell lines, we
transfected miR-137 mimic and inhibitor into cultured U-2 and MC3T3
cells. The results of western blot analysis (Fig. 5A) revealed a signifi-cantly
enhanced LGR4 protein level in U-2 cells transfected with miR-137
inhibitor and reduced LGR4 protein level in the miR-137 mimic
treatment group compared with the scramble controls. As shown in
Fig. 5B, the level of LGR4
protein in MC3T3 cells transfected with miR-137 mimic was clearly
downregulated in comparison with the scramble control, and that of
MC3T3 cells transfected with miR-137 inhibitor was much higher
compared with the scramble control, demonstrating the
downregulation of LGR4 gene by miR-137. The inhibitory effect of
miR-137 was stronger in U-2 cells.
Expression level of LGR4 mRNA and protein
varied in different groups
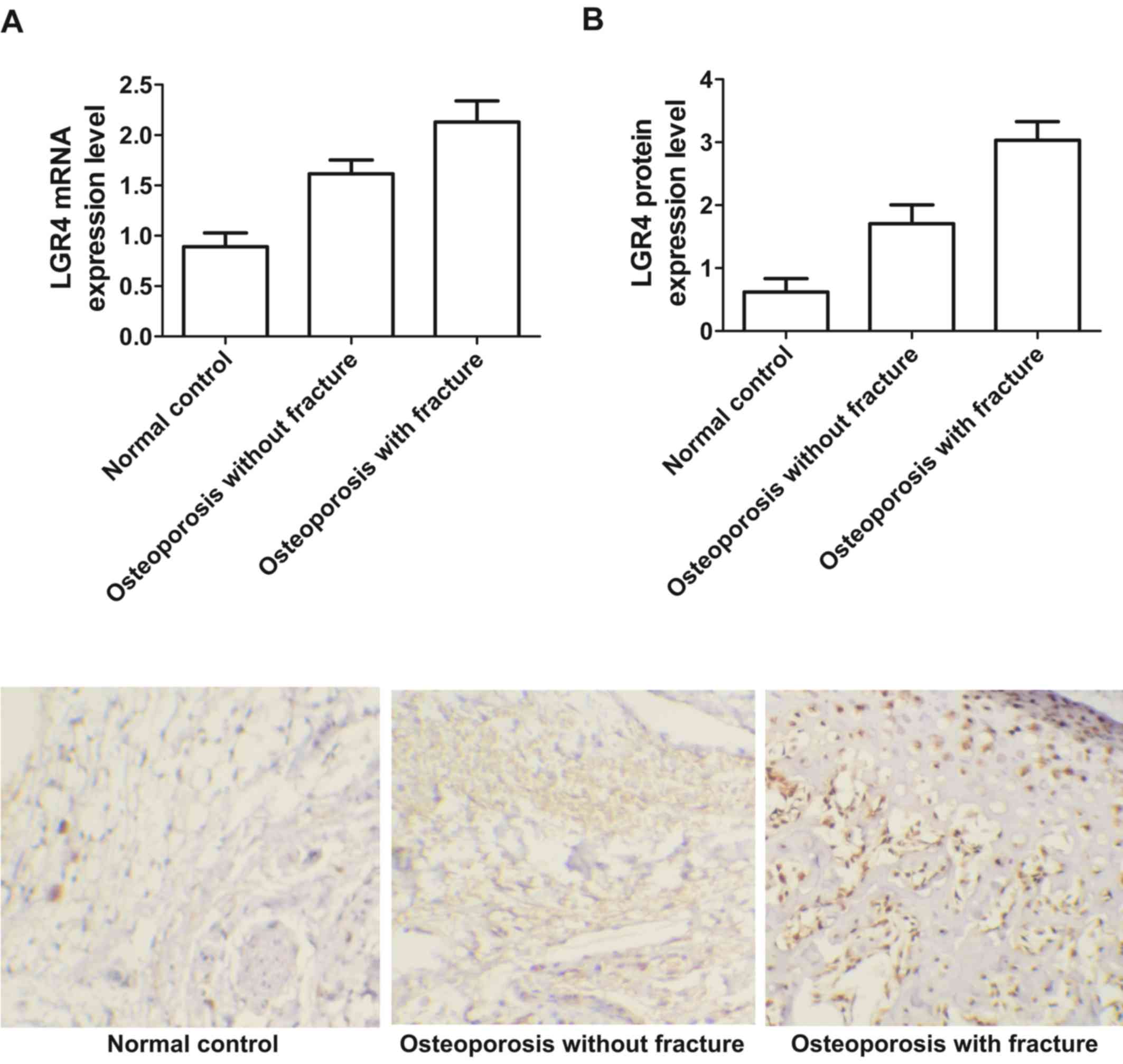
Real-time PCR and immunohistochemistry were used to
detect the expression level of LGR4 mRNA and protein among
osteoporosis patients both with and without fracture and a normal
group of patients. As shown in Fig.
6, the expression of LGR4 mRNA (Fig. 6A) and protein (Fig. 6B) were much lower in osteoporosis
patients without fracture and the normal group compared with
osteoporosis patients with fracture. The expression of LGR4 mRNA
(Fig. 6A) and protein (Fig. 6B) were significantly lower in the
normal group, suggesting that upregulation of LGR4 mRNA and protein
expression may contribute to the formation of osteoporosis and
fracture.
Expression level of ALP and ALP activity
varied in different groups
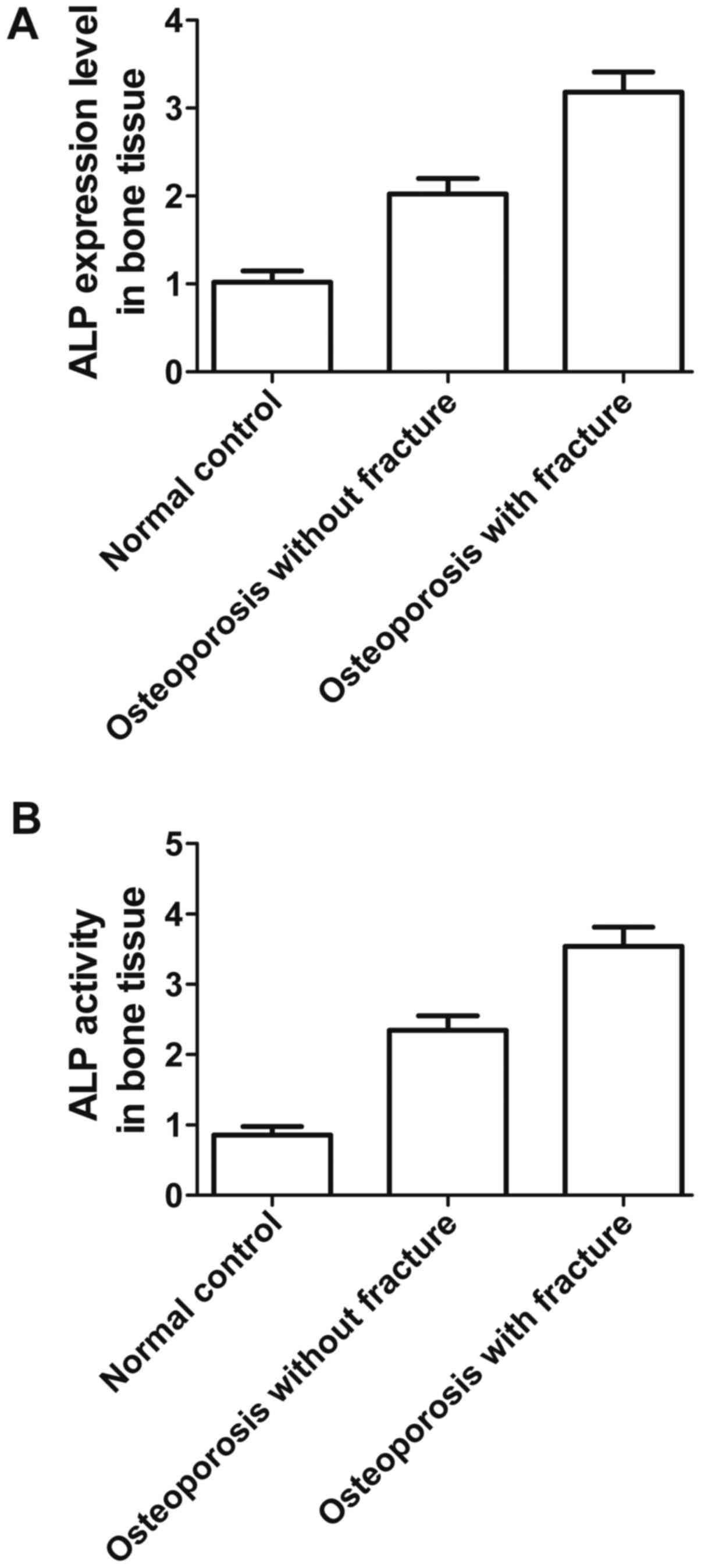
Finally, ELISA was used to measure the expression
level of ALP and to detect ALP activity in osteoporosis patients
with and without fracture, and the normal group. As shown in
Fig. 7, ALP expression (Fig. 7A) and ALP activity (Fig. 7B) in bone tissue were much higher
in osteoporosis patients both with and without fracture than the
normal group, while both ALP expression and ALP activity were
highest in osteoporosis patients with fractures, significantly so,
indicating that upregulation of ALP and ALP activity contributes to
the formation of osteoporosis and fracture.
Discussion
It has been shown in recent studies that miR-137
influences the process of tumorigenesis. For example, it was shown
that the sensitivity of pancreatic cancer cells to chemotherapy,
invasion and growth are modulated by miR-137 (19). It was also revealed that miR-137
inhibited proliferation of melanoma cells by targeting PAK2 and was
considerably downregulated in melanoma (20). Moreover, it was shown that miR-137
in colorectal cancer tissues was reduced and that colony formation,
tumorsphere and cell growth of colon cancer cells were inhibited by
miR-137 through targeting Musashi-1 (21). It was also found that miR-137 in
non-small cell lung cancer (NSCLC) tissues was downregulated and
that NSCLC cell proliferation and motility were inhibited by
miR-137 directly through targeting of SLC22A18 (22). We found that NSCLC cell migration,
invasion and proliferation could be inhibited by overexpression of
miR-137 through the targeting of BMP7. As an important signaling
molecule, bone morphogenetic protein (BMP) was first identified via
its ability to induce the formation of cartilage and bone (23). As the expression of miR-137 is
lost, so the ability of a cell to arrest at the G1 phase is
reduced, which increases its proliferation, and thus may result in
the accumulation of DNA damage which accordingly enhances genomic
instability (24). In this study,
we initially conducted real-time qPCR to estimate the expression of
miR-137 among osteoporosis patients with and without fracture, and
a normal group, and found that the expression was much lower in
osteoporosis patients in both groups compared to normal patients,
while the expression of miR-137 level was the lowest in the
fracture group.
As another potential contributor, miR-137 plays a
possible role in switching from proliferation to cellular
differentiation (25). It has
been reported that the expression level of miR-137 is reduced in
poorly differentiated gliomas and increased in neuronal
differentiation, although it is not currently known whether this
generalizes to other histology (26). Conversely, because of our ability
to detect such a relationship is limited by low statistical power,
no relationship has been discovered between miR-137 promoter
methylation and grade of tumor. Although it involves the control of
cell cycle, no significant relationship between miR-137 promoter
methylation and other squamous cell carcinoma of the head and neck
(SCCHN) prognostic factors, such as tumor size, stage, surgical
tumor margin positivity or nodal involvement has been found
(27). In this study, we searched
a miRNA online database (www.mirdb.org)
to find the target gene of miR-137, and found that LGR4 was a
virtual target of miR-137 with a potential binding site in the
3′UTR of LGR4. Furthermore, we performed a luciferase assay to
confirm that LGR4 is a direct shared target of miR-137, and found
that the luciferase activity of cells transfected with wild-type
LGR4 was significantly downregulated compared with mutant-type LGR4
and the scramble control group, while luciferase activity in cells
transfected with mutant 3′UTR of LGR4 was substantially
downregulated compared with the scramble control.
An orphan receptor can be encoded with the LGR4 gene
and it has been shown in genome-based studies that its genetic
features are associated with low BMD and osteoporotic fractures
(18). It has been reported that
local cytokines negatively regulate the BMP-induced increase in
expression of the LGR4 gene, which are important for bone formation
in addition to TGF beta and FGF (28,29). It was reported that for
osteoporotic patients an orphan receptor was encoded with the LGR4
gene which is identified as a genetic determinant for bone mass. In
osteoblastic cells, the effects of BMP on LGR4 expression were
examined. During culture, the LGR4 gene was expressed in the
osteoblastic cell line MC3T3E1 in a time-dependent manner. The
expression of LGR4 mRNA was partially enhanced by BMP treatment via
transcriptional events. While LGR4 mRNA was downregulated, the
BMP-induced increase in ALP mRNA and ALP activity was inhibited.
FGF suppressed BMP enhancement of LGR4 gene expression, but this
was reversed by dexamethasone. In primary cultures of calvarial
osteoblasts, the expression of LGR4 was also enhanced by BMP. It
can be inferred from these data that BMP regulates the LGR4 gene
which is required for BMP to influence osteoblastic differentiation
(17).
LGR4 is located on chromosome 11 at position
11p14-p13, which is ~107 kb in length. As one of the genes under
active investigation in our laboratory, LGR4 is also known as G
protein-coupled receptor 48 (Gpr48). LGR4 is widely expressed and
critical for the development of many organs including kidneys, the
reproductive tract, heart, cartilage, the nervous system and the
intestines (30). LGR4-knockout
mice demonstrated the importance of LGR4 on the development of
various organs such as the male reproductive tract, kidneys,
eyelids, erythropoiesis, hair placode, gallbladder and cystic duct,
and bones (31-36). A recent study revealed a variant
at position 126 caused by a c.376C<T nonsense mutation of human
LGR4 (37). This mutation
resulted in the complete loss of function of LGR4, and had
important association with risks for osteoporosis, resulting in low
BMD, gallbladder/biliary tract cancer, skin SCC and reduced
testosterone levels (18).
As an end-point clinical outcome of osteoporosis, OF
also has moderate heritability ranging from about 0.5 to 0.7
(8). We now know that GWASs and
their meta-analyses have identified over 20 genes/loci related to
risk of OF and more than 60 genes/loci related to variations in
BMD. Moreover, a rare, novel, nonsense mutation within the LGR4
gene strongly related to low BMD and OF was identified in a
recently published study involving whole-genome sequencing
(18). In the present study, we
performed real-time PCR and western blot analysis to evaluate LGR4
mRNA and protein in U-2 and MC3T3 cells in osteoporosis patients
with and without fracture and normal patients, and found that LGR4
mRNA and protein levels in both cell types were much higher when
transfected with miR-137 inhibitor, while LGR4 mRNA and protein
levels were much lower following transfection with miR-137 mimics
than the scramble control. Finally, we performed real-time PCR and
immunohistochemistry to measure LGR4 mRNA and protein expression,
and ALP expression and ALP activity in bone tissue in osteoporosis
patients with and without fractures, and the normal group, and
found that they were much higher in osteoporosis patients with and
without fractures than the normal group, while they were
significantly the lowest in osteoporosis patients with
fractures.
In conclusion, these data prove that overexpression
of miR-137 is associated with an altered risk of fracture in
patients with osteoporosis, and could be recognized as a biomarker
to predict risk of fracture in osteoporosis.
Acknowledgments
Not applicable.
Funding
No funding was received.
Availability of data and material
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XL planned the study, collected, analyzed and
interpreted the data and prepared the manuscript. XH planned the
study, collected the data, prepared the manuscript and analyzed the
literature. Both authors read and approved the final
manuscript.
Ethics approval and consent to
participate
The institution's Ethics and Research Committees
approved this study. The patients signed informed consent for
participation in the study.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Bruyère O, Cooper C, Pelletier JP, Maheu
E, Rannou F, Branco J, Luisa Brandi M, Kanis JA, Altman RD,
Hochberg MC, et al: A consensus statement on the European Society
for Clinical and Economic Aspects of Osteoporosis and
Osteoarthritis (ESCEO) algorithm for the management of knee
osteoarthritis - From evidence-based medicine to the real-life
setting. Semin Arthritis Rheum. 45(Suppl 4): S3–S11. 2016.
View Article : Google Scholar
|
|
2
|
Melton LJ III, Gabriel SE, Crowson CS,
Tosteson AN, Johnell O and Kanis JA: Cost-equivalence of different
osteoporotic fractures. Osteoporos Int. 14:383–388. 2003.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Adachi JD, Ioannidis G, Pickard L, Berger
C, Prior JC, Joseph L, Hanley DA, Olszynski WP, Murray TM,
Anastassiades T, et al: The association between osteoporotic
fractures and health-related quality of life as measured by the
Health Utilities Index in the Canadian Multicentre Osteoporosis
Study (CaMos). Osteoporos Int. 14:895–904. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Papaioannou A, Morin S, Cheung AM,
Atkinson S, Brown JP, Feldman S, Hanley DA, Hodsman A, Jamal SA,
Kaiser SM, et al Scientific Advisory Council of Osteoporosis
Canada: 2010 clinical practice guidelines for the diagnosis and
management of osteoporosis in Canada: summary. CMAJ. 182:1864–1873.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Orimo H, Nakamura T, Hosoi T, Iki M,
Uenishi K, Endo N, Ohta H, Shiraki M, Sugimoto T, Suzuki T, et al:
Japanese 2011 guidelines for prevention and treatment of
osteoporosis - executive summary. Arch Osteoporos. 7:3–20. 2012.
View Article : Google Scholar :
|
|
6
|
de Almeida Pereira Coutinho M, Bandeira E,
de Almeida JM, Godoi ET, Vasconcelos G and Bandeira F: Low bone
mass is associated with increased carotid intima media thickness in
men with type 2 diabetes mellitus. Clin Med Insights Endocrinol
Diabetes. 6:1–6. 2013.PubMed/NCBI
|
|
7
|
Ralston SH: Genetics of osteoporosis. Rev
Endocr Metab Disord. 2:13–21. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Deng HW, Mahaney MC, Williams JT, Li J,
Conway T, Davies KM, Li JL, Deng H and Recker RR: Relevance of the
genes for bone mass variation to susceptibility to osteoporotic
fractures and its implications to gene search for complex human
diseases. Genet Epidemiol. 22:12–25. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Landgraf P, Rusu M, Sheridan R, Sewer A,
Iovino N, Aravin A, Pfeffer S, Rice A, Kamphorst AO, Landthaler M,
et al: A mammalian microRNA expression atlas based on small RNA
library sequencing. Cell. 129:1401–1414. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Lee RC, Feinbaum RL and Ambros V: The C.
elegans heterochronic gene lin-4 encodes small RNAs with antisense
complementarity to lin-14. Cell. 75:843–854. 1993. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Kozomara A and Griffiths-Jones S: miRBase:
annotating high confidence microRNAs using deep sequencing data.
Nucleic Acids Res. 42(D1): D68–D73. 2014. View Article : Google Scholar :
|
|
12
|
Pauley KM and Cha S: miRNA-146a in
rheumatoid arthritis: a new therapeutic strategy. Immunotherapy.
3:829–831. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Cheng P, Chen C, He HB, Hu R, Zhou HD, Xie
H, Zhu W, Dai RC, Wu XP, Liao EY, et al: miR-148a regulates
osteoclastogenesis by targeting V-maf musculoaponeurotic
fibrosarcoma oncogene homolog B. J Bone Miner Res. 28:1180–1190.
2013. View Article : Google Scholar
|
|
14
|
Wang Y, Li L, Moore BT, Peng XH, Fang X,
Lappe JM, Recker RR and Xiao P: miR-133a in human circulating
monocytes: a potential biomarker associated with postmenopausal
osteoporosis. PLoS One. 7:e346412012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Seeliger C, Karpinski K, Haug AT, Vester
H, Schmitt A, Bauer JS and van Griensven M: Five freely circulating
miRNAs and bone tissue miRNAs are associated with osteoporotic
fractures. J Bone Miner Res. 29:1718–1728. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Garmilla-Ezquerra P, Sañudo C,
Delgado-Calle J, Pérez- Nuñez MI, Sumillera M and Riancho JA:
Analysis of the bone microRNome in osteoporotic fractures. Calcif
Tissue Int. 96:30–37. 2015. View Article : Google Scholar
|
|
17
|
Pawaputanon Na Mahasarakham C, Ezura Y,
Kawasaki M, Smriti A, Moriya S, Yamada T, Izu Y, Nifuji A,
Nishimori K, Izumi Y, et al: BMP-2 enhances Lgr4 gene expression in
osteoblastic cells. J Cell Physiol. 231:887–895. 2016. View Article : Google Scholar
|
|
18
|
Styrkarsdottir U, Thorleifsson G, Sulem P,
Gudbjartsson DF, Sigurdsson A, Jonasdottir A, Jonasdottir A,
Oddsson A, Helgason A, Magnusson OT, et al: Nonsense mutation in
the LGR4 gene is associated with several human diseases and other
traits. Nature. 497:517–520. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Xiao J, Peng F, Yu C, Wang M, Li X, Li Z,
Jiang J and Sun C: microRNA-137 modulates pancreatic cancer cells
tumor growth, invasion and sensitivity to chemotherapy. Int J Clin
Exp Pathol. 7:7442–7450. 2014.
|
|
20
|
Hao S, Luo C, Abukiwan A, Wang G, He J,
Huang L, Weber CE, Lv N, Xiao X, Eichmüller SB, et al: miR-137
inhibits proliferation of melanoma cells by targeting PAK2. Exp
Dermatol. 24:947–952. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Smith AR, Marquez RT, Tsao WC, Pathak S,
Roy A, Ping J, Wilkerson B, Lan L, Meng W, Neufeld KL, et al: Tumor
suppressive microRNA-137 negatively regulates Musashi-1 and
colorectal cancer progression. Oncotarget. 6:12558–12573. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Zhang B, Liu T, Wu T, Wang Z, Rao Z and
Gao J: MicroRNA-137 functions as a tumor suppressor in human
non-small cell lung cancer by targeting SLC22A18. Int J Biol
Macromol. 74:111–118. 2015. View Article : Google Scholar
|
|
23
|
Cheng H, Jiang W, Phillips FM, Haydon RC,
Peng Y, Zhou L, Luu HH, An N, Breyer B, Vanichakarn P, et al:
Osteogenic activity of the fourteen types of human bone
morphogenetic proteins (BMPs). J Bone Joint Surg Am. 85:1544–1552.
2003. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Kozaki K, Imoto I, Mogi S, Omura K and
Inazawa J: Exploration of tumor-suppressive microRNAs silenced by
DNA hypermethylation in oral cancer. Cancer Res. 68:2094–2105.
2008. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Tarantino C, Paolella G, Cozzuto L,
Minopoli G, Pastore L, Parisi S and Russo T: miRNA 34a, 100, and
137 modulate differentiation of mouse embryonic stem cells. FASEB
J. 24:3255–3263. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Silber J, Lim DA, Petritsch C, Persson AI,
Maunakea AK, Yu M, Vandenberg SR, Ginzinger DG, James CD, Costello
JF, et al: miR-124 and miR-137 inhibit proliferation of
glioblastoma multiforme cells and induce differentiation of brain
tumor stem cells. BMC Med. 6:142008. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Langevin SM, Stone RA, Bunker CH,
Lyons-Weiler MA, LaFramboise WA, Kelly L, Seethala RR, Grandis JR,
Sobol RW and Taioli E: MicroRNA-137 promoter methylation is
associated with poorer overall survival in patients with squamous
cell carcinoma of the head and neck. Cancer. 117:1454–1462. 2011.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Zhen G and Cao X: Targeting TGFβ signaling
in subchondral bone and articular cartilage homeostasis. Trends
Pharmacol Sci. 35:227–236. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Itkin T, Kaufmann KB, Gur-Cohen S, Ludin A
and Lapidot T: Fibroblast growth factor signaling promotes
physiological bone remodeling and stem cell self-renewal. Curr Opin
Hematol. 20:237–244. 2013.PubMed/NCBI
|
|
30
|
Du B, Luo W, Li R, Tan B, Han H, Lu X, Li
D, Qian M, Zhang D, Zhao Y, et al: Lgr4/Gpr48 negatively regulates
TLR2/4-associated pattern recognition and innate immunity by
targeting CD14 expression. J Biol Chem. 288:15131–15141. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Mendive F, Laurent P, Van Schoore G,
Skarnes W, Pochet R and Vassart G: Defective postnatal development
of the male reproductive tract in LGR4 knockout mice. Dev Biol.
290:421–434. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Kato S, Matsubara M, Matsuo T, Mohri Y,
Kazama I, Hatano R, Umezawa A and Nishimori K: Leucine-rich
repeat-containing G protein-coupled receptor-4 (LGR4, Gpr48) is
essential for renal development in mice. Nephron Exp Nephrol.
104:e63–e75. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Kato S, Mohri Y, Matsuo T, Ogawa E,
Umezawa A, Okuyama R and Nishimori K: Eye-open at birth phenotype
with reduced keratinocyte motility in LGR4 null mice. FEBS Lett.
581:4685–4690. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Song H, Luo J, Luo W, Weng J, Wang Z, Li
B, Li D and Liu M: Inactivation of G-protein-coupled receptor 48
(Gpr48/Lgr4) impairs definitive erythropoiesis at midgestation
through down-regulation of the ATF4 signaling pathway. J Biol Chem.
283:36687–36697. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Mohri Y, Kato S, Umezawa A, Okuyama R and
Nishimori K: Impaired hair placode formation with reduced
expression of hair follicle-related genes in mice lacking Lgr4. Dev
Dyn. 237:2235–2242. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yamashita R, Takegawa Y, Sakumoto M,
Nakahara M, Kawazu H, Hoshii T, Araki K, Yokouchi Y and Yamamura K:
Defective development of the gall bladder and cystic duct in Lgr4-
hypomorphic mice. Dev Dyn. 238:993–1000. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Luo J, Zhou W, Zhou X, Li D, Weng J, Yi Z,
Cho SG, Li C, Yi T, Wu X, et al: Regulation of bone formation and
remo-deling by G-protein-coupled receptor 48. Development.
136:2747–2756. 2009. View Article : Google Scholar : PubMed/NCBI
|