Introduction
Lumbar disc herniation (LDH) is induced by
mechanical pressure and inflammation in animal models, and displays
the common symptom of radicular pain (1,2).
Inflammatory cytokines concentrate and produce associated pain in
the LDH animal model (3).
Researchers have demonstrated that inflammatory cytokines,
including interleukin (IL)-1 and tumor necrosis factor (TNF)-α, are
closely associated with the impact of nucleus pulposus on nerve
roots in animal models (1). Other
studies have revealed that the affected nerve roots are strongly
associated with high levels of TNF-α in the tissues of patients
with LDH (4,5). Patients with chronic low back pain
that is caused by herniated discs have been reported to have high
expression levels of TNF-α (6).
Additionally, it has been proved that inflammatory cytokines,
including IL-1, IL-6 and TNF-α are detectable in 77 cases of LDH
patients (7). Therefore, the
present study established an inflammatory cytokine-induced LDH
model in rats, and these were used for the current
investigation.
The Wnt1 gene was identified by Nusse and Varmus in
1982, and it acts as an integration site for the mouse mammary
tumor virus in virally-induced mammary tumors (8). Wnt signaling, mediated through
β-Catenin, is identified as a critical regulator in many diseases
(9,10). The Wnt/β-Catenin pathway has been
reported to participate in the progression of multiple types of
cancer, such as ovarian cancer (11), gastric cancer (12), and breast cancer (13). Recently, researchers demonstrated
that the Wnt/β-Catenin pathway participated in the development of
neuropathic pain (14). Other
studies have also reported that the Wnt/β-Catenin pathway is
closely associated with intervertebral disc development (15,16). However, limited knowledge on the
roles of Wnt/β-Catenin pathway in inflammatory cytokine-induced LDH
animal model exists to date.
Cartilage-derived morphogenetic protein-1 (CDMP1),
also known as growth differentiation factor-5 (GDF-5), is a type of
polypeptide growth factor with specific cartilage inducibility.
CDMP1 is expressed in the developmental long bone cartilage core
and in the pre-cartilage matrix in the embryonic stage (17). A previous study revealed that
CDMP1 contributed to the structural and functional maintenance of
the intervertebral disc (18).
Recently, another study suggested that in human intervertebral
annulus cells, CDMP1 could be modulated by IL-1β and TNF-α
(19). Furthermore, researchers
have demonstrated that among Northern European women, CDMP1
expression is association with lumbar disc degeneration (20). Thus, the current study aimed to
investigate the roles and mechanisms of CDMP1 on inflammatory
cytokine-induced LDH animal model.
In the present study, the expression levels of CDMP1
were analyzed in inflammatory cytokine-induced DRG tissues derived
from LDH model rats. In addition, the effects and mechanisms of
CDMP1 overexpression were examined on the Wnt/β-Catenin pathway and
on the apoptosis of inflammatory cytokine-induced DRG cells.
Materials and methods
Animal model
A total of 40 healthy adult male Sprague Dawley (SD)
rats (weight, 110–130 g; age, 5–6 weeks) were obtained from
Guangdong Medical Laboratory Animal Center (Foshan, China), and
kept in cages (22± 6°C and 50±13% humidity) with a 12 h dark/light
cycle. The rats had free access to food and water. The SD rats were
randomly divided into two groups with 20 rats in each group. One
group was left untreated (control/normal group) and the other group
underwent an operation to generate the LDH model (LDH group). The
LDH model was established as follows: The rats in the LDH group
were subjected to abdominal anesthesia (10% Chloral hydrate; 0.35
ml/100 g; XiLongScientific, Shenzhen, China). After being
completely anesthetized, the rats lumbar spine was placed in a
flexion position. A lumbar puncture needle no.9 was used at
L4-L5 spinous process intervals for epidural
puncture. Then 20 µl rat autologous nucleus pulposus
suspension and 30 µl lidocaine (2%; XiLongScientific) were
used for epidural injection. With the effect of anesthesia
decreasing on the rats, the limbs activity of the rats was
observed. The rats’ forelimbs could move normally, however, a loss
of feeling and motion was observed in the hind legs, which was
restored to normal in an additional 30 min. This phenomenon
suggested that the epidural cavity puncture and injection was
successful. The hind legs of the rats (gait or pronation) were
therefore observed in order to determine whether there injury on
the spinal cord was caused in the molding process. If there was no
lower limbs paralysis, heat stimulation for pain-sensitive
detection would then be performed on the lower limbs of rats at 3
days post-operation. If opioid-induced hyperalgesia occurred in
rats, the model establishment for LDH rats was seen as successful.
The experimental protocols involving animals were approved by the
Shanxi Provincial People’s Hospital Ethics Committee (Taiyuan,
China).
Hematoxylin-eosin (HE) staining
DRG tissues were obtained from control and LDH rats.
They were frozen at −80°C and cut into 5-µm-thick sections.
Frozen slices of DRG tissues were dried at room temperature. Slices
were then placed into hematoxylin dye liquor (SolarBio Science
& Technology Co., Ltd., Beijing, China) for 10 min. The slices
were placed into 10% glacial acetic acid for 10 sec and then into
1% ammonia water until they turned blue. Next, the slices were
placed into eosin dye liquor (SolarBio Science & Technology
Co., Ltd.) for 10 sec and gradual alcohol dehydration was performed
at 70, 90, 95 and 100% for 2 min. Afterwards, the slices were
placed into xylene and maintained for 2 min, and this step was
repeated once. Finally, the slices were sealed by neutral gum.
Immunohistochemistry
Prior to the histology analysis, rat DRG tissues
were put into reaction with 4% paraformaldehyde and 30% sucrose
solution for perfusion. Frozen slices of DRG tissues were then
treated with primary antibodies against CDMP1 (1:500; cat. no.
ab93855; Abcam, Cambridge, UK) at 4°C for 1 h, and blocked with
blocking reagent (PBS-Triton X-100 and 0.5% bovine serum albumin
(BSA; Gibco; Thermo Fisher Scientific, Inc., Waltham, MA, USA) at
room temperature for 60 min. The tissue slices were then incubated
with horseradish peroxidase-conjugated secondary antibodies (goat
anti-rabbit; 1:1,000; cat. no. ab150077; Abcam) at room temperature
for 90 min. The tissue samples were observed by fluorescence
microscope (magnification, ×100) after being treated with
diaminobenzidine (Sigma-Aldrich; Merck KGaA, Darmstadt,
Germany).
Cell culture
The DRG tissues were dissected and incubated with
D-Hanks solution (Beyotime Institute of Biotechnology, Shanghai,
China). The mixture was digested with 0.25% trypsin (Beyotime
Institute of Biotechnology) at 37°C for 20 min. A pipette was used
to mix 20 ml Dulbecco’s modified Eagle’s medium with Ham’s F-12
(DMEM/F12; Gibco; Thermo Fisher Scientific, Inc.) in a proportion
of 1:1, and fetal bovine serum (FBS; Sigma-Aldrich; Merck KGaA) was
then added into the mixture to stop the digestion process. Then,
the cell suspension was collected. The suspension was centrifuged
at 224 × g for 15 min, and the supernatant was discarded. A total
of 10 ml DMEM/F12 was added into the precipitate. The cell debris
and residual tissues were sunk, and the cell suspension was
collected. Cells were seeded into a polylysine pre-coated culture
flask (75 cm2, 250 ml) and placed in an incubator with
5% CO2 at 37°C. After 48 h, the cells that attached on
the pre-coated flask were rat DRG neurons.
Cell transfection
The CDMP1 coding sequence (GTTCAAGAGACCCCCGTCTG) was
cloned into the pcDNA3.1(+) empty vector (Invitrogen; Thermo Fisher
Scientific, Inc.). The rat DRG neurons were transfected with 50 nM
pcDNA3.1(+) empty vector or pcDNA3.1-CDMP1 expressing vector using
Lipofectamine® 2000 (Thermo Fisher Scientific, Inc.) for
48 h at 37°C.
Experimental groups
To conduct animal experiments, two treatment groups,
the control group (normal rats) and the LDH group (LDH model rats),
were established in the present study.
For in vitro experiments, eight treatment
groups were prepared, as follows: Control group (DRG cells treated
with 0.1% PBS), NC group (DRG cells transfected with pcDNA3.1 empty
vector), IL-1β group (DRG cells treated with 10 ng/ml IL-1β),
IL-1β+NC group (DRG cells transfected with pcDNA3.1 empty vector
and treated with 10 ng/ml IL-1β), IL-1β+CDMP1 group (DRG cells
transfected with pcDNA3.1-CDMP1 plasmid and treated with 10 ng/ml
IL-1β), TNF-α group (DRG cells treated with 50 ng/ml TNF-α),
TNF-α+NC group (DRG cells transfected with pcDNA3.1 empty vector
and treated with 50 ng/ml TNF-α), and TNF-α+CDMP1 group (DRG cells
transfected with pcDNA3.1-CDMP1 plasmid and treated with 50 ng/ml
TNF-α).
Cell viability analysis
Cell Counting Kit-8 (CCK-8; Beyotime Institute of
Biotechnology) assay was performed to detect cell viability.
Approximately 6×104 cells/ml of DRG neurons were seeded
into 96-well plates and maintained at 37°C and 5% CO2
for 12 h. The cells were treated as indicated. Following treatment,
cells were maintained in the incubator (37°C, 5% CO2)
for 24, 48 and 72 h. Afterwards, 10 µl CCK-8 reagent was
added into each well for 3 h. A microplate reader (Bio-Rad
Laboratories, Inc., Hercules, CA, USA) was used to read the
absorbance at 450 nm. Cell viability was determined as the
proportion of cell survival compared with control.
ELISA
The contents of IL-1β and TNF-α were detected using
IL-1β (cat. no. ml027836) and TNF-α (cat. no. ml0257452) ELISA kits
(Shanghai Enzyme-linked Biotechnology Co., Ltd., Shanghai, China)
according to the manufacturer’s protocol. DRG cells were seeded
into the corresponding wells and maintained at 37°C for 90 min.
Apart from the blank wells, 100 µl biotinylated antibody
fluids were added into each well. The wells were sealed with
adhesive tape and maintained at 37°C for 60 min. PBS was used to
wash the plates, and then 100 µl enzyme solution was added
for 30 min. Next, chromogenic substrate was added into the wells,
excluding the blank wells, for an additional 10–15 min in the dark
at 37°C. Afterwards, stop solution was added into the plates, and
mixed for 10 min. Finally, the absorbance at 450 nm was measured
using a microplate reader (Bio-Rad Laboratories, Inc.).
TdT-mediated dUTP nick-end labeling
(TUNEL) staining assay
Cell apoptosis was measured using a TUNEL assay
staining kit (Roche Diagnostics, Basel, Switzerland) according to
the manufacturer’s protocol. Briefly, the cell suspension was
dripped onto the slide and dried. TdT enzyme reaction liquid was
added to the slide at room temperature for 2–5 min. The slide was
immersed in the washing and stopping reaction buffer which had been
preheated to 37°C. Subsequently, 0.05% DAB was dripped onto the
slide at room temperature for 3–6 min. The slide was then observed
under a fluorescence microscope (magnification, ×100).
Flow cytometry
Flow cytometry was also performed to determine cell
apoptosis. DRG cells were trypsinized with 0.25% trypsin (Beyotime
Institute of Biotechnology), after being washed with PBS. Cells
were then centrifuged at 224 × g for 1 min, and the supernatant was
discarded. Afterwards, cells were suspended in the incubation
buffer at a density of 1×106 cells/ml. Annexin
V-phycoerythrin (PE) and 7-Aminoactinomycin D (7-AAD)
(Sigma-Aldrich; Merck KGaA) or in the dark at room temperature for
15 min. Cell apoptosis was measured by flow cytometry using a
FACSCalibur (BD Biosciences, San Jose, CA, USA). CellQuest software
version 5.1 (BD Biosciences) was used for analysis.
Western blot analysis
Total protein was extracted from tissues or cultured
cells using a radioimmunoprecipitation lysis buffer (SolarBio
Science & Technology Co., Ltd.). The proteins were quantified
using a bicinchoninic acid assay protein quantification kit
(Shanghai Yeasen Biotechnology Co., Ltd., Shanghai, China). A total
of 3 µg protein was loaded per lane and separated by 12%
SDS-PAGE and transferred onto a polyvinylidene fluoride membrane
(Millipore, Billerica, MA, USA). The membrane was blocked using
Tris-buffered saline and Tween-20 solution containing 5% skimmed
milk powder at 37°C for 1 h. The immunoblotting was performed with
the following antibodies at 4°C for 24 h: Anti-CDMP1 (1:1,000; cat.
no. ab93855; Abcam), anti-IL-1β (1:800; cat. no. ab200478; Abcam),
anti-TNF-α (1:1,000; cat. no. ab6671; Abcam), anti-Caspase-3
(1:500; cat. no. ab13847; Abcam), anti-Caspase-8 (1:1,000; cat. no.
ab25901; Abcam), anti-Caspase-9 (1:1,000; cat. no. ab25758; Abcam),
anti-BCL2 associated X (Bax; 1:1,000; cat. no. ab32503; Abcam),
anti-BCL2 apoptosis regulator (Bcl-2; 1:1,000; cat. no. ab59348;
Abcam), anti-Wnt1 (1:1,000; cat. no. ab85060; Abcam),
anti-β-Catenin (1:5,000; cat. no. ab32572; Abcam), anti-Histone
(1:1,000; cat. no. ab1791; Abcam), and anti-β-actin (1:1,000; cat.
no. ab8227; Abcam). Horseradish peroxidase-conjugated secondary
antibodies (1:1,000; cat. no. bs-0293M; BIOSS, Beijing, China) were
used at room temperature for 1 h. Enhanced chemiluminescent
reagents (ECL; Millipore, Billerica, MA, USA) were used for
developing the protein signals on an ECL system (Amersham; GE
Healthcare, Chicago, IL, USA). The quantity one v4.6.2 software
(Bio-Rad Laboratories, Inc.) was used to analyze the protein
signals.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA was extracted from tissues or cultured
cells with TRIzol reagent (Thermo Fisher Scientific, Inc.). RNA was
reverse transcribed to cDNA with the Reverse Transcription kit
(Sigma-Aldrich; Merck KGaA), according to the manufacturer’s
protocol. SYBR Green I (Thermo Fisher Scientific, Inc.) was used
for qPCR. qPCR was conducted on a ABI 7500 Thermocycler (Applied
Biosystems; Thermo Fisher Scientific, Inc.) with the following
thermocycling conditions: Pretreatment at 95°C for 10 min, then 45
cycles of 94°C for 15 sec, 62°C for 45 sec, followed by one cycle
of 94°C for 15 sec, 62°C for 1 min, 95°C for 15 sec, and a final
extension step at 75°C for 10 min and hold at 4°C. The primers were
purchased from Invitrogen (Thermo Fisher Scientific, Inc.) and the
sequences were as follows: CDMP1, forward
5′-GTTCAAGAGACCCCCGTCTG-3′ and reverse 5′-GGAAGGGATGGCACTCGTAG-3′
(product, 195 bp); IL-1β, forward 5′-GGCTTCCTTGTGCAAGTGTC-3′ and
reverse 5′-CACACACTAGCAGGTCGTCA-3′ (product, 376 bp); TNF-α,
forward 5′-CGGAAAGGACACCATGAGCA-3′ and reverse
5′-GGGAGCCCATTTGGGAACTT-3′ (product, 219 bp); Caspase-3, forward
5′-ACCCTGAAATGGGCTTGTGT-3′ and reverse 5′-TTTTCAGGTCCACAGGTCCG-3′
(product, 291 bp); Caspase-8, forward 5′-GTGCCTGATGAGACAGGCTT-3′
and reverse 5′-AGTTCACGCCAGTCAGGATG-3′ (product, 235 bp);
Caspase-9, forward 5′-CCAGCTACCCGAAGACCAAG-3′ and reverse
5′-GAGGGGGCCGAGTACTATCT-3′ (product, 235 bp); Bax, forward
5′-GGCGAATTGGCGATGAACTG-3′ and reverse 5′-ATGGTTCTGATCAGCTCGGG-3′
(product, 217 bp); Bcl-2, forward 5′-TTCTTTCCCCGGAAGGATGG-3′ and
reverse 5′-AGTATCCCACTCGTAGCCCC-3′ (product, 112 bp); β-actin,
forward 5′-ACCCGCGAGTACAACCTTCT-3′ and reverse
5′-AGGGTCAGGATGCCTCTCTT-3′ (product, 263 bp). The 2−ΔΔCq
method was performed to assess the gene expression (21).
Statistical analysis
The statistical analysis was analyzed using IBM SPSS
software version 20 (IBM Corp., Armonk, NY, USA). Results are
presented as mean ± standard error of the mean from at least 3
independent experiments. All of the experimental data were analyzed
by one-way analysis of variance with the Tukey’s post hoc test.
P<0.05 was considered to indicate a statistically significant
difference.
Results
Characterization of DRG tissues and
cells
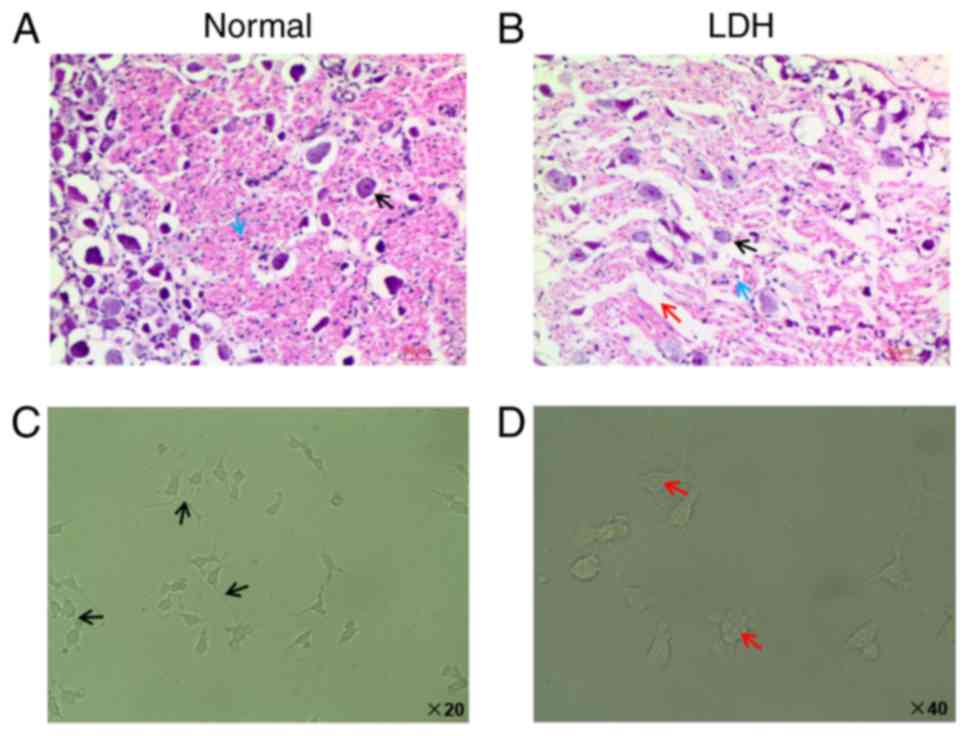
DRG tissues were identified and prepared from normal
rats as well as from LDH model rats. Based on H&E staining
results, the nerve cells in tissue samples from normal rats were
round, homogeneous, and the nucleolus was clear (indicated by the
black arrow in Fig. 1A). In
addition, the cytoplasm was filled with granular Nissl bodies
(indicated by the blue arrow in Fig.
1A). However, in tissue samples from LDH model rats, H&E
staining results revealed shrinking cells, blurry nuclei, a
decrease of the Nissl bodies and widening of the cell gaps
(indicated by the black, blue and red arrows in Fig. 1B, respectively).
Next, DRG cells were harvested from the DRG tissues
of normal rats. During the experiments, the DRG cells were cultured
in DMEM/F12 medium containing 10% FBS. Following 7 days in cell
culture, DRG cells were observed using an inverted microscope. The
microscopy analysis demonstrated that the cell body of the neurons
was gradually enlarged and oval. The neural network of DRG cells
was dense, as indicated by the black arrows in Fig. 1C. The size of DRG cells varied,
and most DRG cells were of small size (Fig. 1C). The cell nucleus of DRG cells
was transparent, and the nucleolus was clear, as indicated by the
red arrows in Fig. 1D. The
formation of the nucleolus was from spherical to multiform and
subsequently became relatively stable.
CDMP1 expression is downregulated and
inflammatory cytokine expression upregulated in DRG tissues from
LDH model rats
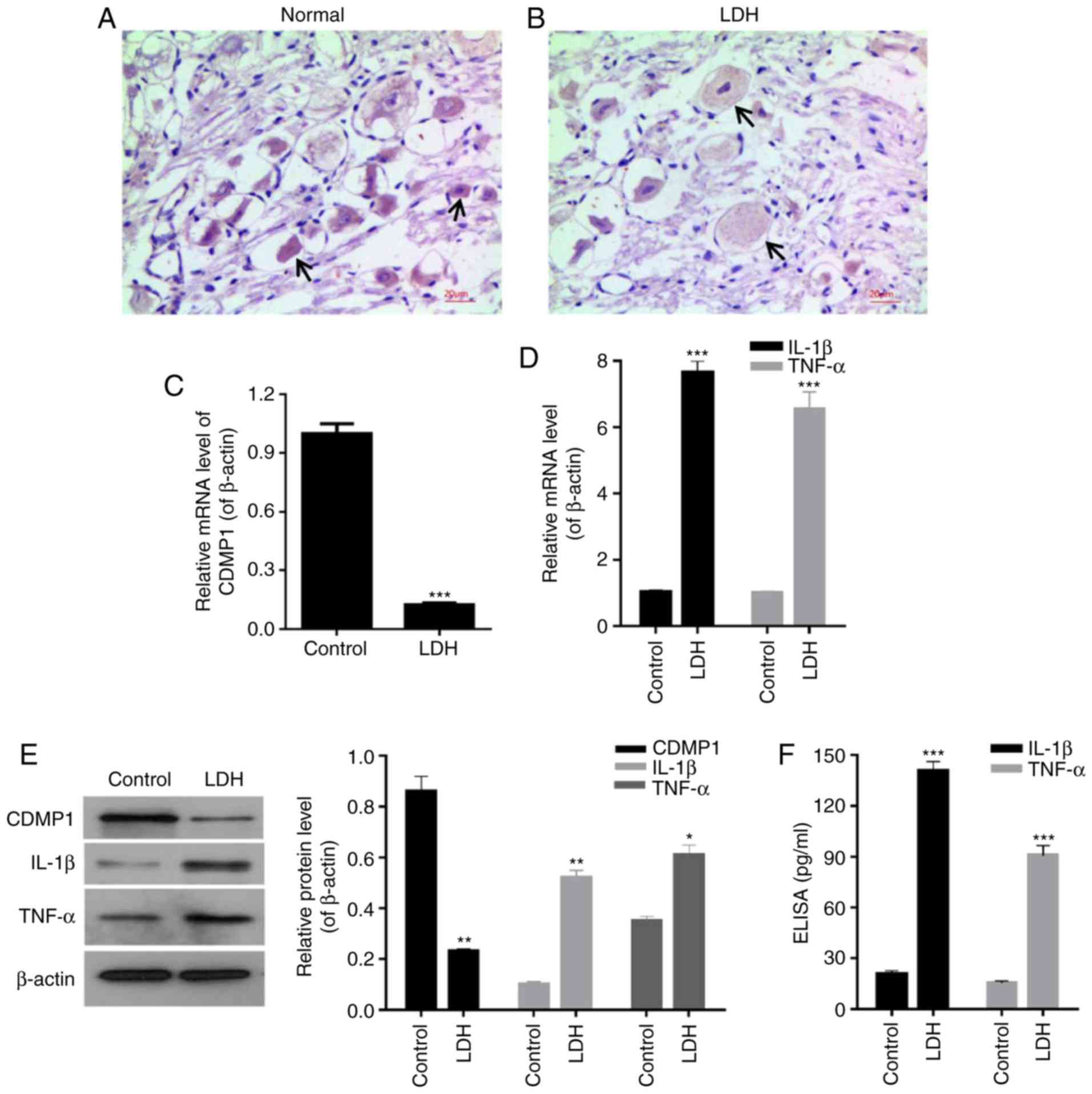
Immunohistochemistry staining results for CDMP1
indicated that, in DRG tissues from LDH model rats, the staining of
cells was lighter compared with the DRG tissues from normal rats
(indicated by the black arrows in Fig. 2A and B), and that the positive
expression rates of CDMP1 in the normal group were evidently higher
compared with the LDH group (Fig. 2A
and B). RT-qPCR and western blot analyses were used to further
determine the CDMP1 expression in DRG tissues, at the mRNA and the
protein level respectively. The results demonstrated significantly
lower expression levels of CDMP1 in DRG tissues from LDH model rats
compared with normal rats (Fig. 2C
and E; P<0.001). In addition, IL-1β and TNF-α expression
were measured by RT-qPCR, western blot and ELISA assays. All the
resulting data revealed that the levels of IL-1β and TNF-α were
upregulated in tissues from the LDH group, compared with the
control group (Fig. 2D–F;
P<0.001). The present findings demonstrated that CDMP1
expression was downregulated, while inflammatory cytokine IL-1β and
TNF-α expression was upregulated in DRG tissues from LDH model
rats.
CDMP1 overexpression enhances the cell
viability of inflammatory cytokine-induced DRG cells
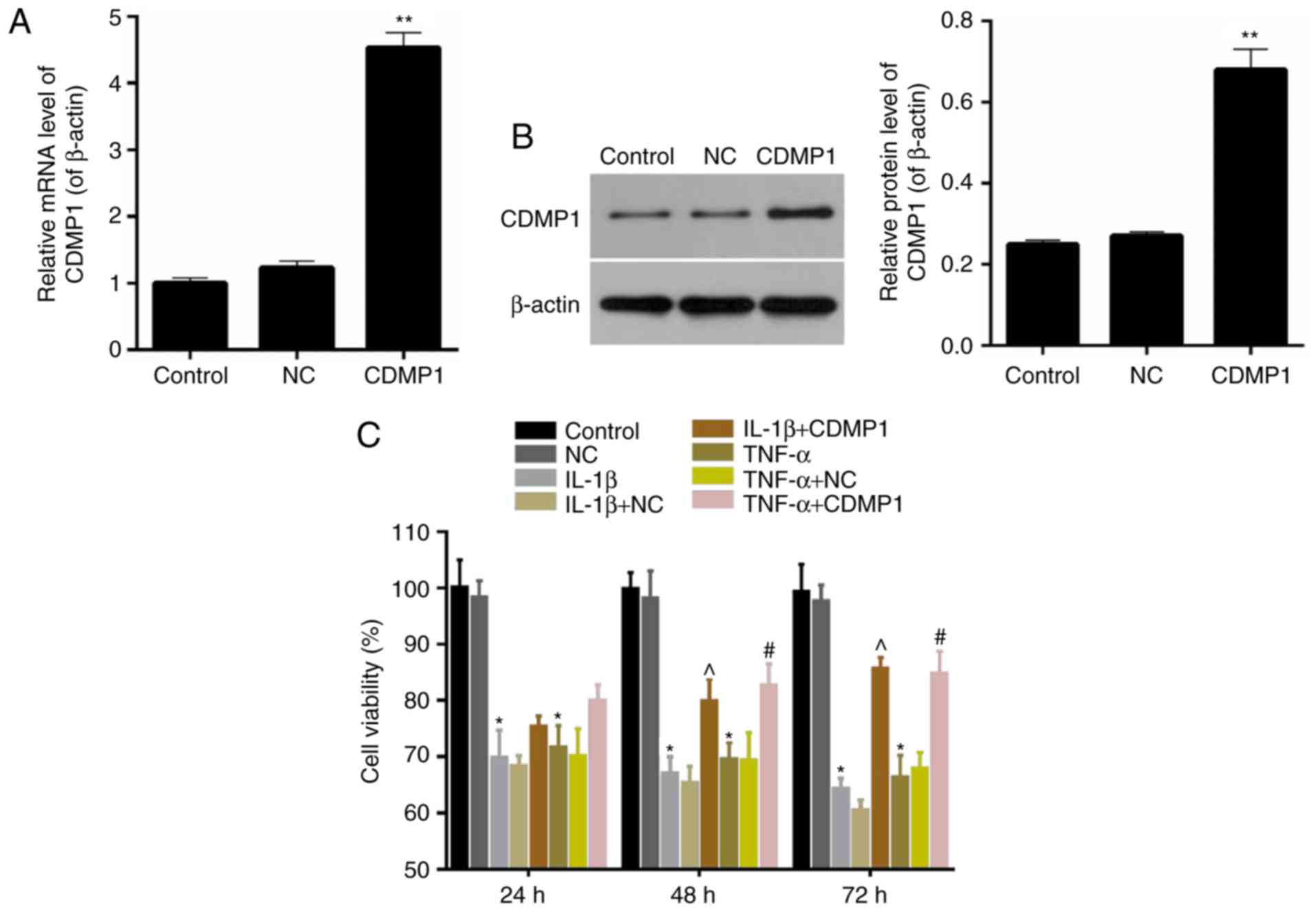
The expression levels of CDMP1 were assessed in DRG
cells that were transfected with empty vector or with a
CDMP1-overexpressing vector. Results from RT-qPCR and western blot
analyses indicated that CDMP1 expression was significantly enhanced
in the CDMP1-tranfected DRG cells compared with empty
vector-transfected DRG cells (Fig. 3A
and B). Thus, DRG cells with CDMP1 overexpression were
successfully generated by transfection with CDMP1-expressing
vector. CCK-8 assay results demonstrated that the cell viability of
inflammatory cytokine-induced DRG cells with CDMP1-expressing
vector was significantly higher compared with inflammatory
cytokine-induced DRG cells that were transfected with empty vector
(Fig. 3C; P<0.05). In
addition, this effect on cell viability was markedly more obvious
at 72 h of the treatment compared with 24 or 48 h. These results
indicated that CDMP1 overexpression enhanced the cell viability of
inflammatory cytokine-induced DRG cells.
CDMP1 overexpression suppresses the
apoptosis of inflammatory cytokine-induced DRG cells through
regulating the expression of apoptosis-associated proteins
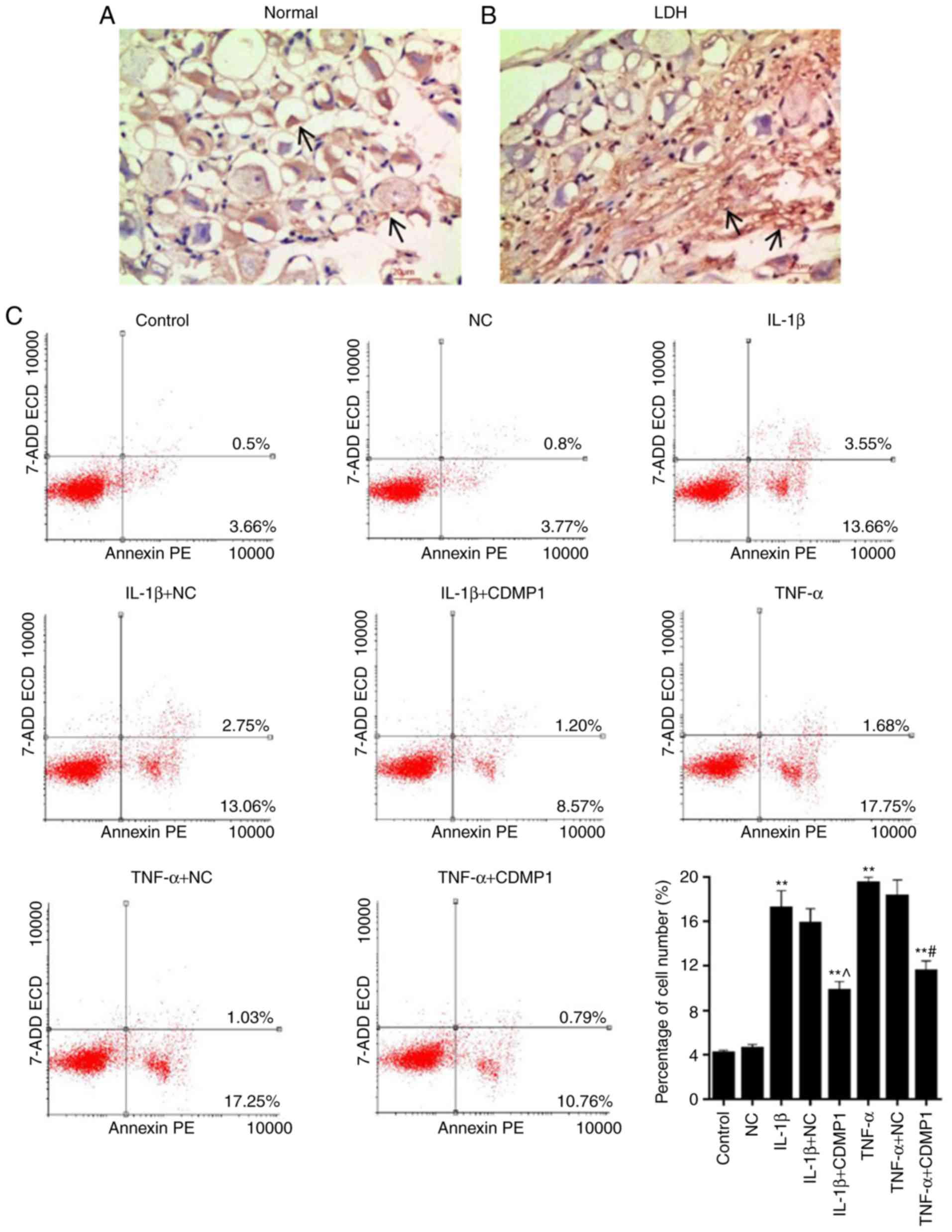
Results from the TUNEL assay revealed that the
number of positive cells in DRG tissues from LDH model rats was
obviously higher compared with normal DRG tissue samples (indicated
by the black arrows in Fig. 4A and
B). This finding suggested that apoptosis of DRG cells was
enhanced in DRG tissues derived from LDH model rats. Cell apoptosis
of DRG cells was further assessed in the present study by flow
cytometry. The results demonstrated that the proportion of
apoptotic DRG cells were as follows: 4.16% (Control group), 4.57%
(NC group), 17.21% (IL-1β group), 15.81% (IL-1β+NC group), 9.77%
(IL-1β+CDMP1 group), 19.43% (TNF-α group), 18.28% (TNF-α+NC group),
and 11.55% (TNF-α+CDMP1 group; Fig.
4C). The proportion of apoptotic DRG cells in the IL-1β+CDMP1
and TNF-α+CDMP1 groups were significantly lower compared with the
IL-1β+NC and TNF-α+NC groups. This finding indicated that CDMP1
overexpression suppressed the apoptosis of inflammatory
cytokine-induced DRG cells.
Next, the apoptosis-related mechanism was
investigated in DRG cells. The mRNA and protein levels of
apoptosis-associated factors Caspase-3/8/9, Bax and Bcl-2 were
respectively evaluated by RT-qPCR and western blotting. The RT-qPCR
data revealed that the Caspase-3/9 and Bax expression in the
IL-1β+CDMP1 and TNF-α+CDMP1 groups were significantly downregulated
compared with the IL-1β+NC and TNF-α+NC groups (Fig. 5A). However, CDMP1 overexpression
reduced the expression levels of Caspase-8 in IL-1β-induced DRG
cells, while it enhanced the Caspase-8 expression in TNF-α-induced
DRG cells (Fig. 5A). CDMP1
overexpression also resulted in high expression levels of Bcl-2 in
IL-1β-induced DRG cells and low Bcl-2 expression in TNF-α-induced
DRG cells (Fig. 5A; P<0.05).
Western blot analysis displayed similar results for the protein
expression levels of apoptosis-associated proteins in DRG cells
from each group (Fig. 5B;
P<0.05). These findings indicated that CDMP1 overexpression
suppressed the apoptosis of inflammatory cytokine-induced DRG cells
by regulating the expression of apoptosis-associated proteins.
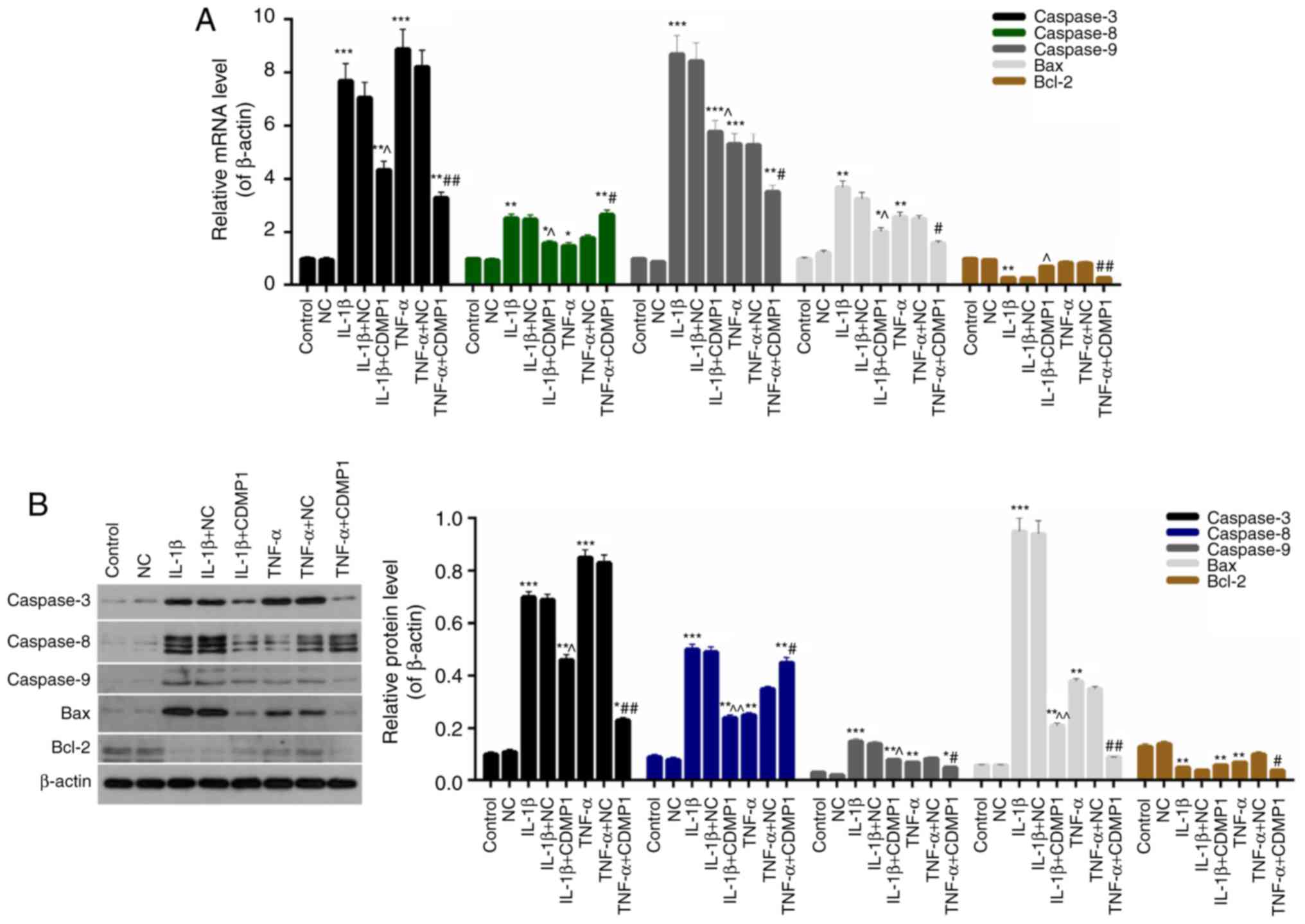 | Figure 5CDMP1 overexpression regulates the
expression of apoptosis-associated proteins in DRG cells. (A) mRNA
and (B) protein expression levels of Caspase-3, Caspase-8,
Caspase-9, Bax and Bcl-2 were examined in control DRG cells, DRG
cells that were transfected with empty vector (NC), and
inflammatory cytokine-induced DRG cells that were transfected with
either empty vector and CDMP1-expressing vector.
*P<0.05, **P<0.01, and
***P<0.001 vs. NC; ^P<0.05 and
^^P<0.01 vs. IL-1β+NC; #P<0.05 and
##P<0.01 vs. TNF-α+NC. CDMP1, cartilage-derived
morphogenetic protein-1; DRG, dorsal root ganglia; Bax, BCL2
associated X; Bcl-2, BCL2 apoptosis regulator; NC, negative control
empty vector transfection group; IL, interleukin; TNF, tumor
necrosis factor. |
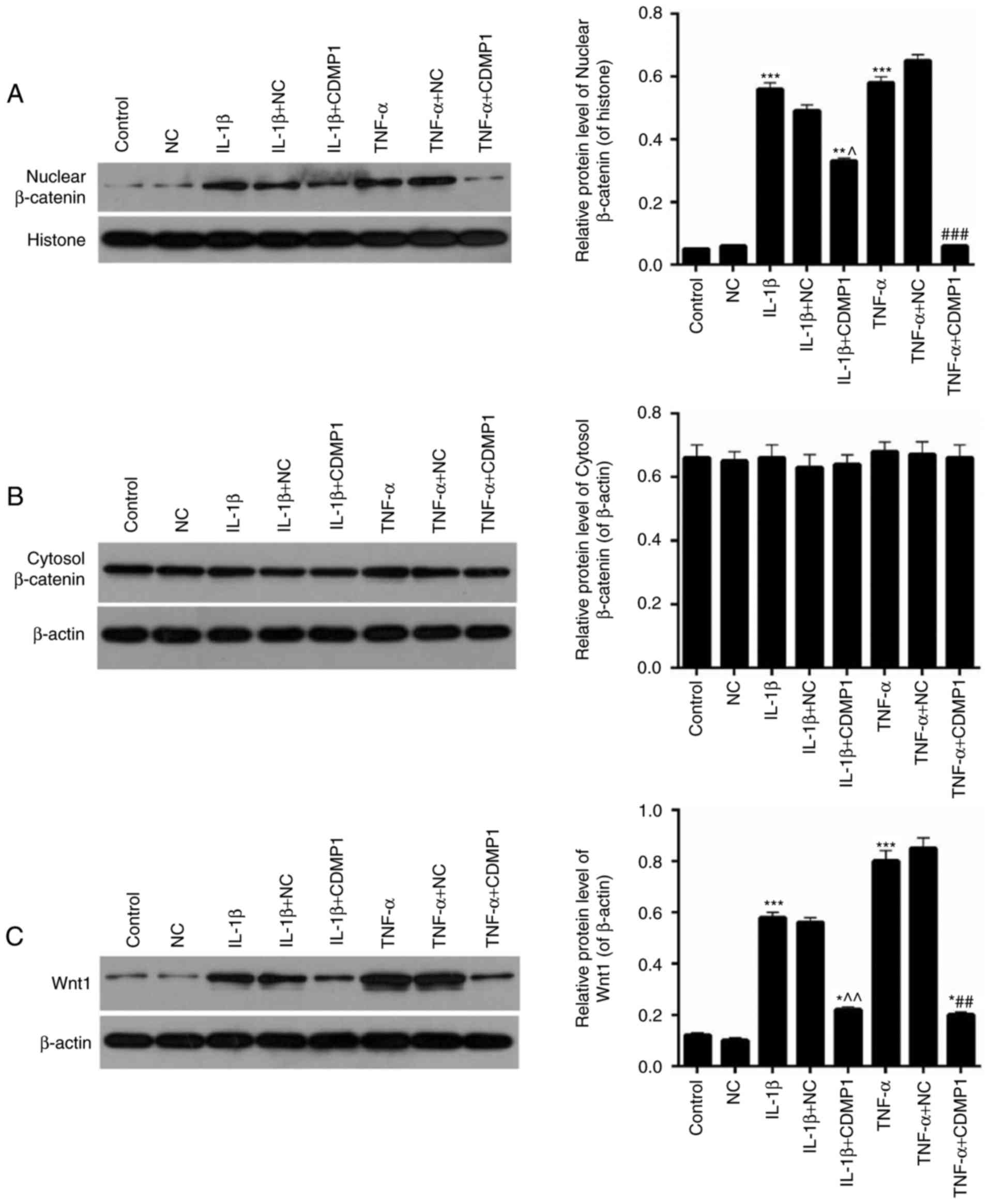
CDMP1 overexpression represses the
Wnt/β-Catenin pathway in inflammatory cytokine-induced DRG
cells
The expression levels of nuclear β-Catenin,
cytosolic β-Catenin and Wnt1 were assessed in DRG cells from each
group. Western blot results indicated that the expression levels of
nuclear β-Catenin were significantly downregulated in inflammatory
cytokine-induced DRG cells following CDMP1 overexpression (Fig. 6A; P<0.05). No significant
difference was observed on the levels of cytosolic β-Catenin in DRG
cells from each group (Fig. 6B).
In addition, CDMP1 overexpression significantly reduced the
expression levels of Wnt1 in inflammatory cytokine-induced DRG
cells (Fig. 6C; P<0.05).
Therefore, CDMP1 overexpression could affect the Wnt/β-Catenin
pathway in inflammatory cytokine-induced DRG cells.
Discussion
Apoptosis is one of the most widely studied
processes of cell death, and it serves an important role in the
development of embryos and the senescence of tissues. Cell
apoptosis in disc degeneration and herniation is a common
phenomenon (22–24). Inflammation, trauma, nitric oxide
and hypoxia can lead to apoptosis of DRG neurons. The mechanisms of
DRG cell apoptosis in disc herniation have been widely studied
(25–27). However, to the best of our
knowledge, no study that examines the regulation of apoptosis of
DRG cells in disc herniation has been reported to date. In the
present study, the aim was to explore the apoptosis pathway of DRG
cells in disc herniation through culture of DRG cells derived from
rats and their treatment with inflammatory cytokines. In addition,
the LDH rat model was established, as previously demonstrated
(28).
CDMP1 is expressed in several organs, such as the
cartilaginous core of long bones, osteoblast-like cells and
articular surfaces, and this indicates that CDMP1 is pivotal to the
development of bone and joints (18). Previous studies have demonstrated
that in animal models CDMP1 had important roles in the development
and progression of inter-vertebral discs (29–31). Among some studies, DRG tissues and
neurons were selected as the research targets (32–34). In the current investigation, it
was demonstrated that, compared with DRG tissues from normal rats,
the CDMP1 expression in DRG tissues from LDH model rats was
significantly downregulated. However, the expression levels of
inflammatory cytokines in DRG tissues from LDH model rats, such as
IL-1β and TNF-α, were markedly enhanced, and these results were in
accordance with previous studies (4–6).
By microscopy analysis, apoptosis of DRG cells was observed to be
promoted in DRG tissues from LDH model rats. These findings
suggested that inflammatory cytokines may induce the neuron
apoptosis in LDH, and that CDMP1 may regulate the induction of
inflammatory cytokines. In order to further explore the potential
effects of CDMP1 in inflammatory cytokine-induced neurons
apoptosis, an in vitro model of inflammatory cytokine (IL-1β
and TNF-α)-induced DRG cells was established, and CDMP1 was
overexpressed in these cells by plasmid transfection. Then, the
effect of CDMP1 overexpression was assessed in the viability and
apoptosis of inflammatory cytokine-induced DRG cells.
The current results demonstrated that CDMP1
overexpression significantly enhanced the cell viability of
inflammatory cytokine-induced DRG cells, particularly following
treatment for 72 h. Flow cytometry data indicated that CDMP1
overexpression significantly reduced the apoptosis of inflammatory
cytokine-induced DRG cells. In addition, CDMP1 overexpression
significantly downregulated the expression levels of Caspase-3/9
and Bax in inflammatory cytokine-induced DRG cells. Following
transfection with the CDMP1-expressing vector, the Caspase-8
expression was reduced in IL-1β-induced DRG cells, but enhanced in
TNF-α-induced DRG cells. CDMP1 overexpression also resulted in a
high Bcl-2 level in IL-1β-induced DRG, but a low Bcl-2 level in
TNF-α-induced DRG cells. Hence, the present results confirmed that
CDMP1 overexpression suppressed the apoptosis of inflammatory
cytokine-induced DRG cells via regulating Caspase-3/8/9, Bax and
Bcl-2.
Previous studies have suggested that the
Wnt/β-Catenin pathway serves as a critical signaling pathway in the
development of lumbar intervertebral disc degeneration and
herniation (35–38). However, very limited knowledge
exists regarding the effect of Wnt/β-Catenin signaling on
inflammatory cytokine-induced DRG cell apoptosis. Hence, the
expression levels of β-Catenin in nuclear and cytosolic extracts of
DRG cells from each group were examined. The results demonstrated
that CDMP1 overexpression markedly downregulated nuclear β-Catenin
expression in inflammatory cytokine-induced DRG cells.
Additionally, there was no significant difference in cytosolic
β-Catenin expression in inflammatory cytokine-induced DRG cells. Of
note, CDMP1 overexpression reduced the expression levels of Wnt1 in
inflammatory cytokine-induced DRG cells. Therefore, CDMP1
overexpression could downregulate the Wnt/β-Catenin pathway in
inflammatory cytokine-induced DRG cells.
In conclusion, the present study demonstrated that
CDMP1 overexpression reduced the apoptosis of inflammatory
cytokine-induced DRG cells by suppressing the Wnt/β-Catenin
pathway. The present findings provide a novel understanding of the
pathogenesis of LDH, and of the effects of CDMP1 in inflammatory
cytokine-induced DRG cells. The current results suggest that CDMP1
might be an effective target for LDH therapies.
Acknowledgments
Not applicable.
Funding
The study was supported by a grant from the National
Natural Science Foundation of China (grant no. 81371987).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors’ contributions
ZJ, XW, QY and LL made substantial contributions to
the conception and design of the study. ZJ, YZ and YS and JY were
responsible for analyzing and interpreting then data. ZJ and JPY
drafted the manuscript. All authors were responsible for giving
final approval of the version to be published.
Ethics approval and consent to
participate
The experimental protocols involving animals were
approved by the Shanxi Provincial People’s Hospital Ethics
Committee (Taiyuan, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Olmarker K and Larsson K: Tumor necrosis
factor alpha and nucleus-pulposus-induced nerve root injury. Spine
(Phila Pa 1976). 23:2538–2544. 1998. View Article : Google Scholar
|
|
2
|
Olmarker K, Rydevik B and Nordborg C:
Autologous nucleus pulposus induces neurophysiologic and histologic
changes in porcine cauda equina nerve roots. Spine (Phila Pa 1976).
18:1425–1432. 1993. View Article : Google Scholar
|
|
3
|
Raghavendra V, Tanga F, Rutkowski MD and
DeLeo JA: Anti-hyperalgesic and morphine-sparing actions of
propentofylline following peripheral nerve injury in rats:
Mechanistic implications of spinal glia and proinflammatory
cytokines. Pain. 104:655–664. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Genevay S, Finckh A, Payer M, Mezin F,
Tessitore E, Gabay C and Guerne PA: Elevated levels of tumor
necrosis factor-alpha in periradicular fat tissue in patients with
radiculopathy from herniated disc. Spine (Phila Pa 1976).
33:2041–2046. 2008. View Article : Google Scholar
|
|
5
|
Lee S, Moon CS, Sul D, Lee J, Bae M, Hong
Y, Lee M, Choi S, Derby R, Kim BJ, et al: Comparison of growth
factor and cytokine expression in patients with degenerated disc
disease and herniated nucleus pulposus. Clin Biochem. 42:1504–1511.
2009. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Kraychete DC, Sakata RK, Issy AM, Bacellar
O, Santos-Jesus R and Carvalho EM: Serum cytokine levels in
patients with chronic low back pain due to herniated disc:
Analytical cross-sectional study. Sao Paulo Med J. 128:259–262.
2010. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Takahashi H, Suguro T, Okazima Y, Motegi
M, Okada Y and Kakiuchi T: Inflammatory cytokines in the herniated
disc of the lumbar spine. Spine (Phila Pa 1976). 21:218–224. 1996.
View Article : Google Scholar
|
|
8
|
Nusse R and Varmus HE: Many tumors induced
by the mouse mammary tumor virus contain a provirus integrated in
the same region of the host genome. Cell. 31:99–109. 1982.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Clevers H: Wnt/beta-catenin signaling in
development and disease. Cell. 127:469–480. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Clevers H and Nusse R: Wnt/beta-catenin
signaling and disease. Cell. 149:1192–1205. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Nagaraj AB, Joseph P, Kovalenko O, Singh
S, Armstrong A, Redline R, Resnick K, Zanotti K, Waggoner S and
DiFeo A: Critical role of Wnt/β-catenin signaling in driving
epithelial ovarian cancer platinum resistance. Oncotarget.
6:23720–23734. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Huang J, Xiao D, Li G, Ma J, Chen P, Yuan
W, Hou F, Ge J, Zhong M, Tang Y, et al: EphA2 promotes
epithelial-mesenchymal transition through the Wnt/β-catenin pathway
in gastric cancer cells. Oncogene. 33:2737–2747. 2014. View Article : Google Scholar
|
|
13
|
Cai J, Guan H, Fang L, Yang Y, Zhu X, Yuan
J, Wu J and Li M: MicroRNA-374a activates Wnt/β-catenin signaling
to promote breast cancer metastasis. J Clin Invest. 123:566–579.
2013.PubMed/NCBI
|
|
14
|
Itokazu T, Hayano Y, Takahashi R and
Yamashita T: Involvement of Wnt/β-catenin signaling in the
development of neuropathic pain. Neurosci Res. 79:34–40. 2014.
View Article : Google Scholar
|
|
15
|
Hiyama A, Sakai D, Risbud MV, Tanaka M,
Arai F, Abe K and Mochida J: Enhancement of intervertebral disc
cell senescence by WNT/β-catenin signaling-induced matrix
metalloproteinase expression. Arthritis Rheum. 62:3036–3047. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Wang M, Tang D, Shu B, Wang B, Jin H, Hao
S, Dresser KA, Shen J, Im HJ, Sampson ER, et al: Conditional
activation of β-catenin signaling in mice leads to severe defects
in intervertebral disc tissue. Arthritis Rheum. 64:2611–2623. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Wu G, Cui Y, Wang YT, Yao M, Hu J, Li JX,
Wang Y and Zhang B: Repair of cartilage defects in BMSCs via CDMP1
gene transfection. Genet Mol Res. 13:291–301. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Feng C, Liu H, Yang Y, Huang B and Zhou Y:
Growth and differentiation factor-5 contributes to the structural
and functional maintenance of the intervertebral disc. Cell Physiol
Biochem. 35:1–16. 2015. View Article : Google Scholar
|
|
19
|
Gruber HE, Hoelscher GL, Ingram JA, Bethea
S and Hanley EN Jr: Growth and differentiation factor-5 (GDF-5) in
the human intervertebral annulus cells and its modulation by IL-1ß
and TNF-α in vitro. Exp Mol Pathol. 96:225–229. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Williams FM, Popham M, Hart DJ, de
Schepper E, Bierma-Zeinstra S, Hofman A, Uitterlinden AG, Arden NK,
Cooper C, Spector TD, et al: GDF5 single-nucleotide polymorphism
rs143383 is associated with lumbar disc degeneration in northern
european women. Arthritis Rheum. 63:708–712. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(−Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
22
|
Ha KY, Koh IJ, Kirpalani PA, Kim YY, Cho
YK, Khang GS and Han CW: The expression of hypoxia inducible
factor-1alpha and apoptosis in herniated discs. Spine (Phila Pa
1976). 31:1309–1313. 2006. View Article : Google Scholar
|
|
23
|
Kohyama K, Saura R, Doita M and Mizuno K:
Intervertebral disc cell apoptosis by nitric oxide: Biological
understanding of intervertebral disc degeneration. Kobe J Med Sci.
46:283–295. 2000.
|
|
24
|
Rannou F, Lee TS, Zhou RH, Chin J, Lotz
JC, Mayoux-Benhamou MA, Barbet JP, Chevrot A and Shyy JY:
Intervertebral disc degeneration: The role of the mitochondrial
pathway in annulus fibrosus cell apoptosis induced by overload. Am
J Pathol. 164:915–924. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Murata Y, Nannmark U, Rydevik B, Takahashi
K and Olmarker K: Nucleus pulposus-induced apoptosis in dorsal root
ganglion following experimental disc herniation in rats. Spine
(Phila Pa 1976). 31:382–390. 2006. View Article : Google Scholar
|
|
26
|
Murata Y, Nannmark U, Rydevik B, Takahashi
K and Olmarker K: The role of tumor necrosis factor-alpha in
apoptosis of dorsal root ganglion cells induced by herniated
nucleus pulposus in rats. Spine (Phila Pa 1976). 33:155–162. 2008.
View Article : Google Scholar
|
|
27
|
Murata Y, Rydevik B, Nannmark U, Larsson
K, Takahashi K, Kato Y and Olmarker K: Local application of
interleukin-6 to the dorsal root ganglion induces tumor necrosis
factor-α in the dorsal root ganglion and results in apoptosis of
the dorsal root ganglion cells. Spine (Phila Pa 1976). 36:926–932.
2011. View Article : Google Scholar
|
|
28
|
Obata K, Tsujino H, Yamanaka H, Yi D,
Fukuoka T, Hashimoto N, Yonenobu K, Yoshikawa H and Noguchi K:
Expression of neurotrophic factors in the dorsal root ganglion in a
rat model of lumbar disc herniation. Pain. 99:121–132. 2002.
View Article : Google Scholar : PubMed/NCBI
|
|
29
|
Chujo T, An HS, Akeda K, Miyamoto K,
Muehleman C, Attawia M, Andersson G and Masuda K: Effects of growth
differentiation factor-5 on the intervertebral disc-in vitro bovine
study and in vivo rabbit disc degeneration model study. Spine
(Phila Pa 1976). 31:2909–2917. 2006. View Article : Google Scholar
|
|
30
|
Li X, Leo BM, Beck G, Balian G and
Anderson GD: Collagen and proteoglycan abnormalities in the
GDF-5-deficient mice and molecular changes when treating disk cells
with recombinant growth factor. Spine (Phila Pa 1976).
29:2229–2234. 2004. View Article : Google Scholar
|
|
31
|
Liang H, Ma SY, Feng G, Shen FH and Joshua
Li X: Therapeutic effects of adenovirus-mediated growth and
differentiation factor-5 in a mice disc degeneration model induced
by annulus needle puncture. Spine J. 10:32–41. 2010. View Article : Google Scholar :
|
|
32
|
Sato J, Inage K, Miyagi M, Sakuma Y,
Yamauchi K, Koda M, Furuya T, Nakamura J, Suzuki M, Kubota G, et
al: Inhibiting vascular endothelial growth factor in injured
intervertebral discs attenuates pain-related neuropeptide
expression in dorsal root ganglia in rats. Asian Spine J.
11:556–561. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Watanabe K, Larsson K, Rydevik B, Konno S,
Nordborg C and Olmarker K: Increase of sodium channels (nav 1.8 and
nav 1.9) in rat dorsal root ganglion neurons exposed to autologous
nucleus pulposus. Open Orthop J. 8:69–73. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Yan J, Zou K, Liu X, Hu S, Wang Q, Miao X,
Zhu HY, Zhou Y and Xu GY: Hyperexcitability and sensitization of
sodium channels of dorsal root ganglion neurons in a rat model of
lumber disc herniation. Eur Spine J. 25:177–185. 2016. View Article : Google Scholar
|
|
35
|
Chen J, Jia YS, Liu GZ, Sun Q, Zhang F, Ma
S and Wang YJ: Role of LncRNA TUG1 in intervertebral disc
degeneration and nucleus pulposus cells via regulating
Wnt/β-catenin signaling pathway. Biochem Biophys Res Commun.
491:668–674. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Jia H, Ma J, Lv J, Ma X, Xu W, Yang Y,
Tian A, Wang Y, Sun L, Xu L, et al: Oestrogen and parathyroid
hormone alleviate lumbar intervertebral disc degeneration in
ovariectomized rats and enhance Wnt/β-catenin pathway activity. Sci
Rep. 6:275212016. View Article : Google Scholar
|
|
37
|
Wang X, Shi SH, Yao HJ, Jing QK, Mo YP, Lv
W, Song LY, Yuan XC, Li ZG and Qin LN: Electroacupuncture at Dazhui
(GV14) and Mingmen (GV4) protects against spinal cord injury: The
role of the Wnt/beta-catenin signaling pathway. Neural Regen Res.
11:2004–2011. 2016. View Article : Google Scholar
|
|
38
|
Xie H, Jing Y, Xia J, Wang X, You C and
Yan J: Aquaporin 3 protects against lumbar intervertebral disc
degeneration via the Wnt/β-catenin pathway. Int J Mol Med.
37:859–864. 2016. View Article : Google Scholar : PubMed/NCBI
|