Introduction
Tongue squamous cell carcinoma (TSCC) is the most
common form of head and neck cancer, and is a major cause of
cancer-related mortality worldwide (1). Although the overall survival rate of
patients with TSCC can reach 5 years or more, numerous patients
experience metastatic disease and a poor prognosis (2). Declines in mortality rates have been
attributed to earlier detection and improvements in therapy.
However, the precise underlying mechanisms of metastasis remain
unclear.
Cancer invasiveness and metastasis are tightly
linked with the acquisition of a migratory phenotype that enables
cancer cells to invade adjacent tissues. Epithelial-mesenchymal
transition (EMT) is known to be a central mechanism in charge of
the invasiveness and metastasis of cancer via the loss of the cell
to cell contact of epithelial cells and the acquisition of
migratory properties (3). It has
been confirmed that the acquisition of the expression of
mesenchymal markers, including vimentin, Slug, Snail and Twist, but
the reduction of the expression of epithelial markers, including
E-cadherin, endows cells with high potential for dissemination
(4). Cell movement is regulated
by members of the regulatory factors and signaling pathways. The
Wnt/β-catenin signaling pathway has been certified to regulate cell
motility and confer cellular metastasis in cancer (5,6).
It has also been proven that β-catenin participates in the
regulation of E-cadherin, Snail and Twist, and that it induces
epithelial cells to become invasive in colon cancer (7).
The transcription factor Sox2 (Sry-box2), a
high-mobility group DNA binding protein, serves an important role
in various phases of embryonic development, and affects cell fate
and differentiation (8). Sox2 has
been studied in several types of human solid tumors, such as breast
cancer, prostate cancer and glioblastoma tumor (9-11).
Li et al (12) suggested
that Sox2 could mediate EMT by inducing β-catenin in breast and
prostate cancer. While the aforementioned cancer types are each
adenocarcinoma, it was uncertain whether β-catenin would be a key
factor in squamous cell carcinoma, particularly TSCC. Sox2 is
overexpressed in cancerous tissues compared with that in
para-tumoral tissues in oral squamous cell carcinoma (OSCC)
(13), and the high expression of
Sox2 in the primary tissues of OSCC has been significantly
correlated with the poor prognosis accompanying lymph node
metastasis (14). However, OSCC
is an umbrella term that included TSCC, buccal mucosa squamous cell
carcinoma, mouth floor squamous cell carcinoma, gingival carcinoma
and carcinoma of the palate. Heterogeneity among these types of
squamous cell carcinoma may exist. The present study aimed to
investigate the function of Sox2 expression with a focus on
TSCC.
Materials and methods
Patients and tissue samples
Specimens were obtained from the patients (35 male
and 26 female patients; mean age, 53.25 years; median age, 54
years) following radical surgery (open operation for the excision
of lesion and part of normal tongue surrounding) between January
2005 and December 2015 at Guanghua Hospital of Stomatology of Sun
Yat-sen University (Guangdong, China). The patients selected had
primary squamous cell carcinoma of the tongue, had not been
subjected to radiotherapy or chemotherapy preoperatively and had no
history of other systemic diseases (e.g. diabetes, hypertension,
etc.). The confirmed diagnosis was tongue squamous cell carcinoma
by pathological examination. Informed consent was obtained on the
use of surgically resected specimens for research purposes,
according to the guidelines for research for human tissues and
samples set by the Institution Review Board (IRB) of Sun Yat-sen
University, who also approved the study. No patients received any
form of adjuvant therapy prior to surgery. These tissues included
61 pairs of TSCC samples and corresponding adjacent non-cancerous
tissues. Of the 61 TSCC samples, there were 31 (50.8%)
well-differentiated and 30 (49.2%) moderately or poorly
differentiated TSCCs, plus 26 (42.6%) tissues with lymph node
metastases. The clinical characteristics of all these patients are
summarized in Table I. Clinical
stages were classified according to the Union for International
Cancer Control (2002) (15).
 | Table IAssociation between Sox2 expression
and clinicopathological features in tongue squamous cell carcinoma
patients. |
Table I
Association between Sox2 expression
and clinicopathological features in tongue squamous cell carcinoma
patients.
| Clinicopathological
features | No. of cases | Sox2 expression, n
(%)
| P-valuea |
|---|
| High | Low |
|---|
| Sex | | | | |
| Male | 35 | 23 (65.7) | 12 (34.3) | 0.737 |
| Female | 26 | 16 (61.5) | 10 (39.5) | |
| Age, years | | | | |
| ≥55 | 29 | 16 (53.3) | 13 (46.7) | 0.175 |
| <55 | 32 | 23 (74.2) | 9 (25.8) | |
|
Differentiation | | | | |
| Well | 31 | 16 (51.6) | 15 (48.4) | 0.042b |
| Moderately +
poorly | 30 | 23 (76.7) | 7 (23.3) | |
| pT stage | | | | |
| T1-2 | 46 | 26 (56.5) | 20 (43.5) | 0.035b |
| T3-4 | 15 | 13 (86.7) | 2 (13.3) | |
| pN stage | | | | |
| N0 | 38 | 20 (52.6) | 18 (47.4) | 0.018b |
| N+ | 23 | 19 (82.6) | 4 (17.4) | |
| pTNM stage | | | | |
| I-II | 31 | 14 (45.2) | 17 (54.8) | 0.002b |
| III-IV | 30 | 25 (83.3) | 5 (16.7) | |
Immunohistochemistry
Expression of Sox2 was measured by
immunohistochemical staining. Paraffin-embedded specimens were cut
into 4-μm thick sections at room temperature. These paraffin
sections were deparaffinized and rehydrated, and then immersed in
3% H2O2 to quench endogenous peroxidase
activity. Subsequent to being treated with 0.01 M citrate buffer,
the sections were blocked with 5% goat serum followed by incubation
with anti-Sox2 antibody (dilution, 1:100; rabbit polyclonal
anti-human; catalog no. 3579; Cell Signaling Technology, Inc.,
Danvers, MA, USA) overnight at 40°C. After washing three times in
phosphate-buffered saline (PBS), the slides were incubated with
ChemMate™ EnVision secondary antibody/horseradish peroxidase (HRP)
for 1 h at room temperature and then visualized with
diaminobenzidine (Dako; Agilent Technologies GmbH, Waldbronn,
Germany). The slides were counterstained with hematoxylin at room
temperature. To confirm the specificity of the immunostaining, a
negative control was included in each run by substituting PBS for
the primary antibody.
Evaluation of immunohistochemical
staining
Five random fields of each section were viewed under
a light microscope (Axioskop 40; Zeiss GmbH, Jena, Germany) at ×400
magnification. The sections were independently examined and scored
by three investigators, who were blinded to the clinical features
and outcomes. The expression of Sox2 was scored by multiplication
of the mean signal intensity (on a scale of 0-3: 0, no staining; 1,
light staining; 2, moderate staining; 3, high staining) and the
percentage of positively stained tumor cells (on a scale of 0-4: 0,
0%; 1, 0-25%; 2, 26-50%; 3, 51-75%; 4, 76-100%). The final
immunoreactive score reported is the mean of the scores from the
three investigators. By receiver operating characteristic analysis,
the cases with a final score >4 were classified as exhibiting
high Sox2 expression (sensitivity 82.9%, specificity 91.6%),
whereas the cases with a score ≤4 were classified as exhibiting low
Sox2 expression.
Cell culture
TSCC cell lines, UM1, UM2, Cal27 and SCC9 (preserved
and managed in Guangdong Provincial Key Laboratory of Stomatology,
Guangzhou, China), and one human oral mucosa epithelial cell
immortalized by insertion of the human telomerase reverse
transcriptase (hTERT) gene (hTERT+-OME) (16) were used in this study. UM1, UM2,
Cal27 and hTERT+-OME cells were maintained in Dulbecco's
modified Eagle's medium (DMEM)/F12 (Sigma-Aldrich; Merck KGaA,
Darmstadt, Germany) containing 10% fetal bovine serum (FBS; Gibco,
New York, NY, USA) at 37°C with 5% CO2. SCC9 cells were
cultured in DMEM/F12 containing 1% hydrocortisone and 10% FBS in
the same conditions as aforementioned. The cells were confirmed to
be free from mycoplasma by testing of the cell lines once every 3
months.
Establishment of Sox2-overexpressing TSCC
cells
Human full-length Sox2 cDNA was cloned into
pSin-EF2-Sox2-Pur (plasmid no. 16577; Addgene, Inc., Cambridge, MA,
USA). Using co-transfection of plasmid DNA with lentivector plus
helper plasmids (pRSV-REV, pMDLg-pRRE and pMD2.G)
(isoconcentration), transfected into 293T cells occurred using
Lipofectamine® 2000 (Invitrogen; Thermo Fisher
Scientific, Inc., Waltham, MA, USA). In order to improve the
transfection efficiency, 6 μg/ml polybrene was added in the
culture medium incubated with TSCC cells and lentivirus. Stable
Sox2-overexpressing TSCC cells were purified using the antibiotic
puromycin.
Construction of Sox2-silenced TSCC
cells
Sox2 short hairpin RNA (shRNA) transduction plasmids
were purchased from Sigma-Aldrich; Merck KGaA (catalog no.
TRCNOO000231641). The sequence of this region was as follows:
5′-CCGGCAACGGCAGCTACAGCATGATCTCGAGATCATGCTGTAGCTGCCGTTGTTTTTG-3′.
MISSION® non-target shRNA control (shRNA-NC)
transduction particles (Sigma-Aldrich; Merck KGaA) were utilized
for the experimental control. Lentivirus production was performed
by co-transfection of plasmid Sox2 shRNA and helper plasmid
(pCM-VSV-G) (isoconcentration) using Lipofectamine 2000
(Invitrogen; Thermo Fisher Scientific, Inc.). Procedures of cell
transfection and purification were as aforementioned.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR)
Total RNA from the cultured cells was prepared using
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.)
according to the manufacturer's protocols. Total RNA (1 μg)
was reverse transcribed to cDNA using a reverse transcription kit
(Toyobo Life Science, Osaka, Japan) according to the manufacturer's
protocols. The amplification was performed in 96-well plates, each
with a total volume of 20 μl, containing 100 nM forward and
reverse primers, 4 mM MgCl2, LightCycler SYBR-Green I
and 2 μl cDNA template. Amplification was performed with an
initial denaturation step at 95°C for 10 min, followed by 40 cycles
of 95°C for 15 sec, 55°C for 30 sec and 72°C for 25 sec. The
reference gene used was β-actin and the primer sequences used were
forward, 5′-TGAAGTGTGACGTGGACATC-3′ and reverse,
5′-GGAGGAGCAATGATCTTGAT-3′. The quantification method used was the
2−∆∆Cq method (17).
All reactions were performed in duplicate three times.
Western blot analysis
Proteins (20 μg per lane) were separated by
sodium dodecyl sulfate-polyacrylamide gel electrophoresis and
transferred onto polyvinylidene difluoride membranes (10%
separation gel and 5% spacer gel). RIPA lysis buffer was the
protein extraction buffer used. BCA was used for protein
determination. Subsequent to blocking using 5% skimmed milk powder
blended with TBST at room temperature for 2 h, the membranes were
then incubated with anti-human antibodies overnight at 40°C. Target
proteins were detected by enhanced chemiluminescence reagents
(Thermo Fisher Scientific, Inc.) following incubation with
secondary antibodies conjugated to HRP [HRP-labeled goat
anti-rabbit IgG (H+L) (A0208) and HRP-labeled goat anti-mouse IgG
(H+L) (A0216) (dilution, 1:1,000; Beyotime Institute of
Biotechnology, Shanghai, China)] for 1 h at room temperature.
Glyceraldehyde 3-phosphate dehydrogenase was used as an internal
control.
Cell proliferation assay
Cell proliferation was analyzed using a colorimetric
(MTT) assay kit. Cells (2×104) were seeded in each well
of a 96-well plate and cultured for 24-72 h at 37°C with 5%
CO2. Next, 10 μl MTT solution was added into the
culture medium, followed by incubation for 4 h at 37°C with 5%
CO2. Dimethyl sulfoxide (100 μl) was added to
dissolve the formazan and the color reaction was analyzed with a
microplate reader set at 560 nm.
Transwell assay
Briefly, 24-well plates containing 8-μm pore
Transwell inserts (EMD Millipore, Billerica, MA, USA) were coated
with Matrigel (BD Biosciences, San Jose, CA, USA). The upper
chambers were filled with a suspension of 1×104 cells
cultured in serum-free DMEM/F12, while the lower chambers were
filled with DMEM/F12 plus 10% FBS. Following 48 h of incubation at
37°C, the filters were fixed with methanol for 30 min and stained
with crystal violet for 20 min at room temperature. The number of
cells which had invaded through the filter pores was counted under
a phase-contrast microscope.
Wound-healing assay
Cultured cells (2×105) were seeded in
each well of a 6-well plate. When the cells reached 90-100%
confluence, a wound was scratched in the central area of the
confluent culture, followed by careful washing to remove detached
cells. Next, the cells were cultured at 37°C with 5%
CO2. Images of the wounded areas were captured using a
phase-contrast microscope at 0 and 24 h.
Plate clone formation assay
Each well of the culture plate was seeded with 200
cells and incubated at 37°C for 2 weeks. The cells were then fixed
with 4% paraformaldehyde for 15 min and then stained with Giemsa
for 15 min at room temperature. The number of visible colonies with
a diameter ≥100 μm was counted under a light microscope
(magnification, ×40; Axioskop 40; Zeiss GmbH).
Statistical analysis
Experimental results presented in the figures are
representative of at least three different repetitions. The data
are presented as the mean ± standard deviation. The associations
between Sox2 expression and clinicopathological parameters were
determined using the χ2 test. Patient survival was
evaluated using the Kaplan-Meier method, and the statistical
significance was examined using the log-rank test. Assessment of
the survival time compared with the clinicopathological variables
was performed by univariate and multivariate analyses using the Cox
regression model. Comparisons between groups were evaluated using
Student's t-test and analysis of variance (one-way ANOVA) followed
by Student-Newman-Keuls (S-N-K) post hoc test. P<0.05 was
considered to indicate a statistically significant difference.
Statistical analysis was performed using SPSS for Windows (version
22.0; IBM SPSS, Inc., Armonk, NY, USA).
Results
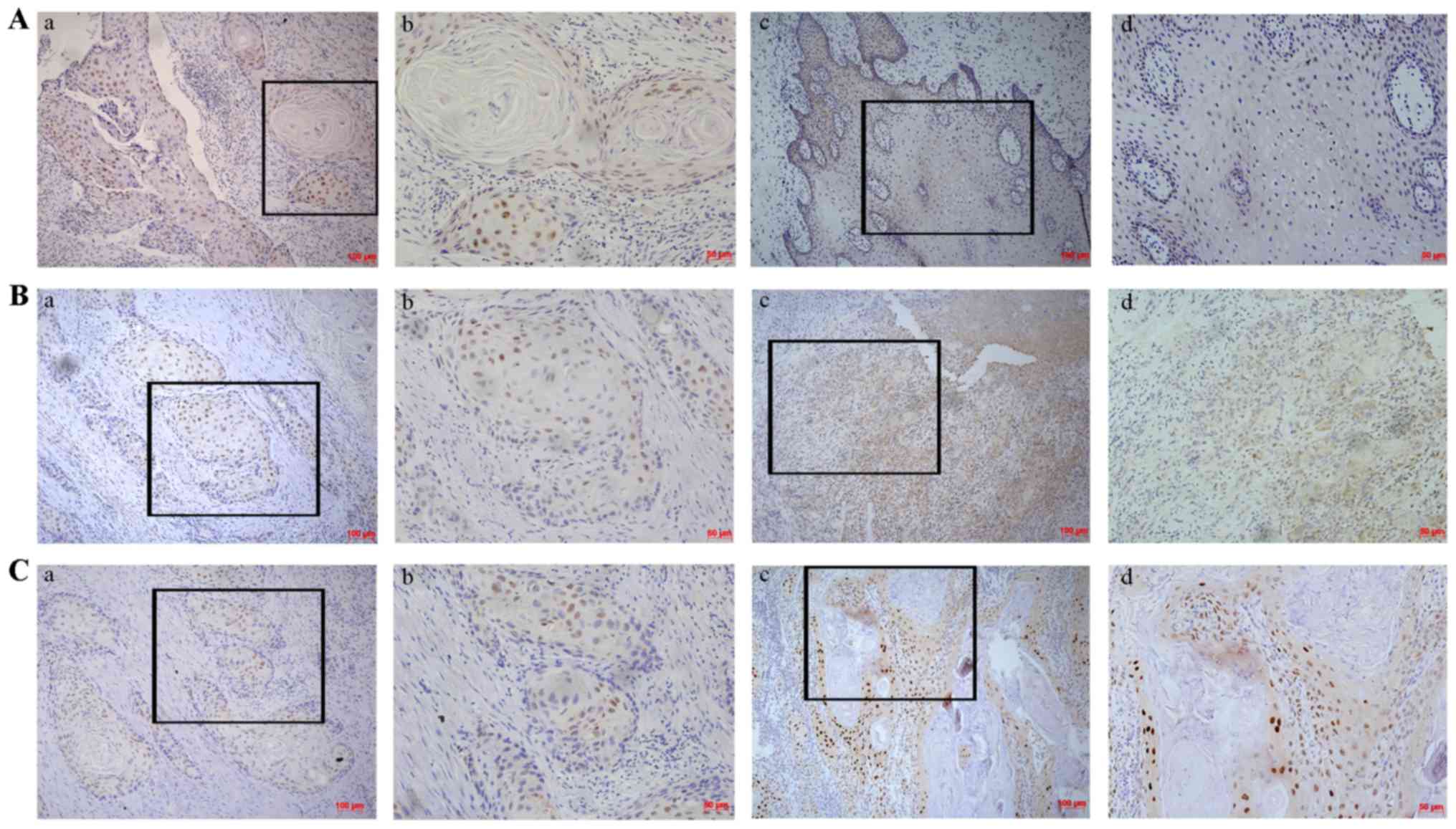
Expression of Sox2 in TSCC
The expression of Sox2 was evaluated by
immunohistochemical staining in 61 pairs of TSCC samples and
corresponding adjacent non-cancerous tissues. As shown in Fig. 1, significantly higher expression
of Sox2 was observed in TSCC samples (83.6%, 51/61) compared with
that in the corresponding adjacent non-cancerous tissues (63.9%,
39/61). In order to evaluate the role of Sox2 in TSCC, associations
between Sox2 expression and the clinicopathological parameters were
investigated (Table I). The
results showed that Sox2 expression was significantly associated
with tumor differentiation (P=0.042), pT stage (P=0.035), pN stage
(P=0.018) and pTNM stage (P=0.002). No significant association was
found between Sox2 expression and sex (P=0.737) or age
(P=0.175).
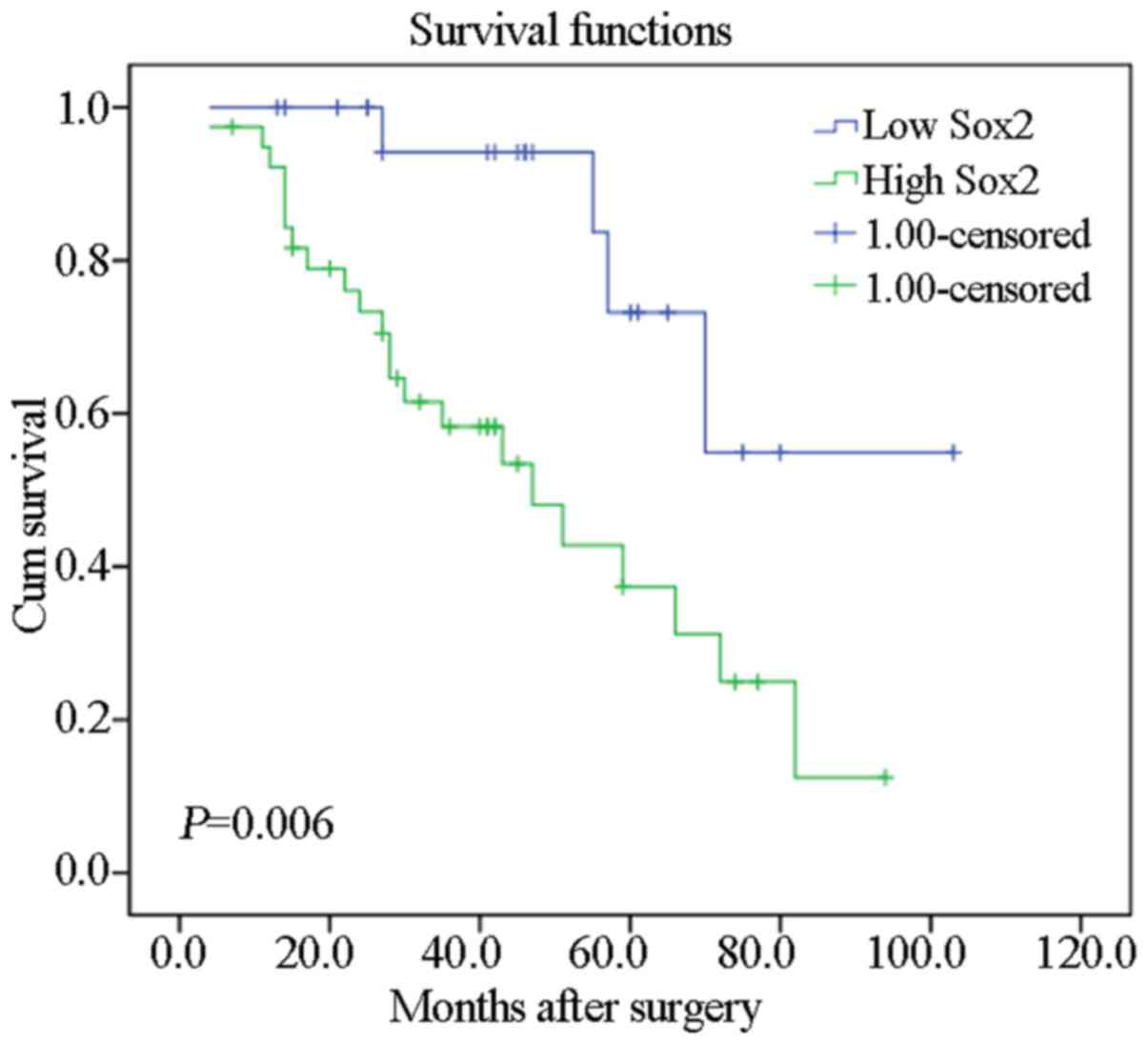
Association between Sox2 expression and
survival in TSCC patients
To investigate the association between Sox2
expression and the clinical outcome of TSCC patients, associations
between patient survival status and Sox2 expression were analyzed.
The results showed that patients with tumors with high Sox2
expression had a significantly worse prognosis than those with low
Sox2 expression (P=0.006; Fig.
2).
To further assess whether Sox2 expression represents
a prognostic parameter in patients with TSCC, regression analysis
using Cox proportional hazards model was performed. Upon univariate
analysis, low Sox2 expression and pN+ stage were
associated with a poor prognosis. Moreover, multivariate analysis
was performed using the significant variables observed in
univariate analysis. The results showed that Sox2 expression was
the only independent prognostic predictor (P=0.033) (Table II). These results strongly
indicated that the downregulated Sox2 expression in TSCC patients
is closely associated with a poor prognosis.
| Table IICox proportional hazards model
analysis of variables affecting survival in tongue squamous cell
carcinoma patients. |
Table II
Cox proportional hazards model
analysis of variables affecting survival in tongue squamous cell
carcinoma patients.
| Variables | Categories | Univariate analysis
| Multivariate
analysis
|
|---|
| HR (95% CI) | P-value | HR (95% CI) | P-value |
|---|
| Gender | Female/male | 0.735
(0.333-1.624) | 0.447 | | |
| Age, years | <55/≥55 | 1.614
(0.738-3.533) | 0.231 | | |
|
Differentiation | Moderately +
poorly/well | 1.349
(0.623-2.925) | 0.448 | | |
| pT stage | T3-4/T1-2 | 1.615
(0.714-3.653) | 0.250 | | |
| pTNM stage | III-IV/I-II | 2.129
(0.961-4.719) | 0.063 | | |
| pN stage |
N+/N0 | 2.319
(1.055-5.095) |
0.036a | 1.582
(0.698-3.583) | 0.272 |
| Sox2 | High/low | 3.977
(1.365-11.582) |
0.011a | 3.366
(1.101-10.294) |
0.033a |
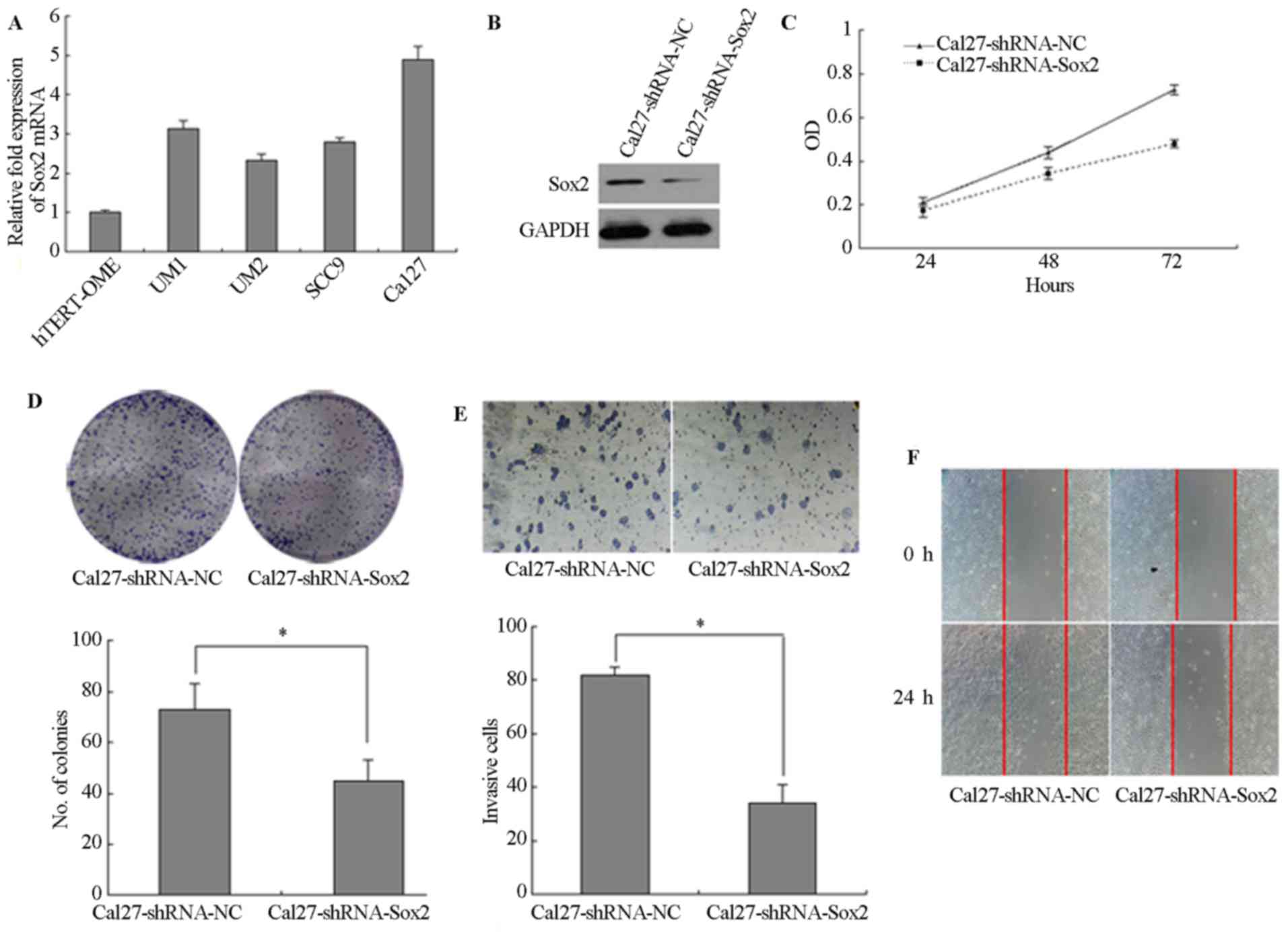
Sox2 regulates malignant phenotypes and
EMT progression in TSCC
The expression of Sox2 was detected by RT-qPCR and
the relative expression level of Sox2 was lower in immortalized
oral mucosal epithelial cells (hTERT+-OME) than that in
the 4 TSCC cells (UM1, UM2, SCC9 and Cal27) (Fig. 3A). To determine the role of Sox2
in TSCC cells, the expression of Sox2 was downregulated in Cal27
cells by transfection with lentiviral vector expressing shRNA and
the expression of Sox2 was lower in Cal27-shRNA-Sox2 than that in
Cal27-shRNA-NC (Fig. 3B).
Silencing of Sox2 significantly inhibited the proliferation of the
Cal27 cells (P<0.05; Fig. 3C).
Following knockdown of Sox2, the colony number was also decreased.
A longer incubation time was required to generate colonies of
equivalent size to those generated by Cal27-shRNA control cells
(P<0.05; Fig. 3D). In
addition, knockdown of Sox2 significantly reduced the cell invasion
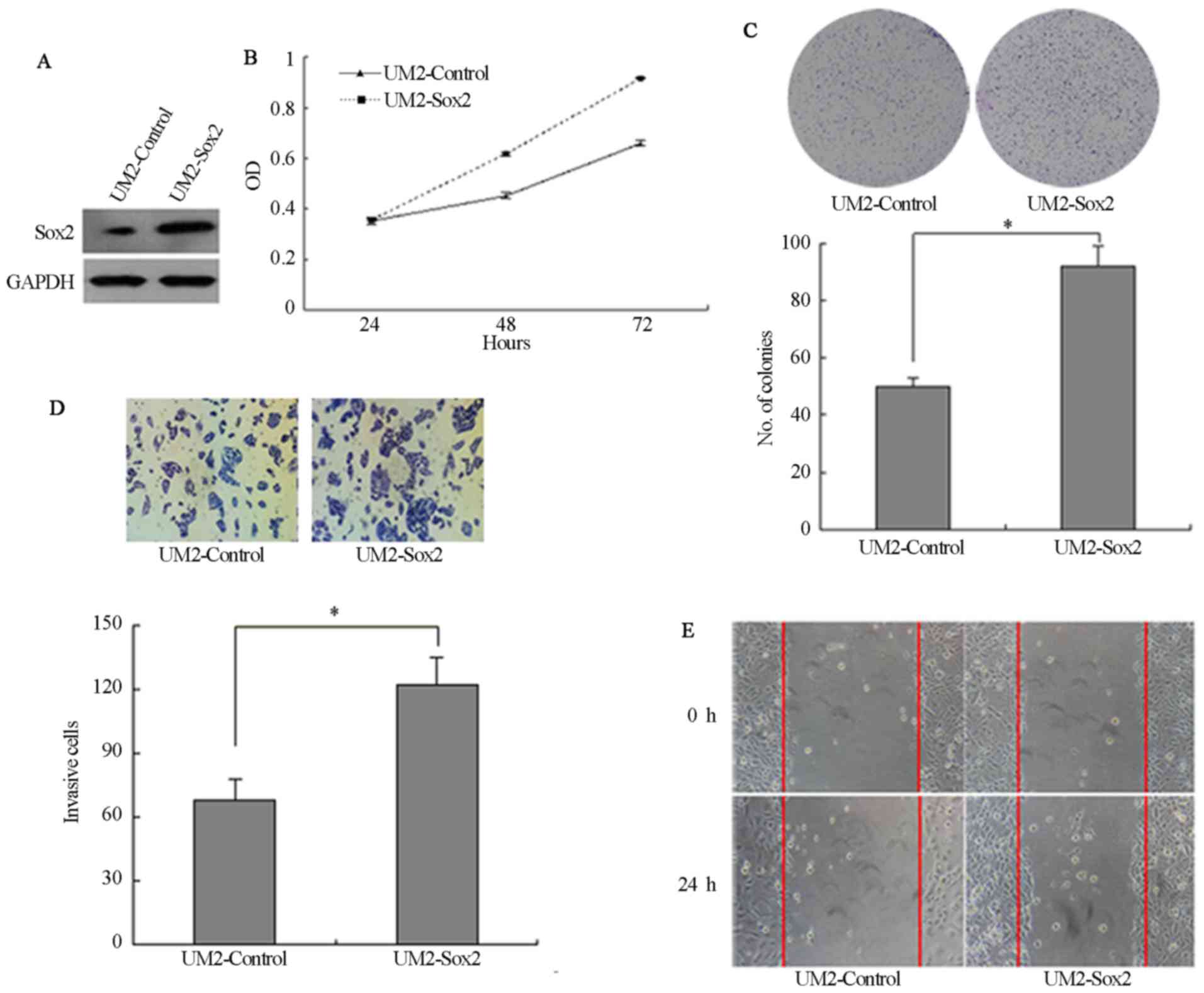
and wound-healing ability (P<0.05; Fig. 3E and F). Furthermore, UM2 cells
were stably transfected with Sox2 cDNA and the expression of Sox2
was higher in UM2-Sox2 than that in Sox2-control (Fig. 4A). UM2 cells transfected with Sox2
(UM2-Sox2) showed significantly higher cell proliferation
(P<0.05; Fig. 4B).
Overexpression of Sox2 in UM2 cells increased the colony numbers by
78.4% (P<0.05; Fig. 4C). Also,
Sox2-overexpressing cells invaded faster than control cells
(P<0.05; Fig. 4D). In
addition, wound-healing assays confirmed these results (Fig. 4E), suggesting that Sox2 affects
the motility in UM2 cells.
To determine if there is an assoicaation between
Sox2 expression and the process of EMT, RT-qPCR and western blot
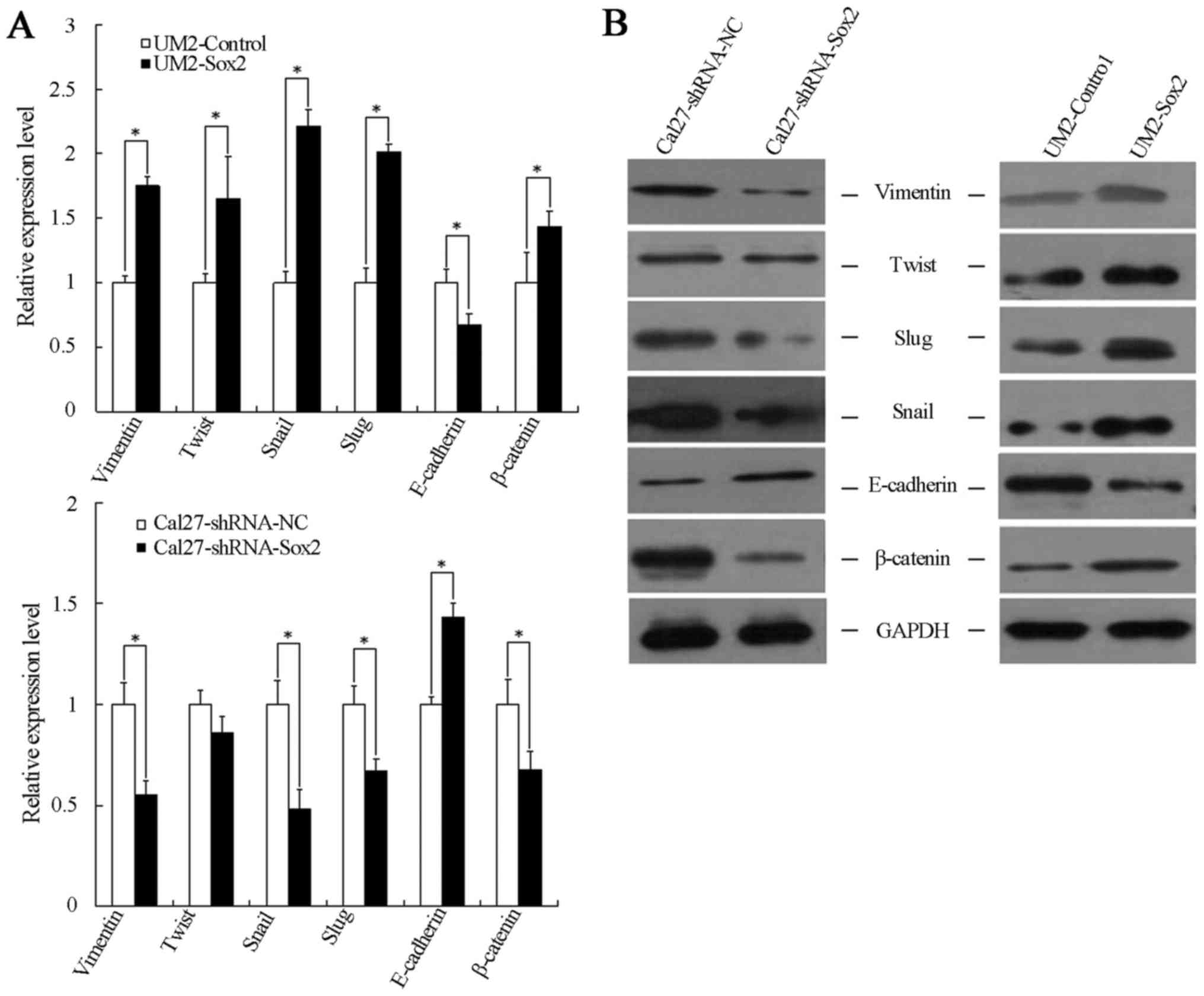
analysis were performed. Compared with the cells transfected with
shRNA-NC, the mesenchymal cell markers (Snail, Twist, vimentin and
Slug) were downregulated, while the epithelial cell marker
(E-cadherin) was upregulated in Cal27-shRNA-Sox2 cells. By
contrast, upregulation of the mesenchymal cell markers and
downregulation of the epithelial marker was found in
Sox2-overexpressing cells (Fig. 5A
and B).
Sox2 modulates β-catenin expression
Since the Wnt/β-catenin signaling pathway is closely
associated with cell tumorigenesis and migration, and it is also an
important route via which to promote the process of EMT,
experiments were designed to interrogate whether Sox2 expression
was closely associated with the expression of β-catenin. As shown
in Fig. 5, β-catenin expression
increased significantly in UM2-Sox2 cells and decreased in
Cal27-shRNA-Sox2 cells, at the mRNA and protein levels. These
results suggest that β-catenin may associated with a higher
malignant phenotype and EMT progression in TSCC cells with
alterations in Sox2 expression.
Discussion
TSCC, as the most common tumor of the oral cavity,
has a high frequency of local recurrence and distant metastatic
characteristics (18). Recent
studies indicated that EMT and the Wnt/β-catenin signaling pathway
were associated with oncogenesis, invasion and metastasis in
various tumors (19-22). Thus, understanding the underlying
molecular mechanism of TSCC aggression and transformation processes
could enhance the treatment outcome of patients with TSCC. The
present study demonstrated that Sox2 can affect tumor
aggressiveness and EMT in TSCC.
Overexpression of Sox2 is often associated with
increased cancer aggressiveness, resistance to chemo-radiation
therapy and decreased survival rate, which have been reported in
various cancer types, including breast, prostate, lung, ovarian and
colon cancer (9,10,23-25). The present study revealed that
Sox2 is vital in the regulation of TSCC motility, invasion and
tumorigenicity. The results indicated that upregulated Sox2
expression was significantly associated with the progression of
TSCC, as we previously described (26). Sox2 expression was significant
higher in primary TSCC tissues, the para-tumoral tissues and oral
epithelial dysplastic (pre-invasive) tissues compared with the oral
normal mucosa (26). Sox2 was
detected in oral pre-invasive lesions, suggesting that Sox2
upregulation may be an early event in TSCC carcinogenesis.
The present data also revealed that Sox2 expression
was higher in UM1 cells than in UM2 cells. UM1 and UM2 cells were
established from the same tongue carcinoma tissue from patients
without any adjuvant treatment. The growth pattern of UM1 was
scattered, while UM2 cells formed colonies with firm adhesion.
Compared with UM2 cells, UM1 cells exhibit a spontaneous EMT
pattern, with higher motility, invasive and metastatic activities
in viv and in vitro (27). From the present results, we
speculate that Sox2 may serve an important role in the aggressive
behavior of TSCC. To confirm this hypothesis, two TSCC cell lines
were constructed to evaluate the role of Sox2 through
lentiviral-mediated overexpression and lentiviral-mediated
knockdown. From this investigation, it was noted that Sox2 could
modulate cell aggression and motility by affecting the capability
of migration, invasion and proliferation in TSCC cells.
EMT is well known for its major role in
embryogenesis and development, which involves growth and
differentiation (28). The
regulatory changes that enable EMT to drive as a normal process of
increased growth and differentiation in developing populations of
cells within an organism. Nevertheless, when the same modifications
occur in cancer cells, metastasis can happen. As an important
factor associated with cancer aggression, EMT is often involved in
invasion and metastasis during tumor progression (29). It has been confirmed that numerous
classical molecular pathways are associated with EMT, including the
Wnt/β-catenin, Snail/Slug, transforming growth factor-β (TGF-β),
Twist and Cripto pathways (30).
Recent studies showed that cytokines, including TGF-β, epidermal
growth factor (EGF) and insulin-like growth factor, mediated the
occurrence of EMT in normal epithelial cells and cancer cells by
autocrine or paracrine signaling (31-33). Nuclear transcription factors,
which include Twist, Snail and Slug, also served important roles in
the process of tumor metastasis by promoting EMT (34-36). In the present study, Sox2 was
shown to possess the capacity to promote the characteristics of EMT
in TSCC cells. Upregulation of Sox2 in UM2 led to the enhanced
expression of Snail, Slug, Twist and vimentin, and the reduction in
the protein level of E-cadherin. In addition, silencing of Sox2
could attenuate the expression of mesenchymal markers, and could
elevate the epithelial marker expression in Cal27 tongue cancer
cells.
Sox2 has been described in certain cases as an
oncogene, and it can control cancer physiology via promoting
oncogenic signaling pathways, such as that of Wnt/β-catenin and
phosphoinositide 3-kinase/protein kinase B (37,38). Meanwhile, Sox2 is mediated by
certain upstream signals to regulate the cell survival,
tumorigenesis, invasion and migration reported in recent
literature. These signals include EGF receptor signals, the
Hedgehog/glioma-associated oncogene homolog zinc finger protein
signaling pathway, and transcription factor mammalian target of
rapamycin and signal transducer and activator of transcription
signals (39-42). Sox2 acts as an important factor in
these signaling pathways.
The Wnt/β-catenin signaling pathway is associated
with malignant cancer progression, and is also an important way to
promote the process of EMT. Abundant evidence has shown that Sox2
promotes tumor metastasis by stimulating EMT via the Wnt/β-catenin
pathway in breast and laryngeal cancer (12,43). β-catenin itself is one of the
vital downstream molecules that mediate EMT induced by Sox2
(12). In the present study, the
results revealed that β-catenin expression was closely associated
with Sox2 expression, which suggests that β-catenin may be
associated with a higher malignant phenotype and EMT progression in
TSCC cells with alteration of Sox2 expression.
In conclusion, the present study illustrated the
pivotal role of Sox2 in the progression of TSCC. The results
provide an explanation for TSCC aggression and metastasis by
linking them with the overexpression of Sox2, and the possible
mechanism by which Sox2 may modulate EMT by targeting β-catenin.
These data can be extended in in viv and in viv
models to further elucidate the role of Sox2 in the carcinogenesis
and metastasis of TSCC. Hence, the study indicates that Sox2 can be
a predictive and therapeutic target for TSCC with aggressive
clinical behavior.
Acknowledgments
Not applicable.
Funding
This study was supported by Shandong Provincial
Natural Science Foundation (grant no. ZR2018PH027) the Natural
Science Foundation of Guangdong, China (grant no. 2014A030313153),
the Science and Technology Plan Projects of Guangzhou (grant no.
201510010268) and the National Natural Science Foundation of China,
with funding from the Youth Foundation of The First Affiliated
Hospital of Zhengzhou University (grant no. 81200796).
Availability of data and materials
The data and materials are available upon
request.
Authors' contributions
XL and QT conceived and designed the study. XL, BQ,
TZ and FH performed the experiments. XL and AKYL wrote the paper.
AKYL and QT reviewed and edited the manuscript. All authors read
and approved the manuscript.
Ethics approval and consent to
participate
Informed consent was obtained on the use of
surgically resected specimens for research purposes, according to
the guidelines for research for human tissues and samples set by
the Institution Review Board (IRB) of Sun Yat-sen University, who
also approved the study.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Siegel RL, Miller KD and Jemal A: Cancer
statistics, 2015. CA Cancer J Clin. 65:5–29. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Mao L, Hong WK and Papadimitrakopoulou VA:
Focus on head and neck cancer. Cancer Cell. 5:311–316. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Tsuji T, Ibaragi S and Hu GF:
Epithelial-mesenchymal transition and cell cooperativity in
metastasis. Cancer Res. 69:7135–7139. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Polyak K and Weinberg RA: Transitions
between epithelial and mesenchymal states: acquisition of malignant
and stem cell traits. Nat Rev Cancer. 9:265–273. 2009. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Zha L, Zhang J, Tang W, Zhang N, He M, Guo
Y and Wang Z: HMGA2 elicits EMT by activating the Wnt/β-catenin
pathway in gastric cancer. Dig Dis Sci. 58:724–733. 2013.
View Article : Google Scholar
|
|
6
|
Wu Y, Ginther C, Kim J, Mosher N, Chung S,
Slamon D and Vadgama JV: Expression of Wnt3 activates Wnt/β-catenin
pathway and promotes EMT-like phenotype in trastuzumab-resistant
HER2-overexpressing breast cancer cells. Mol Cancer Res.
10:1597–1606. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Lin LC, Hsu SL, Wu CL and Hsueh CM: TGFβ
can stimulate the p(38)/β-catenin/PPARγ signaling pathway to
promote the EMT, invasion and migration of non-small cell lung
cancer (H460 cells). Clin Exp Metastasis. 31:881–895. 2014.
View Article : Google Scholar : PubMed/NCBI
|
|
8
|
Kamachi Y, Uchikawa M and Kondoh H:
Pairing SOX off: with partners in the regulation of embryonic
development. Trends Genet. 16:182–187. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Chen Y, Shi L, Zhang L, Li R, Liang J, Yu
W, Sun L, Yang X, Wang Y, Zhang Y, et al: The molecular mechanism
governing the oncogenic potential of SOX2 in breast cancer. J Biol
Chem. 283:17969–17978. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Kregel S, Kiriluk KJ, Rosen AM, Cai Y,
Reyes EE, Otto KB, Tom W, Paner GP, Szmulewitz RZ and Vander Griend
DJ: Sox2 is an androgen receptor-repressed gene that promotes
castration-resistant prostate cancer. PLoS One. 8:e537012013.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Gangemi RM, Griffero F, Marubbi D, Perera
M, Capra MC, Malatesta P, Ravetti GL, Zona GL, Daga A and Corte G:
SOX2 silencing in glioblastoma tumor-initiating cells causes stop
of proliferation and loss of tumorigenicity. Stem Cells. 27:40–48.
2009. View Article : Google Scholar
|
|
12
|
Li X, Xu Y, Chen Y, Chen S, Jia X, Sun T,
Liu Y, Li X, Xiang R and Li N: SOX2 promotes tumor metastasis by
stimulating epithelial-to-mesenchymal transition via regulation of
WNT/β-catenin signal network. Cancer Lett. 336:379–389. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Kokalj Vokač N, Cizmarević B, Zagorac A,
Zagradišnik B and Lanišnik B: An evaluation of SOX2 and hTERC gene
amplifications as screening markers in oral and oropharyngeal
squamous cell carcinomas. Mol Cytogenet. 7:52014. View Article : Google Scholar
|
|
14
|
Michifuri Y, Hirohashi Y, Torigoe T,
Miyazaki A, Kobayashi J, Sasaki T, Fujino J, Asanuma H, Tamura Y,
Nakamori K, et al: High expression of ALDH1 and SOX2 diffuse
staining pattern of oral squamous cell carcinomas correlates to
lymph node metastasis. Pathol Int. 62:684–689. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Sobin LH and Wittekind C: TNM
classification of malignant tumours. 6th edition. New York:
Wiley-Liss; 2002
|
|
16
|
Qiao B, Gopalan V, Chen Z, Smith RA, Tao Q
and Lam AK: Epithelial-mesenchymal transition and
mesenchymal-epithelial transition are essential for the acquisition
of stem cell properties in hTERT-immortalised oral epithelial
cells. Biol Cell. 104:476–489. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Vermeire J, Naessens E, Vanderstraeten H,
Landi A, Iannucci V, Van Nuffel A, Taghon T, Pizzato M and
Verhasselt B: Quantification of reverse transcriptase activity by
real-time PCR as a fast and accurate method for titration of HIV,
lentiand retroviral vectors. PLoS One. 7:e508592012. View Article : Google Scholar
|
|
18
|
Chi AC, Day TA and Neville BW: Oral cavity
and oropharyngeal squamous cell carcinoma an update. CA Cancer J
Clin. 65:401–421. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Heerboth S, Housman G, Leary M, Longacre
M, Byler S, Lapinska K, Willbanks A and Sarkar S: EMT and tumor
metastasis. Clin Transl Med. 4:62015. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Wang J, Zhu X, Hu J, He G, Li X, Wu P, Ren
X, Wang F, Liao W, Liang L, et al: The positive feedback between
Snail and DAB2IP regulates EMT, invasion and metastasis in
colorectal cancer. Oncotarget. 6:27427–27439. 2015.PubMed/NCBI
|
|
21
|
Demirkan B: The roles of
epithelial-to-mesenchymal transition (EMT) and
mesenchymal-to-epithelial transition (MET) in breast cancer bone
metastasis: potential targets for prevention and treatment. J Clin
Med. 2:264–282. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Mulholland DJ, Kobayashi N, Ruscetti M,
Zhi A, Tran LM, Huang J, Gleave M and Wu H: Pte loss and RAS/MAPK
activation cooperate to promote EMT and metastasis initiated from
prostate cancer stem/progenitor cells. Cancer Res. 72:1878–1889.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Belotte J, Fletcher NM, Alexis M, Morris
RT, Munkarah AR, Diamond MP and Saed GM: Sox2 gene amplification
significantly impacts overall survival in serous epithelial ovarian
cancer. Reprod Sci. 22:38–46. 2015. View Article : Google Scholar :
|
|
24
|
Neumann J, Bahr F, Horst D, Kriegl L,
Engel J, Luque RM, Gerhard M, Kirchner T and Jung A: SOX2
expression correlates with lymph-node metastases and distant spread
in right-sided colon cancer. BMC Cancer. 11:5182011. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Yang F, Gao Y, Geng J, Qu D, Han Q, Qi J
and Chen G: Elevated expression of SOX2 and FGFR1 in correlation
with poor prognosis in patients with small cell lung cancer. Int J
Clin Exp Pathol. 6:2846–2854. 2013.PubMed/NCBI
|
|
26
|
Qiao B, He B, Cai J and Yang W: The
expression profile of Oct4 and Sox2 in the carcinogenesis of oral
mucosa. Int J Clin Exp Pathol. 7:28–37. 2013.
|
|
27
|
Nakayama S, Sasaki A, Mese H, Alcalde RE
and Matsumura T: Establishment of high and low metastasis cell
lines derived from a human tongue squamous cell carcinoma. Invasion
Metastasis. 18:219–228. 1999. View Article : Google Scholar
|
|
28
|
Kalluri R and Weinberg RA: The basics of
epithelial-mesenchymal transition. J Clin Invest. 119:1420–1428.
2009. View
Article : Google Scholar : PubMed/NCBI
|
|
29
|
Wang H, Zhang H, Tang L, Chen H, Wu C,
Zhao M, Yang Y, Chen X and Liu G: Resveratrol inhibits
TGF-β1-induced epithelial-to-mesenchymal transition and suppresses
lung cancer invasion and metastasis. Toxicology. 303:139–146. 2013.
View Article : Google Scholar
|
|
30
|
Micalizzi DS, Farabaugh SM and Ford HL:
Epithelial-mesenchymal transition in cancer: parallels between
normal development and tumor progression. J Mammary Gland Biol
Neoplasia. 15:117–134. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Han G, Lu SL, Li AG, He W, Corless CL,
Kulesz-Martin M and Wang XJ: Distinct mechanisms of
TGF-beta1-mediated epithelial-to-mesenchymal transition and
metastasis during skin carcinogenesis. J Clin Invest.
115:1714–1723. 2005. View
Article : Google Scholar : PubMed/NCBI
|
|
32
|
Graham TR, Zhau HE, Odero-Marah VA,
Osunkoya AO, Kimbro KS, Tighiouart M, Liu T, Simons JW and O'Regan
RM: Insulin-like growth factor-I-dependent up-regulation of ZEB1
drives epithelial-to-mesenchymal transition in human prostate
cancer cells. Cancer Res. 68:2479–2488. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
33
|
Lee MY, Chou CY, Tang MJ and Shen MR:
Epithelial-mesenchymal transition in cervical cancer: correlation
with tumor progression, epidermal growth factor receptor
overexpression, and snail up-regulation. Clin Cancer Res.
14:4743–4750. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Vesuna F, van Diest P, Chen JH and Raman
V: Twist is a transcriptional repressor of E-cadherin gene
expression in breast cancer. Biochem Biophys Res Commun.
367:235–241. 2008. View Article : Google Scholar
|
|
35
|
Aigner K, Dampier B, Descovich L, Mikula
M, Sultan A, Schreiber M, Mikulits W, Brabletz T, Strand D, Obrist
P, et al: The transcription factor ZEB1 (deltaEF1) promotes tumour
cell dedifferentiation by repressing master regulators of
epithelial polarity. Oncogene. 26:6979–6988. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Spaderna S, Schmalhofer O, Wahlbuhl M,
Dimmler A, Bauer K, Sultan A, Hlubek F, Jung A, Strand D, Eger A,
et al: The transcriptional repressor ZEB1 promotes metastasis and
loss of cell polarity in cancer. Cancer Res. 68:537–544. 2008.
View Article : Google Scholar : PubMed/NCBI
|
|
37
|
Ye X, Wu F, Wu C, Wang P, Jung K, Gopal K,
Ma Y, Li L and Lai R: β-catenin, a Sox2 binding partner, regulates
the DNA binding and transcriptional activity of Sox2 in breast
cancer cells. Cell Signal. 26:492–501. 2014. View Article : Google Scholar
|
|
38
|
Gen Y, Yasui K, Nishikawa T and Yoshikawa
T: SOX2 promotes tumor growth of esophageal squamous cell carcinoma
through the AKT/mammalian target of rapamycin complex 1 signaling
pathway. Cancer Sci. 104:810–816. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Bora-Singhal N, Perumal D, Nguyen J and
Chellappan S: Gli1-mediated regulation of Sox2 facilitates
self-renewal of stem-like cells and confers resistance to EGFR
inhibitors in non-small cell lung cancer. Neoplasia. 17:538–551.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Singh S, Trevino J, Bora-Singhal N,
Coppola D, Haura E, Altiok S and Chellappan SP: EGFR/Src/Akt
signaling modulates Sox2 expression and self-renewal of stem-like
side-population cells in non-small cell lung cancer. Mol Cancer.
11:732012. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Corominas-Faja B, Cufí S,
Oliveras-Ferraros C, Cuyàs E, López-Bonet E, Lupu R, Alarcón T,
Vellon L, Iglesias JM, Leis O, et al: Nuclear reprogramming of
luminal-like breast cancer cells generates Sox2-overexpressing
cancer stem-like cellular states harboring transcriptional
activation of the mTOR pathway. Cell Cycle. 12:3109–3124. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Bass AJ and Wang TC: An inflammatory
situation: SOX2 and STAT3 cooperate in squamous cell carcinoma
initiation. Cell Stem Cell. 12:266–268. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Yang N, Hui L, Wang Y, Yang H and Jiang X:
Overexpression of SOX2 promotes migration, invasion, and
epithelial-mesenchymal transition through the Wnt/β-catenin pathway
in laryngeal cancer Hep-2 cells. Tumour Biol. 35:7965–7973. 2014.
View Article : Google Scholar : PubMed/NCBI
|