Introduction
Cancer is a global health problem responsible for
one in four mortalities worldwide (1,2).
According to the World Health Organization, breast cancer accounts
for approximately 1/3 of all cancer cases diagnosed in women
(3). There are a number of risk
factors for breast cancer, including hereditary factors, abnormal
hormone levels, smoking history and alcohol consumption (4,5).
Breast cancer is typically treated using chemotherapy, biological
immune treatments and traditional Chinese medicine, however, there
is no definitive therapeutic target for breast cancer treatment
(6). The treatment options used
currently are accompanied by a number of side effects, and thus a
great deal of research has been dedicated to identifying novel
drugs with the therapeutic potential in breast cancer in recent
years (4). However, the
pathogenesis of breast cancer remains unknown (6). Much attention had been given to the
medicinal value of natural compounds, and plant-based drugs have
been successfully used in the treatment of cancer and other
diseases (7-9). Herbal drugs, such as phenolic acids,
flavonoids and sesquiterpene, have been demonstrated to be
cytotoxic to cancer cells, acting via a range of mechanisms
(4).
Inula Helenium L. is a flower of the
Compositae herbaceous family, whose roots have historically been
used as a medicine (10). This
plant has been investigated since the 1970s, and certain studies
have reported that it contains >40 different compounds, with the
terpenoids being the main components (11-13). Alantolactone is a sesquiterpene
lactone extracted from Inula Helenium L. that exhibits a
number of biological effects, including anti-inflammatory,
antibacterial and anticancer activities (14,15). It has previously been reported
that alantolactone induces apoptosis in a number of cancer cell
lines, including in the colorectal cancer RKO cell line via the
mitogen-activated protein kinase (MAPK) pathway (10), and in the liver cancer HepG2 cell
line via regulating the nuclear factor (NF)-κB signaling pathway
(15). Alantolactone has also
been demonstrated to inhibit cell cycle progression in SK-MES-1
cells (16). This compound may
therefore be a promising candidate as a chemotherapeutic drug for
cancer therapy (17).
Although the effects of alantolactone have been
studied in a variety of human cancer cell lines, the potential
anticancer activity of alantolactone in human breast cancer remains
unclear. The aim of the present study was to investigate the
function and molecular mechanisms of alantolactone in repressing
cell proliferation and inducing apoptosis in MCF-7 cells, in order
to provide a theoretical basis for its use as a clinical cancer
treatment.
Materials and methods
Chemicals and materials
Alantolactone (purity, 99%) was purchased from
Chengdu Must Bio-Technology Co., Ltd. (Chengdu, China). Acrylamide,
penicillin, streptomycin, phosphate-buffered saline (PBS), Carnoy's
solution, Giemsa, BCA kit, enhanced chemiluminescence (ECL) and
dimethyl sulfoxide (DMSO) were obtained from Beijing Dingguo
Changsheng Biotechnology Co., Ltd. (Beijing, China). Hoechst 33258
staining kit and Annexin V-FITC cell apoptosis assay kits were
purchased from Nanjing KeyGen Biotech Co., Ltd. (Nanjing, China).
Paraformaldehyde (4%), and antibodies against B-cell lymphoma 2
(Bcl-2; cat. no. sc-509), Bcl-2-associated X protein (Bax; cat. no.
sc-23959), p53 (cat. no. sc-71820), matrix metalloproteinase
(MMP)-2 (cat. no. sc-13594), MMP-7 (cat. no. sc-80205), MMP-9 (cat.
no. sc-21733), nuclear factor erythroid 2-related factor 2 (Nrf2;
cat. no. sc-518033), proliferating cell nuclear antigen (PCNA; cat.
no. sc-25280), caspase-3 (cat. no. sc-271028) and caspase-12 (cat.
no. sc-21747) were obtained from Santa Cruz Biotechnology, Inc.
(Dallas, TX, USA). Anti-p38 (cat. no. sc-271028), anti-phospho-p38
(cat. no. 9215), phospho-inhibitor of NF-κB (p-IκBα; cat. no.
2859), IκBά (cat. no. 9247), anti-NF-κB (p65) antibodies (cat. no.
8242) and β-actin (cat. no. 4970S) were purchased from Cell
Signaling Technology, Inc. (Danvers, MA, USA). Fetal bovine serum,
radioimmunoprecipiation (RIPA) lysis buffer, nuclear protein
extraction kit, horseradish peroxidase (HRP)-conjugated goat
anti-rabbit IgG (cat. no. SE134) and HRP-conjugated goat anti-mouse
IgG secondary antibodies (cat. no. SE131) were obtained from
Beijing Solarbio Science & Technology Co., Ltd. (Beijing,
China). Plates (6 cm in diameter) were obtained from Corning
Incorporated (Corning, NY, USA). Dulbecco's modified Eagle's medium
(DMEM) was purchased from Gibco (Thermo Fisher Scientific. Inc.,
Waltham, MA, USA).
Cell culture
The human breast cancer cell line MCF-7 was obtained
from the Cell Bank of Type Culture Collection of Chinese Academy of
Sciences (Shanghai, China), and cultured in Dulbecco's modified
Eagle's medium containing 10% (v/v) fetal bovine serum, 100 U/ml
penicillin and 100 μg/ml streptomycin. MCF-7 cells were
maintained at 37°C in an incubator with 5% CO2.
Subculture was performed when cells reached 80-90% confluence
(18).
Cell proliferation assay
The harvested cells were seeded at a density of
3×103 cells/well in a 96-well plate. MCF-7 cells were
treated with various concentrations of alantolactone or with 10
mg/ml fluorouracil (5-Fu) for 24 and 48 h, following which cells
were stained with 20 μ1 MTT solution for 4 h at 37°C. The
medium was removed from each well, and the purple formazan crystals
were dissolved in 150 μ1 DMSO for 10 min using vibration.
Absorbance was assessed at 490 nm using a microplate reader
(19). 5-Fu was used to determine
the inhibitory effect of alantolactone on MCF-7 cells. Cells were
observed and images were captured under an Olympus CX22LED
microscope (magnification, ×10; Olympus Corporation, Tokyo, Japan)
at 24 and 48 h, the adherent cells were counted. All measurements
were performed at least in triplicate. MCF-7 cells treated only
with cell medium were used as the control.
Apoptosis assay
Annexin V-FITC/PI double staining was used to
determine the percentage of apoptotic MCF-7 cells. Briefly, cells
were treated with 10, 20 and 30 μM alantolactone for 24 h
and washed twice with PBS, then, 5 μ1 Annexin V-FITC and PI
were added to the cells for 15 min (20). The percentage of apoptotic MCF-7
cells was assessed using flow cytometry (FACSCalibur; BD
Biosciences, Franklin Lakes, NJ, USA). MCF-7 cells treated only
with cell medium were used as the control.
Hoechst 33258 fluorescence staining
MCF-7 cells were seeded at a density of
8×104 cells/well in 12-well plates, then the cells were
treated with 10, 20 and 30 μM alantolactone for 24 h. Next,
cells were obtained and fixed in 4% paraformaldehyde solution for
30 min. The cells were then washed twice with Buffer A from a
Hoechst 33258 staining kit, and stained with 30 μl Hoechst
33258 for 10 min in the dark and observed under a fluorescence
microscope (magnification, ×20) (21). MCF-7 cells treated only with cell
medium were used as the control.
Wound-healing assay
When the cells reached 80-90% confluence, the cell
layer was scratched with a 10 μl sterile pipette tip and the
wells were washed twice with PBS. Subsequently, 10, 20 and 30
μM alantolactone was added to the medium, and cell migration
was observed under a microscope (magnification, ×10) at 24 and 48
h. MCF-7 cells treated only with cell medium were used as the
control.
Colony-forming assay
Cells were seeded at a density of 1×103
cells/plate and cultured with 5 ml medium for 24 h. Next, cells
treated with 10, 20 and 30 μM alantolactone were left to
cultivate for 15 days until visible colonies formed (22). Following fixation with Carnoy's
solution for 30 min and staining with 5 ml Giemsa stain for 10 min,
the number of cell colonies was counted. MCF-7 cells treated only
with cell medium were used as the control.
Western blot assay
Following treatment with alantolactone for 24 h,
cells were harvested and lysed with a RIPA lysis buffer for total
protein and the nuclear protein was prepared with nuclear protein
extraction kit. Protein concentrations were measured using a BCA
protein assay. Equal amounts of protein were separated by 12%
SDS-PAGE and transferred to PVDF membranes. The membranes were
blocked with 5% non-fat milk for 1 h at room temperature, and then
incubated with the primary antibodies (1:2,000) overnight at 4°C,
followed by incubation with the secondary antibodies for 1 h at
room temperature. Finally, bands were observed using an enhanced
chemiluminescence kit. The relative level of total protein was
normalized to β-actin and the relative level of nuclear protein was
normalized to PCNA. The gray intensity was analyzed with ImageJ
software (National Institutes of Health, Bethesda, MD, USA). MCF-7
cells treated only with cell medium were used as control.
Statistical analysis
Data are presented as the mean ± standard
deviation of at least three independent experiments. Statistical
analyses were performed using one-way analysis of variance with
SPSS software, version 20.0 (IBM SPSS, Armonk, NY, USA).
Comparisons between groups were assessed using a post hoc Tukey's
test and correlation analysis. P<0.05 was considered to indicate
a statistically significant difference.
Results
Effect of alantolactone on cell
morphology and viability
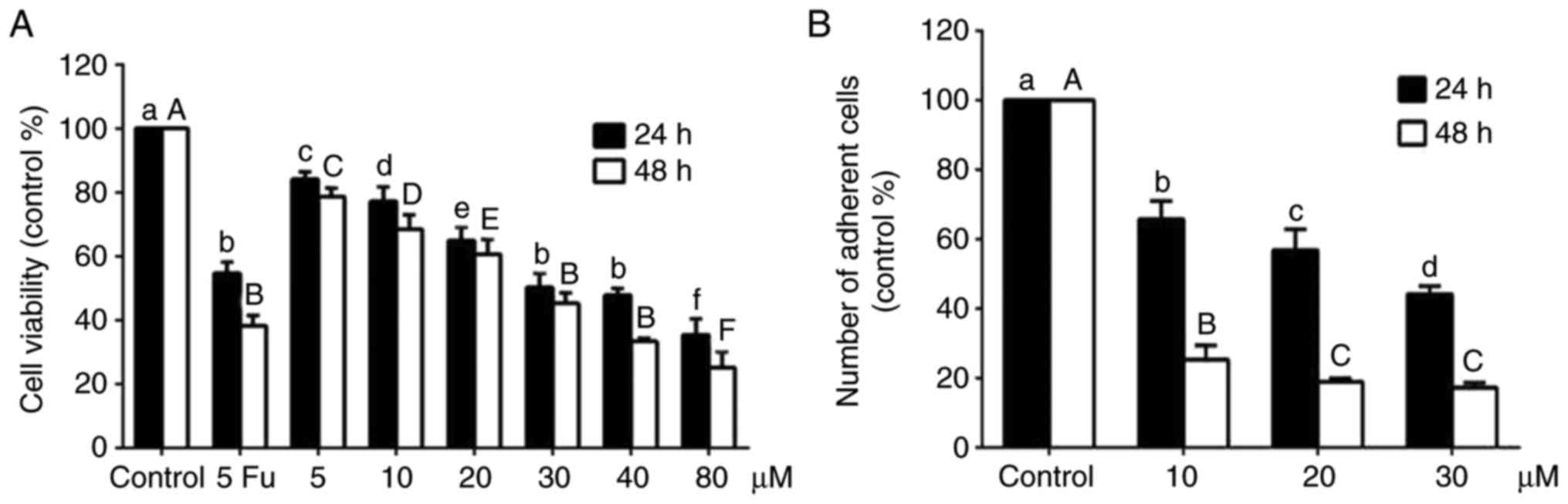
An MTT assay was used to further verify the
inhibitory effect of alantolactone on cell proliferation. As shown
in Fig. 1A, treatment with
different concentrations of alantolactone (5, 10, 20, 30, 40 and 80
μΜ) for 24 and 48 h significantly reduced the cell viability
in a concentration- and time-dependent manner compared with the
control group and the cell viability at 40 μΜ was similar to
the group treated with 5-Fu. The half maximal inhibitory
concentration (IC50) was 35.45 μΜ at 24 h and
24.29 μΜ at 48 h. The number of adherent cells decreased
with alantolactone treatment in a concentration- and time-dependent
manner (Fig. 1B). Therefore, the
concentrations of 10, 20 and 30 μΜ and the treatment time of
24 h were selected for subsequent experiments.
Alantolactone induces apoptosis in MCF-7
cells
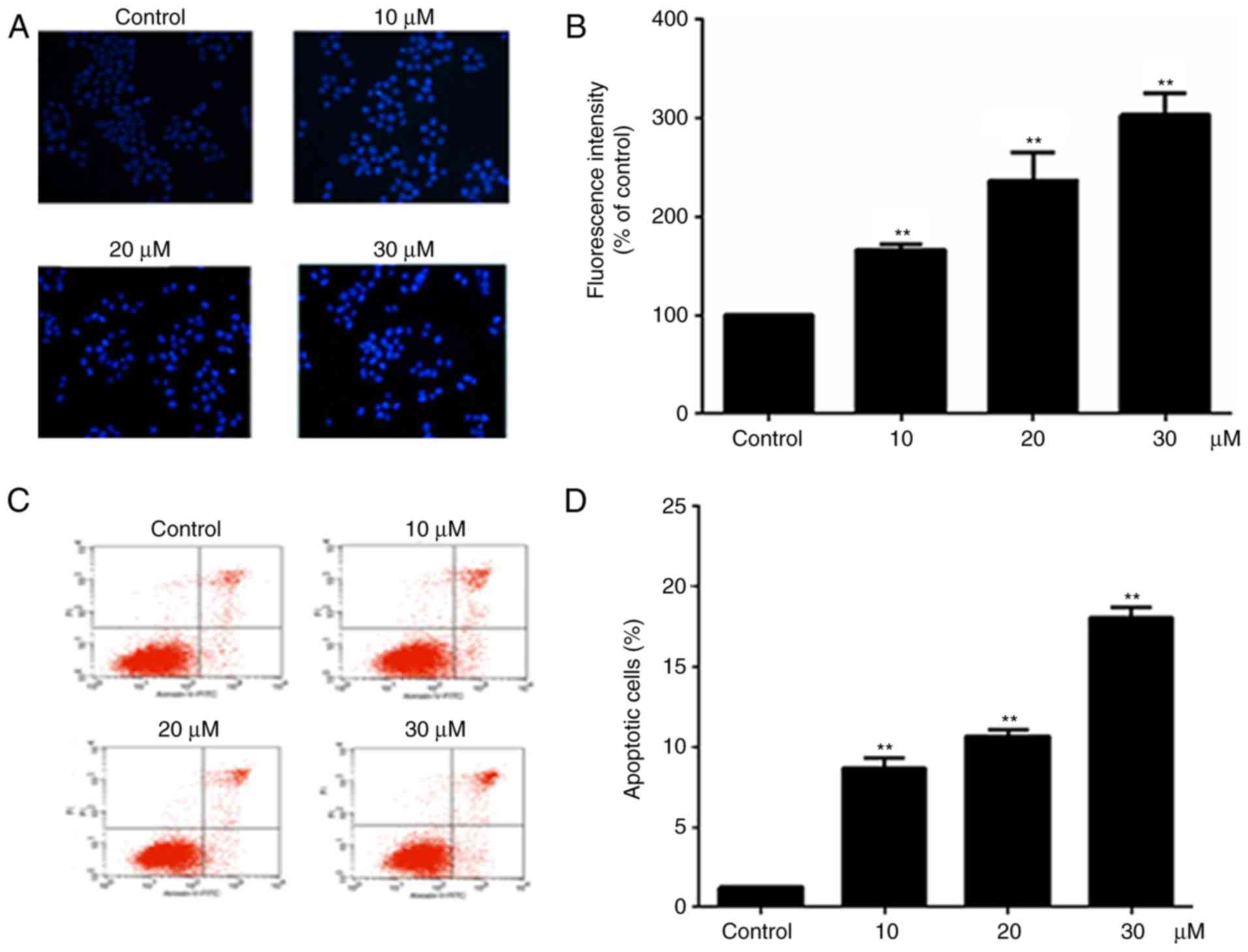
Hoechst 33258 staining was used to examine the cell
apoptosis, since the nuclei of apoptotic cells exhibit fluorescent
blue staining, whereas live cells have uniformly light blue nuclei.
Cells were treated with different concentration of alantolactone
(10, 20 and 30 μΜ) for 24 h, and the number of live cells
was reduced as the concentration of alantolactone increased. The
treated cells exhibited brighter fluorescence in comparison with
the control cells, and the fluorescence intensity of treated cells
decreased significantly as the concentration of alantolactone
increased (Fig. 2B). Furthermore,
the percentage of apoptotic cells was determined using the Annexin
V-FITC/PI double staining method. It was observed that the
percentage of apoptotic cells was increased following treatment
with alantolactone for 24 h in a concentration-dependent manner
compared with that observed in the control cells (Fig. 2C and D).
Alantolactone inhibits the migration of
MCF-7 cells
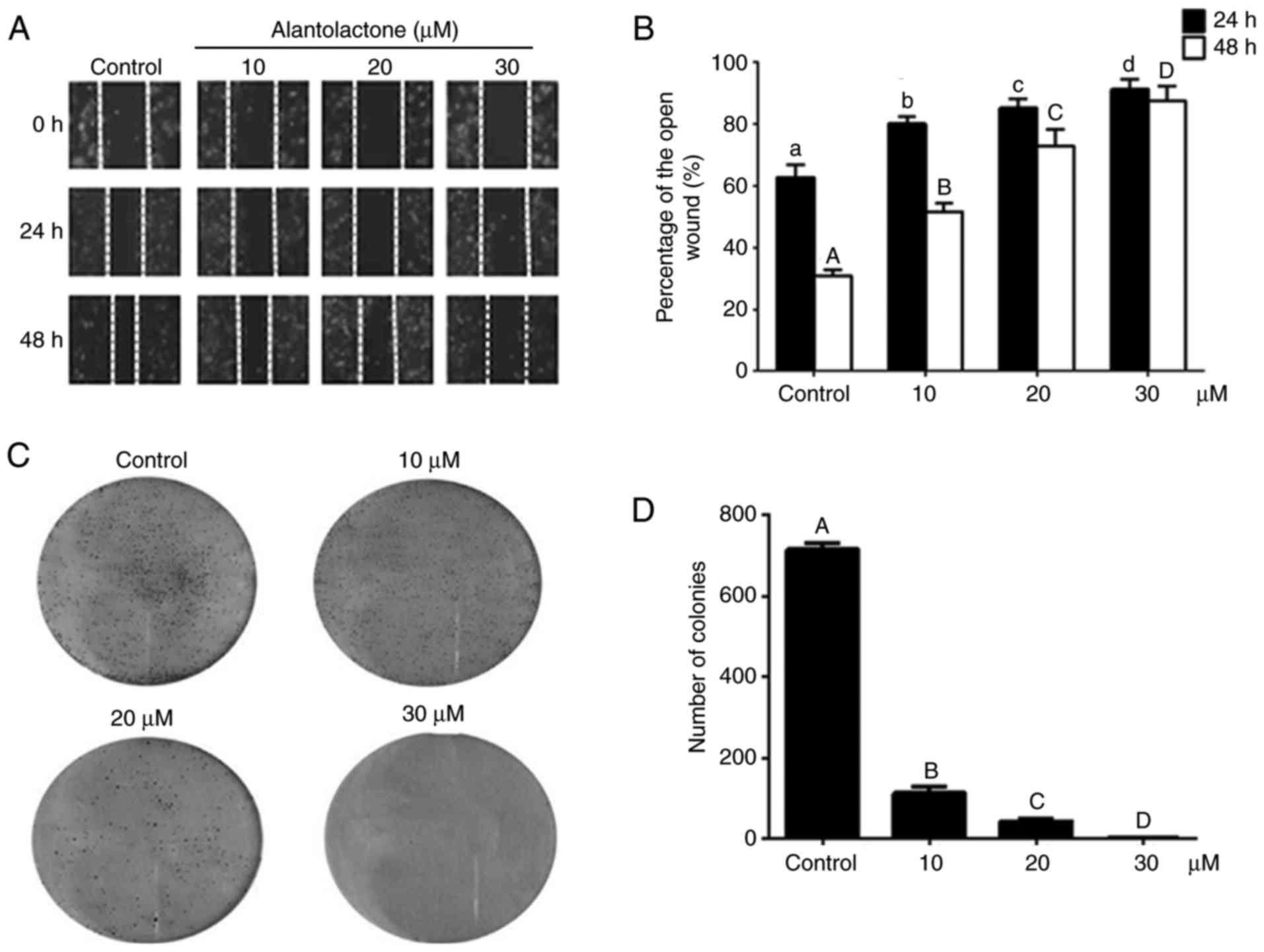
The effect of alantolactone on MCF-7 cell migration
was analyzed using a wound healing assay. Compared with the control
group, alantolactone significantly decreased cell migration in a
dose- and time-dependent manner (P<0.05; Fig. 3A and B).
Colony-forming assay
Compared with the control cells, colony formation
was significantly lower in cells treated with alantolactone
(P<0.01). The colony formation was inhibited by alantolactone in
a concentration-dependent manner (Fig. 3C and D).
Expression levels of apoptosis- and
migration-associated proteins
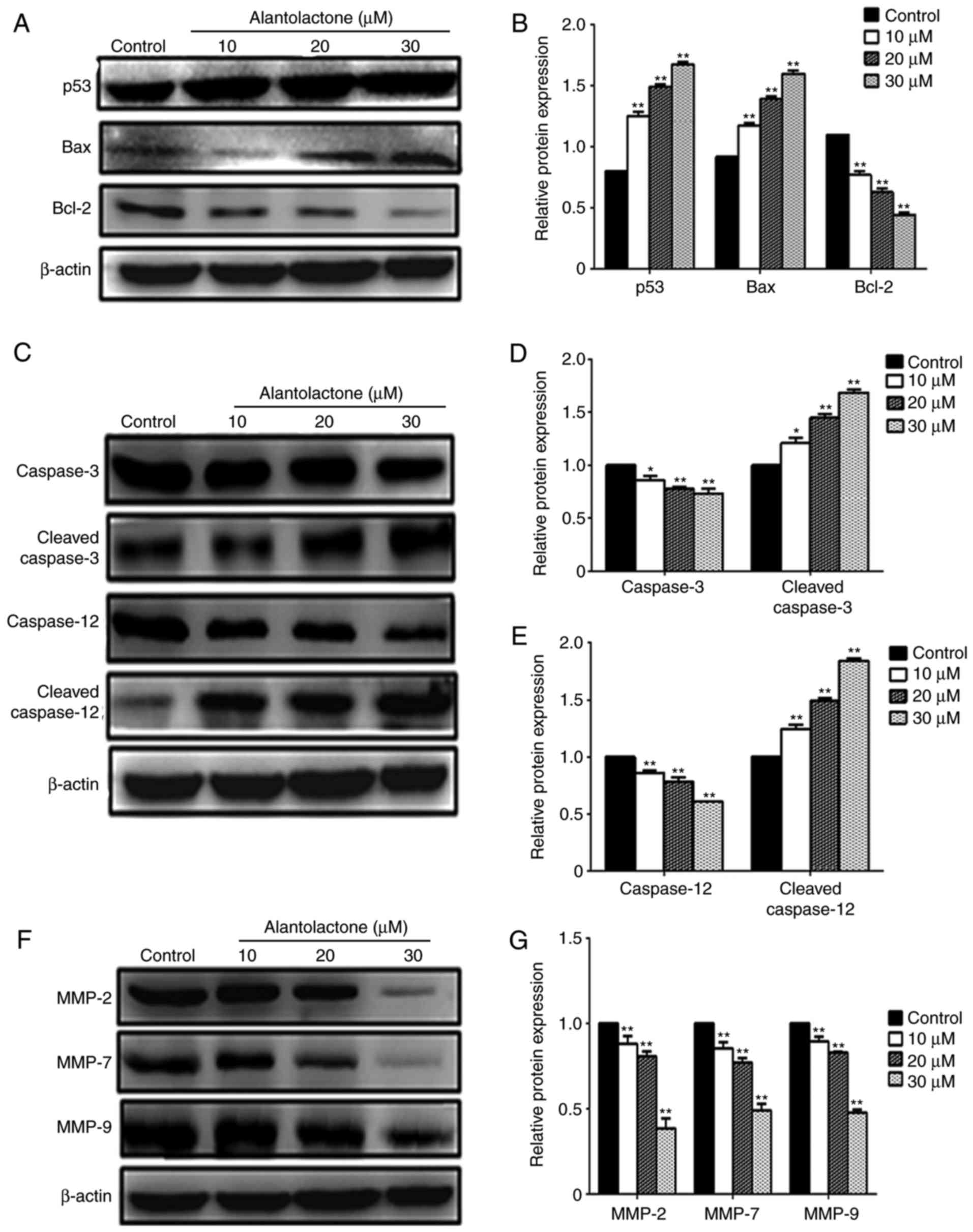
Following treatment with alantolactone, the
expression levels of p53, Bax and Bcl-2 protein in MCF-7 cells were
measured using western blotting. Treatment with 10, 20 or 30
μΜ alantolactone significantly reduced the expression of
Bcl-2 and significantly increased the expression levels of Bax and
p53 compared with those in control cells (Fig. 4A and B). Alantolactone also
decreased the expression of the caspase precursor and significantly
enhanced the expression of cleaved-caspase-3 and cleaved-caspase-12
in a concentration-dependent manner (P<0.05 and P<0.01;
Fig. 4C–E). In addition,
alantolactone at 10, 20 and 30 μΜ significantly
downregulated the expression levels of MMP-2, MMP-7 and MMP-9
protein (Fig. 4F and G).
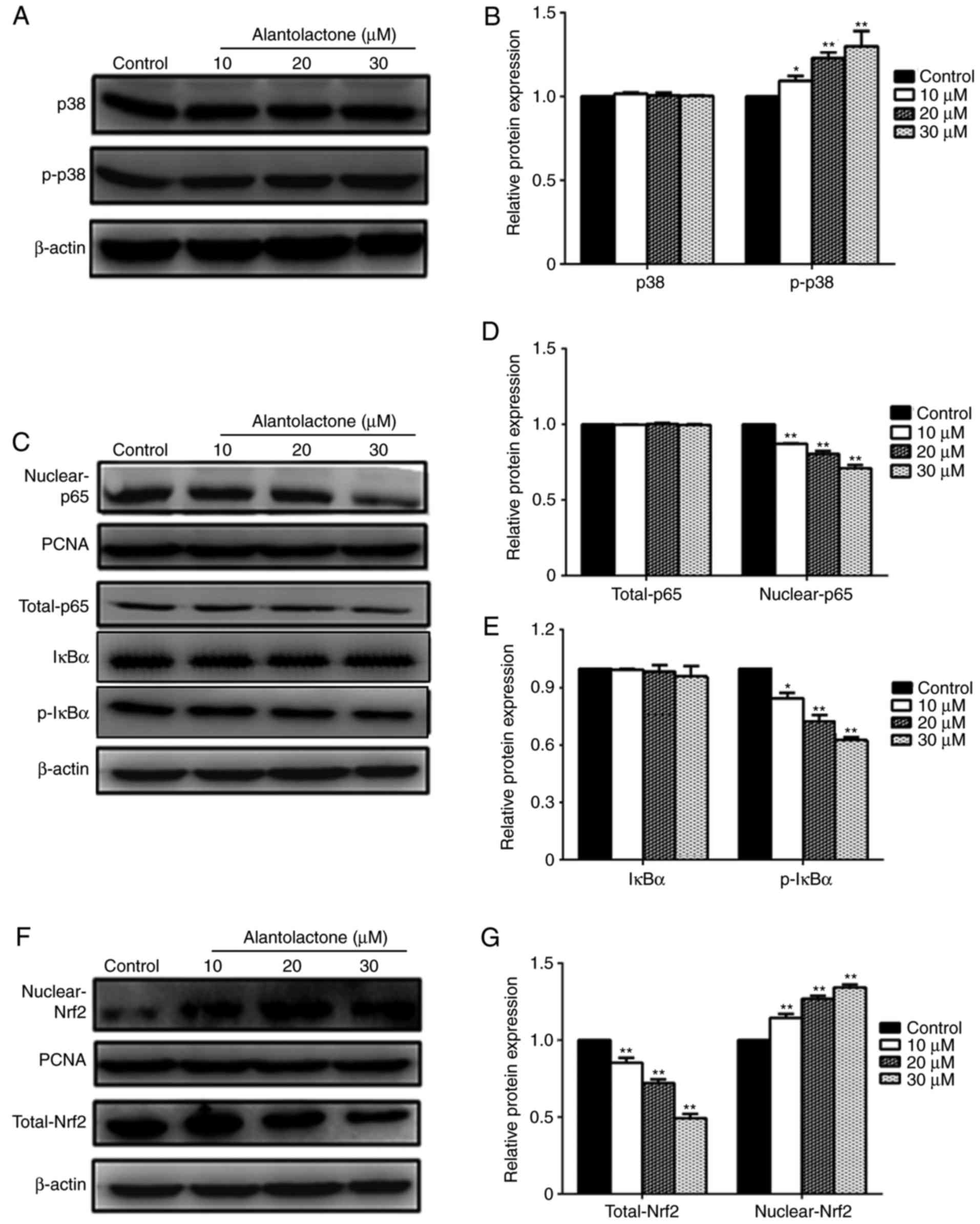
Effect of alantolactone on the p38 MAPK
signaling pathway
To further explore the mechanism by which
alantolactone affects MCF-7 cells, the expression levels of p38 and
p-p38 protein were measured. The results revealed that the
expression of p-p38 was significantly increased by alantolactone in
a dose-dependent manner, while alantolactone had no evident effect
on the expression of p38 (Fig. 5A and
B).
Effect of alantolactone on the NF-κB
signaling pathway
The expression of p65 protein was measured to
explore the role of the NF-κB pathway in alantolactone-mediated
MCF-7 cell damage. Treatment with 10, 20 and 30 μΜ
alantolactone significantly downregulated the nuclear expression of
p65 (Fig. 5C and D). However, at
these concentrations, alantolactone had no significant effect on
the total expression of p65. The expression of p-ΙκΒα was
significantly decreased following treatment with 10, 20 and 30
μM alantolactone, whereas no marked effects on ΙκΒα
expression were observed (Fig. 5C and
E).
Nrf2 signaling pathway serves a role in
alantolactone-regulated proliferation
To investigate the role of the Nrf2 pathway in
alantolactone-mediated MCF-7 cell damage, the expression of Nrf2
protein was measured. As shown in Fig. 5F and G, treatment with 10, 20 and
30 μM alantolactone significantly upregulated the nuclear
expression of Nrf2, while the total Nrf2 expression was
significantly downregulated.
Discussion
Breast cancer is known to be one of the most common
malignant types of cancer affecting women, and the morbidity of
breast cancer has been increasing since the 1970s (14,21). Recently, a great deal of attention
has been paid to the potential of natural compounds as novel
treatments for cancer (5).
Pharmacological agents extracted from plants have been reported to
have good therapeutic effects in the treatment and prevention of
cancer (23). Alantolactone, a
sesquiterpene lactone compound extracted from Inula Helenium
L, had been reported to have anticancer activities (24). In the present study, the
biological effect of alantolactone on MCF-7 cells was investigated
by measuring the levels of apoptosis markers, colony formation and
migration.
To investigate whether alantolactone is able to
inhibit MCF-7 cell growth, the cell morphology was observed and
cell viability was measured. Alantolactone significantly reduced
the viability of MCF-7 cells (Fig.
1A), with an IC50 of 35.45 μM at 24 h and
24.29 μM at 48 h, which indicated that it is a promising
compound for clinical application. Previously, Kumari et al
(25) reported that the
IC50 value of coralyne was 76.4±0.92 μM in
MCF-7 cells for 24 h. Nikhil et al (26) also reported that the
IC50 value of pterostilbene was 65±0.42 μM
in MCF-7 cells for 24 h. Furthermore, tangeretin inhibited the
proliferation of MCF-7 cells, and the IC50 value of
tangeretin was 39.3±1.5 μM (27). Compared with these natural
products, alantolactone is more effective as the IC50
value was lower (25-27).
Changes in the balance between cell proliferation
and apoptosis serve a role in a number of diseases (28). Three types of cell death occur,
including autophagy, apoptosis and cell necrosis (29). Apoptosis serves a vital role in
the evolution of organisms, the stability of internal environments
and the development of multiple systems, particularly in cancer
development (30). Cancer occurs
as a result of insufficient apoptosis (31), and thus apoptosis is a common
target for a number of anticancer treatments (32). Alantolactone has been reported to
induce apoptosis in various cancer cell lines (33). In the present study, Hoechst 33258
and Annexin V/PI staining were used to detect cell apoptosis, and
the results demonstrated that alantolactone significantly increased
the percentage of apoptotic MCF-7 cells (Fig. 2), suggesting that alantolactone
induces apoptosis in human breast cancer cells. Apoptosis occurs
via the extrinsic or intrinsic pathways in mammalian cells, and
mitochondria serve an important role in the intrinsic apoptotic
process (34). The mitochondrial
apoptotic pathway is controlled by the Bcl-2 family proteins,
including pro-apoptotic and anti-apoptotic proteins, such as Bax
and Bcl-2 (35). Alantolactone is
able to induce the apoptosis of HepG2 cells via modulating Bcl-2
family proteins (15). A similar
trend was observed in the present study. The results shown in
Fig. 4A revealed that
alantolactone significantly downregulated the expression of Bcl-2
and significantly upregulated the expression of Bax, suggesting
that alantolactone induces apoptosis via the mitochondrial
apoptotic pathway. In addition, p53 is critical in the evolution
from normal cellular function to tumorigenesis and has been
identified as a common mutated cancer suppressor in human
tumorigenesis (36). In the
present study, p53 expression was increased following treatment
with alantolactone, suggesting that p53 may serve an important role
in alantolactone-induced MCF-7 cell apoptosis via the cellular
apoptotic pathway. The cellular apoptotic pathway is mediated by
caspase family proteins, including caspase-3 and cleaved-caspase-3,
as well as caspase-12 and cleaved-caspase-12. Alantolactone has the
ability to induce apoptosis in HepG2 cells via modulating caspase
family proteins (37). The
current study results demonstrated that alantolactone significantly
enhanced the expression levels of cleaved-caspase-3 and
cleaved-caspase-12 proteins. However, the effect of alantolactone
on the caspase precursor was weak, suggesting that alantolactone
induces cell apoptosis via the apoptotic cellular pathway (Fig. 4C).
Chemotherapy is a commonly used clinical treatment
for cancer, however, the risk of recurrence and metastasis remains
a problem in patients with breast cancer (38). The majority of cancer-associated
mortalities occur as a result of metastatic cancer and tumor growth
at distant sites (39).
Therefore, the migration and invasion inhibiting effects of
plant-based drugs may serve an important role in cancer treatment
(40). To further evaluate the
anticancer effect of alantolactone in MCF-7 cells, colony formation
and migration were assessed in the present study. The results
revealed that alantolactone significantly inhibited colony
formation and migration in breast cancer cells. MMPs, a major
proteinase family associated with tumorigenesis, are key kinases in
cell migration during invasive and metastatic processes (4). A number of studies have reported
that MMP-2, MMP-7 and MMP-9 are able to degrade the basement
membrane and extracellular matrix (18). Therefore, to further investigate
the inhibitive effect of alantolactone on the migration and
invasion of breast cancer cells, the current study measured the
expression levels of MMP-2, MMP-7 and MMP-9. The results (Fig. 4F) revealed that alantolactone
significantly downregulated MMP-2, MMP-7 and MMP-9 in MCF-7 cells,
and blocked cell migration and invasion.
The pathogenic mechanisms of cancer include changes
to signal transduction pathways. As such, molecules involved in
abnormal signaling pathways may be targets for cancer treatments
(2). MAPK is an important signal
transduction pathway that regulates a number of physiological
processes and serves an important role in the induction of cell
damage (40). Three major MAPK
signaling pathways have been identified, including extracellular
signal-regulated kinase, c-Jun N-terminal kinase and p38 MAPK
pathways (41). MAPK activation
leads to the phosphorylation of p38, activates transcription
factors and promotes apoptosis (22). In addition, p38 MAPK has been
identified as the vital signaling pathway that activates Bax
subsequent to its translocation to the mitochondria, and p38 MAPK
signaling pathways are responsible for cell proliferation (42). In the current study (Fig. 5A), alantolactone significantly
upregulated the phosphorylation of p38, suggesting that the
anticancer effects of alantolactone in MCF-7 cells may occur
partially via regulating the p38 MAPK pathway.
MAPKs are closely associated with a number of other
signaling pathways, including NF-κB (p65), which is a downstream
target of p38 MAPK. NF-κB is regarded as an important factor in
physiological and pathological processes, including proliferation
and apoptosis, and a key regulator of oncogenesis (7). Normally, NF-κB is retained in the
cytoplasm in its inactive form, and is released and translocated to
the nucleus when activated. A number of carcinogens are able to
activate the NF-κB signaling pathway, and activated p65 blocks
apoptosis and promotes proliferation (43). It also had been reported that
NF-κB activation is associated with resistance to various
chemotherapeutic agents (44). In
the present study, it was demonstrated that alantolactone
significantly downregulated nuclear NF-κB in cancer cells and
downregulated p-IκBα and its phosphorylation. The results (Fig. 5C) also revealed that alantolactone
induces apoptosis in part by blocking the translocation of NF-κB
from the cytoplasm into the nucleus and promoting the
phosphorylation of IκBα
Nrf2 is an essential signaling molecule that serves
a role in the biological defense mechanism. As a central
transcription factor, Nrf2 is also involved in the suppression of
tumorigenesis, and Nrf2 upregulation is associated with treatment
resistance in cancer (45). In
the present study, the results (Fig.
5F) demonstrated that the expression of Nrf2 was significantly
increased in the nucleus. These findings suggest that alantolactone
promotes the translocation of Nrf2 from the cytoplasm into the
nucleus, and that Nrf2 is a risk factor in breast cancer that may
be considered to be a promising target in cancer diagnosis. In
addition, the results suggested that alantolactone modulates the
NF-κB signaling pathway, resulting in P-IκB downregulation and
reduced migration of MCF-7 cells. Cell death is induced via the p38
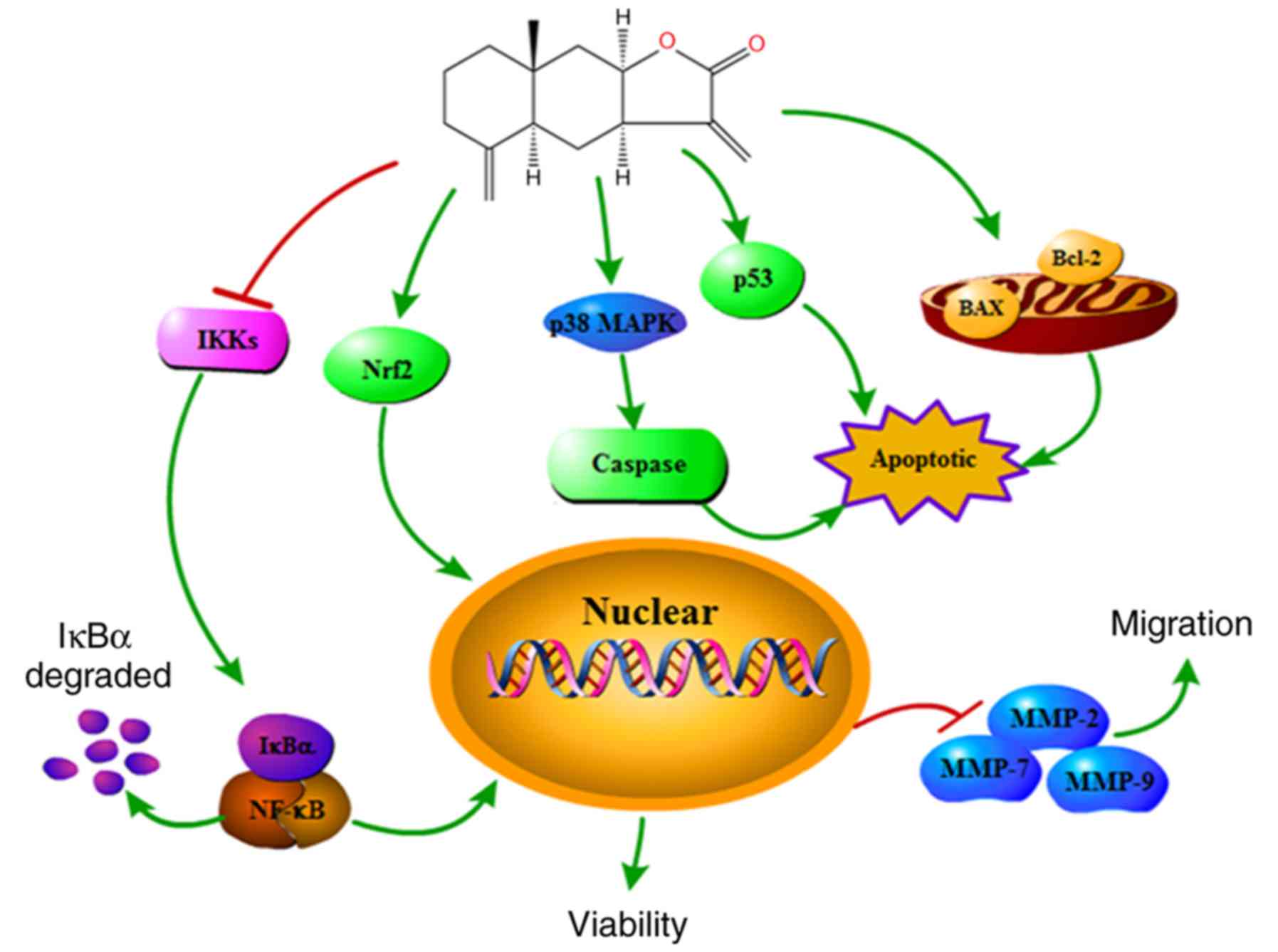
MAPK and Nrf2 pathways in MCF-7 cells (Fig. 6).
In conclusion, the present study demonstrated that
alantolactone has anticancer effects in human breast cancer cells.
The results confirmed that alantolactone is able to inhibit
proliferation and apoptosis, as well as to suppress colony
formation and cell migration. In addition, the mechanism by which
alantolactone acts may involve the p38 MAPK, NF-κB or Nrf2
signaling pathways. Taken together, the results of the present
study suggested that alantolactone is an antineoplastic drug, and
these findings may be used as a clinical basis for the application
of alantolactone as a novel cancer treatment.
Acknowledgments
Not applicable.
Funding
The present study was supported by the National
Natural Science Foundation of china (grant no. 31770017), the
cultivation Plan for Youth Agricultural Science and Technology
Innovative Talents of Liaoning Province (grant no. 2015013), the
Project Supported by Scientific Research Fund of Liaoning
Provincial Education Department (grant no. LQN201714), the Startup
Foundation for doctors of Liaoning Province (grant no. 20170520258)
and the Innovation Team Project from the Education department of
Liaoning Province (grant no. LT2015011).
Availability of data and materials
The datasets used and/or analyzed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
JLL, MJL and XYC conceived and designed the project
and prepared the manuscript. JLL, SW, YH and ZJY conducted the cell
experiments. MJL and YPH analyzed the data. All authors read and
approved the manuscript.
Ethics approval and consent to
participate
Not applicable.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Chung TW, Choi H, Lee JM, Ha SH, Kwak CH,
Abekura F, Park JY, Chang YC, Ha KT, Cho SH, et al: Oldenlandia
diffusa suppresses metastatic potential through inhibiting matrix
metalloproteinase-9 and intercellular adhesion molecule-1
expression via p38 and ERK1/2 MAPK pathways and induces apoptosis
in human breast cancer mcf-7 cells. J Ethnopharmacol. 195:309–317.
2017. View Article : Google Scholar
|
|
2
|
Jiang X, Li T and Liu RH:
2α-Hydroxyursolic acid inhibited cell proliferation and induced
apoptosis in MDA-MB-231 human breast cancer cells through the
p38/MAPK signal transduction pathway. J Agric Food Chem.
64:1806–1816. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Kreike B, Kouwenhove MV, Horlings H,
Weigelt B, Peterse H, Bartelink H and van de Vijver MJ: Gene
expression profiling and histopathological characterization of
triple-negative/basal-like breast carcinomas. Breast Cancer
Research. 9:R652007. View
Article : Google Scholar : PubMed/NCBI
|
|
4
|
Liao YF, Rao YK and Tzeng YM: Aqueous
extract of Anisomeles indica and its purified compound exerts
anti-metastatic activity through inhibition of NF-κB/AP-1-dependent
MMP-9 activation in human breast cancer MCF-7 cells. Food Chem
Toxicol. 50:2930–2936. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Elumalai P, Gunadharini DN, Senthilkumar
K, Banudevi S, Arunkumar R, Benson CS, Sharmila G and Arunakaran J:
Induction of apoptosis in human breast cancer cells by nimbolide
through extrinsic and intrinsic pathway. Toxicol Lett. 215:131–142.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Tan PQ, Zhong YM, Hu ZY and Lou DM: Size
distributions, PAHs and inorganic ions of exhaust particles from a
heavy duty diesel engine using B20 biodiesel with different exhaust
aftertreatments. Energy. 141:898–906. 2017. View Article : Google Scholar
|
|
7
|
Patel DK, Kumar R, Laloo D and Hemalatha
S: Natural medicines from plant source used for therapy of diabetes
mellitus: An overview of its pharmacological aspects. Asian Pac J
Trop D. 2:239–250. 2012. View Article : Google Scholar
|
|
8
|
Wu T, Geng J, Guo W, Gao J and Zhu X:
Asiatic acid inhibits lung cancer cell growth in vitro and in vivo
by destroying mitochondria. Acta Pharm Sin B. 7:65–72. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Luo W, Liu Y, Zhang J, Luo X, Lin C and
Guo J: Andrographolide inhibits the activation of NF-κB and MMP-9
activity in H3255 lung cancer cells. Exp Ther Med. 6:743–746. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Zhang Y, Bao YL, Wu Y, Yu CL, Huang YX,
Sun Y, Zheng LH and Li YX: Alantolactone induces apoptosis in RKO
cells through the generation of reactive oxygen species and the
mitochondrial pathway. Mol Med Rep. 8:967–972. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Huo Y, Shi HM, Li WW, Wang MY and Li XB:
HPLC determination and NMR structural elucidation of sesquiterpene
lactones in. Inula Helenium J Pharm Biomed Anal. 51:942–946. 2010.
View Article : Google Scholar
|
|
12
|
Konishi T, Shimada Y, Nagao T, Okabe H and
Konoshima T: Antiproliferative sesquiterpene lactones from the
roots of. Inula Helenium Biol Pharm Bull. 25:1370–1372. 2002.
View Article : Google Scholar
|
|
13
|
Li Y, Ni ZY, Zhu MC, Dong M, Wang SM, Shi
QW, Zhang ML, Wang YF, Huo CH, Kiyota H and Cong B: Antitumour
activities of sesquiterpene lactones from Inula Helenium and Inula
japonica. Z Naturforsch C. 67:375–380. 2012.PubMed/NCBI
|
|
14
|
Chun J and Kim YS: Alantolactone, a
sesquiterpene lactone isolated from Inula Helenium L. selectively
suppresses STAT3 activation and exhibits anticancer activity in
MDA-MB-231 cells. Planta Med. 81:2015. View Article : Google Scholar
|
|
15
|
Khan M, Li T, Ahmad Khan MK, Rasul A,
Nawaz F, Sun M, Zheng Y and Ma T: Alantolactone induces apoptosis
in HepG2 cells through GSH depletion, inhibition of STAT3
activation, and Mitochondrial dysfunction. Biomed Res Int.
2013:7198582013. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Zhao P, Pan Z, Luo Y, Zhang L, Li X, Zhang
G, Zhang Y, Cui R, Sun M and Zhang X: Alantolactone induces
apoptosis and cell cycle arrest on lung squamous cancer SK-MES-1
cells. J Biochem Mol Toxicol. 29:199–206. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Khan M, Yi F, Rasul A, Li T, Wang N, Gao
H, Gao R and Ma T: Alantolactone induces apoptosis in glioblastoma
cells via GSH depletion, ROS generation, and mitochondrial
dysfunction. IUBMB Life. 64:783–794. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Li Y, Liu B, Yang F, Yu Y, Zeng A, Ye T,
Yin W, Xie Y, Fu Z and Zhao C: Lobaplatin induces BGC-823 human
gastric carcinoma cell apoptosis via ROS-mitochondrial apoptotic
pathway and impairs cell migration and invasion. Biomed
Pharmacother. 83:1239–1246. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Lv J, Cao MQ and Yu JC: Alantolactone
pyrazoline analogue inhibits cancer cell proliferation and induces
apoptosis in non-small cell lung carcinoma cells. Bangl J
Pharmacol. 10:409–415. 2015. View Article : Google Scholar
|
|
20
|
Del Bino G, Darzynkiewicz Z, Degraef C,
Mosselmans R, Fokan D and Galand P: Comparison of methods based on
annexin-V binding, DNA content or TUNEL for evaluating cell death
in HL-60 and adherent MCF-7 cells. Cell Prolif. 32:25–37. 1999.
View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Bielawski K, Bielawska A and Wołczyήski S:
Synthesis, DNA-binding activity and cytotoxicity of carbamate
derivatives of Hoechst 33258 in breast cancer MCF-7 cells. Biol
Pharm Bull. 25:916–919. 2002. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Ning L, Ma H, Jiang Z, Chen L, Li L, Chen
Q and Qi H: Curcumol suppresses breast cancer cell metastasis by
inhibiting MMP-9 via JNK1/2 and Akt-dependent NF-κB signaling
pathways. Integr Cancer Ther. 15:216–225. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Lutterbeck CA, Kern DI, Machado ÊL and
Kümmerer K: Evaluation of the toxic effects of four anticancer
drugs in plant bioassays and its potency for screening in the
context of waste water reuse for irrigation. Chemosphere.
135:403–410. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Min HY, Park HJ, Chung HJ, Lee JW, Kim MS,
Park EJ and Lee SK: Inhibitory effects of alantolactone, a
naturally occurring Sesquiterpenoid, on the growth of human colon
cancer cells. Cancer Res. 64:13912004.
|
|
25
|
Kumari S, Badana AK, Mohan GM, Shailender
Naik G and Malla R: Synergistic effects of coralyne and paclitaxel
on cell migration and proliferation of breast cancer cells lines.
Biomed Pharmacother. 91:436–445. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Nikhil K, Sharan S, Chakraborty A,
Bodipati N, Krishna Peddinti R and Roy P: Role of isothiocyanate
conjugate of Pterostilbene on the inhibition of MCF-7 cell
proliferation and tumor growth in Ehrlich ascitic cell induced
tumor bearing mice. Exp Cell Res. 320:311–328. 2014. View Article : Google Scholar
|
|
27
|
Surichan S, Arroo RR, Tsatsakis AM and
Androutsopoulos VP: Tangeretin inhibits the proliferation of human
breast cancer cells via cyp1a1/cyp1B1 enzyme induction and
cyp1a1/cyp1B1-mediated metabolism to the product 4′ hydroxy
tangeretin. Toxicol In Vitro. 50:274–284. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Hamaratoglu F, Willecke M, Kango-Singh M,
Nolo R, Hyun E, Tao C, Jafar-Nejad H and Halder G: The
tumour-suppressor genes NF2/Merlin and Expanded act through Hippo
signalling to regulate cell proliferation and apoptosis. Nat Cell
Biol. 8:27–36. 2006. View Article : Google Scholar
|
|
29
|
Dolka I, Król M and Sapierzynski R:
Evaluation of apoptosis-associated protein (Bcl-2, Bax, cleaved
caspase-3 and p53) expression in canine mammary tumors: An
immunohistochemical and prognostic study. Res Vet Sci. 105:124–133.
2016. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Zhou N, Zhang Y, Zhang X, Lei Z, Hu R, Li
H, Mao Y, Wang X, Irwin DM, Niu G and Tan H: Exposure of
tumor-associated macrophages to apoptotic MCF-7 cells promotes
breast cancer growth and metastasis. Int J Mol Sci. 16:11966–11982.
2015. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Yan W, Zhu F, Zhao Z, Chai Y, Yue W, Shao
C, Lu F, Li Q and Wang C: Improved PCR-based subtractive
hybridization, a new trategy on cloning differential expression
genes in apoptotic MCF-7 cells. Cell Mol Immunol. 17:35–37. 2001.In
Chinese.
|
|
32
|
Kalid M, Jahanshiri F, Rahman A and Yusoff
K: Gene expression profiling in apoptotic MCF-7 cells infected with
newcastle disease virus. Global Vet. 5:334–340. 2010.
|
|
33
|
Rasul A, Di J, Millimouno FM, Malhi M,
Tsuji I, Ali M, Li J and Li X: Reactive oxygen species mediate
isoalantolactone-induced apoptosis in human prostate cancer cells.
Molecules. 18:9382–9396. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
34
|
Lei JC, Yu JQ, Yin Y, Liu YW and Zou GL:
Alantolactone induces activation of apoptosis in human hepatoma
cells. Food Chem Toxicol. 50:3313–3319. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
35
|
Huang Y, Ray S, Reed JC, Ibrado AM, Tang
C, Nawabi A and Bhalla K: Estrogen increases intracellular p26Bcl-2
to p21Bax ratios and inhibits taxol-induced apoptosis of human
breast cancer MCF-7 cells. Breast Cancer Res Treat. 42:73–81. 1997.
View Article : Google Scholar : PubMed/NCBI
|
|
36
|
Yoshimoto K, Iwahana H, Fukuda A, Sano T,
Saito S and Itakura M: Role of p53 mutations in endocrine
tumorigenesis: Mutation detection by polymerase chain
reaction-single strand conformation polymorphism. Cancer Res.
52:5061–5064. 1992.PubMed/NCBI
|
|
37
|
Tang D, Lahti JM and Kidd VJ: Caspase-8
activation and bid cleavage contribute to MCF7 cellular execution
in a caspase-3-dependent manner during staurosporine-mediated
apoptosis. J Biol Chem. 275:9303–9307. 2000. View Article : Google Scholar : PubMed/NCBI
|
|
38
|
Shah N, Mohammad AS, Saralkar P, Sprowls
SA, Vickers SD, John D, Tallman RM, Lucke-Wold BP, Jarrell KE,
Pinti M, et al: Investigational chemotherapy and novel
pharmacokinetic mechanisms for the treatment of breast cancer brain
metastases. Pharmacol Res. 132:47–68. 2018. View Article : Google Scholar : PubMed/NCBI
|
|
39
|
Aglund K, Rauvala M, Puistola U, Angström
T, Turpeenniemi-Hujanen T, Zackrisson B and Stendahl U: Gelatinases
A and B (MMP-2 and MMP-9) in endometrial cancer-MMP-9 correlates to
the grade and the stage. Gynecol Oncol. 94:699–704. 2004.
View Article : Google Scholar : PubMed/NCBI
|
|
40
|
Chun J and Kim YS: Platycodin D inhibits
migration, invasion, and growth of MDA-MB-231 human breast cancer
cells via suppression of EGFR-mediated Akt and MAPK pathways. Chem
Biol Interact. 205:212–221. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
41
|
Chun J, Choi RJ, Khan S, Lee DS, Kim YC,
Nam YJ, Lee DU and Kim YS: Alantolactone suppresses inducible
nitric oxide synthase and cyclooxygenase-2 expression by
down-regulating nf-kb, MAPK and AP-1 via the MyD88 signaling
pathway in LPS-activated RAW 264.7 cells. Int Immunopharmacol.
14:375–383. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
42
|
Jiang Y, Tang X, Zhou B, Sun T, Chen H,
Zhao X and Wang Y: The ROS-mediated pathway coupled with the
MAPK-p38 signalling pathway and antioxidant system plays roles in
the responses of Mytilus edulis haemocytes induced by BDE-47. Aquat
Toxicol. 187:55–63. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
43
|
Liu Y, Wen PH, Zhang XX, Dai Y and He Q:
Breviscapine ameliorates CCl4-induced liver injury in mice through
inhibiting inflammatory apoptotic response and ROS generation. Int
J Mol Med. 42:755–768. 2018.PubMed/NCBI
|
|
44
|
Yoon H and Liu RH: Effect of selected
phytochemicals and apple extracts on NF-κB activation in human
breast cancer MCF-7 cells. J Agric Food Chem. 55:3167–3173. 2007.
View Article : Google Scholar : PubMed/NCBI
|
|
45
|
Park EJ, Kim YM, Park SW, Kim HJ, Lee JH,
Lee DU and Chang KC: Induction of HO-1 through p38 MAPK/Nrf2
signaling pathway by ethanol extract of Inula Helenium L. reduces
inflammation in LPS-activated RAW 264.7 cells and CLP-induced
septic mice. Food Chem Toxicol. 55:386–395. 2013. View Article : Google Scholar : PubMed/NCBI
|