Introduction
Ischemic stroke accounts for the majority of stroke
cases, and is the leading cause of long-term disability and
mortality worldwide (1). Emerging
evidence has indicated that risk factors, including hypertension,
diabetes and hyperlipidemia, are associated with an increased
mortality rate in patients with ischemic stroke. Among the risk
factors, hyperlipidemia is a major risk factor for ischemic stroke
(2). The demonstrated efficacy of
statin therapy in primary and secondary prevention of stroke by
reducing serum low-density lipoprotein cholesterol (LDL-C) levels
is well established (3,4). However, previous studies have
indicated that high hyperlipidemia is associated with a lower risk
of post-stroke mortality (5–7).
Despite these conflicting results, hyperlipidemia-associated
ischemic stroke remains a major clinical challenge. The regulation
of LDL receptor (LDLR) by proprotein convertase subtilisin/kexin
type 9 (PCSK9) and the contribution of PCSK9 to cholesterol
homeostasis are well accepted. Although PCSK9 was first identified
as a plasma protein associated with hyperlipidemia just over a
decade ago, a rich body of knowledge has been developed since, and
therapeutic strategies inhibiting this target have been approved in
numerous countries (3,4,8).
PCSK9 exhibits high affinity towards LDLR family members, including
LDLR, very LDLR (VLDLR), apolipoprotein E receptor 2 (ApoER2),
LDLR-related protein 1 (LRP1) and β-site amyloid precursor
protein-cleaving enzyme 1 (BACE1) (9–11).
These multiple ligand-receptor interactions have been reported to
serve a well-established role in the systemic control of blood
cholesterol, as well as a less characterized role in neuronal
development and apoptosis (10).
Previous studies revealed that PCSK9 is implicated in oxidized
LDL-C-induced endothelial cell apoptosis, which is attenuated by
PCSK9 silencing (12–14). PCSK9 serves a vital role in
numerous processes resulting from hyperlipidemia, including free
radical generation, lipid oxidation and inflammatory cell
infiltration, which may be also involved in brain damage processes
(1,2). These pathological events ultimately
lead to programmed cell death, also known as apoptosis, which is
activated and maintained for several hours or days following stoke
onset, thus contributing to cerebral ischemia injury (1,15).
Additionally, a series of previous cell studies confirmed the
association between PCSK9 and neuronal apoptosis (16–18). Thus, the present study
hypothesized that PCSK9 may contribute to neuronal apoptosis in
hyperlipidemia-associated ischemic stroke.
ApoER2 is expressed predominantly in the brain, and
its role in neuronal survival depends on the pathological and
physiological conditions. ApoER2 is required for protection against
neuronal cell loss during normal ageing, while it selectively
promotes neuronal cell death upon injury in the adult brain
(19,20). Although both PCSK9 and ApoER2 are
implicated in brain injury, it remains controversial whether PCSK9
regulates the levels of ApoER2, and whether this has a functional
significance in the brain (10).
A number of previous in vitro studies have suggested that
ApoER2 is the mediator of PCSK9-induced neuronal apoptosis
(10), whereas other in
vivo studies have proposed that PCSK9 does not regulate the
levels of ApoER2 in the adult mouse brain (10,11). Therefore, it is particularly
important to determine the role of PCSK9 in
hyperlipidemia-associated ischemic stroke and its impact on ApoER2
levels.
Considering the prevalence of stroke in
hyperlipidemic patients, the present study aimed to clarify whether
PCSK9 contributes to the exacerbation of ischemic brain apoptosis
induced by middle cerebral artery occlusion (MCAO) injury in
hyperlipidemic mice. Therefore, the present study investigated the
influence of the inhibition of PCSK9 via injection of short hairpin
RNA (shRNA) targeting PCSK9 on ischemic brain injury and apoptosis
upon MCAO in hyperlipidemic mice. The study further explored the
underlying mechanisms of action by focusing on the levels of ApoER2
in the hippocampus and cortex. The results suggested that PCSK9
contributed to hyperlipidemia/MCAO-induced brain injury by
promoting neuronal apoptosis in the hippocampus and cortex, and the
protective effect of PCSK9 shRNA was involved in the suppression of
ApoER2.
Materials and methods
Ethics statement
The animal experiments were conducted in accordance
with the National Institutes of Health Guide for the Care and Use
of Laboratory Animals, and all the procedures were approved by the
Animal Ethics Committee of Tianjin Institute of Medical and
Pharmaceutical Sciences (Tianjin, China; approval no.
IMPS-EAEP-Z-W2015KR04). All surgical procedures were performed
under chloral hydrate anesthesia, and all efforts were made to
minimize animal suffering.
High-fat diet (HFD)
Male C57BL/6 mice (age, 9–10 weeks; weight, 24–26 g)
were supplied by Beijing Vital River Laboratory Animal Technology
Co., Ltd. (Beijing, China) and housed in a controlled environment
(25±1°C and 40–70% humidity, with an artificial 12:12 h light/dark
cycle). Mice were randomly assigned to the no-fat diet (NFD; n=8)
or HFD (n=40) groups. NFD mice were fed with a standard chow diet
(cat. no. 11002900022675), while HFD mice were fed with an HFD
consisting of 20% saccharose, 2% cholesterol, 15% lard and 0.3%
cholate (cat. no. 11002900021707; Beijing Keao Xieli Feed Co.,
Ltd., Beijing, China). Food and water were available ad
libitum for 6 weeks prior to surgery.
MCAO
Focal cerebral ischemia was induced by MCAO as
previously described (21).
Briefly, animals were deeply anesthetized with an intraperitoneal
injection 10% chloral hydrate (3.5 ml/kg body weight). Next, a
silicone-coated nylon monofilament was inserted through a small
incision in the right common carotid artery and was then advanced
to ~18 mm distal to the carotid bifurcation through the internal
carotid artery in order to occlude the origin of the middle
cerebral artery. In sham-operated animals, the same procedure was
performed with the exception of inserting the intraluminal
filament. To examine the critical role of PCSK9 in ischemic stroke,
32 HFD mice were randomly divided equally into four groups (n=8),
as follows: HFD-sham, HFD-MCAO, MCAO + LVRH1GP-null
(shRNA-control), and MCAO + LVRH1GP-shRNA-PCSK9 (shRNA-PCSK9).
Administration of the lentivirus expressing shRNA-PCSK9 or controls
was performed immediately following the surgical procedure.
Subsequent to continuous MCAO for 48 h, the mice were sacrificed
for further processing.
Lentivirus production and stereotaxic
injection
Several recombinant lentiviral vectors harboring an
shRNA sequence targeting PCSK9 (LVRH1GP-shRNA-PCSK9) were produced
by GeneCopoeia, Inc. (Guangzhou, China). The lentiviruses harboring
various shRNA sequences (Table I)
against PCSK9 were injected into C57BL/6 mice via the caudal vein
in order to evaluate the interference efficiency in the kidney by
reverse transcription-polymerase chain reaction (RT-PCR) (22). As shown in Table II, the results suggested that the
PCSK9 shRNA sequence 3 provided the most reliable expression, and
therefore this shRNA was selected for use in subsequent
experiments. LVRH1GP-shRNA-PCSK9 or scramble shRNA (LVRH1GP-null)
was delivered to the cortex of C57BL/6 mice via intracortical
injection. The injection site was 1.2-mm anterior to the bregma and
1.2-mm lateral to the midline, and the injection depth was 3.0 mm
(23). A total of 9 µl
purified lentivirus targeting PCSK9 or scramble shRNA was slowly
injected at a speed of 0.5 µl/min. The needle was carefully
withdrawn 10 min after the injection to avoid reflux of the
cerebrospinal fluid.
 | Table IShort hairpin RNA target
sequences. |
Table I
Short hairpin RNA target
sequences.
| Sequence name | Symbol | Location | Length | Target sequence |
|---|
| Sequence 1 | Pcsk9 |
891 | 21 |
ggaacctggagcgaattatcc |
| Sequence 2 | Pcsk9 |
964 | 21 |
ggaggtgtatctcttagatac |
| Sequence 3 | Pcsk9 | 1,228 | 21 |
ggagtttattcggaagagtca |
| Sequence 4 | Pcsk9 |
109 | 21 |
ccgaaacctgatcctttagta |
| Table IIScreening the most effective sequence
among four shRNA sequences against PCSK9 by quantitative reverse
transcription-polymerase chain reaction. |
Table II
Screening the most effective sequence
among four shRNA sequences against PCSK9 by quantitative reverse
transcription-polymerase chain reaction.
| Group | Relative PCSK9 mRNA
expression in kidney
|
|---|
| 24 h | 48 h |
|---|
| NFD-C57BL/6 | 1 | 1 |
| HFD-C57BL/6 | 2.226 | 2.226 |
| HFD+PCSK9 shRNA
control | 1.905 | 1.905 |
| HFD+PCSK9 shRNA
sequence 1 | 1.717 | 1.625 |
| HFD+PCSK9 shRNA
sequence 2 | 1.570 | 1.566 |
| HFD+PCSK9 shRNA
sequence 3 | 1.357 | 1.304 |
| HFD+PCSK9 shRNA
sequence 4 | 1.412 | 1.489 |
Serum parameters
Subsequent to feeding with HFD for 6 weeks, the
concentration of triglycerides (TG), total cholesterol (TC),
high-density lipoprotein cholesterol (HDL-C) and LDL-C in the serum
were measured using the corresponding assay kits according to the
manufacturer's protocol (cat. nos. A110-1, A111-1, A112-1 and
A113-1, respectively; Nanjing Jiancheng Bio-Engineering Institute,
Co., Ltd., Nanjing, China).
RT-quantitative PCR (RT-qPCR)
The RT-qPCR was performed as previously reported
(24). Briefly, total RNA was
extracted from liver, hippocampus and cortex using
TRIzol® reagent (Life Technologies; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) according to the manufacturer's
protocol. The A260/A280 ratio was measured by a spectrophotometer
to determine the purity and concentration of the RNA. Complementary
DNA was synthesized from total RNA using Moloney Murine Leukemia
Virus Reverse Transcriptase (Promega Corporation, Madison, WI,
USA). Subsequently, qPCR was performed using the SYBR Premix Ex
Taq™ II kit (Takara Bio, Inc., Otsu, Japan). The reaction
conditions were as follows: 95°C for 5 min, 95°C for 10 sec, 60°C
for 30 sec, 72°C for 32 sec, 40 cycles; the detection curve was
carried out as follows: 95°C for 15 sec, 60°C for 1 min, 95°C for
15 sec, 60°C for 15 sec. The specific primer pairs were as follows:
PCSK9 forward, 5′-GCATCCACAACACCCCT-3′ and reverse,
5′-CTGCCTCCGGACACTAA-3′ (125 bp); β-actin, forward
5′-ACTCTGTGTGGATTGGTGGC-3′, and reverse,
5′-AGAAAGGGTGTAAAACGCAGC-3′ (155 bp). The messenger RNA (mRNA)
expression levels of PCSK9 were normalized to those of the β-actin
gene. Data analysis was performed using the 2−∆∆Ct
method (25) for relative
quantification.
Brain histopathology and
immunohistochemistry (IHC)
The mouse brain tissues were rapidly isolated and
fixed in 4% paraformaldehyde. Next, the tissues were embedded in
paraffin, and consecutive cross-sections of the brain were cut and
stained with hematoxylin and eosin (H&E). Histological
evaluation was performed by an investigator who was blinded to the
experimental groups. For IHC staining, tissue sections were dewaxed
and rehydrated through a xylene and graded alcohol series, and the
slides were then incubated for 10 min at 95°C in a citrate solution
for antigen retrieval. Peroxidase (POD) clearing was performed
prior to blocking for 2 h with 1% bovine serum albumin in PBS at
room temperature. Subsequently, the tissue sections were incubated
with polyclonal rabbit anti-PCSK9 and rabbit anti-ApoER2 antibodies
(1:1,000; cat. nos. ab31762 and ab204112, respectively; Abcam,
Cambridge, MA USA) at 4°C overnight, followed by incubation with a
secondary antibody conjugated with horseradish POD (cat. no.
SA1028; Boster Biological Technology, Ltd, Wuhan, China) at 37°C
for 45 min. Specific labeling was visualized with a
3,3′-diaminobenzidine (DAB) kit and appeared light yellow or
tan-colored, while cell nuclei were counterstained with hematoxylin
(blue staining). Images were captured with a microscope, and in
each slide, eight random images of the mouse hippocampus and cortex
were obtained to determine the intensity of staining. IHC staining,
expressed as the relative positive expression, was quantified using
gray values by HMIAS-2000 W image system (version 1.0; Wuhan
Qianping Image Technology Co., Ltd., Wuhan, China). A high gray
value indicated low content of protein, while low gray scale
indicated high content.
Western blot analysis
Western blotting was used to measure the hippocampal
and cortical levels of PCSK9 and ApoER2. Briefly, total protein was
extracted using radioimmunoprecipitation assay buffer with a
protease inhibitor (cat. nos. P0013 and ST506, respectively;
Beyotime Institute of Biotechnology, Haimen, China). Protein levels
were quantified using the bicinchoninic acid assay (cat. no. P0010,
Beyotime Institute of Biotechnology, Haimen, China) according to
the manufacturer's protocol. Protein samples were separated by 10%
SDS-PAGE and then transferred to polyvinylidene fluoride membranes.
The membranes were incubated with the aforementioned rabbit
anti-PCSK9 and rabbit anti-ApoER2 primary antibodies (1:1,000) at
4°C overnight. Following washing in TBS with Tween-20, the
membranes were incubated with a POD-conjugated anti-rabbit
immunoglobulin G secondary antibody (1:5,000; cat. no. BA1003;
Wuhan Boster Biological Technology, Ltd., Wuhan, China). Protein
bands were visualized by enhanced chemiluminescence (GE Healthcare
Life Sciences, Little Chalfont, UK). The protein expression levels
were quantitatively analyzed with FluorChem version 2.0 software
(ProteinSimple, San Jose, CA, USA). The protein expression levels
of PCSK9 and ApoER2 were normalized to those of β-actin.
Terminal deoxynucleotidyl transferase
dUTP nick end labeling (TUNEL) assay
A TUNEL assay was applied to identify the apoptotic
cells with nuclear DNA fragmentation in the ischemic cerebral
hemisphere according to the manufacturer's protocol (In Situ
Cell Death Detection kit; cat. no. 11684817910; Roche Diagnostics,
Basel, Switzerland) (22). Upon
deparaffinization and rehydration, tissue sections were
permeabilized with proteinase K (20 µg/ml) at 37°C for 10
min and then incubated with TUNEL reaction mixture at 37°C for 2 h
in a humidified chamber in the dark. Converter POD was next added
and incubated at 37°C for 30 min. Subsequent to rinsing with PBS
(three times; 5 min each), the tissue sections were developed with
DAB tetrahydrochloride and counterstained with Mayer's hematoxylin.
In each section, eight random microscopic fields from the mouse
hippocampus and cortex were analyzed (4 mice in each study group).
The apoptosis ratio was presented as the percentage of
TUNEL-positive cells (brown) in the total number of cell nuclei
(blue). Bar plots for the mouse brain hippocampus and cortex
regions were separately exhibited.
Statistical analysis
Data are expressed as the mean ± standard deviation.
Statistically significant differences were evaluated by unpaired
Student's t-test for comparisons between two groups, or by one-way
analysis of variance followed by a Duncan's multiple range test for
multiple comparisons. P<0.05 was considered to indicate a
statistically significant difference. Statistical analyses were
performed using SPSS version 18.0 software (SPSS Inc., Shanghai,
China).
Results
PCSK9 is upregulated in the liver and
brain of HFD-induced hyperlipidemic mice
After 6 weeks of HFD feeding, the serum TG, TC and
LDL-C levels were significantly higher, whereas the serum HDL-C
level was significantly lower, in HFD mice as compared with the
levels in NFD mice (Fig. 1A). IHC
staining revealed upregulation of PCSK9 in the liver and
hippocampus of HFD mice after 6 weeks of HFD feeding (Fig. 1B and C). Furthermore, the mRNA
levels of PCSK9, as determined by RT-qPCR, were significantly
increased in the hippocampus and cortex of HFD mice (Fig. 1D). Taken together, these data
indicated that there was significant PCSK9 overexpression in the
liver and brain of hyperlipidemic mice.
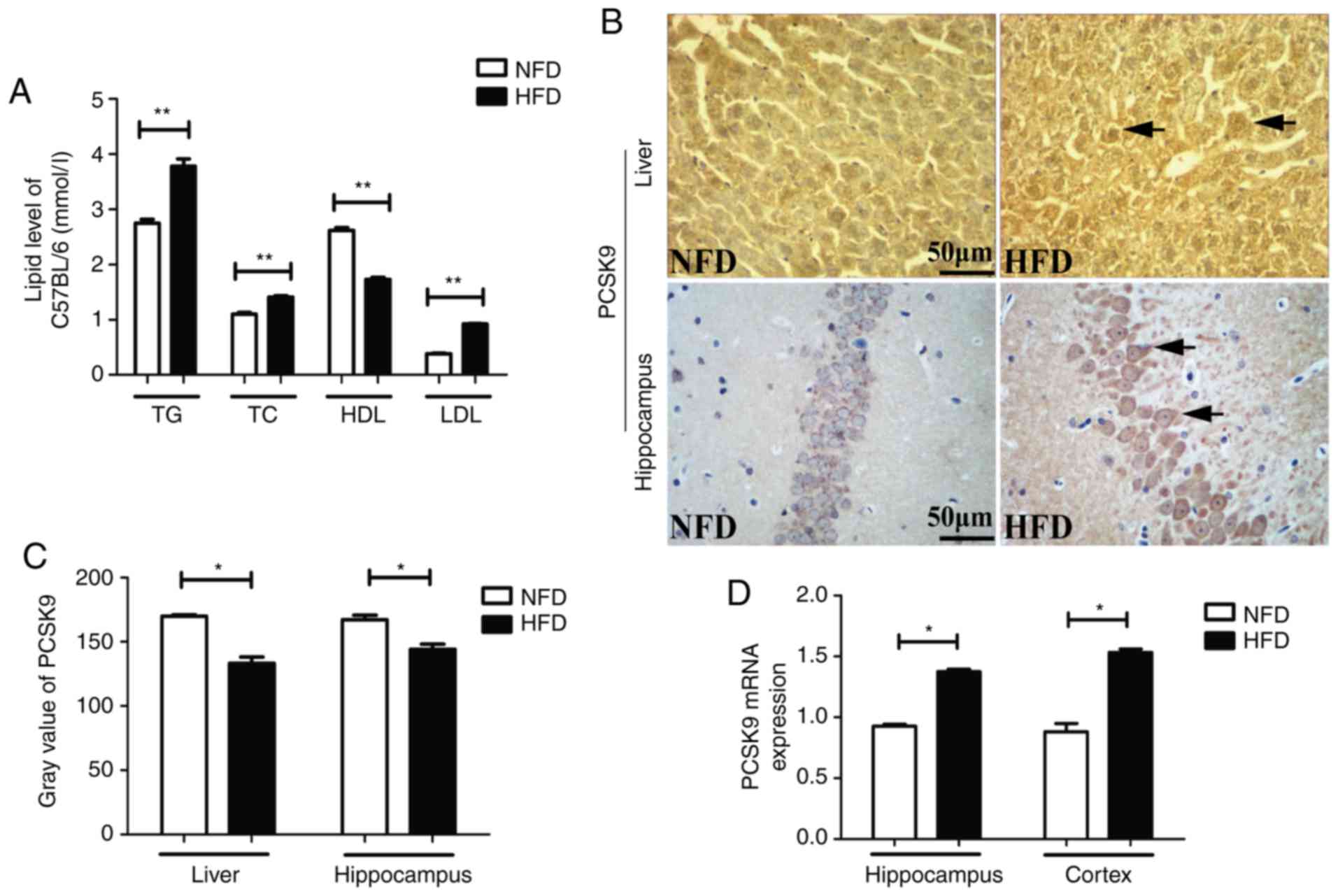 | Figure 1Blood lipids and PCSK9 expression
levels were significantly increased upon HFD feeding. (A) Serum TG,
TC and LDL-C levels were significantly increased, while HDL-C level
was significantly decreased, in HFD mice compared with those in NFD
mice (n=8). (B) Representative PCSK9 IHC staining in the liver and
hippocampus tissues following HFD feeding. (C) Gray value of PCSK9
in IHC analysis was significantly decreased in the liver and
hippocampus upon HFD feeding (n=4). (D) PCSK9 mRNA expression was
significantly increased in the hippocampus and cortex following HFD
feeding, according to reverse transcription-quantitative polymerase
chain reaction analysis (n=4). *P<0.05 and
**P<0.01 vs. NFD mice. PCSK9, proprotein convertase
subtilisin/kexin type 9; HFD, high-fat diet; NFD, no-fat diet; TG,
triglycerides; TC, total cholesterol; LDL-C, low-density
lipoprotein cholesterol; HDL-C, high-density lipoprotein
cholesterol; IHC, immunohistochemistry. |
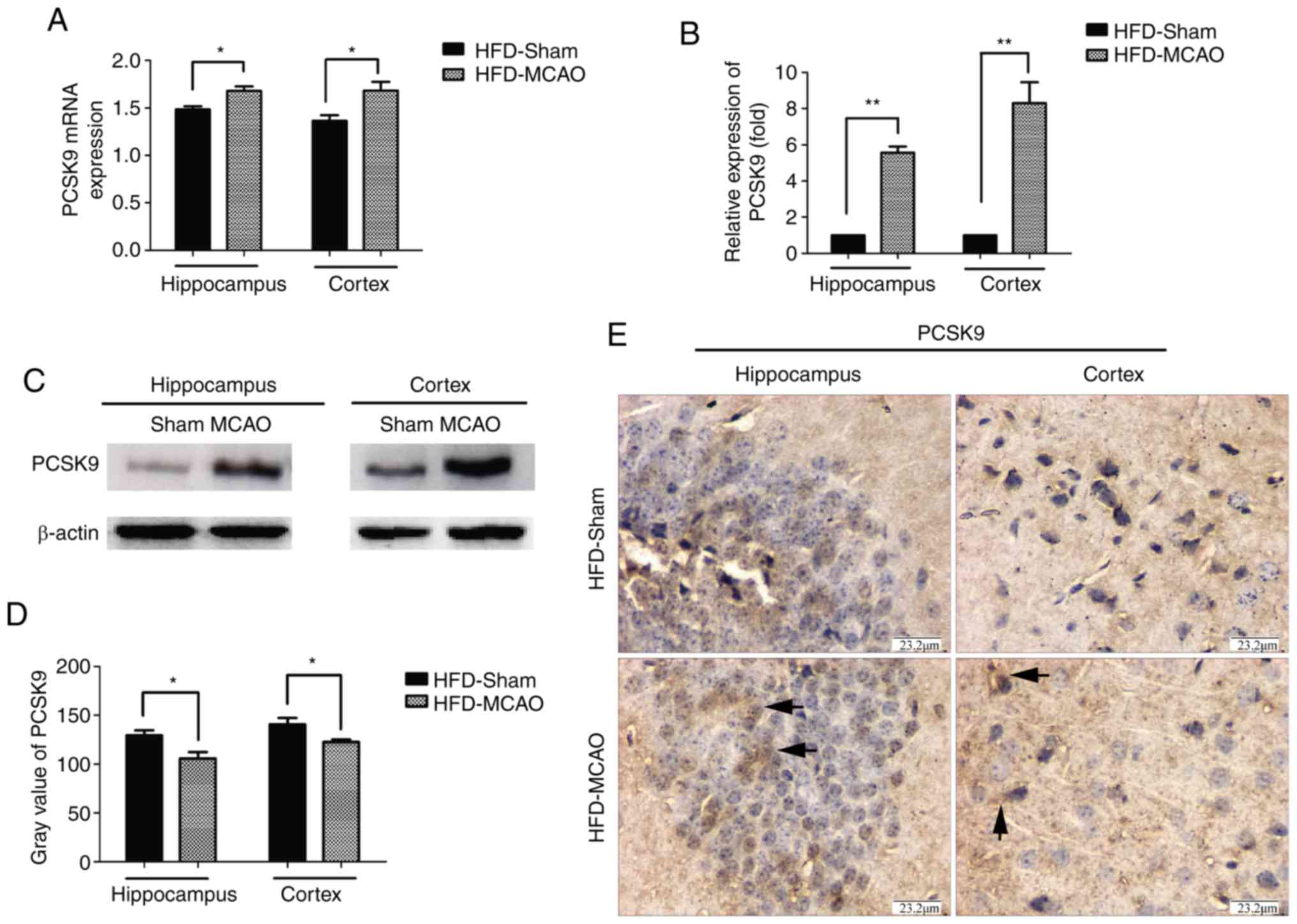
PCSK9 is upregulated following MCAO in
hyperlipidemic mice
Hyperlipidemic mice were subjected to MCAO surgery
in order to imitate ischemic stroke, and the expression of PCSK9 in
the cerebral hemisphere was detected after 48 h of MCAO by RT-qPCR,
western blot and IHC analyses. As shown in Fig. 2, there was significant
upregulation of PCSK9 mRNA and protein levels in the hippocampus
and cortex of HFD-MCAO mice compared with those of HFD-sham mice.
These results indicated that ischemia due to hyperlipidemia
promoted PCSK9 upregulation.
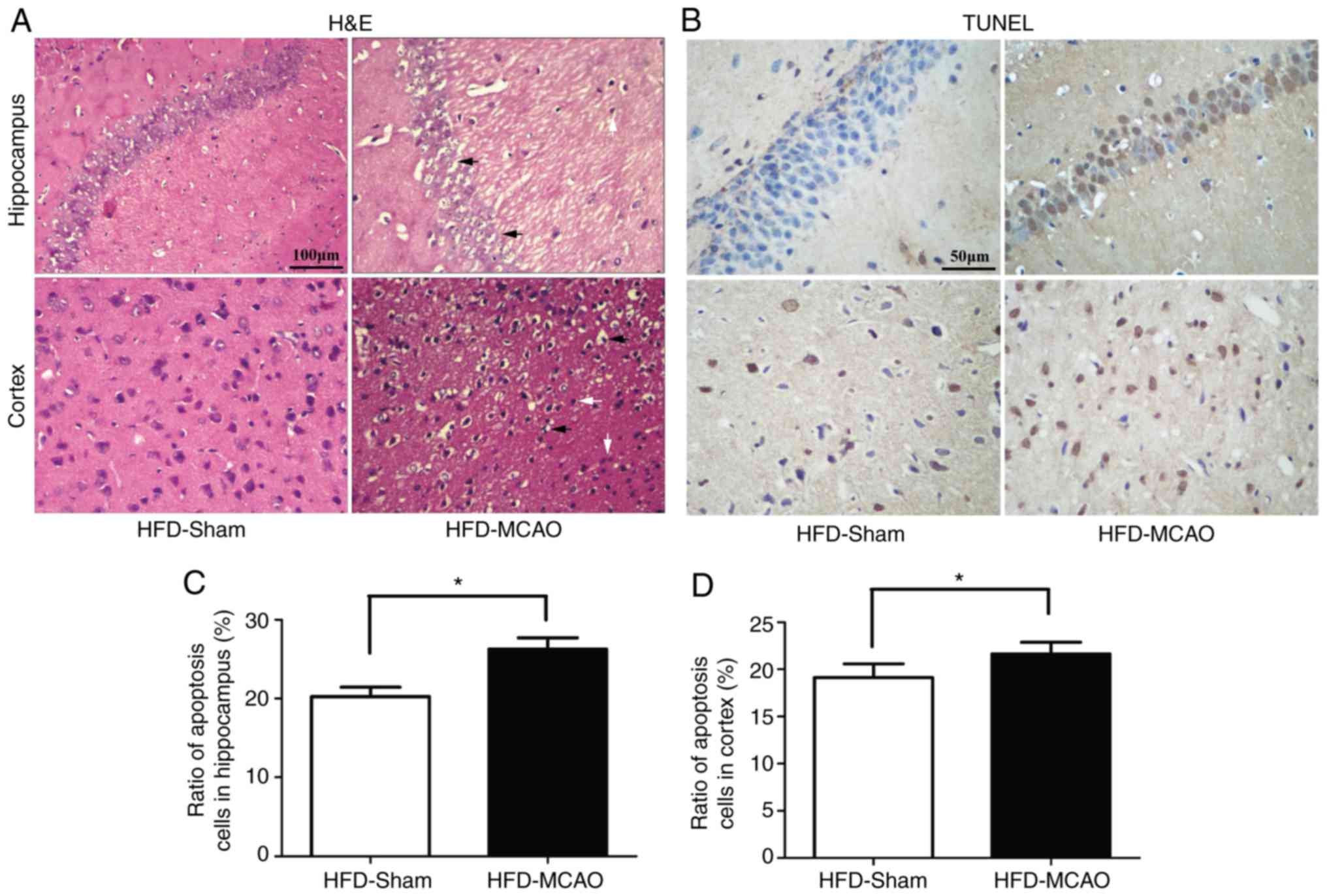
Increased histological injury and
apoptosis following MCAO in hyperlipidemic mice
H&E staining of the brain tissues of mice
subjected to 48 h of ischemia revealed nuclear shrinkage, neuronal
vacuolization, neuronal loss and neutrophil infiltration (Fig. 3A). The quantification of apoptotic
cells in the hippocampus and cortex revealed that MCAO resulted in
a significant increase in apoptosis compared with that observed in
the HFD-sham group (Fig.
3B–D).
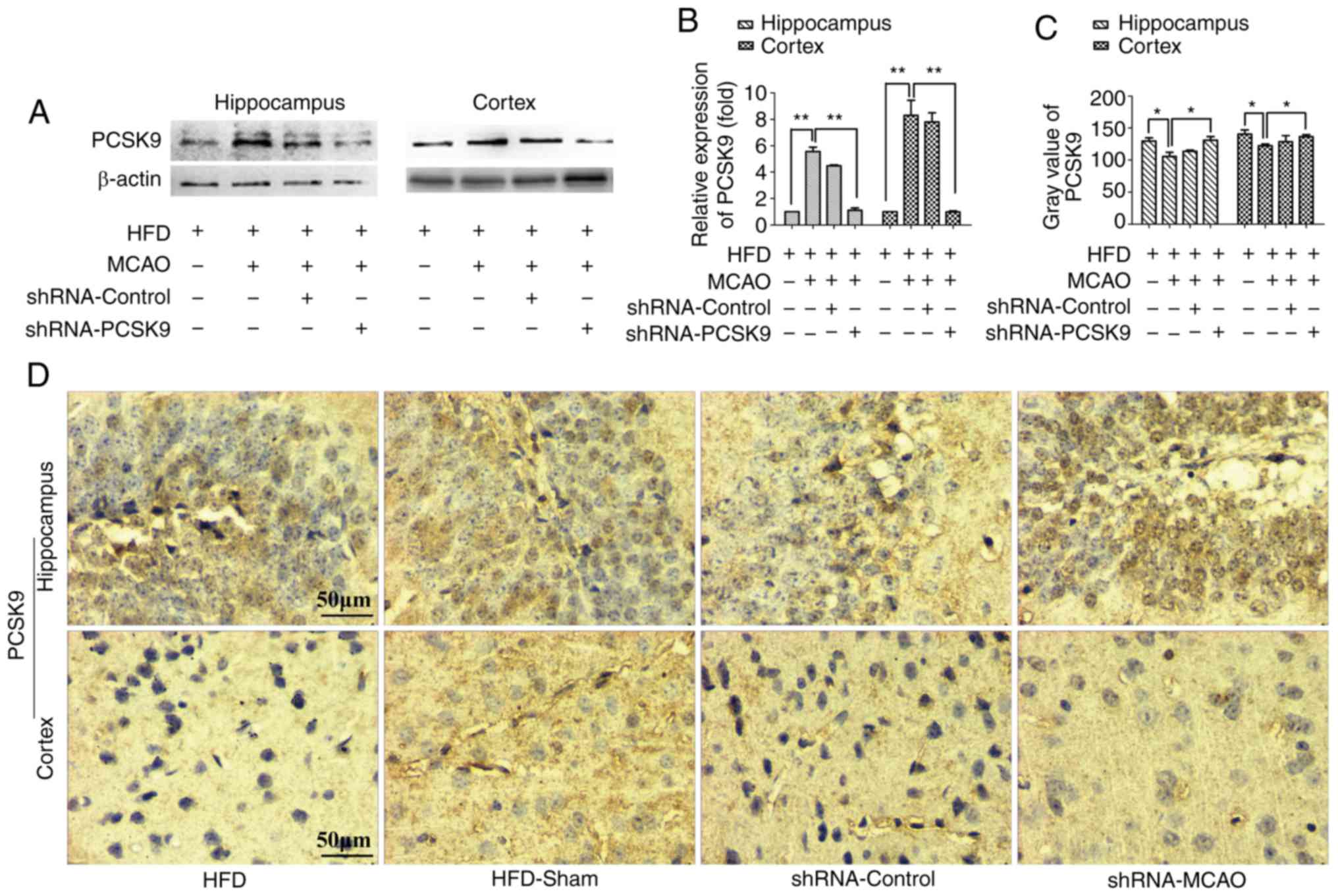
PCSK9 shRNA ameliorates histopathological
damage and apoptosis following MCAO in hyperlipidemic mice
As shown in Fig.
4, PCSK9 shRNA interference markedly decreased the PCSK9
protein levels in the hippocampus and cortex, as demonstrated by
western blotting and IHC staining. Western blotting revealed that
the protein level of PCSK9 was signifi-cantly lower in PCSK9 shRNA
therapy group than those in the HFD-MCAO group and the analysis on
the IHC staining indicated that the gray scale of PCSK9 in the
PCSK9 shRNA pretreated group was higher than in the HFD-MCAO group.
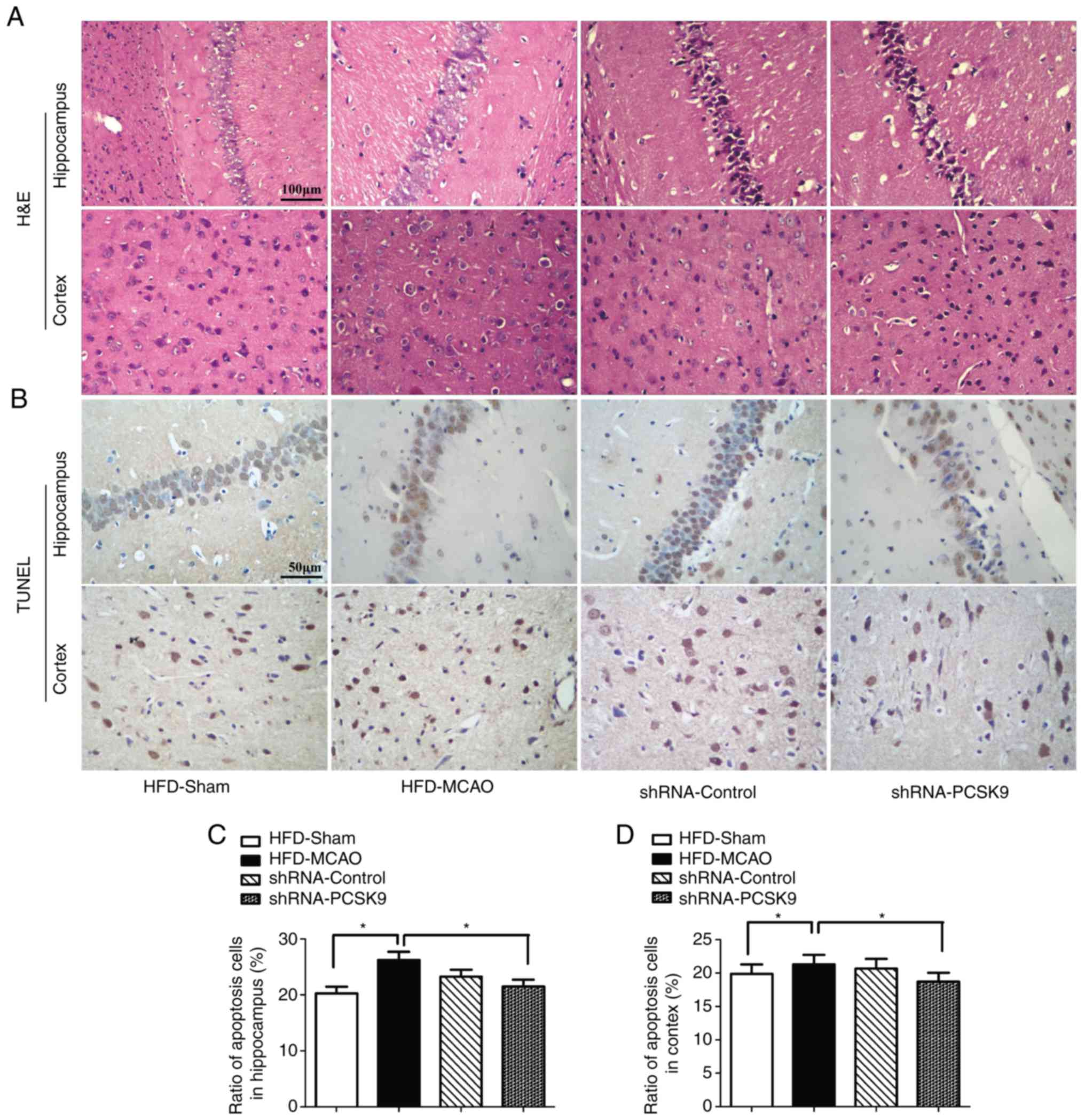
In order to further confirm the effects of PCSK9 shRNA in the
hippocampus and cortex of hyperlipidemic-MCAO mice,
histopathological damage and apoptosis were evaluated using H&E
and TUNEL staining, respectively. Consistently, PCSK9 shRNA
treatment partially attenuated the ischemic damage by decreasing
vacuolization and neutrophil infiltration compared with that
observed in the HFD-MCAO group (Fig.
5A). Since PCSK9 potentiated neuronal apoptosis, suppression of
PCSK9 significantly reduced neuronal death (10,16–18). As expected, in the present study,
neuronal apoptosis resulting from ischemia stroke was significantly
decreased in the hippocampus and cortex of PCSK9 shRNA-pretreated
mice compared with that in HFD-MCAO mice (Fig. 5B–D). Overall, PCSK9 inhibition
contributed to the reduced brain injury and anti-apoptotic effects
observed following MCAO in hyperlipidemic mice.
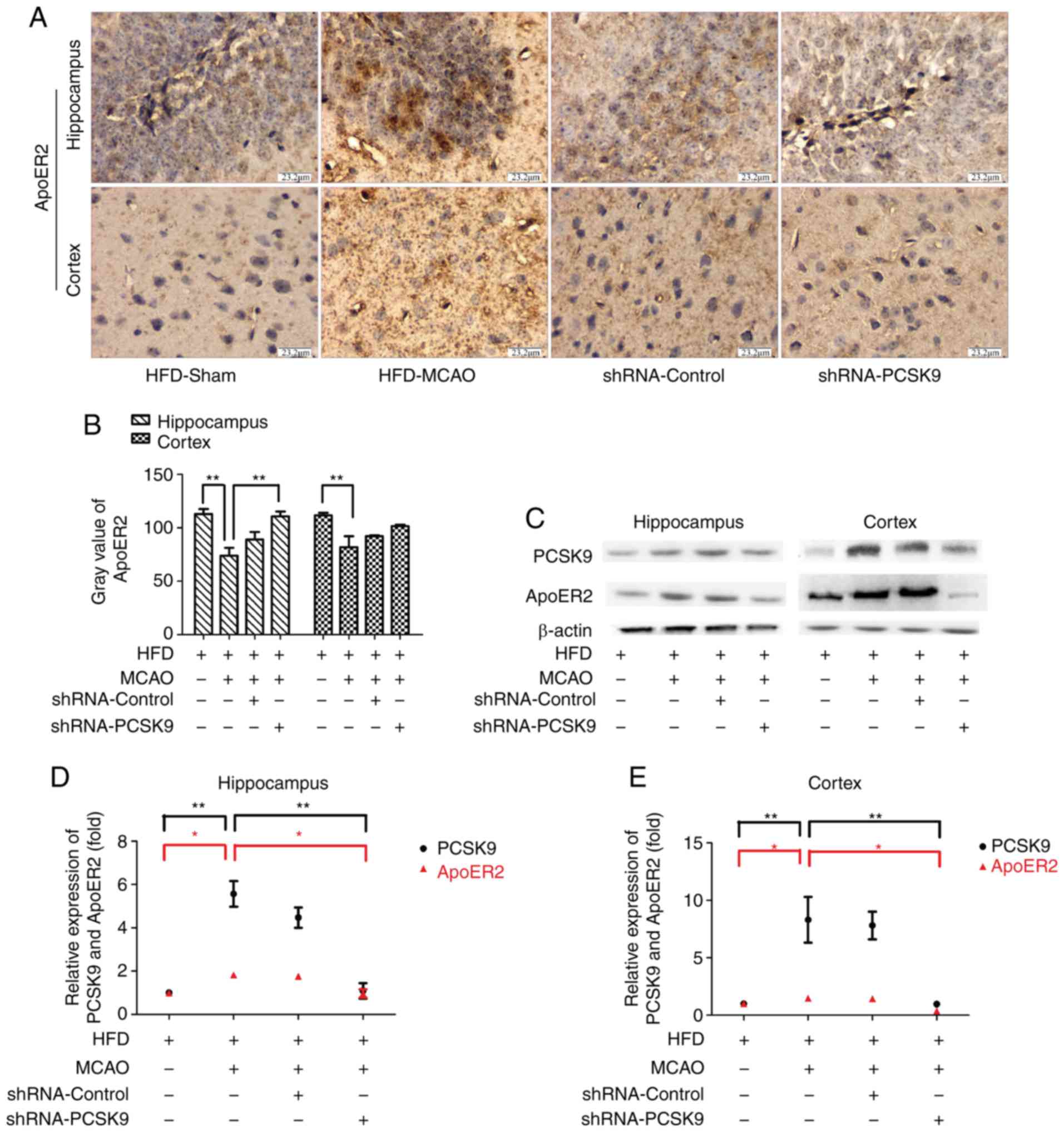
PCSK9 shRNA inhibits the apoptosis in
MCAO mice associated with ApoER2 downregulation
To investigate the role of the PCSK9/ApoER2
signaling pathway in MCAO-induced apoptosis, the expression levels
of PCSK9 and ApoER2 in hippocampus and cortex were further
evaluated. IHC staining demonstrated that pretreatment with PCSK9
shRNA simultaneously inhibited the expression of ApoER2 as
presented in Fig. 6. Consistent
with observations by IHC staining, western blot analysis for PCSK9
and ApoER2 in hippocampus and cortex revealed that the expression
of ApoER2 was significantly decreased in the PCSK9 shRNA treatment
group compared with HFD-MCAO group (Fig. 6). Collectively, these data
indicated that the anti-apoptotic effect triggered by PCSK9 shRNA
interference was associated with ApoER2 downregulation.
Discussion
It has been well established that PCSK9 serves a key
role in maintaining lipid homeostasis by affecting the systemic
levels of LDLR and cholesterol uptake (18). Excessive PCSK9 expression in
hepatocytes is linked to dyslipidemia, and inhibitors/silencers of
PCSK9 are currently being tested in clinical trials for the
treatment of hypercholesterolemia (18). Consistently, the present study
clearly confirmed that HFD-induced hyperlipidemia coincided with
increased PCSK9 expression in the liver. Notably, PCSK9 is also
highly expressed in the central nervous system (CNS) and affects a
wide range of neuronal functions (9–11).
In the hippocampus and cortex, a marked elevation of PCSK9 mRNA
levels was observed in the present study after 6 weeks of HFD
feeding. Accordingly, hyperlipidemia led to PCSK9 overexpression in
both the liver and brain.
Hyperlipidemia is known to be a risk factor for
stroke (2). PCSK9, a proprotein
convertase that binds to ApoER2, VLDLR, LDLR, BACE1 and LRP1, has
been linked to hyperlipidemia and cell apoptosis (9–11).
Previous studies have reported that PCSK9 is increased in the adult
brain following ischemia (18),
while other in vitro studies revealed a key role for PCSK9
in neuronal apoptosis (10,16–18). In the present study, after 6 weeks
of HFD feeding and subsequent establishment of an MCAO mouse model
of ischemic stroke, it was demonstrated that PCSK9-dependent
apoptosis served a major role in ischemic injury. Inhibition of
PCSK9 reduced neuronal apoptosis in the hippocampus and cortex,
which could also decrease brain damage. Thus, these findings
suggest that PCSK9 regulated the apoptosis resulting from ischemic
stroke in hyperlipidemic mice.
There are controversial reports regarding whether
PCSK9 regulates the levels of ApoER2 in vivo possibly via
neuronal apoptosis. A previous study reported that PCSK9 does not
appear to modulate the expression of ApoER2 in the adult mouse
brain (10,11). By contrast, another study revealed
that PCSK9 regulates neuronal apoptosis by adjusting ApoER2
signaling pathway in in vitro experiments using several
types of neuronal cells (10).
The present study demonstrated that suppression of PCSK9 attenuated
neuronal death, which coincided with a reduction in the levels of
ApoER2. This was in disagreement with previously published data
demonstrating that PCSK9 inhibition resulted in elevated levels of
ApoER2 (10). These conflicting
results may be due to differences in the duration of feeding with a
HFD, or time following MCAO when mice were examined, and
experimental conditions, which are implicated in complex, multiple
interactions among various pathological and physiological factors.
Indeed, the role of ApoER2 in neuronal survival depends on
pathophysiological changes. Notably, ApoER2 is required for
protection against neuronal cell loss during normal ageing, but
selectively promotes neuronal cell death upon injury (19,20). In addition, ApoE is a major risk
factor for several neurodegenerative diseases, and ApoER2 may also
be relevant to the pathogenesis of Alzheimer's disease (19). The present results strongly
support the hypothesis that the anti-apoptotic effect of PCSK9
inhibition is associated with downregulation of ApoER2.
However, the present study has several limitations.
Firstly, although there is evidence for a role of hyperlipidemia in
stroke, the results of several studies report that it is also
associated with improved patient outcome following stroke as
compared with that of individuals without hyperlipidemia (3–7).
However, the current study has failed to demonstrate the comparison
between HFD and NFD mice following MCAO with shRNA-PCSK9
interference. Furthermore, the findings of the present study
indicate that the association of PCSK9 and ApoER2 are correlative
and do not support a functional interaction between PCSK9 and
ApoER2. However, this potential interaction remains to be
determined by over expression or silencing of ApoER2 in a further
study. Finally, it is well known that PCSK9 binds to membrane-bound
proteins, including LDLR, VLDLR, ApoER2 and BACE1, which are all
highly expressed in the CNS (9–11).
While downregulation of ApoER2 was observed upon treatment with
shRNA against PCSK9 in the present study, downregulation of VLDLR
and BACE1 may also occur. Thus, further research focusing on VLDLR
and BACE1 would aid to determine whether partial loss of PCSK9
coincides with downregulation of ApoER2 specifically, or whether it
occurs concurrently with VLDLR and BACE1 downregulation.
In conclusion, the findings of the current study
implicate PCSK9 as a pro-apoptotic factor in ischemic brain injury
in hyperlipidemic mice. The present study also demonstrated that
the protective effect of shRNA against PCSK9 is involved in the
suppression of ApoER2 expression. These data warrant further
investigation of the therapeutic potential of PCSK9 inhibition
treatment for ischemic stroke in hyperlipidemic patients.
Acknowledgments
Not applicable.
Funding
The present study was supported by Tianjin Health
Development Planning Commission of Science and Technology Fund
Projects (grant no. 2015KR04) and the National Natural Science
Foundation of China (grant no. 81202801).
Availability of data and materials
All date generated and/or analyzed during the
current study are included in this published article.
Authors' contributions
LW and JS conceived and designed the study. ZW, QJ
and HW performed the experiments. XL and DH analyzed the data and
LW drafted the manuscript. All authors read and approved the final
version of the manuscript.
Ethics approval and consent to
participate
All experiments were approved by the Animal Care and
Use Committee of Tianjin Institute of Medical and Pharmaceutical
Sciences (Tianjin, China).
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
References
|
1
|
Miao Y and Liao JK: Potential serum
biomarkers in the pathophysiological processes of stroke. Expert
Rev Neurother. 14:173–185. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Kim E, Tolhurst AT, Qin LY, Chen XY,
Febbraio M and Cho S: CD36/fatty acid translocase, an inflammatory
mediator, is involved in hyperlipidemia-induced exacerbation in
ischemic brain injury. J Neurosci. 28:4661–4670. 2008. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Nozue T: Lipid lowering therapy and
circulating PCSK9 concentration. J Atheroscler Thromb. 24:895–907.
2017. View Article : Google Scholar : PubMed/NCBI
|
|
4
|
Sun H, Krauss RM, Chang JT and Teng BB:
PCSK9 deficiency reduces atherosclerosis, apolipoprotein B
secretion and endothelial dysfunction. J Lipid Res. 59:207–223.
2018. View Article : Google Scholar
|
|
5
|
Yeramaneni S, Kleindorfer DO, Sucharew H,
Alwell K, Moomaw CJ, Flaherty ML, Woo D, Adeoye O, Ferioli S, de
Los Rios La, Rosa F, et al: Hyperlipidemia is associated with lower
risk of poststroke mortality independent of statin use: A
population-based study. Int J Stroke. 12:152–160. 2017. View Article : Google Scholar :
|
|
6
|
Zhao W, An Z, Hong Y, Zhou G, Guo J, Zhang
Y, Yang Y, Ning X and Wang J: Low total cholesterol level is the
independent predictor of poor outcomes in patients with acute
ischemic stroke: A hospital-based prospective study. BMC Neurol.
16:362016. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Markaki I, Nilsson U, Kostulas K and
Sjöstrand C: High cholesterol levels are associated with improved
long-term survival after acute ischemic stroke. J Stroke
Cerebrovasc Dis. 23:e47–e532014. View Article : Google Scholar
|
|
8
|
Shapiro MD and Fazio S: PCSK9 and
atherosclerosis-lipids and beyond. J Atheroscler Thromb.
24:462–472. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Schulz R and Schlüter KD: PCSK9 targets
important for lipid metabolism. Clin Res Cardiol Suppl. 12(Suppl
1): S2–S11. 2017. View Article : Google Scholar
|
|
10
|
Kysenius K, Muggalla P, Mätlik K, Arumäe U
and Huttunen HJ: PCSK9 regulates neuronal apoptosis by adjusting
ApoER2 levels and signaling. Cell Mol Life Sci. 69:1903–1916. 2012.
View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Liu M, Wu G, Baysarowich J, Kavana M,
Addona GH, Bierilo KK, Mudgett JS, Pavlovic G, Sitlani A, Renger
JJ, et al: PCSK9 is not involved in the degradation of LDL
receptors and BACE1 in the adult mouse brain. J Lipid Res.
51:2611–2618. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Schulz R, Schlüter KD and Laufs U:
Molecular and cellular function of the proprotein convertase
subtilisin/kexin type 9 (PCSK9). Basic Res Cardiol. 110:42015.
View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Li J, Liang X, Wang Y, Xu Z and Li G:
Investigation of highly expressed PCSK9 in atherosclerotic plaques
and ox-LDL-induced endothelial cell apoptosis. Mol Med Rep.
16:1817–1825. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Wu CY, Tang ZH, Jiang L, Li XF, Jiang ZS
and Liu LS: PCSK9 siRNA inhibits HUVEC apoptosis induced by ox-LDL
via Bcl/Bax-caspase9-caspase3 pathway. Mol Cell Biochem.
359:347–358. 2012. View Article : Google Scholar
|
|
15
|
Tang LL, Ye JY, Jiang SN and Zheng JS:
3,4-oxo-isopropylidene-shikimic acid inhibits cerebral
ischemia-induced oxidative stress and neuronal apoptosis in rats.
Am J Transl Res. 9:1764–1773. 2017.PubMed/NCBI
|
|
16
|
Chiang LW, Grenier JM, Ettwiller L,
Jenkins LP, Ficenec D, Martin J, Jin F, DiStefano PS and Wood A: An
orchestrated gene expression component of neuronal programmed cell
death revealed by cDNA array analysis. Proc Natl Acad Sci USA.
98:2814–2819. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Bingham B, Shen R, Kotnis S, Lo CF,
Ozenberger BA, Ghosh N, Kennedy JD, Jacobsen JS, Grenier JM,
DiStefano PS, et al: Proapoptotic effects of NARC 1 (=PCSK9), the
gene encoding a novel serine proteinase. Cytometry A. 69:1123–1131.
2006. View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Rousselet E, Marcinkiewicz J, Kriz J, Zhou
A, Hatten ME, Prat A and Seidah NG: PCSK9 reduces the protein
levels of the LDL receptor in mouse brain during development and
after ischemic stroke. J Lipid Res. 52:1383–1391. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Beffert U, Nematollah Farsian F, Masiulis
I, Hammer RE, Yoon SO, Giehl KM and Herz J: ApoE receptor 2
controls neuronal survival in the adult brain. Curr Biol.
16:2446–2452. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Waltmann MD, Basford JE, Konaniah ES,
Weintraub NL and Hui DY: Apolipoprotein E receptor-2 deficiency
enhances macrophage susceptibility to lipid accumulation and cell
death to augment atherosclerotic plaque progression and necrosis.
Biochim Biophys Acta. 1842:1395–1405. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Morris GP, Wright AL, Tan RP, Gladbach A,
Ittner LM and Vissel B: A comparative study of variables
influencing ischemic injury in the longa and koizumi methods of
intraluminal filament middle cerebral artery occlusion in mice.
PLoS One. 11:e01485032016. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Wang L, Wang Z, Yuan L, Hao D, Lü N and Li
X: Shenfuqiangxin Capsule inhibits apoptosis through
mitogen-activated protein kinase signal pathway in rats with
cardio-renal syndrome induced by infrarenal aortic-clamping. J
Tradit Chin Med. 37:80–87. 2017. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Shi W, Wei X, Wang Z, Han H, Fu Y, Liu J,
Zhang Y, Guo J, Dong C, Zhou D, et al: HDAC9 exacerbates
endothelial injury in cerebral ischaemia/reperfusion injury. J Cell
Mol Med. 20:1139–1149. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Liu L, Wang Y and Yu Q: The PI3K/Akt
signaling pathway exerts effects on the implantation of mouse
embryos by regulating the expression of RhoA. Int J Mol Med.
33:1089–1096. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|