Introduction
Lung cancer is the second most commonly diagnosed
malignant tumor type worldwide, and it causes a higher number of
mortalities than any other cancer type, despite considerable
progress in its early diagnosis and treatment over the past
decades. The overall 5-year survival rate for lung cancer has not
significantly changed over the past decade (1). In China, the morbidity and mortality
associated with lung cancer have increased rapidly in recent years.
Among the different lung cancer types, lung adenocarcinoma (LUAD)
is the most common one; it is the most extensively elucidated
subtype within the family of lung carcinomas and is particularly
prevalent among Asian women (>70% in Japanese females) (2). Most LUADs are associated with a
mutation in an oncogenic driver (3). The mortality rate of LUAD is high,
as its early diagnosis is difficult and the disease progresses
rapidly. Even in patients diagnosed at an early stage of the
disease, when an initially curative surgery is performed, the
5-year survival rate remains <60% (4,5).
Tobacco smoking is the major causative factor for lung cancer;
however, although the etiological role of tobacco is crucial, up to
25% of lung cancers occur in non-smokers (6). In these cases, other risk factors
associated to carcinogens appear to have an important role.
Therefore, identification of novel targets in LUAD may offer novel
viewpoints or ideas for a comprehensive management strategy for
LUAD patients.
Homeobox (HOX)A11, also known as HOX1, HOX1I or
radioulnar synostosis with amegakaryocytic thrombocytopenia 1, has
been proven to have an essential role in numerous diseases,
including renal cell carcinoma (RCC), T-cell acute lymphoblastic
leukemia, glioblastoma and female reproductive system diseases
(7-10). Growing evidence suggests that
HOXA11 acts as a tumor suppressor in a wide range of tumor types.
For instance, Wang et al (7) demonstrated that HOXA11 suppresses
the proliferation, migration and invasion ability of RCC cells,
while inducing cell apoptosis. Xia et al (11) reported that HOXA11
hypermethylation is relevant to the unfavorable prognosis of breast
cancer patients, and the overexpression of HOXA11 suppresses cell
growth. Se et al (8)
indicated that a reduction of HOXA11 expression induces treatment
resistance and leads to a poor prognosis in glioblastoma.
Furthermore, Hwang et al (1) revealed that HOXA11 hypermethylation
promotes non-small cell lung cancer development by enhancing cell
proliferation or migration. However, the current knowledge on the
association between HOXA11 and LUAD remains insufficient. To date,
only one study has reported that HOXA11 may be an early diagnostic
and independent prognostic marker for LUAD (12). The molecular mechanisms of HOXA11
in LUAD remain largely elusive and require further
investigation.
With the rapid development of biological and
computer technology, mass data have been emerging. More and more
public databases have been established and applied for studying the
genomic alterations in malignancies. At present, The Cancer Genome
Atlas (TCGA) database (https://cancerge-nome.nih.gov/) is the biggest
database of cancer-associated genomic alterations, and the analysis
of its data may improve the current understanding of the genetic
basis of various cancer types to then make it possible to diagnose
and treat cancer at an early stage or even prevent it. Oncomine
(https://www.oncomine.org) is the biggest database
of information from oncogene chips in the world, representing an
integrated data mining platform. These two databases collect
information on genomic alterations via different methodologies. To
gain in-depth knowledge regarding the role of HOXA11 in LUAD, the
present study was performed to gather data from TCGA and Oncomine
microarray chips, and produce in-house reverse
transcription-quantitative polymerase chain reaction (RT-qPCR)
data, with the aim of comprehensively investigating the clinical
value of HOXA11 in LUAD in a large sample size and data produced
with different research methods to evaluate whether HOXA11 has any
diagnostic or prognostic value in LUAD. Genes co-expressed with
HOXA11 were also identified by searching the cBioPortal (http://www.cbio-portal.org/) and Multi Experiment
Matrix (MEM; http://biit.cs.ut.ee/mem/index.cgi) databases to
determine the potential molecular mechanisms of HOXA11 in LUAD on a
preliminary basis. The results revealed that HOXA11 may exert its
function through the focal adhesion pathway.
Materials and methods
HOXA11 expression profile mining in
TCGA
Relevant data from TCGA database (http://cancergenome.nih.gov) were downloaded using the
UCSC Cancer Genomics Browser (https://genome-cancer.soe.ucsc.edu/). These data
included the HOXA11 expression levels from 302 LUAD tissues and 22
tumor-adjacent normal control tissues, which were analysed to
determine the diagnostic value of HOXA11 and its capacity to
predict overall survival in LUAD. The HOXA11 values were carefully
checked for each sample and values <1 counts were treated as
missing values. Subsequently, the data were normalized via
logarithmic transformation (log2) for further analysis. The
corresponding clinical parameters of the LUAD patients were also
extracted to investigate their association with HOXA11.
HOXA11 expression profile mining in the
Oncomine database
To increase the reliability of the results, the
online microarray database Oncomine was also used to analyze the
transcript levels of HOXA11 in LUAD (13). The search terms used were as
follows: Analysis type (cancer vs. normal), cancer types (non-small
cell lung carcinoma; lung adenocarcinoma), sample type (clinical
specimen) and data type (HOXA11); the other terms were the system
defaults.
In-house HOXA11 RT-qPCR expression
profiles
A total of 32 pairs of formalin-fixed
paraffin-embedded tissues (tumor and adjacent non-cancerous) from
LUAD patients who underwent curative surgical resection between
January 2012 and February 2014 were collected at the Department of
Pathology of the First Affiliated Hospital of Guangxi Medical
University (Nanning, China). Among these patients, 23 were male and
9 were female, the median age was 56 years (range 33-90 years).
TRIzol reagent (Invitrogen; Thermo Fisher Scientific, Inc.,
Waltham, MA, USA) was used to isolate total RNA following the
manufacturer's protocol; next, the concentration and purity of the
RNA was tested by measuring the absorbance at 260 and 280 nm, and
the qualified RNA was reverse-transcribed in a large volume (20 µl)
with the Prime Script RT Reagent Kit (Takara Bio Inc., Dalian,
China) by using random primers under standard conditions. HOXA11
levels were detected using the SYBR Premix Ex Taq (Takara Bio Inc.)
according to the manufacturer's protocol. GAPDH expression was used
as the internal reference to normalize the results. Real-time
RT-qPCR was performed using the 7900HT PCR system (Applied
Biosystems; Thermo Fisher Scientific, Inc., Waltham, MA, USA) and
the expression of HOXA11 and GAPDH were calculated using the
2−ΔΔCq method (14,15). The temperature protocol of the RT
step was as follows: 16°C for 30 min, 42°C for 30 min and 85°C for
5 min. qPCR was performed using the following thermocycling
conditions: Initial pre-denaturation for 5 min at 95°C, followed by
40 cycles of 95°C for 10 sec, 60°C for 10 sec and 72°C for 10 sec.
All experiments were repeated in triplicate. The HOXA11 forward
primer was 5′-CGC TTC AGA ACT CGT TGC TTT GC-3′ and the reverse
primer was 5′-CGG AAG AAC TGG CAG TCT TTA CCT-3′. The GAPDH forward
primer was 5′-TGA ACG GGA AGC TCA CTG G-3′, and the reverse primer
was 5′-TCC ACC ACC CTG TTG CTG TA-3′.
Meta-analysis
To strengthen the reliability of the results, all
the datasets included were used together to perform a
meta-analysis. The mean value, standard deviation and sample size
of the tumor and normal control groups were calculated using SPSS
Statistics software version 24.0 (IBM Corp., Armonk, NY, USA). The
true positives (TPs), false positives (FPs), false negatives (FNs),
and true negatives (TNs) were also calculated according to the
following equation:
Where SE represents sensitivity; SP represents
specificity; TP+FN is the sample size of the tumour; and FP+TN is
the sample size of the normal control. The meta-analysis was
performed using STATA 12.0 (StataCorp LP, College Station, TX, USA)
for pooling the standardized mean difference (SMD), sensitivity,
specificity and summary ROC of HOXA11 expression (16,17). An I2 test was used to
evaluate heterogeneity, when I2 <50%, a fixed-effects
model was used; while I2 ≥50% the random-effects model
was selected (18).
Identification of co-expressed genes and
pathways associated with HOXA11
The genes co-expressed with HOXA11 and their
associated pathways and functions were then investigated. Using two
online tools, the MEM database and cBioPortal, the genes
co-expressed with HOXA11 were retrieved and extracted. The
candidate co-expressed genes obtained with the two online tools
were then overlapped and displayed in Venn diagrams (http://bioinformatics.psb.ugent.be/webtools/Venn/).
Bioinformatics analyses, including determination of gene ontology
(GO) terms and Kyoto Encyclopedia of Genes and Genomes (KEGG)
pathways, and the generation of the Protein-Protein Interaction
(PPI) network, were performed to investigate the underlying
molecular mechanisms. The GO and KEGG analyses were performed with
the Database for Annotation, Visualization and Integrated Discovery
(DAVID) bioinformatics tool (version 6.8; https://david.ncifcrf.gov/), whereas PPI analysis was
performed using the search tool for the retrieval of interacting
genes/proteins (STRING) version 10.5 (http://www.string-db.org/). The GO analysis comprised
the following three categories: Biological process (BP), cellular
component (CC) and molecular function (MF). In addition, Cytoscape
version 3.4.0 (http://cytoscape.org) provided a
visualization of the functional network between HOXA11 and these
co-expressed genes. Furthermore, mRNA and protein levels of the
molecules in the top three KEGG pathways were confirmed with TCGA
data (http://gepia.cancer-pku.cn/) and
immunohistochemical staining results (https://www.protein-atlas.org/). In addition, an
overall survival analysis of these genes in LUAD was performed
using TCGA data from the GEPIA website (gepia.cancer-pku.cn/),
which is a TCGA project based interactive web server used to
analyse RNA sequencing expression and prognosis.
Statistical analysis
The expression levels of HOXA11 are presented as the
mean ± standard deviation. The unpaired, two-tailed Student's
t-test was used for comparison of data between two groups from TCGA
and Oncomine. The paired t-test was applied for the in-house
RT-qPCR data. Analysis of variance was utilized for identifying
significant differences among three groups. The diagnostic value of
HOXA11 in LUAD was identified by drawing the receiver operating
characteristic (ROC) curve. The correlation between HOXA11
expression and co-expressed genes was analyzed by Pearson
correlation analysis. To assess the ability of HOXA11 to predict
patient survival, Kaplan-Meier survival curves were drawn and
significant differences were assessed using the log-rank test. All
of the abovementioned statistical analyses were performed with SPSS
Statistics software version 24.0 (IBM Corp., Armonk, NY, USA).
GraphPad Prism 7 software (GraphPad Software, Inc., La Jolla, CA,
USA) was used for graphic presentation of data. In addition,
meta-analysis was performed using STATA 12.0 for pooling the SMD,
sensitivity, specificity and summary ROC of HOXA11 expression from
the three data sources. P<0.05 was considered to indicate a
statistically significant difference.
Results
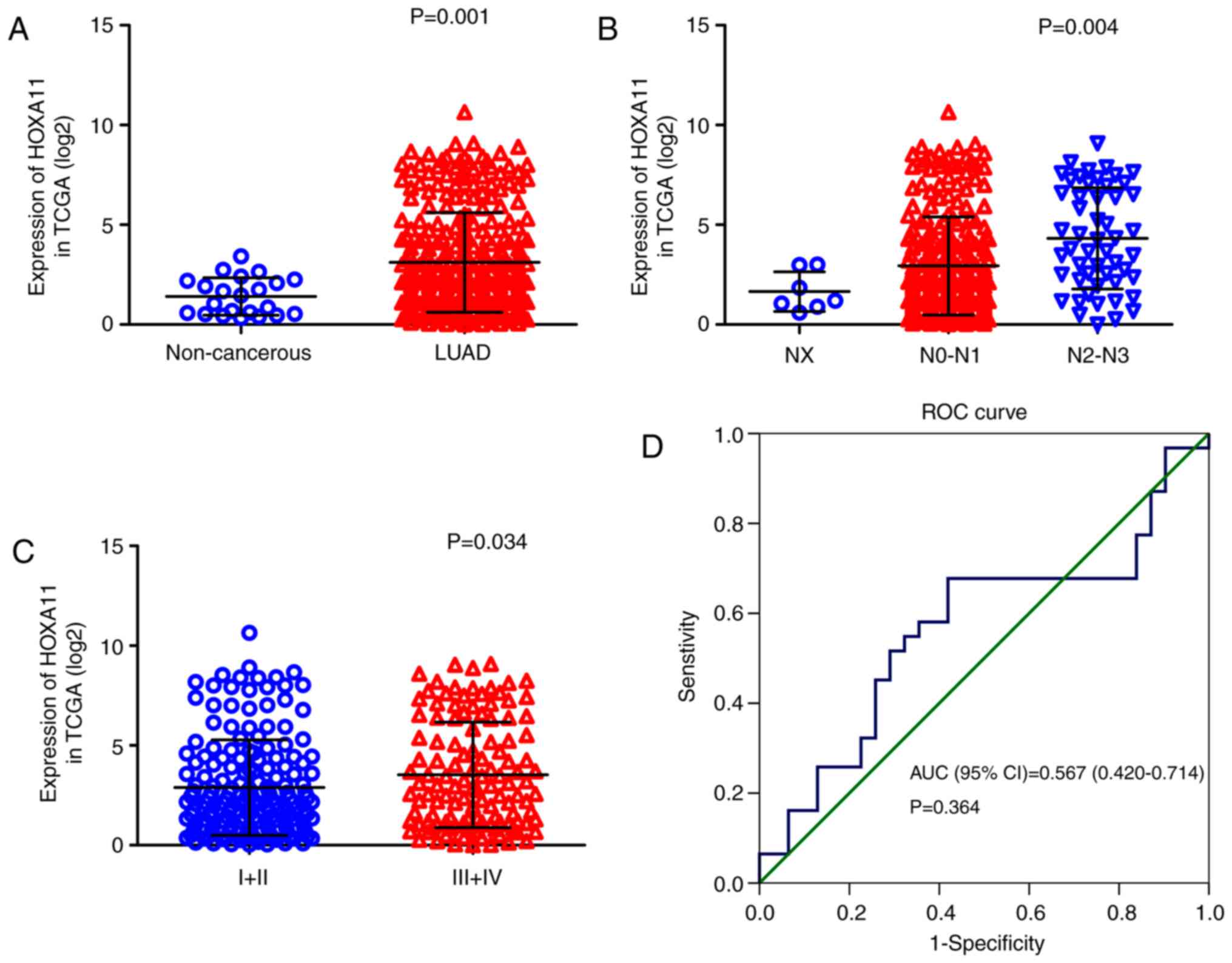
Overexpression of HOXA11 in LUAD
The clinical significance of HOXA11 in LUAD was
validated using data from three different sources. First, the
calculation of HOXA11 expression values with the data from TCGA
(Table I) revealed that HOXA11
was significantly overexpressed in LUAD (3.15±2.506) compared with
that in adjacent, non-tumorous tissue samples (1.41±0.937, P=0.001;
Fig. 1A) and that the level of
HOXA11 in stage III/IV LUAD tissues was significantly higher than
that in stage I/II LUAD tissues (P=0.034; Fig. 1B). The number of lymph node
metastases was positively correlated with the expression levels of
HOXA11 (P<0.001; Fig. 1C). In
addition, the area under the curve (AUC) for upregulated HOXA11 in
LUAD diagnosis was 0.706 [95% confidence interval (CI):
0.620-0.793, P=0.0012; Fig. 1D],
with a cutoff value of 2.747 (sensitivity 44.2% and specificity
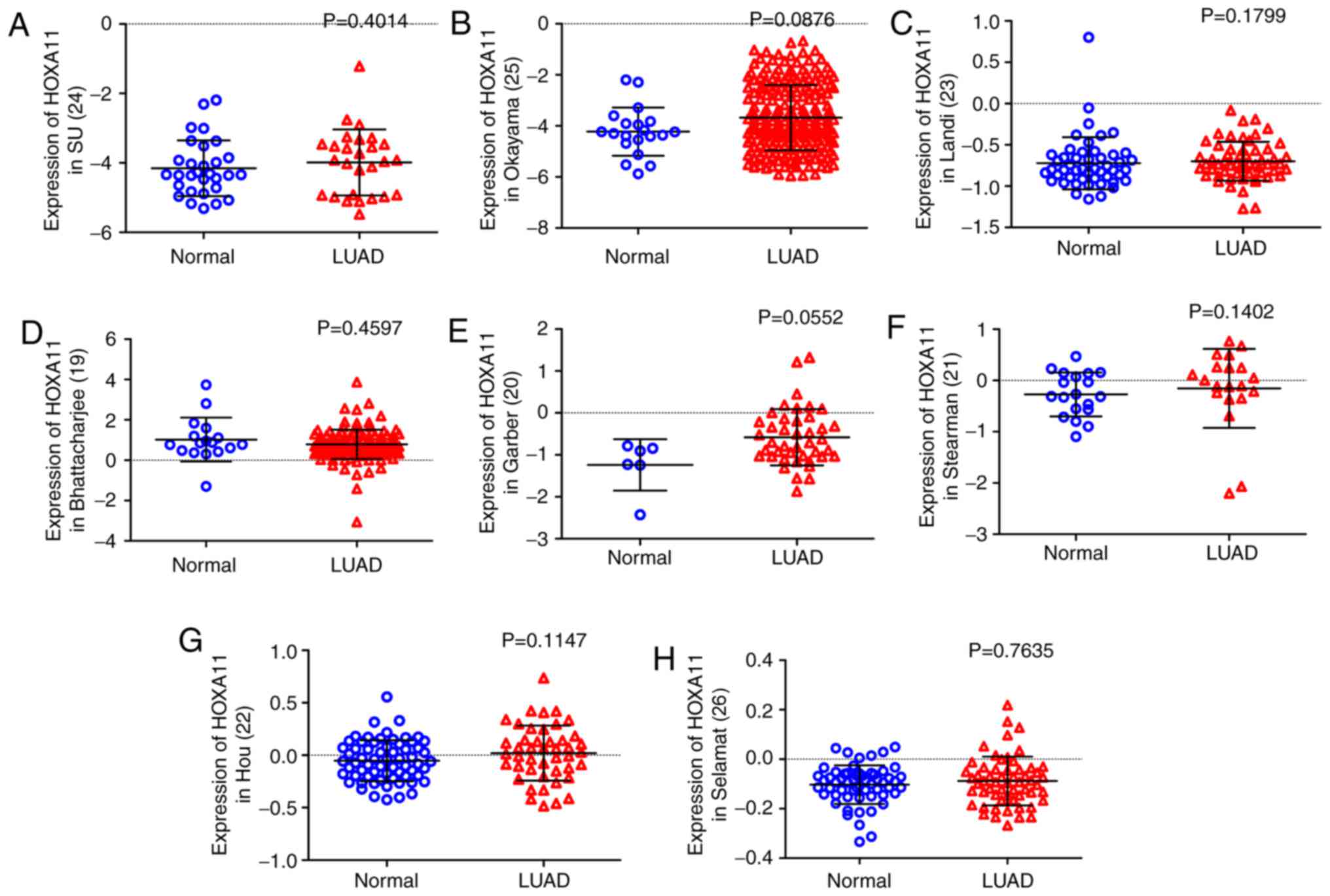
95.5%). Furthermore, except from the data of Bhattacharjee et
al (19), all of the eight
datasets obtained from the Oncomine database indicated that HOXA11
expression in LUAD tissues was higher than that in the normal
controls (20-26), although all the P-values were
>0.05 (Fig. 2). Among the
eight datasets, that by Garber et al (20) revealed that HOXA11 had a moderate
diagnostic value for LUAD, while it was poor in the other studies.
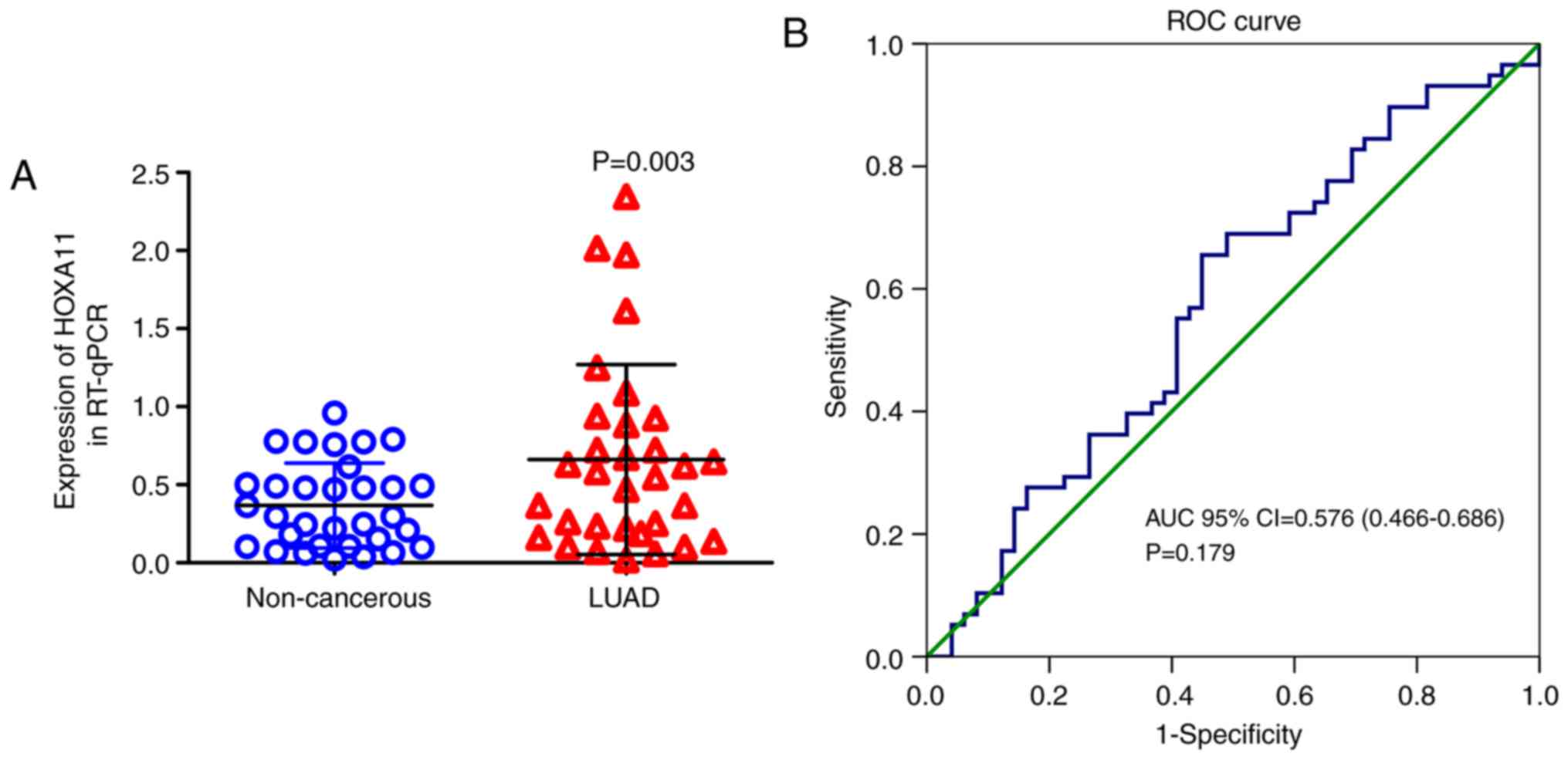
In the present study, HOXA11 expression was also detected in 32
pairs of formalin-fixed, paraffin-embedded tumors and adjacent
non-cancerous tissues by RT-qPCR. These in-house data suggested
that HOXA11 was significantly overexpressed in the LUAD vs. normal
tissue samples (P=0.003) (Table
II). Furthermore, HXOA11 was indicated to be of diagnostic
value for LUAD, as the AUC was 0.632 (95% CI: 0.495-0.770, P=0.069;
Fig. 3), with a cutoff value of
0.5236 (sensitivity 53.1% and specificity 78.1%).
 | Table IExpression of HOXA11 in LUAD based on
the The Cancer Genome Atlas database. |
Table I
Expression of HOXA11 in LUAD based on
the The Cancer Genome Atlas database.
| Clinicopathological
feature | N (%) | HOXA11 expression
(log2) | T- or F-value | P-value |
|---|
| Tissue type | | | | |
| Normal | 22 (6.8) | 1.41±0.937 | T=3.239 | 0.001 |
| LUAD | 302 (93.2) | 3.15±2.506 | | |
| Age (years) | | | | |
| <60 | 96 (31.8) | 3.12±2.644 | T=−0.140 | 0.888 |
| ≥60 | 206 (68.2) | 3.16±2.446 | | |
| Sex | | | | |
| Male | 138 (45.7) | 3.20±2.558 | T=0.318 | 0.750 |
| Female | 164 (54.3) | 3.11±2.469 | | |
| Ethnicity | | | | |
| Caucasian | 226 (87.9) | 3.06±2.526 | F=0.212 | 0.809 |
| Black | 26 (10.1) | 3.29±2.270 | | |
| Asian | 5 (1.9) | 2.55±1.461 | | |
| Tumor stage | | | | |
| T1/T2 | 255 (85.0) | 3.13±2.507 | T=−0.422 | 0.673 |
| T3/T4 | 45 (15.0) | 3.30±2.561 | | |
| N stage | | | | |
| NX | 7 (2.3) | 1.65±0.992 | F=8.111 | <0.001 |
| N0/N1 | 243 (80.5) | 2.94±2.456 | | |
| N2/N3 | 52 (17.2) | 4.32±2.537 | | |
| M stage | | | | |
| MX | 81 (27.0) | 3.12±2.449 | F=0.142 | 0.867 |
| M0 | 202 (67.3) | 3.12±2.532 | | |
| M1 | 17 (5.7) | 3.45±2.655 | | |
| Clinical stage | | | | |
| I/II | 177 (59.0) | 2.89±2.395 | T=−2.127 | 0.034 |
| III/IV | 123 (41.0) | 3.53±2.648 | | |
| Survival
status | | | | |
| Dead | 121 (40.1) | 3.48±2.683 | T=1.827 | 0.069 |
| Alive | 181 (59.9) | 2.93±2.363 | | |
| Recurrence | | | | |
| Distant
metastasis | 53 (58.2) | 3.15±2.559 | F=1.362 | 0.261 |
| Loco-regional
recurrence | 35 (38.5) | 2.61±2.526 | | |
| New primary
tumor | 3 (3.3) | 0.97±1.053 | | |
| Table IIExpression of HOXA11 in LUAD tissues
based on RT-qPCR in house. |
Table II
Expression of HOXA11 in LUAD tissues
based on RT-qPCR in house.
| Clinicopathological
features | N (%) | HOXA11 expression
(2 −ΔΔCq)
| P-value |
|---|
| Mean ± SD | T- or F-value |
|---|
| Tissues | | | | 0.003 |
| Non-tumor | 32 (50.0) | 0.3667±0.27144 | T=3.201 | |
| LUAD | 32 (50.0) | 0.6616±0.60821 | | |
| Size | | | | 0.877 |
| ≤3 cm | 9 (28.1) | 0.6888±0.71826 | T=0.156 | |
| >3 cm | 23 (71.9) | 0.6509±0.57727 | | |
| TNM | | | | 0.615 |
| I-II | 19 (59.4) | 0.7073±0.70770 | T=0.508 | |
| III-IV | 13 (40.6) | 0.5947±0.44298 | | |
| Sex | | | | 0.786 |
| Male | 23 (71.9) | 0.6429±0.65476 | T=−0.274 | |
| Female | 9 (28.1) | 0.7093±0.50092 | | |
| Age | | | | 0.659 |
| <60 years | 20 (62.5) | 0.6240±0.52890 | T=−0.446 | |
| ≥60 years | 12 (37.5) | 0.7243±0.74329 | | |
| Smoking | | | | 0.793 |
| No | 18 (56.3) | 0.6361±0.43031 | T=−0.265 | |
| Yes | 14 (43.7) | 0.6943±0.79871 | | |
| Vascular
invasion | | | | 0.154 |
| No | 30 (93.8) | 0.6217±0.57800 | T=−1.463 | |
| Yes | 2 (6.2) | 1.2598±1.00785 | | |
| LNM | | | | 0.920 |
| No | 18 (56.3) | 0.6518±0.73299 | T=−0.102 | |
| Yes | 14 (43.7) | 0.6742±0.42335 | | |
| Grade | | | | 0.383 |
| I | 5 (15.6) | 0.4371±0.24929 | F=0.991 | |
| II | 24 (75.0) | 0.7480±0.67319 | | |
| III | 3 (9.4) | 0.3440±0.17608 | | |
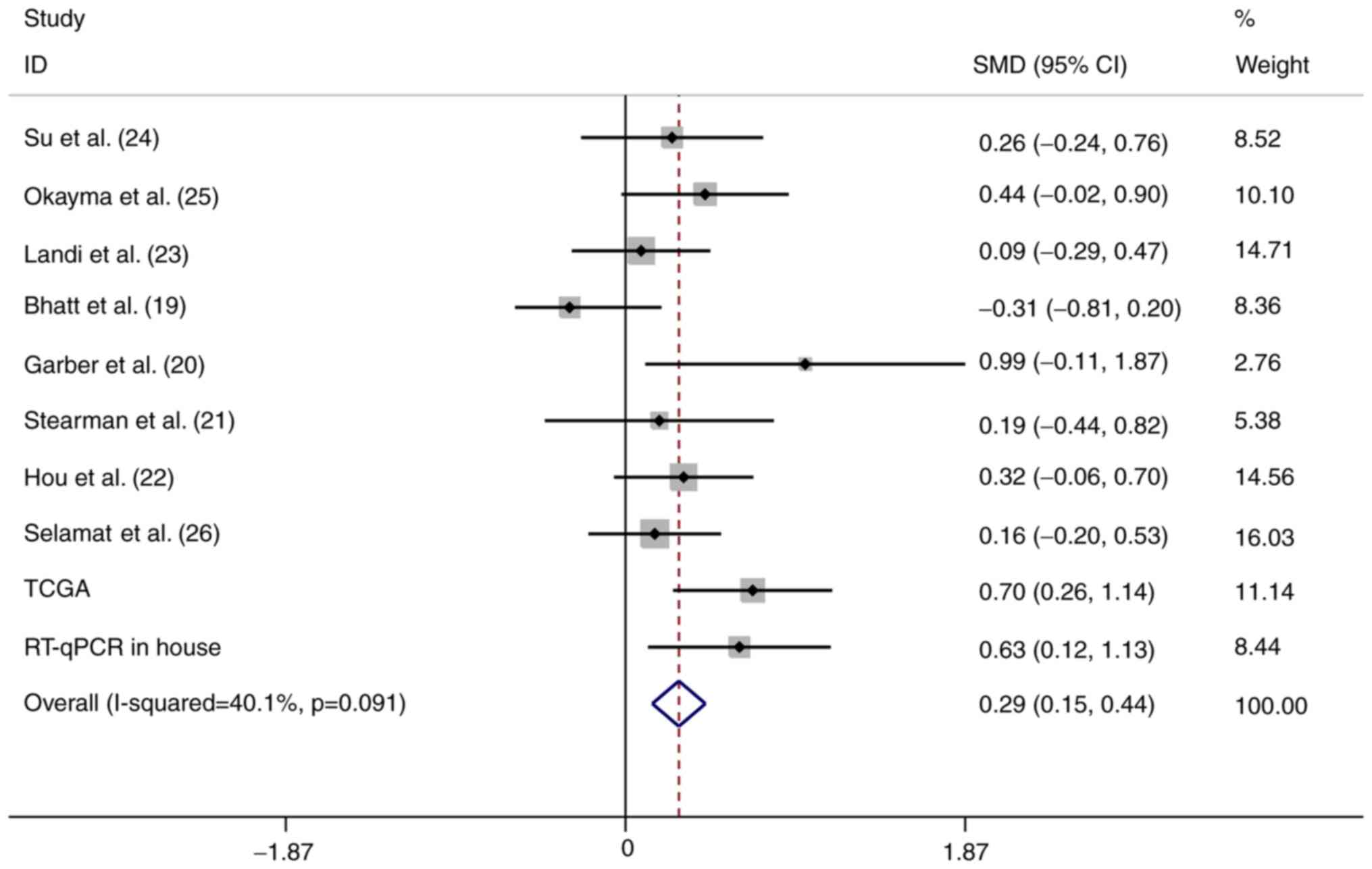
Meta-analysis
To strengthen the reliability of the results, a
meta-analysis of the data from the three different sources was
performed. In total, 934 LUAD and 319 normal control samples (159
were normal tissues from healthy subjects and 160 were
tumor-adjacent tissues) were included. A fixed-effects model was
selected and the pooled SMD of the 10 studies was 0.29 (95% CI:
0.15-0.44). A significant difference was identified in the
expression of HOXA11 between LUAD and normal tissues, and the
heterogeneity among the individual datasets was low
(I2=40.1%, P=0.091; Fig.
4). This result further proved that HOXA11 was overexpressed in
LUAD. In addition, the potential diagnostic value of HOXA11 in LUAD
was analyzed using the merged data, and the meta-analysis revealed
that the pooled AUC for the diagnostic value of HOXA11 for LUAD was
0.69 (95% CI: 0.64-0.73; Fig.
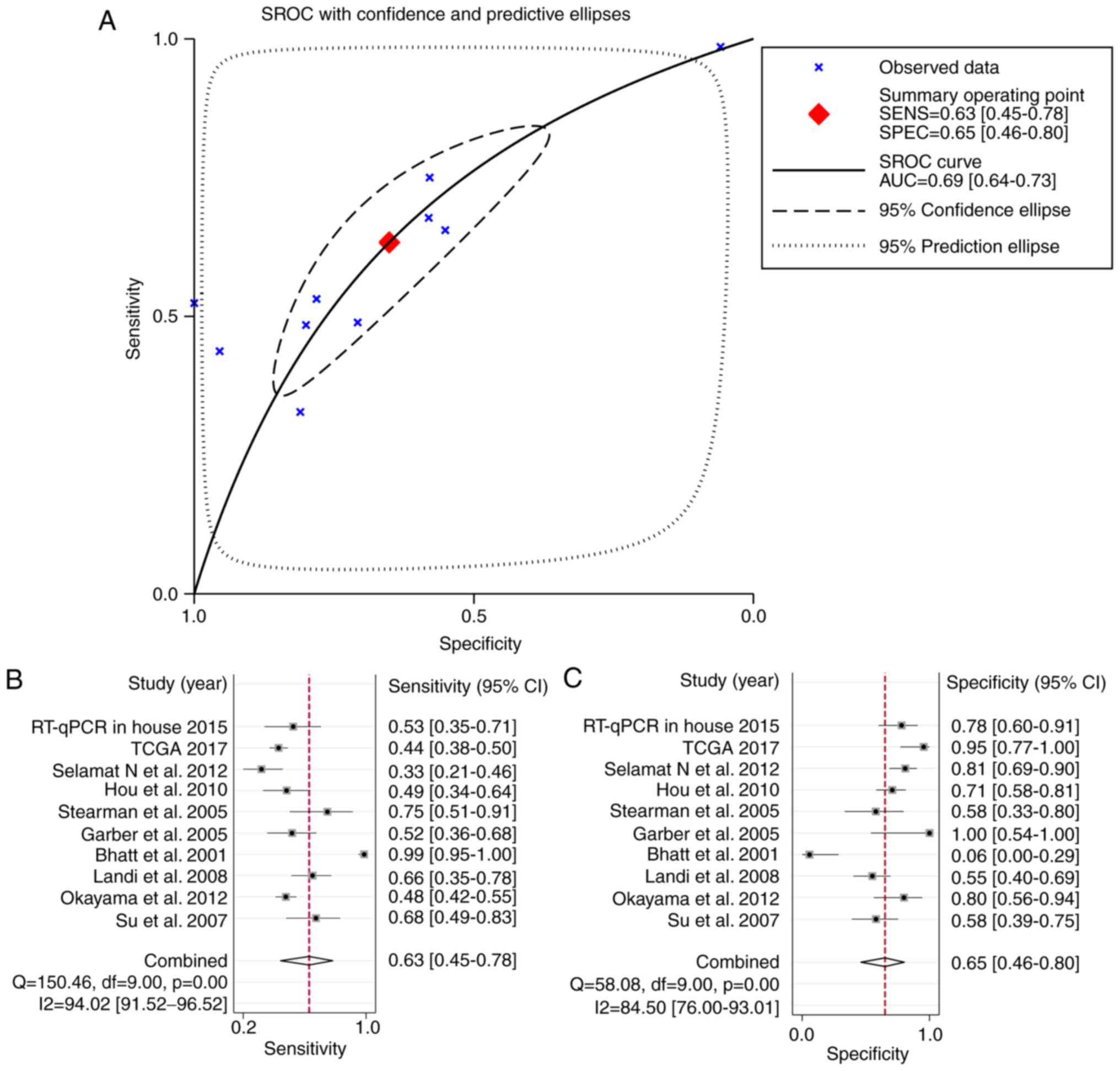
5A). The combined sensitivity and specificity were 0.63 (95%
CI: 0.45-0.78) and 0.65 (95% CI: 0.46-0.80), respectively (Fig. 5A and B), indicating that HOXA11
probably has a role in the tumorigenesis of LUAD.
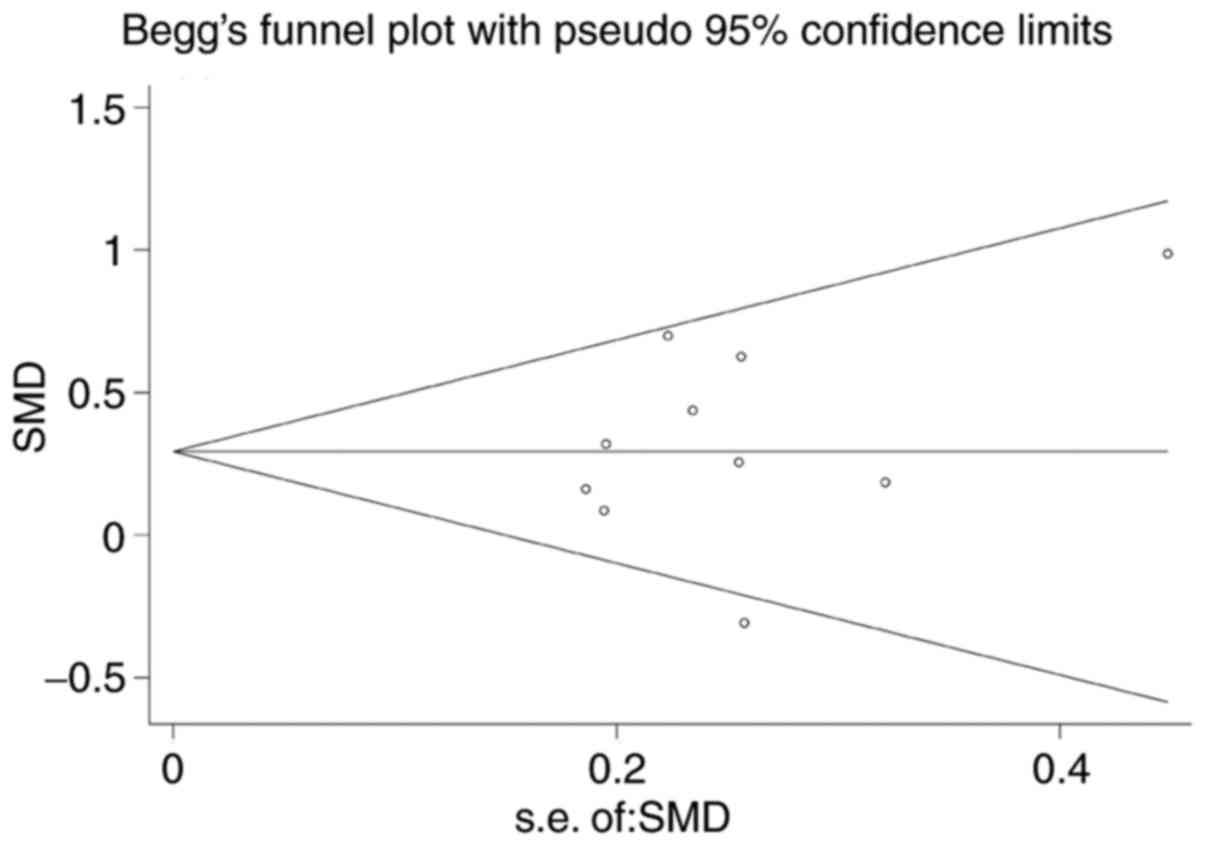
Begg's funnel plot was performed for all of the
datasets (19-26) to assess publication bias, and the
results yielded P=0.138. This result suggested the absence of
publication bias in the present study (Fig. 6).
Genes co-expressed with HOXA11
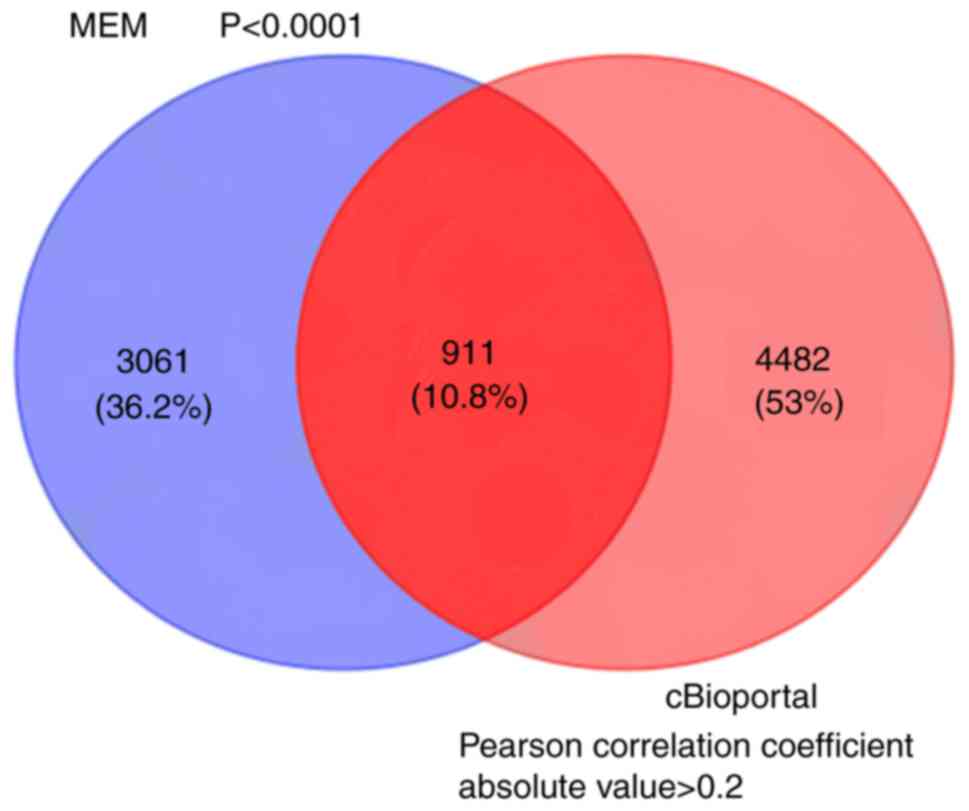
A total of 3,972 genes co-expressed with HOXA11 were
extracted from the MEM database with two independent gene probe
tests. According to the co-expression analysis in the cBioPortal
database, the mRNAs of 5,393 genes were identified as being
co-expressed with HOXA11. Ultimately, to more accurately obtain
genes co-expressed with HOXA11, the co-expressed genes extracted
from the MEM and cBioPortal databases were overlapped and 911 were
thereby identified for further analysis (Fig. 7).
Bioinformatics analysis
To explore the mechanisms and pathways of HOXA11 and
its co-expressed genes, the 911 co-expressed genes selected were
subjected to in silico analysis using the DAVID and STRING
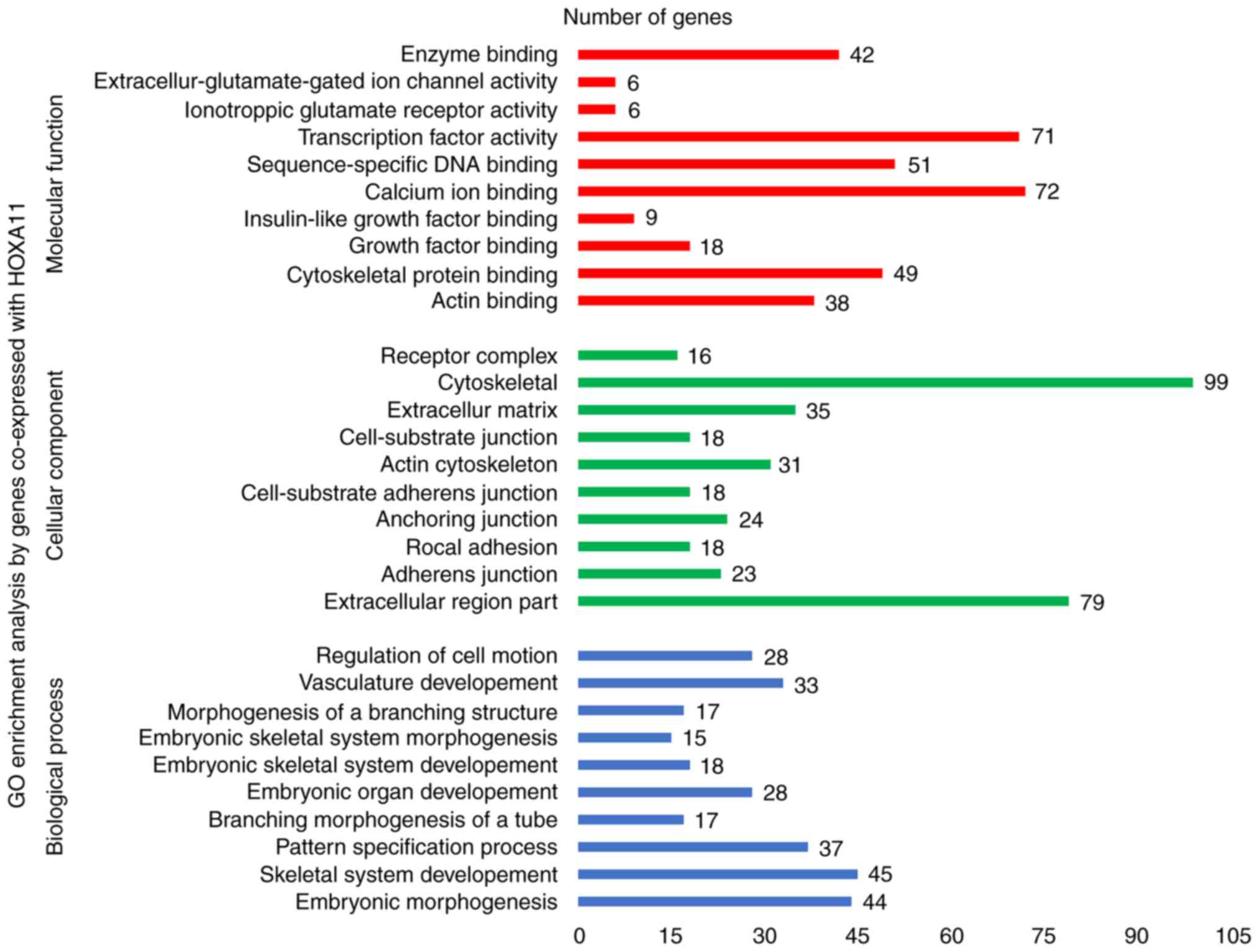
online tools. According to the GO enrichment analysis, these
co-expressed genes were mainly enriched in the terms 'vasculature
development' (GO:0001944, P=9.3×10−7), 'regulation of
cell motion' (GO:0051270, P=1.08×10−6), 'cell adhesion'
(GO:0007155, P=6.52×10−6) and 'biological adhesion'
(GO:0022610, P=6.58×10−6) in the category BP; 'adherens
junction' (GO:0005912, P=7×10−6), 'anchoring junction'
(GO:0070161, P=1.2×10−5), 'receptor complex'
(GO:0043235, P=5.62×10−4) and 'cell junction'
(GO:0030054, P=1.86×10−3) in the category CC; and 'actin
binding' (GO:0003779, P=2×10−6), 'growth factor binding'
(GO:0019838, P=1.3×10−5), 'calcium ion binding'
(GO:0005509, P=8.8×10−5) and 'transcription regulator
activity' (GO:0030528, P=9.5×10−3) in the category MF.
The GO functional annotation results are presented in Table III and Fig. 8 with the top 10 significantly
enriched terms by the co-expressed genes provided for each
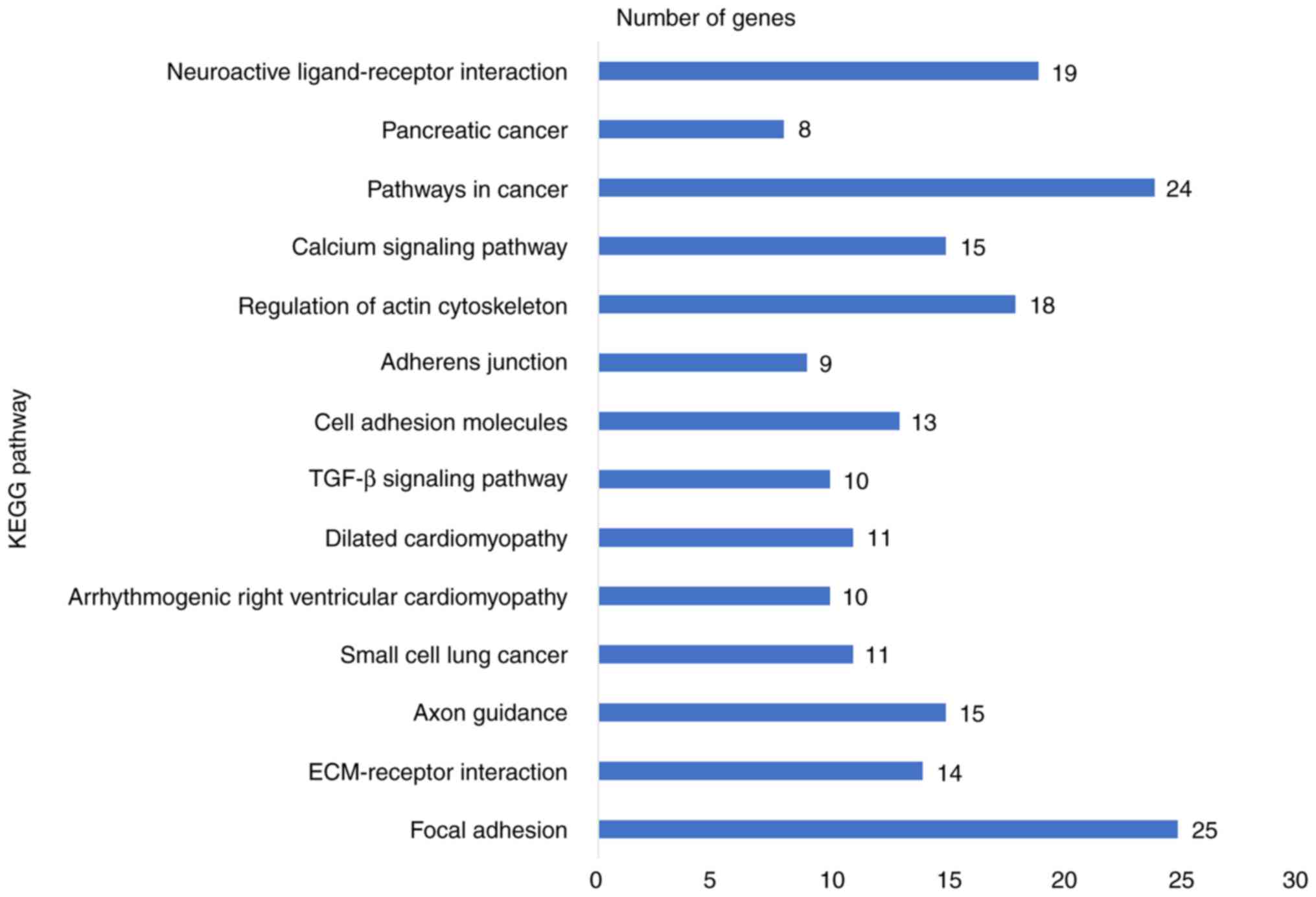
category. KEGG pathway analysis revealed a significant enrichment
of the co-expressed genes in the pathways 'focal adhesion'
(hsa04510; P=6.5×10−5), 'extracellular matrix
(ECM)-receptor interaction' (hsa04512; P=2.7×10−4),
'axon guidance' (hsa04360; P=5.4×10−3) and 'small-cell
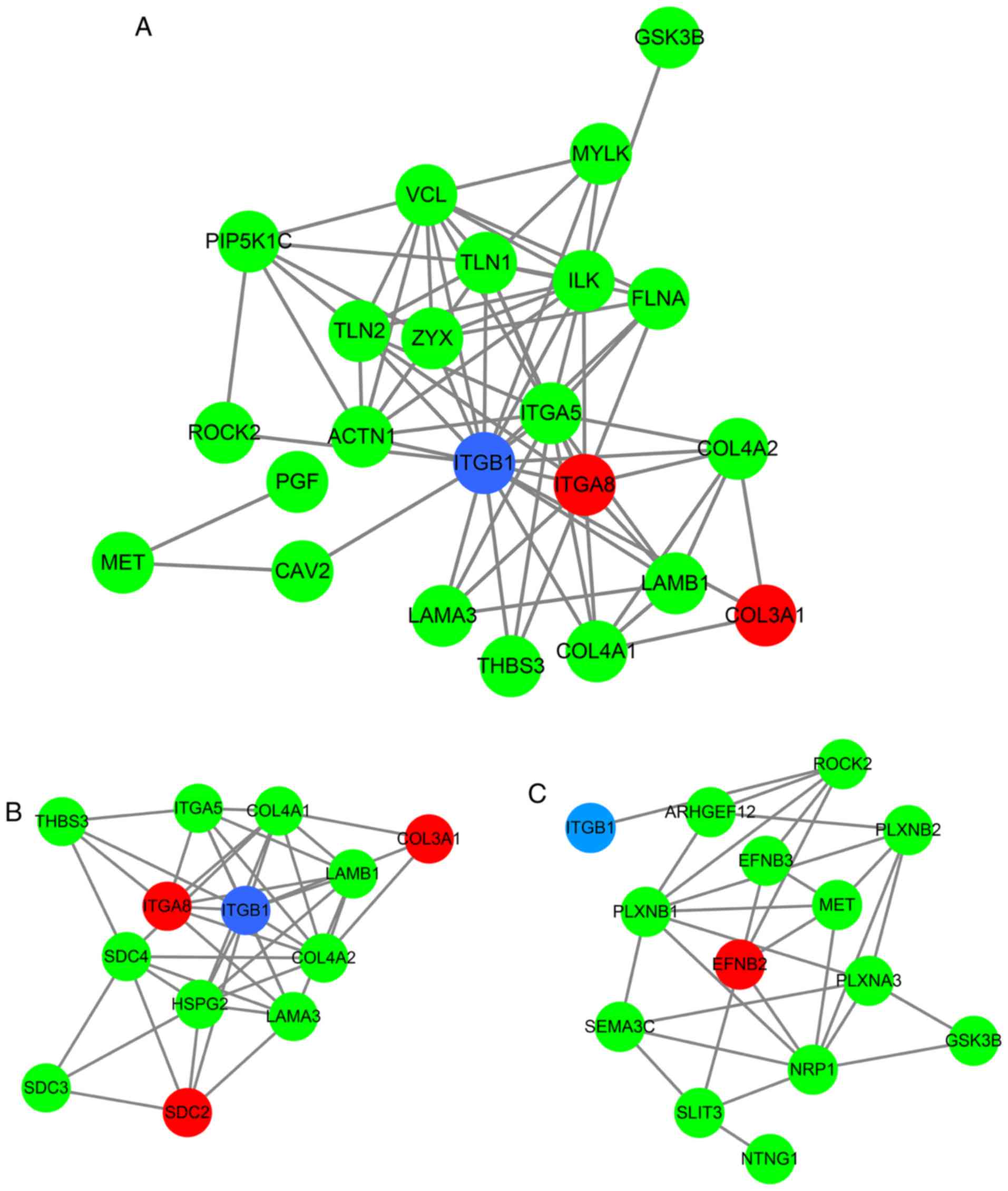
lung cancer' (hsa05222; P=9.5×10−3; Table IV and Fig. 9). To investigate the interaction
of the genes enriched in the top three pivotal KEGG pathways (focal
adhesion, ECM-receptor interaction and axon guidance), a PPI
network was constructed (Fig.
10). The network analysis revealed that integrin β1 (ITGB1) and
neuropilin 1 (NRP1) are the most important hub genes in the three
pivotal pathways mentioned above. Pearson analysis of the
correlation between HOXA11 and these two hub genes was performed,
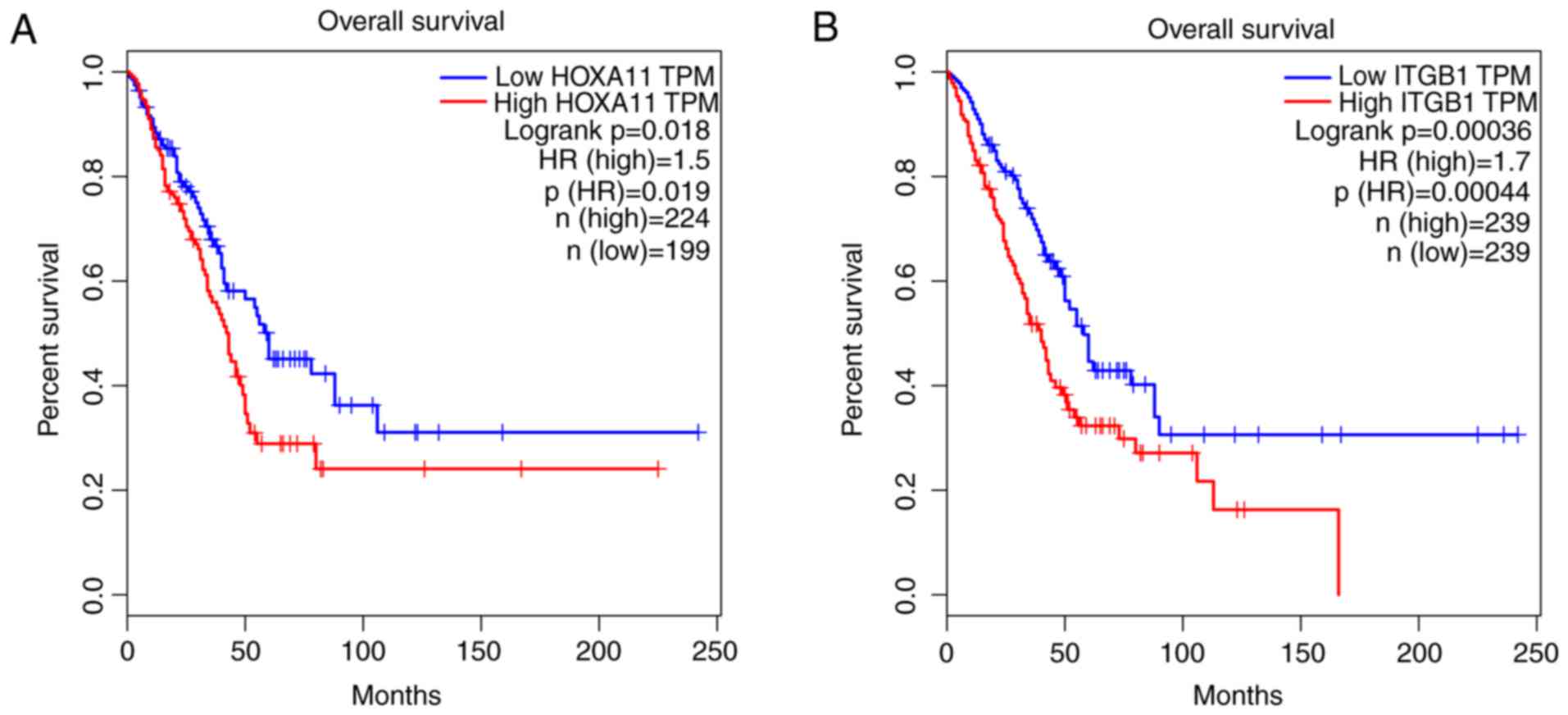
and ITCB1 was identified to be negatively correlated with HOXA11.
Thus, an analysis of the influence of HOXA11 and ITGB1 on the
overall survival of LUAD patients was then performed. Of note, LUAD
patients with low expression levels of HOXA11 or ITGB1 in the
tumour tissues had better overall survival rates than those with
high expression (Fig. 11).
Furthermore, high-throughput sequencing data and
immunohistochemical staining confirmed that the mRNA and protein
levels of several molecules in the top three KEGG pathways were
dysregulated. Collagen type III α1 chain (COL3A1) was significantly
upregulated in the 'ECM-receptor interaction' pathway, ephrin B2
(EFNB2) and syndecan 2 (SDC2) were separately downregulated in the
'axon guidance' and 'ECM-receptor interaction' pathways, and ITGA8
was significantly and concurrently downregulated in the 'focal
adhesion' and 'ECM-receptor interaction' pathways (Fig. 12). The aberrant expression of
these genes may cause abnormal changes in the above three signaling
pathways to induce the tumorigenesis and progression of LUAD.
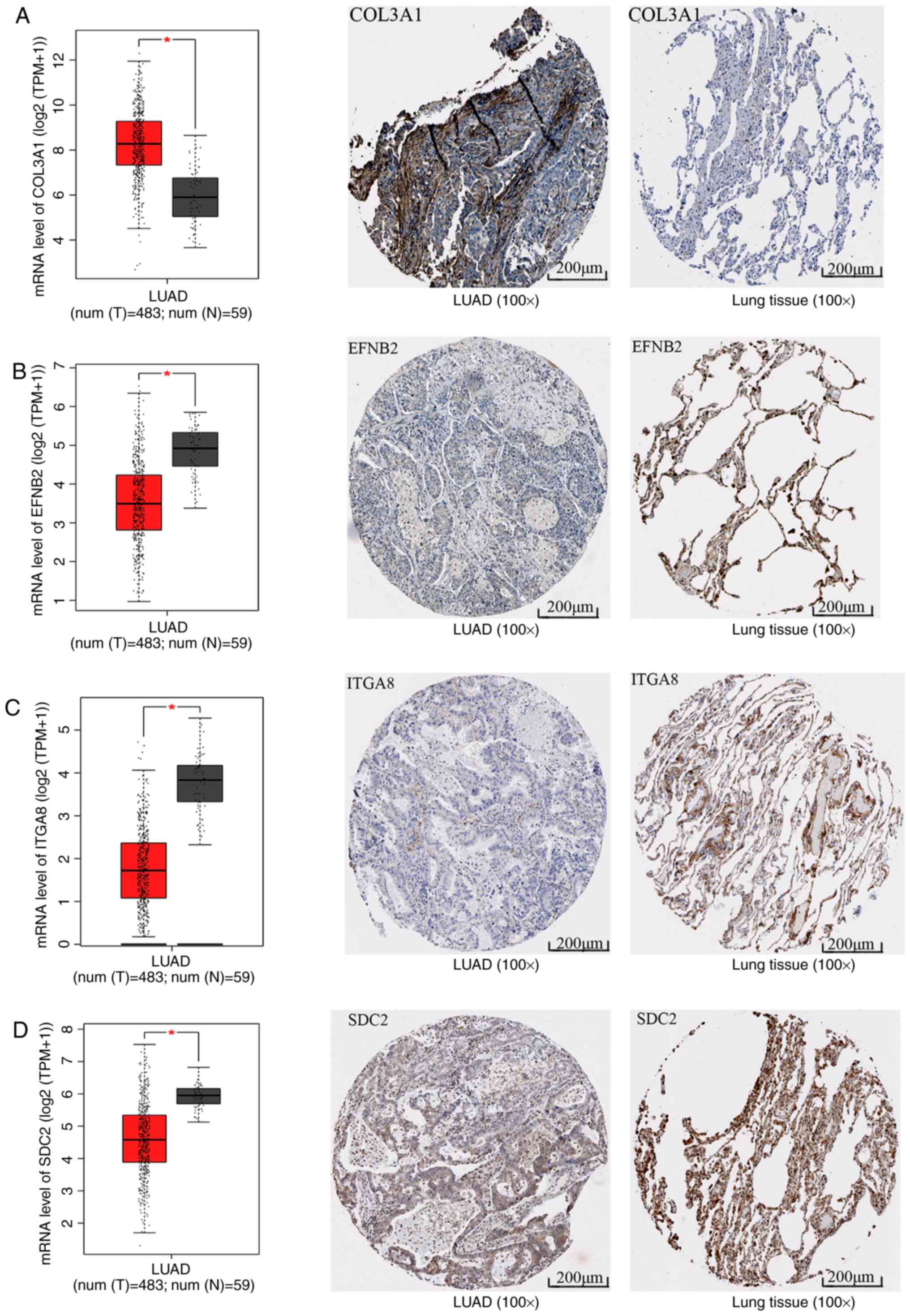 | Figure 12mRNA and protein levels of molecules
in the top three Kyoto Encyclopedia of Genes and Genomes pathways.
The box plots represent the mRNA levels of the molecules; the red
boxes represent the LUAD group and the gray boxes the normal
controls. The five horizontal lines in the box plots from top to
bottom represent the upper limit, upper quartile, median value,
lower quartile and lower limit, respectively. Each black dot
represents a tissue sample. In the immunohistochemical staining
images, the left panel represents LUAD and the right panel
represents the normal control. (A) COL3A1, (B) EFNB2, (C) ITGA8 and
(D) SDC2. *P<0.05. LUAD, lung adenocarcinoma; COL3A1,
collagen type III α1 chain; EFNB2, ephrin B2; ITGA8, integrin
subunit α8; SDC, syndecan. |
| Table IIITop 10 significantly enriched GO
terms in three categories by genes co-expressed with homeobox
A11. |
Table III
Top 10 significantly enriched GO
terms in three categories by genes co-expressed with homeobox
A11.
| Category/term | Count | P-value | FDR |
|---|
| Biological
process | | | |
|
GO:0048598-Embryonic morphogenesis | 44 |
6.79×10−10 | 0.000001 |
|
GO:0001501-Skeletal system
development | 45 |
7.03×10−10 | 0.000001 |
| GO:0007389-Pattern
specification process | 37 |
4.66×10−8 | 0.000084 |
|
GO:0048754-Branching morphogenesis of a
tube | 17 |
7.79×10−8 | 0.000140 |
|
GO:0048568-Embryonic organ
development | 28 |
1.02×10−7 | 0.000183 |
|
GO:0048706-Embryonic skeletal system
development | 18 |
1.71×10−7 | 0.000306 |
|
GO:0048704-Embryonic skeletal system
morphogenesis | 15 |
5.22×10−7 | 0.000938 |
|
GO:0001763-Morphogenesis of a branching
structure | 17 |
5.32×10−7 | 0.000955 |
|
GO:0001944-Vasculature development | 33 |
9.30×10−7 | 0.001669 |
|
GO:0051270-Regulation of cell motion | 28 |
1.08×10−6 | 0.001945 |
| Cellular
component | | | |
|
GO:0044421-Extracellular region part | 79 |
7.0×10−6 | 0.009522 |
|
GO:0005912-Adherens junction | 23 |
7.0×10−6 | 0.010227 |
| GO:0005925-Rocal
adhesion | 18 |
9.0×10−6 | 0.013065 |
|
GO:0070161-Anchoring junction | 24 |
1.2×10−6 | 0.017438 |
|
GO:0005924-Cell-substrate adherens
junction | 18 |
1.6×10−6 | 0.022116 |
| GO:0015629-Actin
cytoskeleton | 31 |
2.5×10−6 | 0.035229 |
|
GO:0030055-Cell-substrate junction | 18 |
3.2×10−6 | 0.046211 |
|
GO:0031012-Extracellular matrix | 35 |
9.3×10−6 | 0.133109 |
|
GO:0005856-Cytoskeleton | 99 |
1.1×10−5 | 0.161576 |
|
GO:0043235-Receptor complex | 16 |
5.6×10−5 | 0.798459 |
| Molecular
function | | | |
| GO:0003779-Actin
binding | 38 |
2.0×10−6 | 0.002752 |
|
GO:0008092-Cytoskeletal protein
binding | 49 |
8.0×10−6 | 0.012857 |
| GO:0019838-Growth
factor binding | 18 |
1.3×10−6 | 0.020705 |
|
GO:0005520-Insulin-like growth factor
binding | 9 |
1.7×10−6 | 0.026080 |
| GO:0005509-Calcium
ion binding | 72 |
8.8×10−6 | 0.135240 |
|
GO:0043565-Sequence-specific DNA
binding | 51 |
2.3×10−5 | 0.357942 |
|
GO:0003700-Transcription factor
activity | 71 |
8.2×10−5 | 1.256619 |
|
GO:0004970-Ionotropic glutamate receptor
activity | 6 |
1.4×10−4 | 2.177531 |
|
GO:0005234-Extracellular-glutamate-gated
ion channel activity | 6 |
1.8×10−4 | 2.828013 |
| GO:0019899-Enzyme
binding | 42 |
2.2×10−4 | 3.271727 |
| Table IVKyoto Encyclopedia of Genes and
Genomes pathways enriched by genes co-expressed with homeobox
A11. |
Table IV
Kyoto Encyclopedia of Genes and
Genomes pathways enriched by genes co-expressed with homeobox
A11.
| Pathway | n | P-value | FDR |
|---|
| hsa04510: Focal
adhesion | 25 |
6.50×10−6 | 0.077743 |
| hsa04512:
ECM-receptor interaction | 14 |
2.68×10−5 | 0.318914 |
| hsa04360: Axon
guidance | 15 |
5.41×10−4 | 6.249372 |
| hsa05222: Small
cell lung cancer | 11 |
9.48×10−4 | 10.714508 |
| hsa05412:
Arrhythmogenic right ventricular cardiomyopathy | 10 |
1.40×10−3 | 15.453584 |
| hsa05414: Dilated
cardiomyopathy | 11 |
1.74×10−3 | 18.830973 |
| hsa04350: TGF-β
signaling pathway | 10 |
3.12×10−3 | 31.416058 |
| hsa04514: Cell
adhesion molecules | 13 |
3.47×10−3 | 34.317283 |
| hsa04520: Adherens
junction | 9 |
4.01×10−3 | 38.569481 |
| hsa04810:
Regulation of actin cytoskeleton | 18 |
4.43×10−3 | 41.646034 |
| hsa04020: Calcium
signaling pathway | 15 |
6.18×10−3 | 53.213195 |
| hsa05200: Pathways
in cancer | 24 |
6.82×10−3 | 56.834134 |
| hsa05212:
Pancreatic cancer | 8 |
7.11×10−3 | 58.417821 |
| hsa04080:
Neuroactive ligand-receptor interaction | 19 |
9.72×10−3 | 70.392280 |
Discussion
HOX genes have been reported to encode for
transcription factors that have essential roles in the embryonic
development and differentiation of adult cells, and their altered
expression has also been identified in cancer (27-29). Several studies also reported that
HOXA11 was hypermethylated in lung cancer (1,30,31). However, the expression of HOXA11
in cancer and whether altered HOXA11 expression may contribute to
the genesis of lung cancer remains to be fully elucidated. In the
present study, the expression of HOXA11 in LUAD patients was
determined using data obtained from TCGA database, revealing that
HOXA11 was overexpressed in LUAD samples compared with its
expression in adjacent, non-tumorous tissues. To gain further
insight into the role of HOXA11 in LUAD, a total of 8 microarrays
collected from the Oncomine database, as well as in-house RT-qRCR
data, were used to validate the results from TCGA. Most of the
microarray data indicated that HOXA11 expression in LUAD was higher
than that in the normal samples, but only the results of two
microarrays exhibited statistical significance. This may be for two
possible reasons: The sample size of LUAD patients and
tumor-adjacent tissues was small, while another likely cause is
that, for certain microarrays, the sample size was unbalanced
between the two compared groups (tumor and normal groups). However,
the RT-qRCR data of the present study also suggested that HOXA11
was significantly upregulated in LUAD. Furthermore, to verify
whether HOXA11 was differentially expressed between LUAD and normal
lung tissue, comprehensive data produced by various research
methods were used and a huge amount of data was analyzed to enhance
the reliability of the results.
The meta-analysis of data gathered from three
sources, with 934 LUAD and 319 normal controls, also revealed that
HOXA11 was significantly overexpressed in LUAD [SMD=0.29 and 95%CI:
0.15-0.44], which indicated that HOXA11 has a role in the genesis
of LUAD. In addition, analysis of the association between HOXA11
expression and clinical parameters indicated that the levels of
HOXA11 were significantly higher in patients with lymphoid
metastasis and advanced clinical stage, two important clinical
parameters in cancer progression. For the other parameters,
including tumor size, metastasis and survival status, no
statistical significance was identified for their association with
HOXA11 expression, while a higher level of HOXA11 was noted in risk
groups (large tumor size, distant metastasis and deceased
patients). The uneven sample size between the two compared groups
may be the major cause for the insignificant difference
(P>0.05). A further limitation of the present study is that the
normal controls in included tissues from healthy individuals and
tumor-adjacent samples. The tumor-adjacent samples of LUAD patients
are located by inflammatory lesions, which may affect gene
expression. Therefore, sample selection may be another reason for
the insignificant differences observed. However, the results of the
present study still suggested that HOXA11 may act as an oncogene in
the initiation and progression of LUAD. HOXA11 may be used as a
clinical predictor for the survival of LUAD patients. In future
studies, it is required to further verify the role of HOXA11
expression in LUAD development/progression with a larger sample
size and to identify the underlying regulatory mechanisms.
Due to the insufficient number of studies
investigating the role of HOXA11 in cancer, the modes and molecular
pathways via which upregulated HOXA11 exerts its function in LUAD
have remained to be elucidated. Hence, the present study attempted
to predict the potential mechanisms of the role of HOXA11 in LUAD
in silico. With the development of high-throughput
sequencing and novel computational approaches, accumulating
evidence has proven that multiple genes always function together
and then regulate tumor initiation and progression (32). To investigate the function of
HOXA11, the cBioPortal and MEM databases were used in the present
study to identify the genes co-expressed with HOXA11. The 911
co-expressed genes obtained were inputted into the DAVID online
tool for predicting the pathways through which these genes are
involved in LUAD progression. The KEGG pathway analysis results
suggested that HOXA11 and its co-expressed genes were significantly
enriched in the focal adhesion, ECM-receptor interaction and axon
guidance pathways. The focal adhesion pathway has been reported to
be correlated with cellular behavior, including migration and
metastasis (33). From these
results it may be deduced that HOXA11 promotes the development and
progression of LUAD by affecting cancer cell adhesion and
migration.
The genes enriched in the above pathways (focal
adhesion, ECM-receptor interaction and axon guidance) were used to
construct PPI networks, in which ITGB1 and NRP1 were identified as
the hub genes. ITGB1 is a member of the integrin family and forms
various heterodimeric receptors for cell adhesion to ECM proteins.
Integrins are known to have important roles in regulating
proliferation, cell migration, invasion and survival (34). Previous studies reported that the
expression of ITGB1 is associated with the prognosis of patients
with lung cancer (35),
colorectal cancer (34) and
breast cancer (36). However, its
value as a prognostic marker, as well as its correlation with the
expression of other genes in LUAD patients, has rarely been
studied. In the present study, a Pearson correlation analysis was
performed, which indicated that ITGB1 expression is positively
correlated with HOXA11 in LUAD. Further investigation of the
prognostic value with Kaplan-Meier analysis and a log-rank test
also revealed that low expression levels of HOXA11 and ITGB1 were
associated with a better overall survival rate of LUAD patients.
However, logistic regression analysis has not been performed in the
current study, it would be assessed in a future study. The mRNA
(analysed using TCGA data) and protein levels (from the human
protein atlas database) of COL3A1 were identified to be upregulated
in LUAD tissues, while those of EFNB2, ITGA8 and SDC2 were
significantly downregulated in LUAD. Therefore, it may be
speculated that HOXA11 probably binds with ITGB1, COL3A1, EFNB2,
ITGA8 and SDC2, which then get involved in the focal adhesion
pathway to promote the initiation and progression of LUAD. Further
experimental verification is required to investigate whether these
potential targets were transcriptionally activated by HOXA11
protein binding with their activation sequence in the 3′UTR, which
may promote the development and progression of LUAD.
In the present study the expression and potential
clinical value of HOXA11 in LUAD patients was comprehensively
analyzed using various data sources and methods. The results
revealed that HOXA11 is overexpressed in LUAD. In addition, the
bioinformatics analysis suggested that HOXA11 probably combines
with ITGB1, COL3A1, EFNB2, ITGA8 and SDC2, which get involved in
the focal adhesion pathway and then regulate the emergence and
development of LUAD. However, the present study has certain
limitations. First, the expression of HOXA11 was detected in only
32 pairs of LUAD and adjacent non-tumorous tissues. The normal
controls in the current study included tissues from healthy
individual combined with tumor-adjacent samples. Further
investigations with an even sample size and even distribution of
data are required to confirm the value of HOXA11 as a potential
target for LUAD. In addition, the mechanism of HOXA11 in LUAD was
predicted in silico. Further studies are required to
experimentally investigate the molecular mechanisms of HOXA11 in
LUAD.
Funding
The study was supported by funds from the National
Natural Science Foundation of China (grant nos. NSFC81560469 and
NSFC81360327), the Natural Science Foundation of Guangxi, China
(grant nos. 2017GXNSFAA198016 and 2015GXNSFCA139009), the Guangxi
Medical University Training Program for Distinguished Young
Scholars (Gang Chen) and a Medical Excellence Award funded by the
Creative Research Development Grant from the First Affiliated
Hospital of Guangxi Medical University (Gang Chen).
Availability of data and materials
The datasets used and/or analysed during the current
study are available from the corresponding author on reasonable
request.
Authors' contributions
XY and YD designed the study, performed the
experiments, analyzed and interpreted the data and wrote the
manuscript. They contributed equally to the present study. XY, RQH
and XJL performed the statistical analysis, and designed and
completed the figures and tables. YD and XJL recruited the patients
to obtain their specimens and performed the mRNA isolation and
RT-qPCR. JM, GC and XHH participated in the design of the study,
supervised the experiments and corrected the manuscript. All of the
authors read and approved the final manuscript.
Ethics approval and consent to
participate
All experiments using patient tissues were
authorized by the Ethics Committee of the First Affiliated Hospital
of Guangxi Medical University (Guangxi, China).
Patient consent for publication
All subjects gave their informed consent prior to
their inclusion in the study. Details that may disclose their
identity were omitted.
Competing interests
The authors declare that they have no competing of
interest.
Acknowledgments
Not applicable.
References
|
1
|
Hwang JA, Lee BB, Kim Y, Park SE, Heo K,
Hong SH, Kim YH, Han J, Shim YM, Lee YS and Kim DH: HOXA11
hypermethylation is associated with progression of non-small cell
lung cancer. Oncotarget. 4:2317–2325. 2013. View Article : Google Scholar : PubMed/NCBI
|
|
2
|
Calvayrac O, Pradines A, Pons E, Mazières
J and Guibert N: Molecular biomarkers for lung adenocarcinoma. Eur
Respir J. 49:16017342017. View Article : Google Scholar : PubMed/NCBI
|
|
3
|
Sholl LM: Biomarkers in lung
adenocarcinoma: A decade of progress. Arch Pathol Lab Med.
139:469–480. 2015. View Article : Google Scholar
|
|
4
|
Wallerek S and Sørensen JB: Biomarkers for
efficacy of adjuvant chemotherapy following complete resection in
NSCLC stages I-IIIA. Eur Respir Rev. 24:340–355. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Qi L, Li Y, Qin Y, Shi G, Li T, Wang J,
Chen L, Gu Y, Zhao W and Guo Z: An individualised signature for
predicting response with concordant survival benefit for lung
adenocarcinoma patients receiving platinum-based chemotherapy. Br J
Cancer. 115:1513–1519. 2016. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Chalela R, Curull V, Enriquez C, Pijuan L,
Bellosillo B and Gea J: Lung adenocarcinoma: From molecular basis
to genome-guided therapy and immunotherapy. J Thorac Dis.
9:2142–2158. 2017. View Article : Google Scholar :
|
|
7
|
Wang L, Cui Y, Sheng J, Yang Y, Kuang G,
Fan Y, Jin J and Zhang Q: Epigenetic inactivation of HOXA11, a
novel functional tumor suppressor for renal cell carcinoma, is
associated with RCC TNM classification. Oncotarget. 8:21861–21870.
2017.PubMed/NCBI
|
|
8
|
Se YB, Kim SH, Kim JY, Kim JE, Dho YS, Kim
JW, Kim YH, Woo HG, Kim SH, Kang SH, et al: Underexpression of
HOXA11 is associated with treatment resistance and poor prognosis
in glioblastoma. Cancer Res Treat. 49:387–398. 2017. View Article : Google Scholar
|
|
9
|
Whitcomb BP, Mutch DG, Herzog TJ, Rader
JS, Gibb RK and Goodfellow PJ: Frequent HOXA11 and THBS2 promoter
methylation, and a methylator phenotype in endometrial
adenocarcinoma. Clin Cancer Res. 9:2277–2287. 2003.PubMed/NCBI
|
|
10
|
Speleman F, Cauwelier B, Dastugue N, Cools
J, Verhasselt B, Poppe B, Van Roy N, Vandesompele J, Graux C,
Uyttebroeck A, et al: A new recurrent inversion, inv(7)(p15q34),
leads to transcriptional activation of HOXA10 and HOXA11 in a
subset of T-cell acute lymphoblastic leukemias. Leukemia.
19:358–366. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
11
|
Xia B, Shan M, Wang J, Zhong Z, Geng J, He
X, Vu T, Zhang D and Pang D: Homeobox A11 hypermethylation
indicates unfavorable prognosis in breast cancer. Oncotarget.
8:9794–9805. 2017.
|
|
12
|
Li Q, Chen C, Ren X and Sun W: DNA
methylation profiling identifies the HOXA11 gene as an early
diagnostic and prognostic molecular marker in human lung
adenocarcinoma. Oncotarget. 8:33100–33109. 2017.PubMed/NCBI
|
|
13
|
Rhodes DR, Yu J, Shanker K, Deshpande N,
Varambally R, Ghosh D, Barrette T, Pandey A and Chinnaiyan AM:
ONCOMINE: A cancer microarray database and integrated data-mining
platform. Neoplasia. 6:1–6. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-Delta Delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
15
|
Chen G, Kronenberger P, Teugels E, Umelo
IA and De Grève J: Targeting the epidermal growth factor receptor
in non-small cell lung cancer cells: The effect of combining RNA
interference with tyrosine kinase inhibitors or cetuximab. BMC Med.
10:282012. View Article : Google Scholar : PubMed/NCBI
|
|
16
|
Luo YH, Tang W, Zhang X, Tan Z, Guo WL,
Zhao N, Pang SM, Dang YW, Rong MH and Cao J: Promising significance
of the association of miR-204 5p expression with
clinicopathological features of hepatocellular carcinoma. Medicine
(Baltimore). 96:e75452017. View Article : Google Scholar
|
|
17
|
Yang X, Pang YY, He RQ, Lin P, Cen JM,
Yang H, Ma J and Chen G: Diagnostic value of strand-specific
miRNA-101 3p and miRNA-101 5p for hepatocellular carcinoma and a
bioinformatic analysis of their possible mechanism of action. FEBS
Open Bio. 8:64–84. 2017. View Article : Google Scholar
|
|
18
|
Yang X, Zeng Z, Hou Y, Yuan T, Gao C, Jia
W, Yi X and Liu M: MicroRNA-92a as a potential biomarker in
diagnosis of colorectal cancer: A systematic review and
meta-analysis. PLoS One. 9:e887452014. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Bhattacharjee A, Richards WG, Staunton J,
Li C, Monti S, Vasa P, Ladd C, Beheshti J, Bueno R, Gillette M, et
al: Classification of human lung carcinomas by mRNA expression
profiling reveals distinct adenocarcinoma subclasses. Proc Natl
Acad Sci USA. 98:13790–13795. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
20
|
Garber ME, Troyanskaya OG, Schluens K,
Petersen S, Thaesler Z, Pacyna-Gengelbach M, van de Rijn M, Rosen
GD, Perou CM, Whyte RI, et al: Diversity of gene expression in
adenocarcinoma of the lung. Proc Natl Acad Sci USA. 98:13784–13789.
2001. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Stearman RS, Dwyer-Nield L, Zerbe L,
Blaine SA, Chan Z, Bunn PA Jr, Johnson GL, Hirsch FR, Merrick DT,
Franklin WA, et al: Analysis of orthologous gene expression between
human pulmonary adenocarcinoma and a carcinogen-induced murine
model. Am J Pathol. 167:1763–1775. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Hou J, Aerts J, den Hamer B, van Ijcken W,
den Bakker M, Riegman P, van der Leest C, van der Spek P, Foekens
JA, Hoogsteden HC, et al: Gene expression-based classification of
non-small cell lung carcinomas and survival prediction. PLoS One.
5:e103122010. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Landi MT, Dracheva T, Rotunno M, Figueroa
JD, Liu H, Dasgupta A, Mann FE, Fukuoka J, Hames M, Bergen AW, et
al: Gene expression signature of cigarette smoking and its role in
lung adenocarcinoma development and survival. PLoS One.
3:e16512008. View Article : Google Scholar : PubMed/NCBI
|
|
24
|
Su LJ, Chang CW, Wu YC, Chen KC, Lin CJ,
Liang SC, Lin CH, Whang-Peng J, Hsu SL, Chen CH and Huang CY:
Selection of DDX5 as a novel internal control for Q-RT-PCR from
microarray data using a block bootstrap re-sampling scheme. BMC
Genomics. 8:1402007. View Article : Google Scholar : PubMed/NCBI
|
|
25
|
Okayama H, Kohno T, Ishii Y, Shimada Y,
Shiraishi K, Iwakawa R, Furuta K, Tsuta K, Shibata T, Yamamoto S,
et al: Identification of genes upregulated in ALK-positive and
EGFR/KRAS/ALK-negative lung adenocarcinomas. Cancer Res.
72:100–111. 2012. View Article : Google Scholar
|
|
26
|
Selamat SA, Chung BS, Girard L, Zhang W,
Zhang Y, Campan M, Siegmund KD, Koss MN, Hagen JA, Lam WL, et al:
Genome-scale analysis of DNA methylation in lung adenocar-cinoma
and integration with mRNA expression. Genome Res. 22:1197–1211.
2012. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Shah N and Sukumar S: The Hox genes and
their roles in oncogenesis. Nat Rev Cancer. 10:361–371. 2010.
View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Javed S and Langley SE: Importance of HOX
genes in normal prostate gland formation, prostate cancer
development and its early detection. BJU Int. 113:535–540. 2014.
View Article : Google Scholar
|
|
29
|
Lopez-Romero R, Marrero-Rodriguez D,
Romero-Morelos P, Villegas V, Valdivia A, Arreola H, Huerta-Padilla
V and Salcedo M: The role of developmental HOX genes in cervical
cancer. Rev Med Inst Mex Seguro Soc. 53(Suppl 2): S188–S193.
2015.In Spanish.
|
|
30
|
Nelson HH, Marsit CJ, Christensen BC,
Houseman EA, Kontic M, Wiemels JL, Karagas MR, Wrensch MR, Zheng S,
Wiencke JK and Kelsey KT: Key epigenetic changes associated with
lung cancer development: Results from dense methylation array
profiling. Epigenetics. 7:559–566. 2012. View Article : Google Scholar : PubMed/NCBI
|
|
31
|
Bibikova M, Lin Z, Zhou L, Chudin E,
Garcia EW, Wu B, Doucet D, Thomas NJ, Wang Y, Vollmer E, et al:
High-throughput DNA methylation profiling using universal bead
arrays. Genome Res. 16:383–393. 2006. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Yu W, Zhao S, Wang Y, Zhao BN, Zhao W and
Zhou X: Identification of cancer prognosis-associated functional
modules using differential co-expression networks. Oncotarget.
8:112928–112941. 2017. View Article : Google Scholar
|
|
33
|
Eke I and Cordes N: Focal adhesion
signaling and therapy resistance in cancer. Semin Cancer Biol.
31:65–75. 2015. View Article : Google Scholar
|
|
34
|
Liu QZ, Gao XH, Chang WJ, Gong HF, Fu CG,
Zhang W and Cao GW: Expression of ITGB1 predicts prognosis in
colorectal cancer: A large prospective study based on tissue
microarray. Int J Clin Exp Pathol. 8:12802–12810. 2015.
|
|
35
|
Oshita F, Kameda Y, Ikehara M, Tanaka G,
Yamada K, Nomura I, Noda K, Shotsu A, Fujita A, Arai H, et al:
Increased expression of integrin beta1 is a poor prognostic factor
in small-cell lung cancer. Anticancer Res. 22:1065–1070.
2002.PubMed/NCBI
|
|
36
|
Yao ES, Zhang H, Chen YY, Lee B, Chew K,
Moore D and Park C: Increased beta1 integrin is associated with
decreased survival in invasive breast cancer. Cancer Res.
67:659–664. 2007. View Article : Google Scholar : PubMed/NCBI
|