Introduction
Type 1 diabetes is an increasing public health
problem with huge economical and social burdens (1-3).
The primary therapeutic method for hyperglycemia is insulin
injection, but several daily injections are required for optimal
glycemic regulation, which is difficult to achieve (4). The application of alternative
treatments, such as cell replacement therapy using β cells, are
restricted by donor shortages and immune rejection (5,6);
however, stem cells provide a promising strategy to reconstitute
pancreatic endocrine function. Human embryonic stem cells (hESCs)
are able to differentiate into all cell types, including
insulin-producing cells (IPCs) (7,8).
Thus far, the use of these cells is burdened by the increased risk
of tumor formation as well as by ethical considerations (9).
Adult stem cells, such as mesenchymal stem cells
(MSCs), are multipotent cells with self-renewal capability
(10). Among the MSCs, adipose
MSCs (hADSCs) are easily obtained (11), and amniotic MSCs (hAMSCs) are
superior due to their relatively high yield of younger cells from a
naive source (12). The potential
of these MSCs to differentiate into IPCs has been evaluated
previously. It was previously reported that IPCs may be obtained
from hADSCs using a three-stage protocol (13) or specific differentiation media
(14). Additionally, hADSC
differentiation into IPCs using pancreas/duodenum homeobox protein
1 (PDX-1) gene transfer has also been successfully described
(15). hAMSCs have been revealed
to differentiate into IPCs using stepwise differentiation protocols
(16). However, the current
strategy involves exogenous overexpression mediated by viruses, and
its safety and stability have motivated the search for optimized
methods for more efficient differentiation. Superparamagnetic iron
oxide (SPIO) nanoparticles (NPs) are non-viral gene delivery
reagents that have been investigated for DNA and RNA delivery, and
PEI@Fe3O4 NPs have demonstrated promising
results clinically (17,18). In our recent study,
PEI@Fe3O4 NPs were used in the small
interfering (siR)NA silencing of MSCs (19). Herein, novel reagent,
PEI@Fe3O4 NP, was adopted to differentiate
MSCs into IPCs.
Neuronal restrictive silencing factor (NRSF) was
first recognized due to its ability to block the transcription of
neuronal differentiation genes in non-neuronal cells or in neural
stem cells by binding to neuron-restrictive silencer element (NRSE)
(20). Target genes of NRSF
associated with islet cell development have been reported (21,22), including hepatocyte nuclear factor
4a (HNF4A), paired homeobox 4 (PAX4), neurogenin-3 (NGN3) and
neuronal differentiation 1 (NEUROD1). These data suggest that the
suppression of NRSF may promote islet cell development. Further
studies demonstrated that the downregulation of NRSF is required
for islet development (23,24), but its effect is weak unless
coexpressed with other key transcription factors. In addition,
sonic hedgehog (SHH) signaling exerts opposite roles in neuron and
pancreas development. While the expression of SHH promotes neuronal
development (25), it also
inhibits pancreas specification (26,27).
The present study aimed to investigate whether
hADSCs and hAMSCs could differentiate into IPCs using
PEI@Fe3O4 NP-mediated NRSF and SHH silencing,
which may offer a potentially effective therapeutic approach to
cell therapy for type 1 diabetes.
Materials and methods
hADSC culture
Samples from human adipose-derived tissues were
obtained from The First Hospital of China Medical University
(Shenyang, China), and all samples were collected following written
informed consent being obtained. The present study was approved by
the Ethical Committee of the First Hospital of China Medical
University. Adipose tissues were washed with PBS and incubated with
collagenase I (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany) at
37°C for 30 min. Enzyme activity was neutralized with 10% fetal
bovine serum (FBS; Hyclone; GE Healthcare Life Sciences, Logan, UT,
USA) at room temperature immediately, and the cells were filtered
through a 70-µm cell sieve and collected by centrifugation at 180 x
g for 10 min at room temperature. The hADSCs were cultured in
Dulbecco's modified Eagle medium (DMEM)/F12 (Gibco; Thermo Fisher
Scientific, Inc., Waltham, MA, USA) at 37°C with 5% CO2.
Once 80-90% confluence had been achieved, the cells were
trypsinized and resuspended in DMEM/F12 containing 10% FBS. hADSCs
that had been passaged thrice were used for subsequent
experiments.
hAMSC culture
Samples from human amnion tissues were obtained from
the First Hospital of China Medical University, and all samples
were collected once written informed consent was obtained. Freshly
isolated amniotic membranes were washed with 0.9% saline
supplemented with gentamicin and amphotericin B to remove the blood
and then immersed in PBS for 10 min at room temperature. The amnion
was transferred into 0.25% trypsin solution and incubated at 37°C
for 30 min; then, the supernatant was discarded, and this process
was repeated three times. Enzyme activity was neutralized with 10%
FBS at room temperature immediately. Next, the amnion was incubated
with PBS containing 1 mg/ml collagenase IV (Sigma-Aldrich; Merck
KGaA) and 0.1 mg/ml DNase I (Takara Biotechnology Co., Ltd.,
Dalian, China) at 37°C for 1 h, then the cells were filtered
through a 70-µm cell sieve and collected by centrifugation at 180 ×
g for 10 min at room temperature. The hAMSCs were cultured in
DMEM/F12 supplemented with 10% FBS at 37°C with 5% CO2.
Once 80-90% confluence had been achieved, the cells were
trypsinized and resuspended in DMEM/F12 containing 10% FBS. hADSCs
that had been passaged thrice were used for subsequent
experiments.
Flow cytometric characterization of
MSCs
Flow cytometry was used to detect MSC surface
markers on the cultured cells. The cells were trypsinized and
collected in regular mesenchymal media, then centrifuged at 180 × g
for 10 min at room temperature. Subsequently, the cells were
resuspended in PBS, and cells incubated with monoclonal
phycoerythrin-conjugated antibodies directed against cluster of
differentiation (CD)45 (1:100; cat. no. 368509), CD34 (1:100; cat.
no. 343505), human leukocyte antigen-antigen D related (HLA-DR)
(1:100; cat. no. 307605), CD90 (1:100; cat. no. 32810) and CD105
(1:100; cat. no. 323205) (all from BD Biosciences, San Jose, CA,
USA) for 30 min on ice. The cells were washed twice with PBS and
then analyzed by flow cytometry. Data were analyzed using FCS
Express 6.0 software (BD Biosciences).
PEI@Fe3O4
NP-mediated siRNA transfection
In our previous study,
PEI@Fe3O4 NPs were successfully synthesized
(19). In the present study,
PEI@Fe3O4 NPs were used to deliver siRNA into
MSCs. NRSF siRNA, SHH siRNA and siRNA FAM-conjoined NC (all from
Shanghai GenePharma Co., Ltd., Shanghai, China) were applied to the
MSCs using PEI@Fe3O4 NPs. The following siRNA
sequences were used: NRSF siRNA, 5′-GGC CUC UAA UCA ACA UGA ATT-3′;
SHH siRNA, 5′-GGU GUA AGG ACA AGU UGA ATT-3′; and FAM-conjoined
siRNA NC, 5′-UUG UAC UAC ACA AAA GUA CUG-3′. The weight ratio of
PEI@Fe3O4 NPs and siRNA was 4:1, with 6 µl 6
ng/ml PEI@ Fe3O4 NPs and 2 µl 1.5 ng/ml siRNA
transfected, as previously reported (19). The MSCs were seeded in 6-well
plates, and once 80% confluence was achieved, the medium was
changed. The PEI@Fe3O4 NPs were prepared and
mixed with siRNA at room temperature for 30 min, and then added to
the wells. Following incubation overnight, the culture media was
replaced. Cells were transfected with FAM-conjoined siRNA NC for
transfection efficiency analysis. The resulting cells were analyzed
by flow cytometry as aforementioned.
Prussian blue staining
The MSCs were fixed with 4% formaldehyde for 30 min
at room temperature, washed with PBS and then immersed in a
solution consisting of equal parts of 20% HCl and 10% ferrocyanide
for 30 min at room temperature, and then washed in PBS three
times.
Cell viability assay
A total of 1×105 cells/well were seeded
in 96-well plates and transfected with 2 µg FAM-conjoined siRNA NC
using PEI@Fe3O4 NPs or Lipofectamine 2000
(Gibco; Thermo Fisher Scientific, Inc.) and incubated at 5%
CO2 and 37°C for 24-48 h. MTT stock solution was added
to each well, and the culture was continued for 4 h. Subsequently,
the culture solution in the wells was removed. DMSO was added to
each well, and then the 96-well plate was agitated at a low speed
for 10 min. The absorbance of each well was measured at 570 nm as
the optical density.
Reverse transcription-quantitative
polymerase chain reaction (RT-qPCR) analysis
Total RNA was extracted from MSCs using TRIzol
reagent (Invitrogen; Thermo Fisher Scientific, Inc.). The
concentration of extracted RNA was determined using an ultraviolet
spectrophotometer. cDNA was synthesized using PrimeScript™ RT
reagent (Takara Biotechnology Co., Ltd.) according to the
manufacturer's protocol. The primers used are shown in Table I, and the reactions were performed
using the SYBR PrimeScript RT-PCR kit (Takara Biotechnology Co.,
Ltd.) with an ABI 7500 Sequence Detection system (Applied
Biosystems; Thermo Fisher Scientific, Inc.). The PCR thermocycling
conditions were as follows: 95°C for 30 sec; followed by 45 cycles
of 95°C for 5 sec and 60°C for 34 sec. Each experiment was
performed three times. As an internal control, levels of GAPDH were
quantified in parallel with the target genes. Normalization and
fold changes were calculated using the ΔΔCq method (28).
 | Table IPrimers for RT-qPCR analysis. |
Table I
Primers for RT-qPCR analysis.
| Gene name | Primer sequence
5′-3′ |
|---|
| PDX1 | |
| Forward |
ACTCCACCTTGGGACCTGTTTAGA |
| Reverse |
CGAGTAAGAATGGCTTTATGGCAGA |
| Insulin | |
| Forward |
GCCGCAGCCTTTGTGAA |
| Reverse |
CGGGTCTTGGGTGTGTAGAAG |
| NGN3 | |
| Forward |
TGCTCATCGCTCTCTATTCTTTTG |
| Reverse |
GGCAGGTCACTTCGTCTTCC |
| PAX4 | |
| Forward |
GGCTGTGTGAGCAAGATCCTAGGA |
| Reverse |
TTGCCAGGCAAAGAGGGCTGGAC |
| NRSF | |
| Forward |
ATTGAAGTTGGCTTAGTG |
| Reverse |
TATGGGTAGATTCGTTGA |
| SHH | |
| Forward |
GGCTGGATTCGACTGGGTCTACTA |
| Reverse |
AACTTGGTGCCACCCTGCTC |
| GAPDH | |
| Forward |
GCACCGTCAAGGCTGAGAAC |
| Reverse |
TGGTGAAGACGCCAGTGGA |
Western blot analysis
Cells were lysed with radio immunoprecipitation
assay (Beyotime Institute of Biotechnology, Haimen, China) buffer
containing a complete protease inhibitor cocktail tablet. The
protein concentration was determined using a BCA protein assay. A
total of 20 µg protein/lane was separated using 10% SDS-PAGE and
transferred to polyvinylidene difluoride membranes. The membranes
were incubated with primary antibodies, including rabbit anti-NRSF
(1:1,000; cat. no. 21635; Abcam, Cambridge, MA, USA), rabbit
anti-SHH (1:1,000; cat. no. 22075; Cell Signaling Technology, Inc.,
Danvers, MA, USA) and mouse anti-GAPDH (1:5,000; cat. no. sc-47727;
Santa Cruz Biotechnology, Inc., Dallas, TX, USA) overnight at 4°C.
Following incubation with goat anti-rabbit IgG horseradish
peroxidase-conjugated secondary antibody (1:10,000; cat. no.
sc-2004; Santa Cruz Biotechnology, Inc.) at room temperature for 1
h, the results were visualized using Amersham ECL Prime Western
Blotting Detection reagent (GE Healthcare, Chicago, IL, USA) and a
Tanon-5200 chemiluminescence detection system (Tanon Science and
Technology Co., Ltd., Shanghai, China). GAPDH was used as an
internal control.
Immunofluorescence
Cells were washed with PBS twice, fixed with 4%
paraformaldehyde for 10 min at room temperature and then blocked
with 1% bovine serum albumin (Amresco, LLC, Solon, OH, USA) for 60
min at room temperature. Subsequently, cells were permeabilized
with 0.1% Triton X-100 for 5 min at room temperature. Cells were
incubated with the rabbit anti-insulin (1:1,000; cat. no. 3014S)
and rabbit anti-glucagon primary antibodies (1:1,000; cat. no.
2760S) (both from Cell Signaling Technology, Inc.) at 4°C
overnight. Subsequently, the cells were washed with PBS and
incubated with the mouse anti-rabbit fluorescence-labeled secondary
antibody (1:400; cat. no. SC-2359; Santa Cruz Biotechnology, Inc.)
for 1 h at room temperature. The labeled cells were visualized
under a fluorescence microscope at ×10 magnification.
Glucose-stimulated insulin secretion
assay
The cells were tested for insulin secretion at basal
(5.5 mM) and stimulated (25 mM) glucose concentrations. The
differentiated cells in the plates were washed with PBS and
incubated in 1 ml of serum-free DMEM containing 5.5 mM glucose for
5 h at 37°C. The media were collected and stored at −20°C; then,
fresh media with 25 mM glucose was added. After 5 h of incubation
at 37°C, the media were collected and stored. The stored media were
then analyzed for insulin content using a direct human insulin
ELISA kit (cat. no. KAQ1251; Invitrogen; Thermo Fisher Scientific,
Inc.). Non-induced MSCs were used as controls.
Statistical analysis
Data are expressed as the mean ± standard deviation
following three independent experiments. The statistical analyses
were performed using the Student's t-test or one-way analysis of
variance followed by Tukey's post hoc test with GraphPad Prism 5.0
software (GraphPad Software, Inc., La Jolla, CA, USA). P<0.05
was considered to indicate a statistically significant
difference.
Results
Characterization of the cultured MSCs
from different tissues
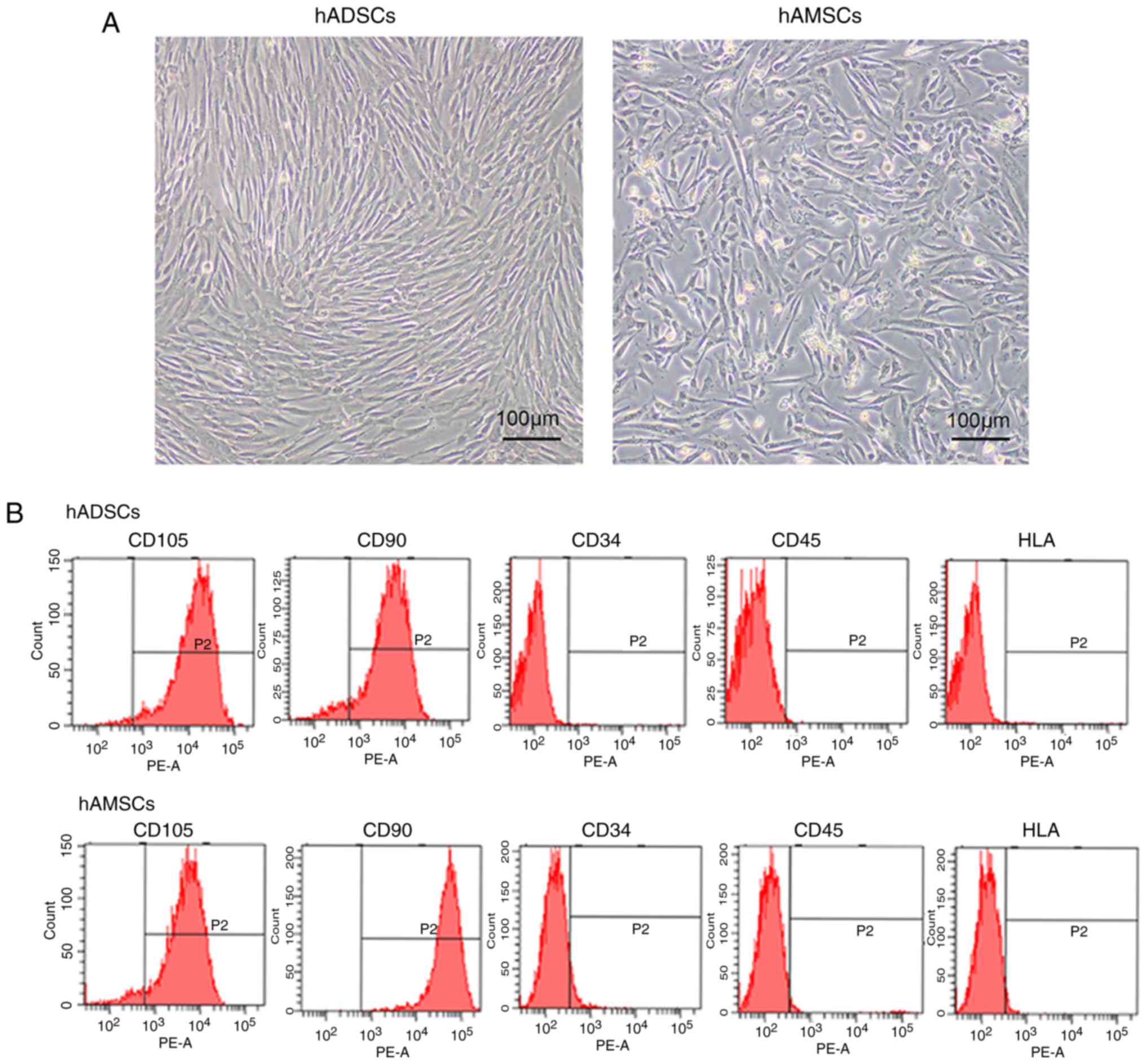
MSCs were isolated from human adipose tissues and
amnion tissues. They were expanded in a culture flask and exhibited
similar fibroblast-like morphologies. The morphology of the hADSCs
was longer compared with the hAMSCs, and hADSCs exhibited a shorter
doubling time (Fig. 1A). The
associated cells surface markers were assessed by flow cytometry to
characterize isolated MSCs. All types of MSCs expressed high levels
of CD105 and CD90, but negligible levels of CD34, CD45 and HLA-DR,
indicating a satisfactory level of homogeneity of the MSCs in the
culture (Fig. 1B).
Transfection efficiency of
PEI@Fe3O4 NPs/siRNA
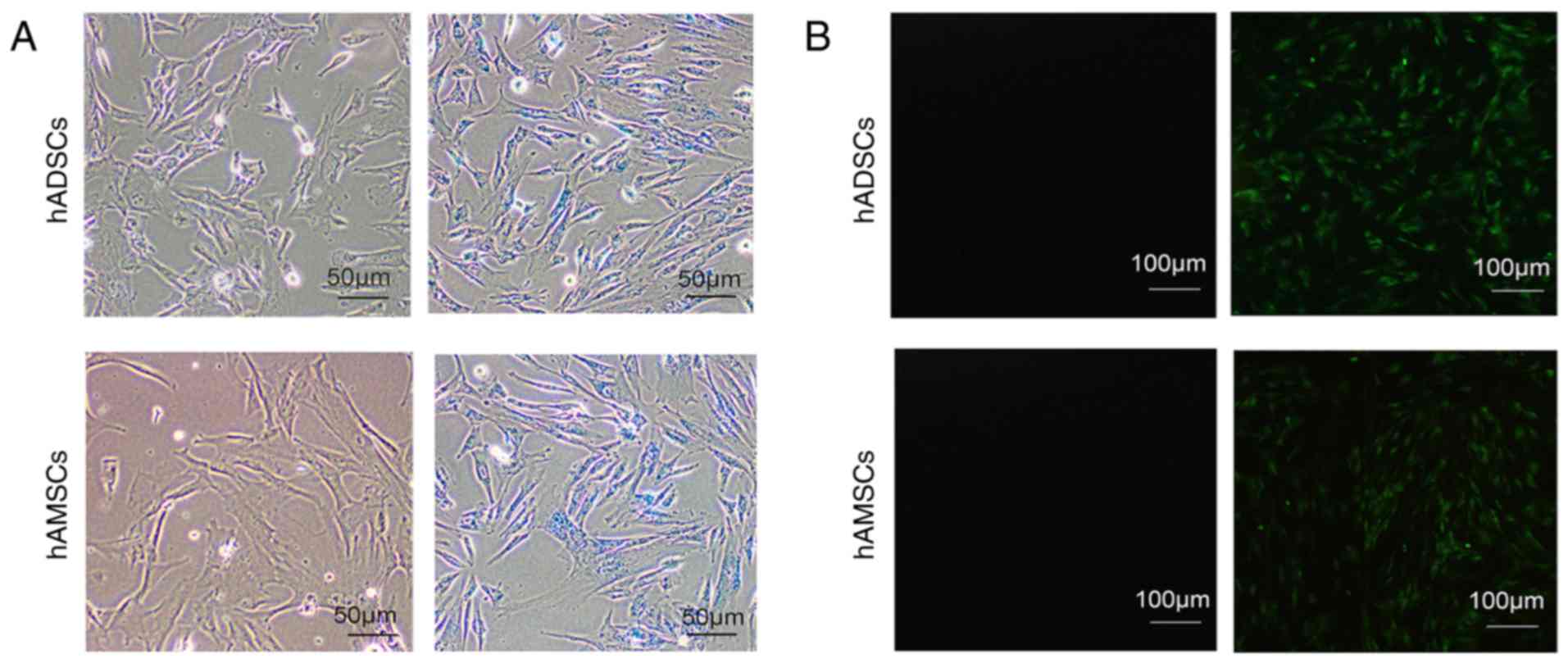
To evaluate the transfection efficiency of the
PEI@Fe3O4 NPs/siRNA complex, fluorescence
microscopy and Prussian blue staining was used. According to
Prussian blue staining, more than 96% of the cells were positive
(Fig. 2A). Furthermore, according
to the fluorescence microscopic analysis, >80% of the cells were
FAM-positive (Fig. 2B). Thus,
PEI@Fe3O4 NPs were considered efficient siRNA
delivery reagents in MSCs.
Comparison of the cytotoxicity of
PEI@Fe3O4 NPs/siRNA and Lipofectamine
2000/siRNA
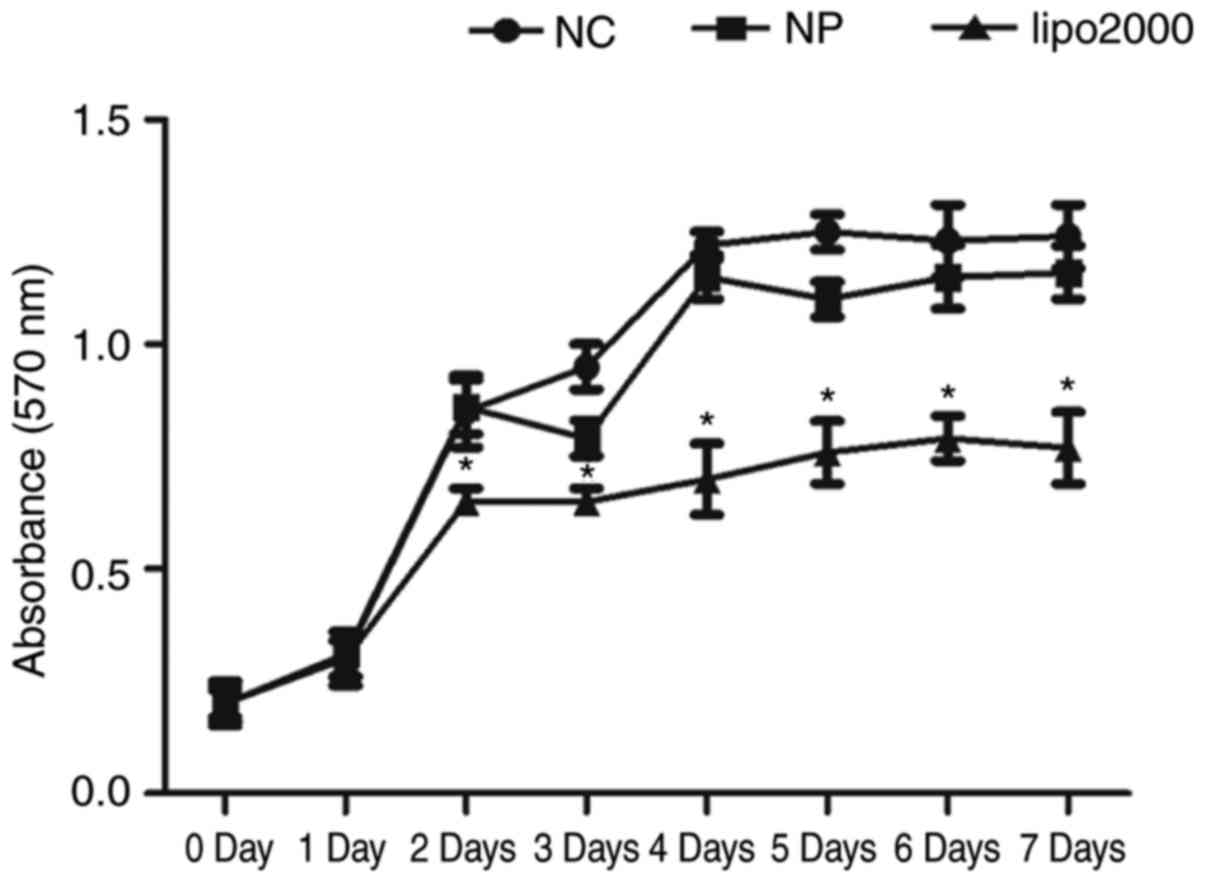
In addition to transfection efficiency, cytotoxicity
is also an important aspect for gene delivery reagents. The
majority of the non-viral delivery reagents are nanoparticles and
liposomes, and Lipofectamine 2000 is the most commonly used
liposome. The present study compared the cytotoxicity of
Lipofectamine 2000 with the PEI@ Fe3O4 NPs.
As demonstrated by the MTT cell proliferation assay, the
PEI@Fe3O4 NPs exhibited similar activity, and
Lipofectamine 2000 exhibited significantly lower activity compared
with the non-transfected cells after transfection for 48 h
(Fig. 3).
Expression of NRSF and SHH is
downregulated in MSCs by PEI@Fe3O4 NPs
Concurrent with successful transfection, it is
important to demonstrate the interference efficiency. To determine
the interference efficiency of PEI@Fe3O4 NPs
in the MSCs, RT-qPCR and western blot analysis were used to detect
the expression level of NRSF and SHH in the MSCs following
transfection for 2 days (Fig. 4A and
B). Using western blot analysis, markedly reduced NRSF and SHH
protein expression was observed in the MSCs following
siRNA-targeted knockdown compared with the NC control (Fig. 4A). In addition, the mRNA level of
NRSF and SHH was significantly silenced to 30% of the control in
MSCs using PEI@Fe3O4 NPs (Fig. 4B). Furthermore, on the 14th day of
the transfection, RT-qPCR was used to detect the NRSF and SHH
expression levels in MSCs. The mRNA levels of NRSF and SHH were
silenced to 70% of the control (Fig.
4C). These results suggested that the
PEI@Fe3O4 NPs were highly efficient in
silencing NRSF and SHH, and the interference efficacy lasted for 14
days.
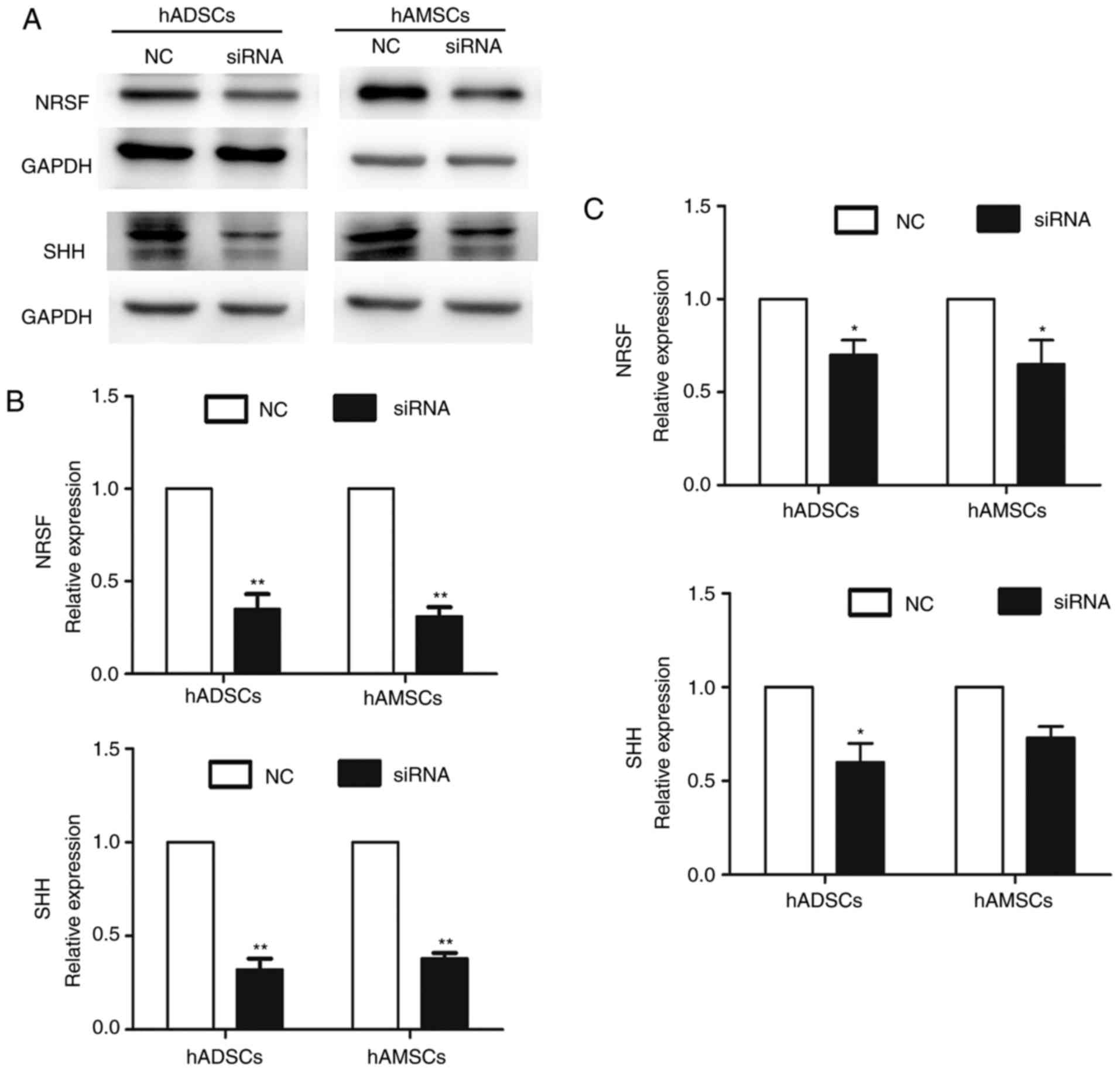 | Figure 4Interference efficiency. After 2 days
of transfection, NRSF and SHH expression levels in MSCs were
detected by (A) western blot analysis and (B) RT-qPCR. (C) A total
of 14 days after transfection, NRSF and SHH expression levels in
MSCs were detected by RT-qPCR. Data are presented as the mean ±
standard deviation (n=3). *P<0.05 and
**P<0.01. RT-qPCR, reverse transcription-quantitative
polymerase chain reaction. NRSF, neuronal restrictive silencing
factor; SHH, sonic hedgehog; MSCs, mesenchymal stem cells; siRNA,
small interfering RNA; NC, negative control; hADSCs, human
adipose-derived stem cells; hAMSCs, human amniotic mesenchymal stem
cells. |
Differentiation of MSCs into IPCs
To examine whether NRSF and SHH silencing promote
MSCs to differentiate into IPCs, the expression levels of several
genes specific for islet cells were detected by RT-qPCR. NRSF and
SHH knockdown for 7 days and 14 days significantly enhanced the
islet progenitor expression of NGN3, PAX4, PDX1 and insulin
compared with the NC control. The expression of these genes was
significantly higher in hADSCs compared with that in hAMSCs, with
the exception of PAX4 (Fig. 5A).
Immunofluorescence staining was performed to detect the glucagon
and insulin proteins in the induced cells following transfection
for 14 days. The results demonstrated that the differentiated MSCs
expressed glucagon and insulin (Fig.
5B). To determine the differentiation ability of the induced
cells following transfection for 14 days, the glucose-stimulated
insulin secretion assay was performed. The differentiated MSCs were
treated with 5.5 and 25 mM glucose, and insulin concentrations were
measured using immunoassays. The induced hADSCs and hAMSCs secreted
insulin, but hADSCs demonstrated a significantly increased ability
to secrete insulin compared with hAMSCs following stimulation with
25 mM glucose (Fig. 5C). In
summary, the present results indicate that the induced MSCs
acquired the characteristics of functional IPCs, and hADSCs
exhibited a greater potential compared with hAMSCs for diabetic
treatment.
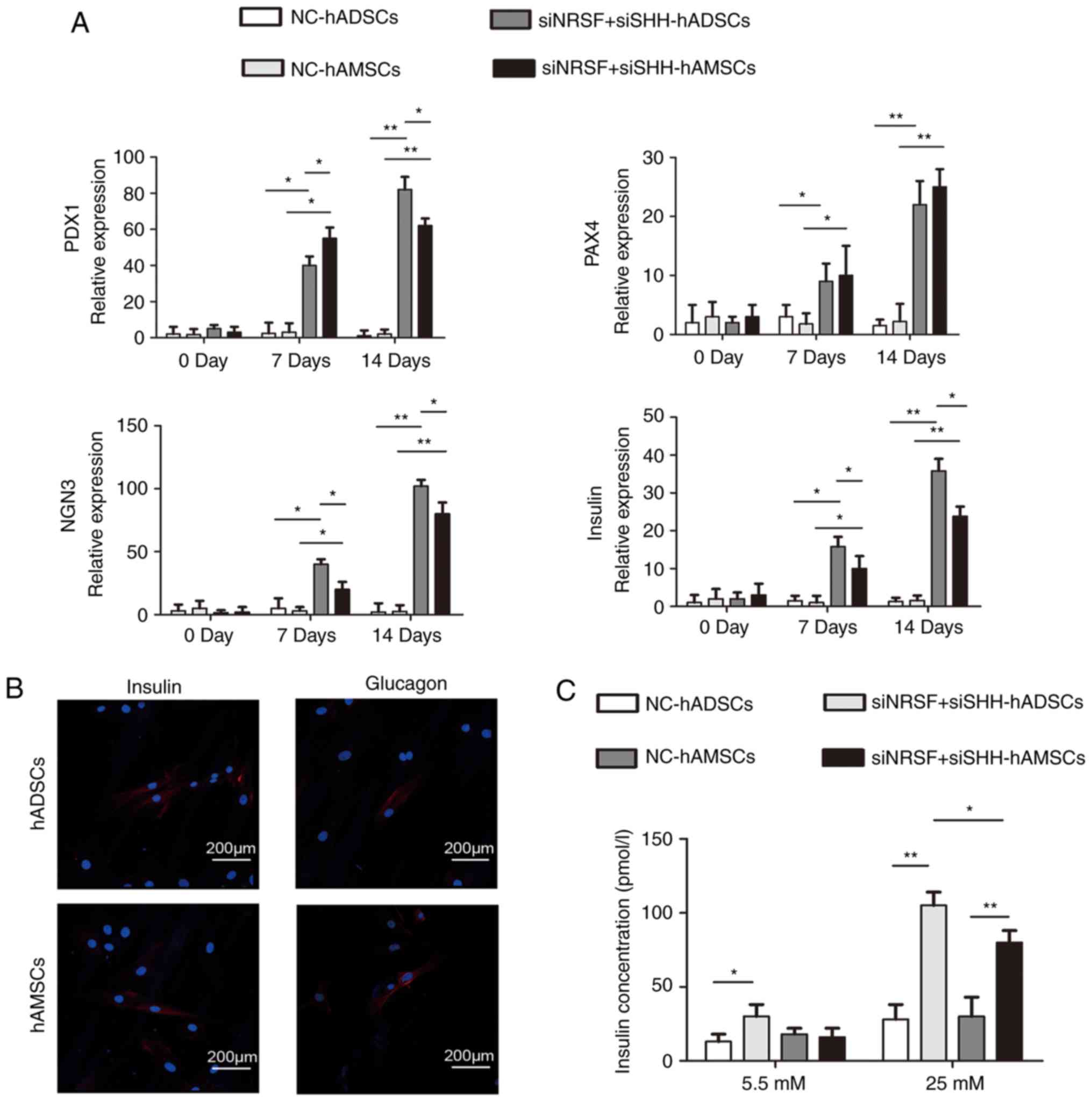 | Figure 5Detection of the islet
cell-associated genes in the induced MSCs. (A) Reverse
transcription-quantitative polymerase chain reaction was used to
detect the insulin, PDX1, PAX4 and NGN3 in the NC (D0), and in the
differentiated MSCs (D7, D14). *P<0.05 and
**P<0.01. (B) Immunofluorescence analysis for insulin
and glucagon in the differentiated MSCs (D14). (C) Insulin
concentrations were detected in NC and the differentiated MSCs
(D14). Insulin concentrations were determined with an
ultrasensitive insulin ELISA assay. Data are presented as the mean
± standard deviation (n=3). *P<0.05 and
**P<0.01. PDX1, pancreas/duodenum homeobox protein 1;
PAX4, paired homeobox 4; NGN3, neurogenin-3; MSCs, mesenchymal stem
cells; siRNA/si, small interfering RNA; NC, negative control;
hADSCs, human adipose-derived stem cells; hAMSCs, human amniotic
mesenchymal stem cells. |
Discussion
The present study investigated the differentiation
of hADSCs and hAMSCs into IPCs. It was demonstrated that hADSCs and
hAMSCs were able to differentiate into IPCs using
PEI@Fe3O4 NP-mediated NRSF and SHH silencing.
The differentiated MSCs exhibited elevated glucose-stimulated
insulin secretory abilities. The differentiation of these MSCs into
IPCs may provide an alternative renewable therapeutic strategy for
diabetes treatment.
Currently, genetic delivery reagents are viral or
synthetic non-viral. Although the transfection efficiency of the
viral reagents is higher, there are several issues, including the
induction of host immune responses and high tumorigenicity rates
(29). Studies have reported MSC
differentiation into IPCs using lentivirus (23,24,30). The lack of reports on MSC
differentiation into IPCs using non-viral gene delivery reagents
led us to focus on a safer and more efficient reagent. The majority
of the non-viral delivery reagents are nanoparticles and liposomes.
The present study compared the cytotoxicity of liposomes with the
PEI@Fe3O4 NPs, and confirmed that PEI@
Fe3O4 NPs exhibited lower cytotoxicity levels
and were labeled by Prussian Blue staining. This indicates that
PEI@Fe3O4 NPs may be a better delivery
reagent for cell therapy in the clinical setting. To the best of
our knowledge, the present study is the first to differentiate
hADSCs and hAMSCs into IPCs via NP-mediated gene silencing.
The mechanism responsible for islet cell
differentiation from stem cells remains to be elucidated. During
the development of islet cells, NRSF represses the human insulin
gene by binding to NRSE in its promoter region (31). Several studies have demonstrated
that following the downregulation of NRSF, MSCs are able to
differentiate into IPCs (22.32). However, hADSCs and hAMSCs cannot
differentiate into IPCs through the suppression of NRSF alone.
Although SHH is not normally expressed in the pancreatic domain, a
study reported that SHH is required for the initiation of pancreas
gene expression in the posterior foregut (25). Consistently, hADSCs and hAMSCs
cannot differentiate into IPCs via suppression of SHH alone. It has
been reported that human amniotic fluid-derived stem cells are able
to differentiate into IPCs using differentiation medium and the
repression of NRSF (23).
Cytokines in the differentiation medium have been reported to
mediate the suppression of SHH signaling (23). Thus, in the present study, the
combined suppression of NRSF and SHH in hADSCs and hAMSCs using
PEI@Fe3O4 NPs was performed. The
downregulation of NRSF and SHH may trigger a derepression of NRSF-
and SHH-regulated genes, subsequently promoting hADSCs and hAMSCs
to differentiate into IPCs, and enhancing the islet progenitor
expression of NGN3, PAX4, PDX1 and insulin. The mRNA expression of
these genes was significantly higher in hADSCs compared with that
in hAMSCs, with the exception of PAX4. Furthermore, hADSCs
demonstrated an increased ability to secrete insulin compared with
hAMSCs in a glucose-responsive manner. These data suggest that NRSF
and SHH may serve dominant roles in enhancing the differentiation
of the MSCs into IPCs, and that hADSCs exhibit a greater potential
compared with hAMSCs for use in diabetic treatment.
In conclusion, the results of the present study
demonstrated that the suppression of NRSF and SHH using
PEI@Fe3O4 NPs may promote hADSCs and hAMSCs
to differentiate into IPCs, and these differentiated cells are able
to secrete insulin in response to glucose stimulation. These
results support the further evaluation of the use of hADSCs and
hAMSCs in cell therapy in diabetes.
Funding
The present study was funded by the National Natural
Science Foundation of China (grant no. 81703102), Shenyang
Transformation Project of Major Scientific and Technological
Achievements (grant no. Z17-5-039), Shenyang Key Laboratory Project
(grant no. F15-157-1-00) and Foundation of China Medical University
(grant no. XZR20160022).
Availability of data and materials
All data generated or analyzed during this study are
included in this published article.
Authors' contributions
RW and DZ performed the experiments and wrote the
manuscript. TZ and FZ contributed to data analysis and
interpretation. HL and XL contributed to collection and assembly of
data. XP designed the study and provided final approval of
manuscript.
Ethics approval and consent to
participate
Samples from human adipose-derived tissues and human
amnion tissues were obtained from The First Hospital of China
Medical University (Shenyang, China), and all samples were
collected following written informed consent being obtained. The
present study was approved by the Ethical Committee of the First
Hospital of China Medical University.
Patient consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing
interests.
Acknowledgments
Not applicable.
References
|
1
|
Shaw JE, Sicree RA and Zimmet PZ: Global
estimates of the prevalence of diabetes for 2010 and 2030. Diabetes
Res Clin Pract. 87:4–14. 2010. View Article : Google Scholar
|
|
2
|
Hematti P, Kim J, Stein AP and Kaufman D:
Potential role of mesenchymal stromal cells in pancreatic islet
transplantation. Transplant Rev (Orlando). 27:21–29. 2013.
View Article : Google Scholar
|
|
3
|
American Diabetes Association: Standards
of medical care in diabetes-2013. Diabetes Care. 36(Suppl 1):
S11–S66. 2013. View Article : Google Scholar
|
|
4
|
Larrañaga A, Docet MF and Garcia-Mayor RV:
Disordered eating behaviors in type 1 diabetic patients. World J
Diabetes. 2:189–195. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
5
|
Ryan EA, Paty BW, Senior PA, Bigam D,
Alfadhli E, Kneteman NM, Lakey JR and Shapiro AM: Five-year
follow-up after clinical islet transplantation. Diabetes.
54:2060–2069. 2005. View Article : Google Scholar : PubMed/NCBI
|
|
6
|
Miyazaki S, Yamato E and Miyazaki J:
Regulated expression of pdx-1 promotes in vitro differentiation of
insulin-producing cells from embryonic stem cells. Diabetes.
53:1030–1037. 2004. View Article : Google Scholar : PubMed/NCBI
|
|
7
|
Kroon E, Martinson LA, Kadoya K, Bang AG,
Kelly OG, Eliazer S, Young H, Richardson M, Smart NG, Cunningham J,
et al: Pancreatic endoderm derived from human embryonic stem cells
generates glucose-responsive insulin-secreting cells in vivo. Nat
Biotechnol. 26:443–452. 2008. View
Article : Google Scholar : PubMed/NCBI
|
|
8
|
Soria B, Skoudy A and Martin F: From stem
cells to beta cells: New strategies in cell therapy of diabetes
mellitus. Diabetologia. 44:407–415. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
9
|
Lumelsky N, Blondel O, Laeng P, Velasco I,
Ravin R and McKay R: Differentiation of embryonic stem cells to
insulin-secreting structures similar to pancreatic islets. Science.
292:1389–1394. 2001. View Article : Google Scholar : PubMed/NCBI
|
|
10
|
Krampera M, Franchini M, Pizzolo G and
Aprili G: Mesenchymal stem cells: From biology to clinical use.
Blood Transfus. 5:120–129. 2007.PubMed/NCBI
|
|
11
|
Gimble JM, Katz AJ and Bunnell BA:
Adipose-derived stem cells for regenerative medicine. Circ Res.
100:1249–1260. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
12
|
Diaz-Prado S, Muiños-López E,
Hermida-Gómez T, Cicione C, Rendal-Vázquez ME, Fuentes-Boquete I,
de Toro FJ and Blanco FJ: Human amniotic membrane as an alternative
source of stem cells for regenerative medicine. Differentiation.
81:162–171. 2011. View Article : Google Scholar : PubMed/NCBI
|
|
13
|
Gabr MM, Zakaria MM, Refaie AF, Khater SM,
Ashamallah SA, Ismail AM, El-Halawani SM and Ghoneim MA:
Differentiation of human bone marrow-derived mesenchymal stem cells
into insulin-producing cells: Evidence for further maturation in
vivo. Biomed Res Int. 2015:5758372015. View Article : Google Scholar : PubMed/NCBI
|
|
14
|
Sun Y, Zhang M, Ji S and Liu L: Induction
differentiation of rabbit adipose-derived stromal cells into
insulin-producing cells in vitro. Mol Med Rep. 12:6835–6840. 2015.
View Article : Google Scholar : PubMed/NCBI
|
|
15
|
Kajiyama H, Hamazaki TS, Tokuhara M, Masui
S, Okabayashi K, Ohnuma K, Yabe S, Yasuda K, Ishiura S, Okochi H
and Asashima M: Pdx1-transfected adipose tissue-derived stem cells
differentiate into insulin-producing cells in vivo and reduce
hyperglycemia in diabetic mice. Int J Dev Biol. 54:699–705. 2010.
View Article : Google Scholar
|
|
16
|
Okere B, Alviano F, Costa R, Quaglino D,
Ricci F, Dominici M, Paolucci P, Bonsi L and Iughetti L: In vitro
differentiation of human amniotic epithelial cells into
insulin-producing 3D spheroids. Int J Immunopathol Pharmacol.
28:390–402. 2015. View Article : Google Scholar : PubMed/NCBI
|
|
17
|
Watanabe M, Yoneda M, Morohashi A, Hori Y,
Okamoto D, Sato A, Kurioka D, Nittami T, Hirokawa Y, Shiraishi T,
et al: Effects of Fe3O4 magnetic
nanoparticles on A549 cells. Int J Mol Sci. 14:15546–15560. 2013.
View Article : Google Scholar : PubMed/NCBI
|
|
18
|
Figuerola A, Di Corato R, Manna L and
Pellegrino T: From iron oxide nanoparticles towards advanced
iron-based inorganic materials designed for biomedical
applications. Pharmacol Res. 62:126–143. 2010. View Article : Google Scholar : PubMed/NCBI
|
|
19
|
Zhang D, Wang J, Wang Z, Wang R, Song L,
Zhang T, Lin X, Shi P, Xin H and Pang X: Polyethyleneimine-coated
Fe3O4 nanoparticles for efficient siRNA
Delivery to human mesenchymal stem cells derived from different
tissues. Sci Adv Mater. 7:1058–1064. 2015. View Article : Google Scholar
|
|
20
|
Hermanson O: Stem cells have different
needs for Rest. PloS Biol. 6:e2712008. View Article : Google Scholar : PubMed/NCBI
|
|
21
|
Johnson DS, Mortazavi A, Myers RM and Wold
B: Genome-wide mapping of in vivo protein-DNA interactions.
Science. 316:1497–1502. 2007. View Article : Google Scholar : PubMed/NCBI
|
|
22
|
Martin D, Tawadros T, Meylan L,
Abderrahmani A, Condorelli DF, Waeber G and Haefliger JA: Critical
role of the transcriptional repressor neuron-restrictive silencer
factor in the specific control of connexin36 in insulin-producing
cell lines. J Biol Chem. 278:53082–53089. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
23
|
Li B, Wang S, Liu H, Liu D, Zhang J, Zhang
B, Yao H, Lv Y, Wang R, Chen L, et al: Neuronal restrictive
silencing factor silencing induces human amniotic fluid-derived
stem cells differentiation into insulin-producing cells. Stem Cells
Dev. 20:1223–1231. 2011. View Article : Google Scholar
|
|
24
|
Li H, Jiang F, Shi P, Zhang T, Liu XY, Lin
XW and Pang XN: In vitro reprogramming of rat bone marrow-derived
mesen-chymal stem cells into insulin-producing cells by genetically
manipulating negative and positive regulators. Biochem Bioph Res
Commun. 420:793–798. 2012. View Article : Google Scholar
|
|
25
|
Dayer D, Tabar MH, Moghimipour E, Tabandeh
MR, Ghadiri AA, Bakhshi EA, Orazizadeh M and Ghafari MA: Sonic
hedgehog pathway suppression and reactivation accelerates
differentiation of rat adipose-derived mesenchymal stromal cells
toward insulin-producing cells. Cytotherapy. 19:937–946. 2017.
View Article : Google Scholar : PubMed/NCBI
|
|
26
|
Hebrok M, Kim SK and Melton DA: Notochord
repression of endodermal Sonic hedgehog permits pancreas
development. Genes Dev. 12:1705–1713. 1998. View Article : Google Scholar : PubMed/NCBI
|
|
27
|
Schwitzgebel VM, Mamin A, Brun T,
Ritz-Laser B, Zaiko M, Maret A, Jornayvaz FR, Theintz GE, Michielin
O, Melloul D and Philippe J: Agenesis of human pancreas due to
decreased half-life of insulin promoter factor 1. J Clin Endocrinol
Metab. 88:4398–4406. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
28
|
Livak KJ and Schmittgen TD: Analysis of
relative gene expression data using real-time quantitative PCR and
the 2(-delta delta C(T)) method. Methods. 25:402–408. 2001.
View Article : Google Scholar
|
|
29
|
Yin H, Kanasty RL, Eltoukhy AA, Vegas AJ,
Dorkin JR and Anderson DG: Non-viral vectors for gene-based
therapy. Nat Rev Genet. 15:541–555. 2014. View Article : Google Scholar : PubMed/NCBI
|
|
30
|
Allahverdi A, Abroun S, Jafarian A,
Soleimani M, Taghikhani M and Eskandari F: Differentiation of human
mesenchymal stem cells into insulin producing cells by using a
lentiviral vector carrying PDX1. Cell J. 17:231–242.
2015.PubMed/NCBI
|
|
31
|
Kemp DM, Lin JC and Habener JF: Regulation
of Pax4 paired homeodomain gene by neuron-restrictive silencer
factor. J Biol Chem. 278:35057–35062. 2003. View Article : Google Scholar : PubMed/NCBI
|
|
32
|
Li HT, Jiang FX, Shi P, Zhang T, Liu XY,
Lin XW, San ZY and Pang XN: In vitro reprogramming of rat bmMSCs
into pancreatic endocrine-like cells. In Vitro Cell Dev Biol Anim.
53:157–166. 2017. View Article : Google Scholar
|